

Citizen science, big data, and a habitat marketplace enable dynamic habitat for migratory birds in California’s Central Valley.
Abstract
In an era of unprecedented and rapid global change, dynamic conservation strategies that tailor the delivery of habitat to when and where it is most needed can be critical for the persistence of species, especially those with diverse and dispersed habitat requirements. We demonstrate the effectiveness of such a strategy for migratory waterbirds. We analyzed citizen science and satellite data to develop predictive models of bird populations and the availability of wetlands, which we used to determine temporal and spatial gaps in habitat during a vital stage of the annual migration. We then filled those gaps using a reverse auction marketplace to incent qualifying landowners to create temporary wetlands on their properties. This approach is a cost-effective way of adaptively meeting habitat needs for migratory species, optimizes conservation outcomes relative to investment, and can be applied broadly to other conservation challenges.
INTRODUCTION
The pace and magnitude of global change test the resilience of ecological and social systems on many fronts (1, 2). Climatic change is upending traditional land uses and creating unpredictability across the spectrum of urban, agricultural, and natural ecosystems (3–7). Reports of changes in species range shifts and phenologies are now ubiquitous [for example, study by Chen et al. (8)]. Existing protected areas may be increasingly insufficient to provide the amount and distribution of habitat needed by species (9). Migratory species may be especially vulnerable, given their reliance on finding suitable habitat across vast areas that may also span an array of human use intensities (10). A recent study has shown that just 9% of 1451 migratory bird species are adequately covered by protected areas across all stages of their annual cycle (11). Inadequate protection, along with habitat loss and degradation and climate change, is a contributing factor to the decline of more than half of the migratory bird species across all major flyways in the last 30 years (12).
The need to complement networks of permanently protected areas with suitable habitat outside of those areas is well established (13). For many transient species, this may also require creating, enhancing, or retaining habitat conditions on a temporary basis, such as by engineering adequate hydrologic flows at critical life stages of anadromous fish (14). However, to be able to adaptively provision habitat when and where migratory species most need it, conservationists need to surmount at least three substantive challenges. They must be able to (i) predict where the species will be over the course of the year, (ii) identify areas that are suitable for the migrants or that can be modified to make them suitable, and (iii) create cost-effective mechanisms to ensure that the habitat will be there when the species arrive.
Fortunately, recent advances in big data analytics, remote sensing, citizen science, and market-based conservation interventions can be deployed to unlock more dynamic approaches to adaptively meet conservation needs that are transient in space and time (15–18). These strategies, for example, can seek to “rent” habitat from private landowners through short-term agreements, rather than having government agencies or conservation organizations “own” the habitat and all of its associated management costs. Dynamic conservation strategies have been implemented in marine systems [for example, the studies reviewed by Lewison et al. (19)], but have had limited application in terrestrial systems [notable examples include Australia’s Bush Tender Program (20) and the U.S. Department of Agriculture’s Conservation Reserve Program (21)]. Dynamic conservation may offer important advantages over static protected area strategies (22), especially for migratory species (23). Potential advantages include the ability to adjust the timing, extent, and location of provisioned habitat to better match species’ full life-cycle needs (for example, breeding, migration, stopover), and to adapt to climate change, droughts, habitat conversion, and other threats. Dynamic conservation strategies may also have greater scalability and cost-effectiveness because temporary habitat enhancements can be less expensive than permanent protection. With increased cost-effectiveness short-term conservation investments can be adapted to fluctuations in funding and need. Moreover, private landowners may be more willing to enter short-term conservation agreements than to sell permanent interests in their property.
Here, we provide evidence that a dynamic approach, when coupled with precise information about the species’ distributions and habitat, can be a cost-effective, scalable, and adaptable conservation strategy for migratory birds. We conducted our study in the Sacramento Valley, the northernmost region of the Central Valley of California (USA), with rice farmers who were paid to create temporary habitat for migratory shorebirds, a globally declining group of species (24) that have unmet habitat needs in our study area (25).
The Central Valley is typical of wetland-dominated temperate regions in both its importance to migrating and wintering waterbirds (shorebirds, waterfowl, and wading birds) and its high degree of alteration (Fig. 1). It is among the most important regions for waterbirds in North America (25, 26), with nearly 3 million ducks, 1 million geese, and 500,000 shorebirds overwintering annually, and many more using the region as a migratory stopover (26, 27). With the loss of over 90% of the original 16,000 km2 of wetlands and riparian areas, largely to agriculture and urbanization (25), migratory waterbirds in the Central Valley are dependent on a limited network of permanently protected refuges and, increasingly, on compatible management of agricultural lands for habitat (28, 29).
Fig. 1. California’s Sacramento Valley wetlands, rice, and protected areas.
