

Abstract
Glioblastoma is the most aggressive type of brain tumor with a very poor prognosis. Therefore, it is always of great importance to explore and develop new potential treatment for glioblastoma. Quercetin, a flavonoid present in a variety of human foods, has been shown to inhibit various tumor cell proliferation. In this study, we found that treating human glioblastoma U251 cells with 10 μg/mL quercetin for 24 hours, a concentration that was far below the IC50 (113.65 μg/mL) and at which quercetin failed to inhibit cell proliferation, inhibited cell migration (30%) and cell invasion as examined by wound scratch assay and transwell assay, respectively. We further showed that 10 μg/mL quercetin inhibited cell migration and tube formation of human umbilical vein endothelial cells induced by the conditioned medium derived from U251 cell culture. The inhibitory effect of quercetin on migration and angiogenesis is possibly mediated through the downregulation of protein levels of VEGFA, MMP9, and MMP2 as detected by Western blot. Our findings demonstrated that low concentration of quercetin antagonized glioblastoma cell invasion and angiogenesis in vitro.
Keywords: glioblastoma, quercetin, angiogenesis, invasion, metastasis
Introduction
Glioblastoma, also called glioblastoma multiforme (GBM), is the most invasive type of glioma. The incidence of GBM is two to three per 100,000 adults per year, and it accounts for about 17% of all the brain tumors.1,2 GBM is composed of several different cell types, and most of these tumors occur in the cerebral hemisphere. Glioblastoma usually grows aggressively and spreads rapidly into the nearby brain tissue.3 The median survival time for adults with glioblastoma treated with concurrent temozolomide and radiation therapy is only around 14.6 months.4 The most common treatment for GBM is surgery, followed by radiation and chemotherapy. However, it is impossible to remove the tumor entirely by surgery since GBM usually migrates and invades the surrounding normal brain tissue. The major problem in chemotherapy is the difficulty to deliver drugs into the brain.5 So far, no significant improvement has been achieved to increase the survival rates;6,7 therefore, researchers are urged to develop new treatments for GBM.
Quercetin (3,3′,4′,5,7-pentahydroxy-flavone) is one of the most abundant flavonoids found in a variety of human foods such as apples, cherries, grapes, spinach, and onions.8,9 Quercetin has the ability to scavenge free radicals and bind transition metal ions, thus functioning as a powerful antioxidant.10,11 For instance, the inhibition of peroxidation of low-density lipoprotein by quercetin potentially reduces the risk of heart disease and atherosclerosis.12,13 Quercetin has also been reported to have anti-inflammatory, antimicrobial, antiviral, antiplatelet, and anti-proliferative activities.14–17 Interestingly, quercetin inhibits the proliferation of a wide range of cancers such as lung, breast, liver, colon, and prostate,18–22 via inducing cell cycle arrest and/or promoting cell apoptosis.23–25 Recent studies also showed that quercetin is able to inhibit DNA topoisomerases I and II26 and modulate the PI3K/AKT and NF-κB signaling pathways,27,28 indicating that quercetin has various anticancer mechanisms.
It is interesting to note that quercetin can also efficiently inhibit the growth of glioblastoma cells and induce apoptosis by suppressing the NF-κB, Ras/MAPK/ERK, and PI3K/AKT signaling pathways.29–31 The minimal inhibitory concentration of quercetin on various glioma cells ranges from 20 μg/mL to 50 μg/mL. Our previous work showed that quercetin inhibits the growth of glioblastoma U251 cells in a dose-dependent manner with an IC50 of 113.65 μg/mL. At 10 μg/mL, quercetin failed to inhibit the cell growth (Liu et al, unpublished data, 2017). In this study, we investigated the effect of 10 μg/mL quercetin on cell migration and invasion and found that low concentration of quercetin inhibited glioblastoma cell invasion and angiogenesis in vitro.
