

Abstract
Porcine infections are currently not the state-of-the-art model to study human diseases. Nevertheless, the course of human and porcine toxoplasmosis is much more comparable than that of human and murine toxoplasmosis. For example, severity of infection, transplacental transmission, and interferon-gamma-induced antiparasitic effector mechanisms are similar in pigs and humans. In addition, the severe immunosuppression during acute infection described in mice does not occur in the experimentally infected ones. Thus, we hypothesise that porcine Toxoplasma gondii infection data are more representative for human toxoplasmosis. We therefore suggest that the animal model chosen must be critically evaluated for its assignability to human diseases.
1. Introduction
Toxoplasma gondii (T. gondii) is one of the most prevalent parasites worldwide. This is due to the fact that T. gondii is able to chronically infect all warm-blooded animals including humans. Furthermore, its lifelong persistence in the host increases the chance of transmission. Definitive hosts are members of the Felidae family, which eventually shed environmentally resistant oocysts that are taken up by new intermediate or aberrant hosts (e.g., mice, pigs, or humans) via soil, food, or water [1, 2].
Since pork is the most frequently consumed meat in Europe, the T. gondii infection rate of pigs is of great interest [1, 3]. The prevalence of toxoplasmosis in pigs is commonly determined by the detection of anti-T. gondii antibodies in sera and meat juice. Detection rates of these porcine anti-T. gondii antibodies vary worldwide, ranging from 45% (n = 402) of pigs, analysed in Mexico [4], to 6.2%(n = 1368) positives in a seroprevalence study in Austria [5]. About 25% (n = 1200) positives were measured in Spain [6] and about 19% (n = 2004) in Germany [7].
A “meat juice serology” study, performed in Germany, demonstrated a prevalence of T. gondii of about 10% [8], whereas a prevalence of about 3% (n = 1549) was found in cardiac fluids in a French study [9]. Therefore, setting a benchmarking with the studies conducted so far is exceedingly difficult, since seroprevalence depends on the geographic region. The observed seroprevalence has also been influenced by other variables, such as the detection method, the age of the animals, the species, and the type of specimen. Moreover, prevalence studies performed within individual herds varied from 0 to 100%. Hence, the need for standardized test systems is growing.
One method to standardize serologic studies is the use of recombinant antigens as targets. T. gondii surface antigens (SAG) such as SAG1 [10] or secreted antigens have been successfully employed to analyse murine and porcine probes. Among the latter are the dense granule proteins (GRA proteins) which are secreted into the parasitophorous vacuole (PV) and integrated into the PV membrane (PVM) where they interact with host cell proteins and organelles [11]. For example, a GRA7-based enzyme-linked immunosorbent assay has emerged as a promising tool to study the prevalence of toxoplasmosis in pigs [12]. Furthermore, the important role of GRA antigens in immunity against T. gondii has also been underlined by the efficiency with which a GRA1-GRA7 DNA vaccine elicits cellular and humoral immune responses [13].
To summarize, several publications deal with the porcine humoral immune response after T. gondii infection, but data analysing cellular immune responses are relatively rare. Nevertheless, it is clear that toxoplasmosis initially results in the induction of a type 1 T helper cell (TH1) response in infected swine. Seroconversion was detected 20 days post infection (dpi) in orally infected mini pigs. An enhanced cellular immune response (indicated by increased CD25 expression on lymphocytes) was already detectable on day three postinfection, and significant IFN-γ production was detectable on day six postinfection [14]. Verhelst et al. described an upregulation of interferon response factor 1 (IRF1) and interferon-gamma (IFN-γ) gene expression in infected swine [12]. Dawson et al. were able to ascribe the elevated IFN-γ expression in T. gondii-infected pigs to peripheral blood mononuclear cells (PBMC) [15]. IFN-γ is the most prominent TH1 cytokine and known to be essential for a sufficient defence against T. gondii in different host species and cell types. Nevertheless, the effector mechanisms that are activated by IFN-γ in porcine cells have been unclear up to now.
Here, we show that infected pigs have a strong immune response against recombinant GRA antigens (rGRA), which is concomitant with a strong IFN-γ production by T cells mediating antiparasitic effects. We demonstrate that the growth control of T. gondii tachyzoites is mediated by the IFN-γ-induced expression of the tryptophan-degrading enzyme indoleamine 2,3-dioxygenase (IDO) in porcine choroid plexus epithelial (PCP-R) cells.