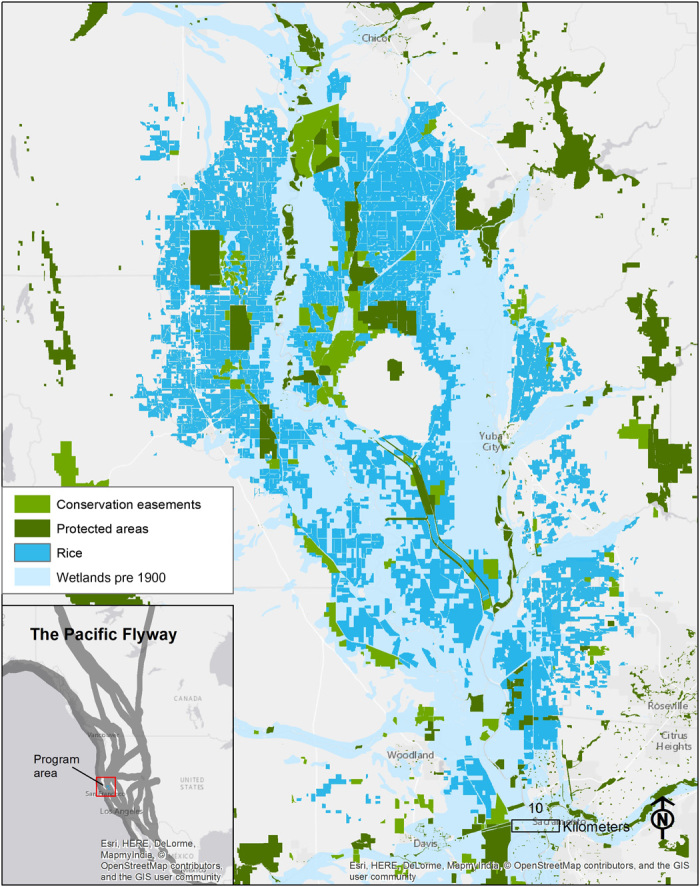
Habitat auction program area showing historic wetlands, current rice-growing extent, and protected areas. Inset shows the Sacramento Valley location within California and the Pacific Flyway.
Rice is grown on over 2000 km2 of the Sacramento Valley, accounting for about 23% of land coverage (Fig. 1) (25). Rice fields are flooded during the growing season; many are also flooded during the fall and winter to decompose rice stubble from the previous year’s crop (30). Shallow water during flood-up and draw-down periods for stubble management can help meet habitat needs of shorebirds and other waterbirds during migration (28).
RESULTS
Understanding temporal variation in species’ distributions is critical for developing dynamic conservation strategies (31). To prioritize potential conservation actions in space and time, we used predictive models of shorebird species’ occurrences and abundances developed with data from eBird, an online checklist program of citizen science bird observations (32–34). Predicted shorebird occurrences, abundances, distribution ranges, and arrival and departure dates in the Sacramento Valley were highly seasonal, as expected based on natural history, with peaks during fall and spring migration (Fig. 2).
Fig. 2. Average estimated seasonal shorebird abundance in the Sacramento River Valley.
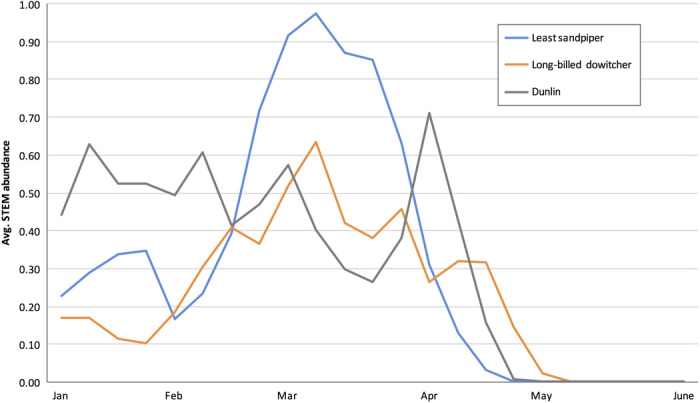
Weekly STEM model abundance estimates for the least sandpiper (blue), long-billed dowitcher (orange), and dunlin (gray) averaged across the Sacramento River Valley. Abundance estimates are reported in units of the expected number of birds of the given species reported by a typical eBird participant on a search from 0700 to 0800 hours while traveling 1 km.
Shorebirds depend on shallow water (<10 cm deep) habitats for foraging (28, 35). Analysis of satellite imagery shows that surface water in the Sacramento Valley is relatively plentiful in winter, declines in early spring, and is scarce in summer and fall (36). This pattern is driven largely by postharvest flooding of rice fields and management of duck clubs and wildlife refuges for hunting and waterfowl food production (25). Recent drought conditions in California have reduced surface water availability in fall and spring by more than 20% on average, further restricting waterbird habitat (36).
We combined predicted shorebird abundance values with predicted surface water for winter through early spring (January to April) to identify times and locations that temporary wetlands could deliver potentially high-value shorebird habitat (Fig. 3). Model results indicated that the highest conservation values for shorebird habitat enhancements were in late winter and early spring (February through April) and in three general areas within the Sacramento Valley.
Fig. 3. Estimated shorebird abundance, surface water availability, and conservation value in the Sacramento Valley for February.

(A) Relative abundance of shorebirds. (B) Probability of surface water based on NASA Landsat imagery. (C) Ranked conservation value in terms of potential shorebird habitat. Areas of high shorebird predictions and low water availability represent highest conservation value in terms of potential shorebird habitat return on investment. Highest conservation value (dark blue), moderate conservation value (light blue), existing refuges, and maintain status quo areas (green).
In 2014, The Nature Conservancy (TNC), a nongovernmental conservation organization, implemented a program to encourage rice farmers to flood fields, and thereby provide habitat, during time periods when our models predicted high shorebird abundances and limited water availability for habitat. The program used a reverse auction (20, 37) in which a single buyer (in this case, TNC) purchases goods or services from multiple suppliers (that is, the farmers). TNC invited farmers to submit bids representing their costs to flood fields at a specified depth (5 to 10 cm) and duration (4-, 6-, or 8-week periods, each beginning 1 February 2014). The reverse auction allowed farmers to set their own price for providing habitat, and the differences in bids likely reflected cost differences farmers bear to provision water depending on their water rights, management practices, water districts, and on-farm infrastructure. The reverse auction approach enabled TNC to select the greatest amount and duration of high-quality habitat for the lowest total cost.