Materials and methods
Reagent
Quercetin was purchased from Sigma-Aldrich Co. (St Louis, MO, USA), and Matrigel was obtained from BD Biosciences (San Jose, CA, USA). Transwell with 8.0 μm pore polycarbonate membrane insert was obtained from Corning Incorporated (Corning, NY, USA). Anti-VEGFA (Cat No sc-7269) was purchased from Santa Cruz Biotechnology Inc. (Dallas, TX, USA). Anti-MMP2 (Cat No 10373-2-AP) and anti-MMP9 antibodies (Cat No 10375-2-AP) were purchased from Proteintech, Chicago, China. Anti-α-tubulin antibody was obtained from Sigma-Aldrich Co. Radioim-munoprecipitation assay (RIPA) lysis buffer was purchased from Beyotime (Nantong, China). This study was approved by the Institutional Review Board of Ethics Committee of Renmin Hospital, Hubei University of Medicine.
Cell culture
Human glioblastoma cell line U251 and human umbilical vein endothelial cell line CRL-1730 were provided by the Cell Center of Institute of Neurosurgery, Shiyan Renmin Hospital at Hubei University of Medicine. Both cell lines were cultured in DMEM, high glucose medium, supplemented with 10% fetal bovine serum (FBS). No antibiotics were added to the medium. The cells were grown in 5% CO2 and humidity saturation at 37°C in culture flasks. Cells were trypsinized with 0.25% trypsin and split at 80% confluence.
Cell migration and invasion assay
The capacity of cell migration was measured by the scratch wound healing assay. Briefly, U251 cells were plated and allowed to grow to the mid-log phase. Cell monolayer was gently scratched with a new 200 μL pipette tip across the center of the dish. The wells were then washed twice with PBS, replenished with serum-free DMEM containing 10 μg/mL quercetin, and cultured for 24 hours or 48 hours. Cells were then fixed with 3.7% paraformaldehyde, stained with 1% crystal violet, and examined by microscope. The gap distance was quantitatively evaluated using IPP 6.0 software.
Transwell cell invasion and migration assays were also conducted. Briefly, transwell inserts were coated with matrigel which were 1:6 diluted with serum-free medium. U251 cells were trypsinized, washed, and resuspended in medium containing 1% bovine serum albumin with or without 10 μg/mL quercetin (1×105/mL). Then, 100 μL of cell suspensions was added to the upper transwell compartment, and 600 μL of DMEM with 10% FBS was added to the lower compartment. Cells in the transwell plate were cultured at 37°C with 5% CO2 for 24 hours. Cells that remained on the upper side of the filter membrane were gently removed with a cotton swab, and cells on the lower side of the insert filter were fixed with 10% methanol and stained with 0.1% crystal violet. Images of the cells on the lower side of the insert filter were taken under a microscope. A total of 16 different 200× fields were randomly chosen, and the number of cells was counted.
Preparation of tumor-conditioned medium
U251 cells were cultured in DMEM, high glucose medium, supplemented with 10% FBS. When the cell density reached 60%–70% confluency, cells were washed with PBS for three times after removing the old culture medium and cultured in serum-free medium for 24 hours. The medium was harvested, centrifuged to remove cell debris, filtered with 0.22 μm membrane, and stored in freezer for later use.
U251 cell-induced migration of endothelial cells
CRL-1730 cells were resuspended in serum-free DMEM with or without 10 μg/mL quercetin at a density of 1×105/mL. Then, 100 μL of cells was added to the upper compartment of the transwell, and 600 μL of U251-conditioned medium was added to the lower compartment. After culturing for 8 hours, cells on the lower side of the insert filter were fixed with 10% methanol, stained with 1% crystal violet, and examined by microscope.
Endothelial cell tube formation assay
Matrigel was thawed at 4°C overnight and then added to a 96-well plate (50 μL/well). The plate was incubated for 30 minutes at 37°C to solidify the gel. CRL-1730 cells were resuspended in U251-conditioned medium with or without 10 μg/mL quercetin and were gently added at 2×105/mL to the gel-coated wells. Cells were cultured for 6 hours and examined by microscope for tubing formation.