Ensuring confidentiality of bids was essential to building farmers’ trust in this program. In addition, because this program is ongoing, we cannot report actual awards to farmers, as this would compromise future competitive bidding processes. Thus, here, we have transformed actual costs to a relative cost distribution where average price per bid was standardized to equal 100 “cost units” for comparisons.
TNC received 55 bids across all three duration options, totaling 49.9 km2 of potential wetland habitat (Table 1). Within each enrollment duration, TNC selected the winning bids based on the modeled habitat value (predicted shorebird abundance and surface water availability) per dollar invested versus a simple price per acre. TNC included additional fields in the final set of winning bids to ensure representation across the Sacramento Valley and each of the three duration options.
Table 1. Spring 2014 habitat auction participation.
| n farmers | n bids* | Total (km2) | Average (km2 per bid) | |
| 4-Week | 30 | 37 | 31.5 | 0.85 |
| 6-Week | 11 | 12 | 12.6 | 1.05 |
| 8-Week | 6 | 6 | 5.8 | 0.97 |
| Total | 37* | 55 | 49.9 | 0.91 |
*Individual farmers could submit bids for multiple duration options.
TNC accepted 44 of the 55 bids (80.0%; Table 2) and contracted for 38.9 km2 of habitat. Bid prices per square kilometer varied by 67% between the lowest and highest bids (Table 2). Average price per square kilometer of accepted bids was significantly less than that by rejected bids by 60% (mean price accepted versus rejected bids = 54.2 versus 132.5 relative cost units; F test for two sample variances, F = 0.03, P < 0.01, df = 7, 43). There were fewer bids, and bid prices per square kilometer were higher for the 6- and 8-week programs than for the 4-week program, reflecting both the higher value of shorebird habitat at this time (and hence TNC’s willingness to pay more) and the increasing risk to farmers of providing habitat later in the season, which has the potential to delay planting and decrease crop value [average bid price per square kilometer 4-week program, 89.7 relative cost units; 6-week program, 96.9 relative cost units; and 8-week program, 170.0 relative cost units; analysis of variance (ANOVA), F = 4.09, P < 0.02, df = 54; Table 2].
Table 2. Relative cost per square kilometers per bid.
| n |
Average relative cost per bid |
SD | Coefficient of variation | |
| Bids | 55 | 100.00 | 67.41 | 67.41 |
| Bids accepted | 44 | 80.12 | 22.21 | 27.72 |
| Bids rejected | 11 | 331.74 | 554.96 | 167.29 |
To evaluate shorebird responses to the temporary flooding program, we surveyed shorebird use of enrolled fields (participating in temporary flood management for shorebirds) and a sample of unenrolled fields (we asked participating farmers for access to unenrolled fields to monitor bird use; necessary permission limited our sample size of unenrolled fields). These unenrolled fields were simply comparable fields not involved in the temporary flooding program and included a range of conditions from dry to flooded. We observed high bird use of temporary wetlands and recorded more than 180,000 observations of birds representing 57 different species using the enrolled fields in spring of 2014. On average, shorebird species richness was more than three times greater and average shorebird density was five times greater on enrolled fields than unenrolled fields [mean shorebird species richness per survey = 0.21 versus 0.67 (P < 0.001) and mean density per hectare = 0.12 versus 0.58 (P < 0.001 per survey for unenrolled and enrolled fields, respectively); bootstrap significance test as described in Materials and Methods; sample sizes = 170 survey points in 38.9 km2 of enrolled fields surveyed 8.6 ± 2.9 SD times and 48 survey points in 9.9 km2 of unenrolled fields surveyed 10.1 ± 3.3 SD times] (Fig. 4). Shorebird densities documented in our program were an order of magnitude greater than those reported for mid-winter flooded rice fields (35), indicating the high bird conservation value of provisioning habitat during spring migration.
Fig. 4. Observed shorebird densities and species richness.
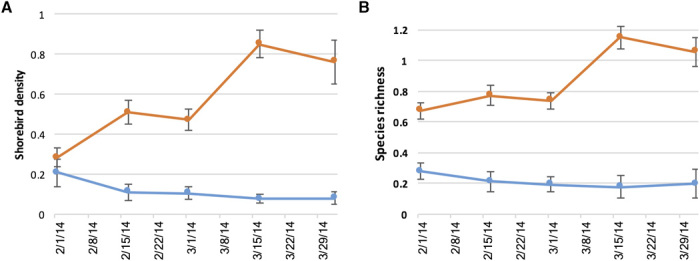
Biweekly shorebird densities per hectare (A) and species richness (B) in enrolled fields (orange) and unenrolled fields (blue) for 2 February to 12 April 2014 (mean ± SEM).
To contrast the performance of this dynamic conservation strategy with a more traditional protected land strategy, we compared the cost of providing 38.9 km2 of habitat for 8 weeks during February through March each year using a temporary habitat reverse auction, as we did in our pilot, against the estimated cost of purchasing 38.9 km2 of the Sacramento Valley rice-farming land and retiring it from production. The highest possible estimate of the total cost per year of the dynamic approach is $1.4 million. This figure is based on the average bid received, which is significantly higher than the average winning bid (which was the price paid by TNC). In contrast, the estimated cost to buy the same area of land based on current land values is roughly $150 million, the estimated cost of restoration is roughly $25 million, and the estimated ongoing annual maintenance costs are roughly $100,000 (property maintenance and staffing) (38). Assuming a U.S. standard constant discount rate of 7.0%, the real, pretax average return on private investment (39) used for cost-benefit analysis by the U.S. Office of Management and Budget (40), the cost of providing temporary habitat for 8 weeks every year for 25 years represents just 9.9% of the cost of acquiring, restoring, and maintaining those acres for in-perpetuity conservation management (11.7% for 50 years and 12.1% for 100 years). Even at a 3% discount rate, the consumption rate of discount, the cost of renting is less than 15% of the cost of purchasing. The dynamic approach is significantly cheaper because payments are spread over a much longer period, whereas static conservation approaches typically require a large up-front capital investment. In addition, the dynamic program compensates farmers only for the targeted conservation needs, without a loss of agricultural production, whereas static conservation usually involves purchase of the entire suite of property rights.