Western blot
U251 cells were harvested, washed with cold PBS twice, and lysed with the lysis buffer containing the protease inhibitor for 30 minutes at 4°C. After centrifugation, the supernatant was fractionated on a 10% sodium dodecyl sulfate poly-acrylamide gel electrophoresis (SDS-PAGE) gel (20 μg total protein/lane). Proteins were transferred to polyvinylidene difluoride (PVDF) membrane, blocked with 5% fat-free milk for 1 hour, and incubated with primary antibodies (anti-VEGFA: 1:500; anti-MMP9: 1:500; anti-MMP2: 1:500; anti-α-tubulin: 1:1,000) at 4°C overnight. Membranes were washed with PBS containing 0.1% Tween for three times and then incubated with goat anti-rabbit secondary antibodies (1:10,000) for 2 hours. Western blots were developed by using the enhanced chemiluminescence (ECL) kit, and the images were scanned and analyzed by using Quantity One software.
Statistical analysis
All the experiments were repeated three times, and the data were presented as mean ± standard deviation . Statistical analysis was conducted by using SPSS16.0 software (SPSS Inc., Chicago, IL, USA). Student’s t-test was used for two sample comparisons. P-value <0.05 was considered statistically significant.
Results
Effects of quercetin on migration and invasion of U251 cells
To study the effect of quercetin on cell migration, we conducted in vitro scratch assay. Control and quercetin-treated glioblastoma U251 cells were scratched and allowed to migrate for 24 hours or 48 hours (Figure 1A). The migration distance of cells treated with 10 μg/mL quercetin for 24 hours and 48 hours was 70.10 ± 1.18% and 67.38 ± 0.48% of those of the control cells, respectively (P<0.05). Moreover, transwell cell migration assay showed that the cell number passing the membrane for quercetin-treated cells (65±12/field) was significantly lower than that of the control group (87±5/field; P<0.05, Figure 1B and C), indicating that quercetin treatment significantly inhibited U251 cell migration and invasion.
Figure 1.

Quercetin inhibits U251 cell migration and invasion.
Notes: (A) Control and quercetin-treated confluent monolayer of U251 cells were scratched (0 hour) and allowed to migrate for 24 hours and 48 hours, respectively. (B) Transwell experiments showing control and quercetin-treated cells passing the membrane. (C) Number of cells passed the membrane for the control and quercetin-treated groups. *P<0.05. Magnification (A) ×100, (B) ×200.
The effect of quercetin on U251-induced endothelial cell tube formation
We next used human umbilical vein endothelial cell line CRL-1730 to examine the effect of quercetin on U251-induced cell migration and angiogenesis. CRL-1730 cells, treated with control or quercetin in the transwell experiments, were allowed to invade into the bottom well for 8 hours, where U251-conditioned medium was present. In contrast to control cells, of which 41±7 cells/field passed the membrane, there were only 17±3/field quercetin-treated cells in the bottom well (P<0.05, Figure 2A and B). The data strongly indicated that quercetin inhibited U251 cell-induced cell invasion. Endothelial cell tube formation was also performed with U251-conditioned medium to examine the effects of quercetin on tumor cell-induced angiogenesis. Consistently, we found an inhibitory effect of quercetin on endothelial cell tube formation (7.67±1.53 in the quercetin group vs 13.00±2.65 in the control group, P<0.05, Figure 2C and D).
Figure 2.

Quercetin inhibits U251-induced endothelial cell invasion and tube formation.
Notes: (A) Transwell experiments showing the control and quercetin-treated cells passing to the bottom wells, where U251-conditioned medium was applied. (B) Quantification of cell numbers for the control and quercetin-treated groups. (C) Endothelial cell tube formation in the presence of control and 10 μg/mL quercetin. (D) Quantification of tube formation for the control and quercetin-treated groups. *P<0.05.
Quercetin-regulated proteins involved in cell migration and angiogenesis
Proangiogenic signals such as VEGF are involved in activating endothelial cells to degrade the basement membrane, and proliferate, migrate, and form tubes in angiogenesis.32 Matrix metalloproteinases, MMP9 and MMP2, have been shown to be involved in cell migration by degrading the matrix.33 Western blot shows that quercetin treatment significantly downregulated the levels of VEGF, MMP9, and MMP2 (Figure 3), suggesting that the inhibitory effects of quercetin on cell migration and angiogenesis are partially due to the deregulation of proangiogenic signals and matrix metalloproteinases.
Figure 3.
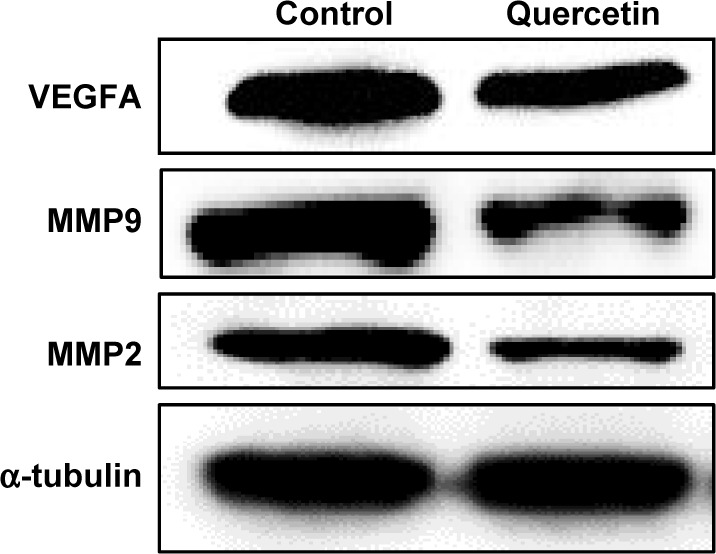
Quercetin inhibits VEGFA, MMP9, and MMP2 expression.
Note: Immunoblot showing protein levels of VEGFA, MMP9, and MMP2 in U251 cells in response to 0 μg/mL (control) and 10 μg/mL quercetin.
Discussion
In this study, we have found that quercetin inhibited glioblastoma cell migration and angiogenesis by downregulating the expression of VEGFA, MMP2, and MMP9. Tumor invasion and angiogenesis are very complex and dynamic processes that are made of multistep cascades.34 A number of molecules such as growth factors, cell adhesion molecules, and extracellular matrix have been demonstrated to be involved in these processes. For example, VEGF is a key mediator of cancer angiogenesis.35,36 It stimulates the growth of new blood vessels and allows tumors to get nutrients and oxygen for growing. MMPs can degrade and remodel the extracellular matrix, facilitating the invasion and metastasis of tumor cells.33,37 MMP2 and MMP9, two major MMPs that proteolyze IV-type collagen, have been recognized as prognostic markers for malignant glioma.38,39 Quercetin has been reported to inhibit human prostate tumor angiogenesis by targeting VEGFR2-regulated AKT/mTOR/P70S6K signaling pathways.40 Quercetin also inhibits migration and invasion of human oral cancer cells through suppressing MMP2/9 expression.41 Moreover, Santos et al29 demonstrated that quercetin delayed cell migration in human GL-15 glioblastoma cells, possibly by reducing the expression of metalloproteinase, MMP2, as well as increasing the expression of fibronectin and laminin. In this study, we found that quercetin inhibited glioblastoma cell invasion and angiogenesis at 10 μg/mL, which was the lowest concentration reported so far for quercetin-mediated anti-invasion effects in vitro. In the future, we will examine the in vivo anticancer effects of quercetin using animal models and test any potential synergistic effects of quercetin with other chemotherapy drugs on cell migration and angiogenesis.
Conclusion
Our findings demonstrated that low concentration of quercetin antagonized glioblastoma cell invasion and angiogenesis in vitro.
Acknowledgments
This work was supported by the Foundation of Hubei Educational Committee (B2015171).
Footnotes
Disclosure
The authors report no conflicts of interest in this work.