Even with the obvious economic efficiency, we caution that dynamic conservation is best considered as an additional tool and not a replacement for permanent protection, especially in areas of high, year-round conservation value. If land use in the region were to change to agricultural or other practices not compatible with seasonal flooding, then permanent protection strategies, such as land acquisition and habitat restoration, may be required. However, we note that programs providing payment for ecosystem services could help ensure the economic viability of conservation-compatible land uses (41).
DISCUSSION
The dynamic strategy we piloted offers a promising new “tool in the toolbox” for advancing conservation of biodiversity in a changing world. It may be particularly valuable for developing and implementing conservation plans for migratory species, whose dispersed habitat needs often test the limits of more traditional conservation approaches. The precision and efficiency of dynamic approaches, such as ours, are made possible by access to near real-time, high-quality citizen science and remotely sensed data streams, and the ability to rapidly process those data to develop predictive models. Those models can be used to generate near-term habitat priorities that, in turn, market-based tools, such as reverse auctions, can cost-effectively and temporarily secure. In concert, these elements comprise a strategy that can be readily scaled. Since 2014, TNC’s temporary migratory bird habitat program has been expanded to over 200 km2 of wetland habitat in California including multiple seasons, and additional species groups, regions, and crop types.
Similar to what the sharing economy has done in transportation and lodging (42), our model of renting habitat complements the more traditional approach of permanent conservation, by meeting temporary needs using an adaptive and agile approach. We expect dynamic conservation strategies to become increasingly important for biodiversity conservation, especially as a means of facilitating adaptation to climate change and its concomitant variability and extremes, such as extended drought (43, 44). Spatially and temporally targeted conservation outcomes similar to those that we provided for birds could benefit myriad species with large-scale and transient habitat requirements, such as anadromous fish and migratory marine and land mammals (14, 19, 45). Dynamic conservation strategies also can help extend the impact of scarce conservation dollars and, importantly, engage citizen scientists and private landowners, all of which may be necessary if we are to achieve conservation outcomes at the scale needed to protect migratory phenomena.
MATERIALS AND METHODS
Bird species abundance modeling
We estimated the relative abundance of three shorebird species (long-billed dowitcher Limnodromus scolopaceus, least sandpiper Calidris minutilla, and dunlin Calidris alpina) based on bird-monitoring data from eBird (32) and local land cover and elevation data from NASA satellites. The eBird data used for analysis were restricted to searches from “stationary” or “traveling” protocols within the state of California collected between 1 January 2007 and 31 December 2012, where observers recorded counts of all bird species they detected and identified during each search (n = 232,175 search checklists, of which 23,131 were withheld for model validation). Each checklist was associated with spatial and temporal covariates to describe the ecological processes governing population abundance and search effort covariates to describe the observation process. To account for variation in detectability associated with search effort, we included covariates for the search duration, the distance traveled during the search, whether the observer was stationary or traveled, and the number of people in the search party. Environmental covariates were elevation (46) and annual land cover from the Cropland Data Layer (47), the latter of which also indirectly reflects climatic variation. This land cover data set contains 131 crop and other land cover classifications (including surface water and wetlands), enough to model habitat relationships for a broad range of species. Annual land cover data were summarized as proportional cover of each category within 3 × 3 km squares centered on each checklist location. Time was included at two scales: the time of day, capturing differences in behavior of birds and availability for detection, and the day of the year, describing seasonal trends including migratory bird movements.
We used the spatiotemporal exploratory model (STEM) to estimate species’ abundance because of its ability to adapt to spatial and temporal variation in predictor-response relationships modeled from large sets of irregularly distributed observational data (33, 48). STEM is an ensemble of local regression models generated by repeatedly partitioning the study extent into 200 randomized grids of spatiotemporal blocks, called stixels, and then fitting independent regression models, called base models, within all stixels. To capture the spatial variation among stixels at the scale of the Sacramento Valley and the temporal variation throughout the annual cycle, we used stixel dimensions of 3° longitude, 3° latitude, and 40 contiguous days. Within each stixel, species’ abundance was assumed to be stationary, and we fit a zero-inflated boosted regression tree model (34) as the base model to capture the spatial and temporal variation in abundance as a function of the covariate data within the stixel. Using the fact that stixels from this randomized design overlap, abundance predictions at a specified location and time were made by taking an average across all base models that include location and time. Combining estimates across the ensemble controls for intermodel variability and adapts to nonstationary predictor-response relationships.
We separately modeled abundances of each of the three species and averaged abundance predictions. Predictions of abundance were calculated as the expected number of birds of a given species by a typical eBird participant on a search from 0700 to 0800 hours while traveling 1 km at a given location and day of the year. Predictive performance was assessed on validation data and found to have good ability to rank observed abundance [see the study by Johnston et al. (34) for details]. Abundance predictions were computed across a 3 × 3–km grid of locations spanning the Sacramento Valley at weekly intervals. These predictions were used to create spatial surfaces of conservation value for each of the three shorebirds every week. All models and predictions were run in R (49) with package “gbm” (49, 50). Johnston et al. (34) present the complete abundance modeling methodology.