References
- 1.Burger PC, Vogel FS, Green SB, Strike TA. Glioblastoma multiforme and anaplastic astrocytoma. Pathologic criteria and prognostic implications. Cancer. 1985;56(5):1106–1111. doi: 10.1002/1097-0142(19850901)56:5<1106::aid-cncr2820560525>3.0.co;2-2. [DOI] [PubMed] [Google Scholar]
- 2.Rousseau A, Mokhtari K, Duyckaerts C. The 2007 WHO classification of tumors of the central nervous system – what has changed? Curr Opin Neurol. 2008;21(6):720–727. doi: 10.1097/WCO.0b013e328312c3a7. [DOI] [PubMed] [Google Scholar]
- 3.Reardon DA, Wen PY. Glioma in 2014: unravelling tumour heterogeneity-implications for therapy. Nat Rev Clin Oncol. 2015;12(2):69–70. doi: 10.1038/nrclinonc.2014.223. [DOI] [PubMed] [Google Scholar]
- 4.Stupp R, Mason WP, van den Bent MJ, et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N Engl J Med. 2005;352(10):987–996. doi: 10.1056/NEJMoa043330. [DOI] [PubMed] [Google Scholar]
- 5.Fadul CE, Wen PY, Kim L, Olson JJ. Cytotoxic chemotherapeutic management of newly diagnosed glioblastoma multiforme. J Neurooncol. 2008;89(3):339–357. doi: 10.1007/s11060-008-9615-4. [DOI] [PubMed] [Google Scholar]
- 6.Stupp R, Hegi ME, Mason WP, et al. Effects of radiotherapy with concomitant and adjuvant temozolomide versus radiotherapy alone on survival in glioblastoma in a randomised phase III study: 5-year analysis of the EORTC-NCIC trial. Lancet Oncol. 2009;10(5):459–466. doi: 10.1016/S1470-2045(09)70025-7. [DOI] [PubMed] [Google Scholar]
- 7.Woehrer A, Bauchet L, Barnholtz-Sloan JS. Glioblastoma survival: has it improved? Evidence from population-based studies. Curr Opin Neurol. 2014;27(6):666–674. doi: 10.1097/WCO.0000000000000144. [DOI] [PubMed] [Google Scholar]
- 8.Bischoff SC. Quercetin: potentials in the prevention and therapy of disease. Curr Opin Clin Nutr Metab Care. 2008;11(6):733–740. doi: 10.1097/MCO.0b013e32831394b8. [DOI] [PubMed] [Google Scholar]
- 9.Formica JV, Regelson W. Review of the biology of Quercetin and related bioflavonoids. Food Chem Toxicol. 1995;33(12):1061–1080. doi: 10.1016/0278-6915(95)00077-1. [DOI] [PubMed] [Google Scholar]
- 10.Hollman PC, Katan MB. Absorption, metabolism and health effects of dietary flavonoids in man. Biomed Pharmacother. 1997;51(8):305–310. doi: 10.1016/s0753-3322(97)88045-6. [DOI] [PubMed] [Google Scholar]
- 11.Sakanashi Y, Oyama K, Matsui H, et al. Possible use of quercetin, an antioxidant, for protection of cells suffering from overload of intracellular Ca2+: a model experiment. Life Sci. 2008;83(5–6):164–169. doi: 10.1016/j.lfs.2008.05.009. [DOI] [PubMed] [Google Scholar]
- 12.Boots AW, Haenen GR, Bast A. Health effects of quercetin: from antioxidant to nutraceutical. Eur J Pharmacol. 2008;585(2–3):325–337. doi: 10.1016/j.ejphar.2008.03.008. [DOI] [PubMed] [Google Scholar]
- 13.Loke WM, Proudfoot JM, McKinley AJ, et al. Quercetin and its in vivo metabolites inhibit neutrophil-mediated low-density lipoprotein oxidation. J Agric Food Chem. 2008;56(10):3609–3615. doi: 10.1021/jf8003042. [DOI] [PubMed] [Google Scholar]
- 14.Davis JM, Murphy EA, McClellan JL, Carmichael MD, Gangemi JD. Quercetin reduces susceptibility to influenza infection following stressful exercise. Am J Physiol Regul Integr Comp Physiol. 2008;295(2):R505–R509. doi: 10.1152/ajpregu.90319.2008. [DOI] [PubMed] [Google Scholar]