We chose these species because they are wide-ranging and relatively common, which increased the quantity of citizen science data and, consequently, the robustness of the predictive models. We assumed that these species, which span the range of body sizes and physiological requirements for shorebirds—from relatively large (long-billed dowitcher), medium (dunlin), to small size (least sandpiper)—would represent the habitat needs of the >15 species of shorebirds that depend on stopover and wintering habitat in the Central Valley (27).
Surface water habitat modeling
We acquired 30-m resolution satellite imagery of the Central Valley taken by the Landsat 5 Thematic Mapper (http://earthexplorer.usgs.gov/). We downloaded all seven bands of five scene footprints collected in January through April 2000 and 2011. To develop predictive models of open-water occurrence, we used 11,675 ground survey observations of agricultural fields [n = 3904 (35, 51, and Point Blue Conservation Science, unpublished data)] and wetlands (n = 7771; Sacramento Valley National Wildlife Refuge Complex, unpublished data), as well as 8690 manually classified points to specify known areas of open surface water (lakes, rivers/streams, and canals; n = 2094) and areas with no open water (streets, parking lots, buildings, forests, and mountains; n = 6596). We matched all the ground-truth data with a satellite scene from the closest date to the observation and spatially extracted the observed percentage of water coverage from the ground survey area (polygons) to each spatially corresponding pixel in the satellite scene to develop a data set to train and then test our models through cross-validation (52).
We developed a boosted regression tree model (53), where the response variable was 1 if the estimated percentage from the ground-truth data was ≥75% open water or 0 if <10% open water. The predictor variables were the mean spectral reflectance values for each band across all pixels within the ground survey area (most survey areas had 10 to 20, 30-m pixels). We weighted the observed data in the model by the inverse of the number of days between the field observation and the most recent satellite scene.
On the basis of preliminary comparisons, we pooled data from all subregions and years to calibrate our model and then used the full model to predict the probability of open surface water to all pixels in all scenes. Image analysis and classification modeling were performed primarily using the “raster” and “dismo” packages in R (54, 55) and Arc Map 9.3.1 (Esri Inc. 2009). Reiter et al. (36) present complete methods for surface water habitat modeling.
Estimating habitat conservation value for shorebirds
We ranked areas of the Sacramento Valley as high, medium, or low for predicted shorebird abundance for each week from January to April. We similarly ranked surface water habitat availability. The shorebird abundance models, which include coarse wetland and surface water variables, estimate where numbers of shorebirds could be at a given date. The surface water habitat models, which incorporate a large amount of ground-truth data and satellite image analysis, provide an accurate estimate of surface water habitat at a given location and date. Combining these layers, we identified times and places of potential shorebird habitat conservation value in which high predicted shorebird abundance coincided with low predicted average surface water habitat availability.
Habitat auction design
TNC implemented a two-round, closed, discriminatory price auction in which a farmer’s bid was evaluated in terms of habitat value per dollar invested, using the bid price and TNC’s habitat rankings. Prices were not revealed to other bidders, and for accepted bids, farmers were paid the value of their bids. This contrasts with a uniform payment scheme in which all winning farmers receive the same price.
Relative bid cost distribution
To ensure confidentiality of bid prices in the habitat auction and to allow for comparison between prices for accepted and rejected bids and among program duration options, we converted actual prices bid in USD/km2 to a relative cost unit where the average price per bid was standardized to equal 100 cost units (for example, Xi rel = (Xi/Xmean) × 100, where Xi rel is the bid price in “relative cost units,” Xi is the bid price i in USD/km2, and Xmean is the average bid price in USD/km2).
Monitoring shorebird response
To evaluate shorebird responses to the temporary flooding program, we monitored responses of shorebirds in enrolled fields (participating in temporary flood management for shorebirds) and unenrolled fields. We asked participating farmers for access to unenrolled fields to monitor bird use. These unenrolled fields were simply comparable fields not involved in the temporary flooding program and included a range of conditions from dry to flooded. Necessary permission limited our access to a smaller number of unenrolled fields than enrolled fields. We used a spatially balanced survey design (Generalized Random Tessellation Stratified Sampling) (56) of sampling locations arranged systematically across enrolled and unenrolled fields. Sample points were visited at least weekly between 2 February and 11 April 2014, and species and numbers of birds were counted within 200 m of sample points using standard shorebird census protocols (35). We monitored a total of 38.9 km2 of enrolled fields and 9.9 km2 of unenrolled fields with 218 survey points (170 points in enrolled fields and 48 points in unenrolled fields) that were surveyed 1934 times (mean number of times surveyed 8.6 ± 2.9 SD times for enrolled fields and 10.1 ± 3.3 SD times for unenrolled fields).
Shorebird density and species richness
Observations and identifications of shorebird species were recorded from each survey point. The density of shorebirds was computed as the total number of observations of all shorebirds divided by the area (in hectares) of the survey polygon around each survey point. Density was then transformed using the natural logarithm [log(x + 1)] for linearization of data. Species richness was computed using the “specnumber” function of the “vegan” package in R (49).
We created a bootstrapping algorithm to test the effects of the program on each shorebird response metric (density and species richness). Because our data were effectively time series data from an open population, we needed a test that did not require any a priori assumptions to be met but could still test the main effect of the program. Therefore, we created our own null distribution that would not require any assumptions, using as the test statistic the total number of days in which the average bird response was greater on enrolled fields than on unenrolled fields. We sampled data that were pooled by date, with replacement, per the number of surveys taken on each date for enrolled and unenrolled fields, respectively. The bird response data were then averaged for each group. Next, we scored the total number of days in which there was a higher average value (of density or species richness) on enrolled versus unenrolled fields. This process was repeated 1000 times to create a null distribution of the number of days that enrolled fields were greater than unenrolled fields in whichever bird response variable was being tested. If the observed value occurred less than 5% of the time due to random chance, that value was considered as statistically significant. Shorebird densities in enrolled fields exceeded those in unenrolled fields on 52 of 61 survey days (P < 0.001; critical value at 95% = 37 days). Shorebird species richness in enrolled fields exceeded that in unenrolled fields on 52 of 61 survey days (P < 0.001; critical value at 95% = 36 days).
Acknowledgments
We thank S. Heard and M. Ochoa for help with the analyses, G. LeBuhn for reviewing a previous version of the manuscript and the participating farmers, the California Rice Commission, Audubon California, TNC’s field assistants, and the legion of citizen scientists who contribute data to eBird. Funding: Funding for this project was provided by TNC and the S. D. Bechtel, Jr. Foundation, with additional support to Cornell Lab of Ornithology from the Leon Levy Foundation, the Seaver Foundation, the Wolf Creek Foundation, NASA grant (NNH12ZDA001N-ECOF), and NSF grants ABI-7388506 and CCF-1522054 with NSF computing support from CNS-1059284 and DEB-110008. Author contributions: M.D.R., E.H., S.M., P.S., and S.A.M. conceived the conservation program. B.L.S., D.F., A.J., W.M.H., N.E.B., S.K., and J.W.F. developed the eBird models. G.H.G. led the field data collection. M.E.R., S.V., N.E., and C.H. developed the surface water habitat availability models. L.M. helped design the auction. M.D.R., G.H.G., C.L., and E.H. performed the analyses, and M.D.R. wrote the manuscript. D.F., M.M., and C.M. assisted with key elements of the analysis. All authors contributed ideas to the manuscript. Competing interests: The authors declare that they have no competing interests. Data and materials availability: The data reported in this paper are tabulated and archived at TNC, Point Blue Conservation Science, and Cornell Lab of Ornithology and are available from the authors upon request.
REFERENCES AND NOTES
- 1.Crutzen P. J., Geology of mankind. Nature 415, 23 (2002). [DOI] [PubMed] [Google Scholar]
- 2.Liu J., Dietz T., Carpenter S. R., Alberti M., Folke C., Moran E., Pell A. N., Deadman P., Kratz T., Lubchenco J., Ostrom E., Ouyang Z., Provencher W., Redman C. L., Schneider S. H., Taylor W. W., Complexity of coupled human and natural systems. Science 317, 1513–1516 (2007). [DOI] [PubMed] [Google Scholar]
- 3.Intergovernmental Panel on Climate Change, Climate Change, 2014–Impacts, Adaptation and Vulnerability: Regional Aspects (Cambridge Univ. Press, 2014). [Google Scholar]
- 4.Grimm N. B., Faeth S. H., Golubiewski N. E., Redman C. L., Wu J., Bai X., Briggs J. M., Global change and the ecology of cities. Science 319, 756–760 (2008). [DOI] [PubMed] [Google Scholar]
- 5.Vermeulen S. J., Campbell B. M., Ingram J. S. I., Climate change and food systems. Annu. Rev. Env. Resour. 37, 195–222 (2012). [Google Scholar]
- 6.Zabel F., Putzenlechner B., Mauser W., Global agricultural land resources – A high resolution suitability evaluation and its perspectives until 2100 under climate change conditions. PLOS ONE 9, e107522 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Thomas C. D., Cameron A., Green R. E., Bakkenes M., Beaumont L. J., Collingham Y. C., Erasmus B. F. N., de Siqueira M. F., Grainger A., Hannah L., Hughes L., Huntley B., van Jaarsveld A. S., Midgley G. F., Miles L., Ortega-Huerta M. A., Peterson A. T., Phillips O. L., Williams S. E., Extinction risk from climate change. Nature 427, 145–148 (2004). [DOI] [PubMed] [Google Scholar]
- 8.Chen I.-C., Hill J. K., Ohlemüller R., Roy D. B., Thomas C. D., Rapid range shifts of species associated with high levels of climate warming. Science 333, 1024–1026 (2011). [DOI] [PubMed] [Google Scholar]
- 9.Mora C., Sale P. F.. Ongoing global biodiversity loss and the need to move beyond protected areas: A review of the technical and practical shortcomings of protected areas on land and sea. Mar. Ecol. Prog. Ser. 434, 251–266 (2011). [Google Scholar]
- 10.Runge C., Martin T. G., Possingham H. P., Willis S. G., Fuller R. A., Conserving mobile species. Front. Ecol. Environ. 12, 395–402 (2014). [Google Scholar]
- 11.Runge C. A., Watson J. E. M., Butchart S. H. M., Hanson J. O., Possingham H. P., Fuller R. A., Protected areas and global conservation of migratory birds. Science 350, 1255–1258 (2015). [DOI] [PubMed] [Google Scholar]
- 12.Kirby J. S., Stattersfield A. J., Butchart S. H. M., Evans M. I., Grimmett R. F. A., Jones V. R., O’Sullivan J., Tucker G. M., Newton I., Key conservation issues for migratory land- and waterbird species on the world’s major flyways. Bird Conserv. Int. 18 (suppl 1), S49–S73 (2008). [Google Scholar]
- 13.Poiani K. A., Richter B. D., Anderson M. G., Richter H. E., Biodiversity conservation at multiple scales: Functional sites, landscapes, and networks. BioScience 50, 133–146 (2000). [Google Scholar]
- 14.Jager H. I., Rose K. A., Designing optimal flow patterns for fall chinook salmon in a Central Valley, California, River. N. Am. J. Fish. Manage. 23, 1–21 (2003). [Google Scholar]
- 15.Hampton S. E., Strasser C. A., Tewksbury J. J., Gram W. K., Budden A. E., Batcheller A. L., Duke C. S., Porter J. H., Big data and the future of ecology. Front. Ecol. Environ. 11, 156–162 (2013). [Google Scholar]
- 16.Turner W., Spector S., Gardiner N., Fladeland M., Sterling E., Steininger M., Remote sensing for biodiversity science and conservation. Trends Ecol. Evol. 18, 306–314 (2003). [Google Scholar]
- 17.Bonney R., Cooper C. B., Dickinson J., Kelling S., Phillips T., Rosenberg K. V., Shirk J., Citizen science: A developing tool for expanding science knowledge and scientific literacy. BioScience 59, 977–984 (2009). [Google Scholar]