- 15.Kumazawa Y, Kawaguchi K, Takimoto H. Immunomodulating effects of flavonoids on acute and chronic inflammatory responses caused by tumor necrosis factor alpha. Curr Pharm Des. 2006;12(32):4271–4279. doi: 10.2174/138161206778743565. [DOI] [PubMed] [Google Scholar]
- 16.Ono K, Nakane H, Fukushima M, Chermann JC, Barre-Sinoussi F. Differential inhibitory effects of various flavonoids on the activities of reverse transcriptase and cellular DNA and RNA polymerases. Eur J Biochem. 1990;190(3):469–476. doi: 10.1111/j.1432-1033.1990.tb15597.x. [DOI] [PubMed] [Google Scholar]
- 17.Gryglewski RJ, Korbut R, Robak J, Swies J. On the mechanism of antithrombotic action of flavonoids. Biochem Pharmacol. 1987;36(3):317–322. doi: 10.1016/0006-2952(87)90288-7. [DOI] [PubMed] [Google Scholar]
- 18.Kuo PC, Liu HF, Chao JI. Survivin and p53 modulate quercetin-induced cell growth inhibition and apoptosis in human lung carcinoma cells. J Biol Chem. 2004;279(53):55875–55885. doi: 10.1074/jbc.M407985200. [DOI] [PubMed] [Google Scholar]
- 19.Vijayababu MR, Arunkumar A, Kanagaraj P, Venkataraman P, Krishnamoorthy G, Arunakaran J. Quercetin downregulates matrix metalloproteinases 2 and 9 proteins expression in prostate cancer cells (PC-3) Mol Cell Biochem. 2006;287(1–2):109–116. doi: 10.1007/s11010-005-9085-3. [DOI] [PubMed] [Google Scholar]
- 20.Kim JY, Kim EH, Park SS, Lim JH, Kwon TK, Choi KS. Quercetin sensitizes human hepatoma cells to TRAIL-induced apoptosis via Sp1-mediated DR5 up-regulation and proteasome-mediated c-FLIPS down-regulation. J Cell Biochem. 2008;105(6):1386–1398. doi: 10.1002/jcb.21958. [DOI] [PubMed] [Google Scholar]
- 21.Mense SM, Hei TK, Ganju RK, Bhat HK. Phytoestrogens and breast cancer prevention: possible mechanisms of action. Environ Health Perspect. 2008;116(4):426–433. doi: 10.1289/ehp.10538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Ranelletti FO, Maggiano N, Serra FG, et al. Quercetin inhibits p21-RAS expression in human colon cancer cell lines and in primary colorectal tumors. Int J Cancer. 2000;85(3):438–445. [PubMed] [Google Scholar]
- 23.Chien SY, Wu YC, Chung JG, et al. Quercetin-induced apoptosis acts through mitochondrial- and caspase-3-dependent pathways in human breast cancer MDA-MB-231 cells. Hum Exp Toxicol. 2009;28(8):493–503. doi: 10.1177/0960327109107002. [DOI] [PubMed] [Google Scholar]
- 24.Galluzzo P, Martini C, Bulzomi P, et al. Quercetin-induced apoptotic cascade in cancer cells: antioxidant versus estrogen receptor alpha-dependent mechanisms. Mol Nutr Food Res. 2009;53(6):699–708. doi: 10.1002/mnfr.200800239. [DOI] [PubMed] [Google Scholar]
- 25.Lee TJ, Kim OH, Kim YH, et al. Quercetin arrests G2/M phase and induces caspase-dependent cell death in U937 cells. Cancer Lett. 2006;240(2):234–242. doi: 10.1016/j.canlet.2005.09.013. [DOI] [PubMed] [Google Scholar]
- 26.Yamashita N, Kawanishi S. Distinct mechanisms of DNA damage in apoptosis induced by quercetin and luteolin. Free Radic Res. 2000;33(5):623–633. doi: 10.1080/10715760000301141. [DOI] [PubMed] [Google Scholar]
- 27.Gulati N, Laudet B, Zohrabian VM, Murali R, Jhanwar-Uniyal M. The antiproliferative effect of Quercetin in cancer cells is mediated via inhibition of the PI3K-Akt/PKB pathway. Anticancer Res. 2006;26(2A):1177–1181. [PubMed] [Google Scholar]