- 18.Kroeger T., Casey F., An assessment of market-based approaches to providing ecosystem services on agricultural lands. Ecol. Econ. 64, 321–332 (2007). [Google Scholar]
- 19.Lewison R., Hobday A. J., Maxwell S., Hazen E., Hartog J. R., Dunn D. C., Briscoe D., Fossette S., O’Keefe C. E., Barnes M., Abecassis M., Bograd S., Bethoney N. D., Bailey H., Wiley D., Andrews S., Hazen L., Crowder L. B., Dynamic ocean management: Identifying the critical ingredients of dynamic approaches to ocean resource management. BioScience 65, 486–498 (2015). [Google Scholar]
- 20.Stoneham G., Chaudri V., Ha A., Strappazzon L., Auctions for conservation contracts: An empirical examination of Victoria’s BushTender trial. Aust. J. Agr. Resour. Ec. 47, 477–500 (2003). [Google Scholar]
- 21.USDA-FSA, Conservation Reserve Program Annual Summary and Enrollment Statistics FY 2012 (USDA-FSA, 2012).
- 22.Bengtsson J., Angelstam P., Elmqvist T., Emanuelsson U., Folke C., Ihse M., Moberg F., Nyström M., Reserves, resilience and dynamic landscapes. Ambio 32, 389–396 (2003). [DOI] [PubMed] [Google Scholar]
- 23.Bull J., Suttle K., Singh N. J., Milner-Gulland E., Conservation when nothing stands still: Moving targets and biodiversity offsets. Front. Ecol. Environ. 11, 203–210 (2013). [Google Scholar]
- 24.BirdLife International, Threatened Birds of the World 2008 (CD-ROM. BirdLife International, 2008).
- 25.Central Valley Joint Venture, Central Valley Joint Venture 2006 Implementation Plan: Conserving Bird Habitat (U.S. Fish and Wildlife Service, 2006).
- 26.D. P. Collins, C. A. Palmer, R. E. Trost, Pacific Flyway Data Book 2011: Waterfowl Harvests and Status, Hunter Participation and Success in the Pacific Flyway and United States (US Fish and Wildlife Service Division of Migratory Bird Management, 2011). [Google Scholar]
- 27.Shuford W., Page G. W., Kjelmyr J., Patterns and dynamics of shorebird use of California’s Central Valley. Condor 100, 227–244 (1998). [Google Scholar]
- 28.Elphick C. S., Oring L. W., Winter management of Californian rice fields for waterbirds. J. Appl. Ecol. 35, 95–108 (1998). [Google Scholar]
- 29.Stralberg D., Cameron D. R., Reynolds M. D., Hickey C. M., Klausmeyer K., Busby S. M., Stenzel L. E., Shuford W. D., Page G. W., Identifying habitat conservation priorities and gaps for migratory shorebirds and waterfowl in California. Biodivers. Conserv. 20, 19–40 (2011). [Google Scholar]
- 30.Bird J. A., Pettygrove G. S., Eadie J. M., The impact of waterfowl foraging on the decomposition of rice straw: Mutual benefits for rice growers and waterfowl. J. Appl. Ecol. 37, 728–741 (2000). [Google Scholar]
- 31.R. Greenberg, P. Marra, Birds of Two Worlds: The Ecology and Evolution of Migration (Johns Hopkins Univ. Press, 2005). [Google Scholar]
- 32.Sullivan B. L., Aycrigg J. L., Barry J. H., Bonney R. E., Bruns N., Cooper C. B., Damoulas T., Dhondt A. A., Dietterich T., Farnsworth A., Fink D., Fitzpatrick J. W., Fredericks T., Gerbracht J., Gomes C., Hochachka W. M., Iliff M. J., Lagoze C., La Sorte F. A., Merrifield M., Morris W., Phillips T. B., Reynolds M., Rodewald A. D., Rosenberg K. V., Trautmann N. M., Wiggins A., Winkler D. W., Wong W.-K., Wood C. L., Yu J., Kelling S., The eBird enterprise: An integrated approach to development and application of citizen science. Biol. Conserv. 169, 31–40 (2014). [Google Scholar]
- 33.Fink D., Hochachka W. M., Zuckerberg B., Winkler D. W., Shaby B., Munson M. A., Hooker G., Riedewald M., Sheldon D., Kelling S., Spatiotemporal exploratory models for broad-scale survey data. Ecol. Appl. 20, 2131–2147 (2010). [DOI] [PubMed] [Google Scholar]
- 34.Johnston A., Fink D., Reynolds M. D., Hochachka W. M., Sullivan B. L., Bruns N. E., Hallstein E., Merrifield M. S., Matsumoto S., Kelling S., Abundance models improve spatial and temporal prioritization of conservation resources. Ecol. Appl. 25, 1749–1756 (2015). [DOI] [PubMed] [Google Scholar]
- 35.Strum K., Reiter M. E., Hartman C. A., Iglecia M. N., Kelsey T. R., Hickey C. M., Winter management of California’s rice fields to maximize waterbird habitat and minimize water use. Agric. Ecosyst. Environ. 179, 116–124 (2013). [Google Scholar]
- 36.Reiter M. E., Elliott N., Veloz S., Jongsomjit D., Hickey C. M., Merrifield M., Reynolds M. D., Spatio-temporal patterns of open surface water in the Central Valley of California 2000-2011: Drought, land cover, and waterbirds. J. Am. Water Resour. Assoc. 51, 1722–1738 (2015). [Google Scholar]
- 37.Latacz-Lohmann U., Van der Hamsvoort C., Auctioning conservation contracts: A theoretical analysis and an application. Am. J. Agric. Econ. 79, 407–418 (1997). [Google Scholar]
- 38.C. Loesch, D. Hellerstein, D. Nulph, J. Williamson, L. Hansen, C. Loesch, M. Ribaudo, W. Crumpton, “Targeting investments to cost effectively restore and protect wetland ecosystems: Some economic insights” (Economic Research Report ERR-183, U.S. Department Agriculture, Economic Research Service, 2015). [Google Scholar]
- 39.Arrow K., Cropper M., Gollier C., Groom B., Heal G., Newell R., Nordhaus W., Pindyck R., Pizer W., Portney P., Sterner T., Toll R. S. J., Weitzman M., Determining benefits and costs for future generations. Science 341, 349–350 (2013). [DOI] [PubMed] [Google Scholar]
- 40.U. S. Office of Management and Budget, Circular A-4: Regulatory Analysis (OMB, 2003).
- 41.Daily G. C., Matson P. A., Ecosystem services: From theory to implementation. Proc. Natl. Acad. Sci. U.S.A. 105, 9455–9456 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Cramer J., Krueger A. B., Disruptive change in the taxi business: The case of Uber. Am. Econ. Rev. 106, 177–182 (2016). [Google Scholar]
- 43.Dawson T. P., Jackson S. T., House J. I., Prentice I. C., Mace G. M., Beyond predictions: Biodiversity conservation in a changing climate. Science 332, 53–58 (2011). [DOI] [PubMed] [Google Scholar]
- 44.Bonebrake T. C., Brown C. J., Bell J. D., Blanchard J. L., Chauvenet A., Champion C., Chen I., Clark T. D., Colwell R. K., Danielsen F., Dell A. I., Donelson J. M., Evengård B., Ferrier S., Frusher S., Garcia R. A., Griffis R. B., Hobday A. J., Jarzyna M. A., Lee E., Lenoir J., Linnetved H., Martin V. Y., McCormack P. C., McDonald J., McDonald-Madden E., Mitchell N., Mustonen T., Pandolfi J. M., Pettorelli N., Possingham H., Pulsifer P., Reynolds M., Scheffers B. R., Sorte C. J. B., Strugnell J. M., Tuanmu M., Twiname S., Vergés A., Villanueva C., Wapstra E., Wernberg T., Pecl G. T., Management of climate-driven species redistribution consequences requires the integration of ecology, conservation and social science. Biol. Rev. 10.1111/brv.12344 (2017). [DOI] [PubMed] [Google Scholar]
- 45.Berger J., The last mile: How to sustain long-distance migration in mammals. Conserv. Biol. 18, 320–331 (2004). [Google Scholar]
- 46.T. Tachikawa, M. Hato, M. Kaku, A. Iwasaki, The characteristics of ASTER GDEM version 2, in Proceedings of the 2011 IEEE International Geoscience and Remote Sensing Symposium (IGARSS) (IEEE, 2011), pp. 3657–3660. [Google Scholar]
- 47.USDA-NASS, USDA National Agricultural Statistics Service Cropland Data Layer (USDA, 2013).
- 48.Fink D., Damoulas T., Bruns N. E., La Sorte F. A., Hochachka W. M., Gomes C. P., Kelling S., Crowdsourcing meets ecology: Hemisphere-wide spatiotemporal species distribution models. AI Mag. 35, 19–30 (2014). [Google Scholar]
- 49.R Core Team, R: A Language and Environment for Statistical Computing (R Foundation for Statistical Computing, 2015).
- 50.G. Ridgeway, G. Generalized boosted regression models. R package version 2.1.1, http://CRAN.R-project.org/package=gbm [accessed 22 July 2015].
- 51.Fleskes J. P., Perry W. M., Petrik K. L., Spell R., Reid F., Change in area of winter-flooded and dry rice in the northern Central Valley of California determined by satellite imagery. Calif. Fish Game 91, 207–215 (2005). [Google Scholar]
- 52.Arlot S., Celisse A., A survey of cross-validation procedures for model selection. Stat. Surv. 4, 40–79 (2010). [Google Scholar]
- 53.Elith J., Leathwick J. R., Hastie T., A working guide to boosted regression trees. J. Anim. Ecol. 77, 802–813 (2008). [DOI] [PubMed] [Google Scholar]
- 54.R. J. Hijmans, L. Guarino, C. Bussink, P. Mathur, M. Cruz, I. Barrentes, E. Rojas, DIVA-GIS 7.5. A geographic information system for the analysis of species distribution data. (2012); www.diva-gis.org.
- 55.R. J. Hijmans, J. van Etten, raster: Geographic analysis and modeling with raster data. R package version 2.0–12 (R Found. Stat. Comput., Vienna, 2012); http://CRAN.R-project.org/package=raster.
- 56.Stevens D. L. Jr, Olsen A. R., Spatially balanced sampling of natural resources. J. Am. Stat. Assoc. 99, 262–278 (2004). [Google Scholar]