- 28.Ruiz PA, Braune A, Holzlwimmer G, Quintanilla-Fend L, Haller D. Quercetin inhibits TNF-induced NF-kappaB transcription factor recruitment to proinflammatory gene promoters in murine intestinal epithelial cells. J Nutr. 2007;137(5):1208–1215. doi: 10.1093/jn/137.5.1208. [DOI] [PubMed] [Google Scholar]
- 29.Santos BL, Oliveira MN, Coelho PL, et al. Flavonoids suppress human glioblastoma cell growth by inhibiting cell metabolism, migration, and by regulating extracellular matrix proteins and metalloproteinases expression. Chem Biol Interact. 2015;242:123–138. doi: 10.1016/j.cbi.2015.07.014. [DOI] [PubMed] [Google Scholar]
- 30.Kiekow CJ, Figueiro F, Dietrich F, et al. Quercetin derivative induces cell death in glioma cells by modulating NF-kappaB nuclear translocation and caspase-3 activation. Eur J Pharm Sci. 2016;84:116–122. doi: 10.1016/j.ejps.2016.01.019. [DOI] [PubMed] [Google Scholar]
- 31.Pan HC, Jiang Q, Yu Y, Mei JP, Cui YK, Zhao WJ. Quercetin promotes cell apoptosis and inhibits the expression of MMP-9 and fibronectin via the AKT and ERK signalling pathways in human glioma cells. Neurochem Int. 2015;80:60–71. doi: 10.1016/j.neuint.2014.12.001. [DOI] [PubMed] [Google Scholar]
- 32.Ferrara N. VEGF and the quest for tumour angiogenesis factors. Nat Rev Cancer. 2002;2(10):795–803. doi: 10.1038/nrc909. [DOI] [PubMed] [Google Scholar]
- 33.Murphy G, Nagase H. Progress in matrix metalloproteinase research. Mol Aspects Med. 2008;29(5):290–308. doi: 10.1016/j.mam.2008.05.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Friedl P, Wolf K. Tumour-cell invasion and migration: diversity and escape mechanisms. Nat Rev Cancer. 2003;3(5):362–374. doi: 10.1038/nrc1075. [DOI] [PubMed] [Google Scholar]
- 35.Ferrara N, Gerber HP, LeCouter J. The biology of VEGF and its receptors. Nat Med. 2003;9(6):669–676. doi: 10.1038/nm0603-669. [DOI] [PubMed] [Google Scholar]
- 36.Simons M, Gordon E, Claesson-Welsh L. Mechanisms and regulation of endothelial VEGF receptor signalling. Nat Rev Mol Cell Biol. 2016;17(10):611–625. doi: 10.1038/nrm.2016.87. [DOI] [PubMed] [Google Scholar]
- 37.Roy R, Yang J, Moses MA. Matrix metalloproteinases as novel biomarkers and potential therapeutic targets in human cancer. J Clin Oncol. 2009;27(31):5287–5297. doi: 10.1200/JCO.2009.23.5556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Ranuncolo SM, Armanasco E, Cresta C, Bal De Kier Joffe E, Puricelli L. Plasma MMP-9 (92 kDa-MMP) activity is useful in the follow-up and in the assessment of prognosis in breast cancer patients. Int J Cancer. 2003;106(5):745–751. doi: 10.1002/ijc.11288. [DOI] [PubMed] [Google Scholar]
- 39.La Rocca G, Pucci-Minafra I, Marrazzo A, Taormina P, Minafra S. Zymographic detection and clinical correlations of MMP-2 and MMP-9 in breast cancer sera. Br J Cancer. 2004;90(7):1414–1421. doi: 10.1038/sj.bjc.6601725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Pratheeshkumar P, Budhraja A, Son YO, et al. Quercetin inhibits angio-genesis mediated human prostate tumor growth by targeting VEGFR-2 regulated AKT/mTOR/P70S6K signaling pathways. PLoS One. 2012;7(10):e47516. doi: 10.1371/journal.pone.0047516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Lai WW, Hsu SC, Chueh FS, et al. Quercetin inhibits migration and invasion of SAS human oral cancer cells through inhibition of NF-kappaB and matrix metalloproteinase-2/-9 signaling pathways. Anticancer Res. 2013;33(5):1941–1950. [PubMed] [Google Scholar]