2. Materials and Methods
2.1. Infection of Pigs
All animal experiments were performed by Prof. A. M. Tenter (Institute for Parasitology, University of Veterinary Medicine, Hannover). The animal study, permit number DEC 2008.III.30.023, was reviewed and approved by the local animal ethics committee according to the recommendations of the EU directive 86/609/EEC. The number of animals used and their suffering was minimized. In a first study, sera were harvested from 10 orally infected animals (T. gondii; DX) and 7 uninfected controls. In the second part of this study, eight 7- to 9-week-old pigs were infected with T. gondii ME49 oocysts according to the protocol of Bokken et al. [16] and T cell responses were analysed. To simulate a natural infection, pigs were infected orally with 105 oocysts in 2 ml drinking water per animal. Blood samples were taken at different time points (0, 7, 21, 28, 48, 98, and 159 days postinfection (dpi)). Heparinised blood samples were processed within 24 h and peripheral blood mononuclear cells (PBMC) were obtained by Ficoll gradient centrifugation and used immediately.
2.2. Production of Recombinant Antigens
The His-tagged recombinant antigens SAG1 and BAG1 were obtained from the Institute of Medical Microbiology of the University Medical Center Göttingen [17]. The recombinant antigens GRA1, GRA2, GRA7, and GRA9 were produced in the Institute of Medical Microbiology and Hospital Hygiene of the Heinrich Heine University in Düsseldorf as described previously [17]. All recombinant antigens were purified using a nickel-nitrilotriacetic acid (Ni-NTA) matrix under denaturating conditions and tested by SDS-PAGE, Coomassie blue staining, and Western blot analysis using anti-His antibody (QUIAGEN, Hilden, Germany). Protein concentrations were determined using a bicinchoninic acid (BCA) detection assay according to the manufacturer's instruction (Pierce, Rockford, IL).
2.3. Western Blot Analysis
For Western blot analysis, 10% NuPAGE Novex Bis-Tris Mini gels and the appropriate electrophoresis system from Invitrogen (Karlsruhe, Germany) were used. Recombinant proteins (2 or 6 μg) were separated in the SDS-containing gels by electrophoresis for more than one hour at 160 V. Seeblue Plus2 marker was used as molecular mass standard. After the proteins were semidry blotted on nitrocellulose membranes (CarboGlas, Schleicher & Schüll, Dassel, Germany), membranes were blocked in 5% (w/v) skim milk powder in PBS for 1 h at room temperature. Then, the pig sera were used as primary antibody in a 1 : 100 dilution in 5% (w/v) skim milk powder in PBS. After incubation overnight at 4°C, membranes were washed three times in PBS containing 0.2% Tween for 5 minutes. Thereafter, membranes were incubated for 45 minutes at room temperature with goat anti-pig alkaline phosphatase-conjugated IgG (1 : 000 Dianova, Hamburg, Germany), diluted in 5% (w/v) skim milk powder in PBS. After three additional washing steps, bands were detected by the addition of substrate buffer for alkaline phosphatase (100 mM Tris-HCl, pH 9.5; 100 mM NaCl; and 10 mM MgCl2) and staining solution (33 μl BCIP-T and 44 μl NBT in 10 ml substrate buffer). The reaction was stopped by the addition of aqua dest. And membranes were laminated.
2.4. T Cell Proliferation Experiments
Peripheral blood mononuclear cells (PBMC) were prepared from the heparinised blood of infected and noninfected pigs after Ficoll density gradient centrifugation. For proliferation experiments, 1.5 × 105 porcine PBMC per well were incubated in 200 μl Iscove's modified Dulbecco's medium (IMDM; Gibco, Grand Island, USA) containing 5% foetal calf serum (FCS) and 1% PenStrep (Biochrom, Berlin, Germany).
Cells were stimulated with toxoplasma lysate antigen (TLA) (106 lysed RH T. gondii parasites per ml), recombinant antigens (1 μg/ml), concanavalin A (ConA; 1 μg/ml), or left untreated as negative control. After three to five days of incubation (dependent on the result of microscopic examination), 0.2 mCi [3H]-thymidine was added for 24 h and T cell proliferation was determined using liquid scintillation spectrometry (1205 Betaplate, PerkinElmer, Jügesheim, Germany).
2.5. IFN-γ Assay
After a five-day stimulation period with TLA or recombinant antigens the amount of IFN-γ in supernatants of porcine PBMC cultures was determined using the porcine DuoSet ELISA (R&D Systems, Minnesota, USA) according to the manufacturer's instructions.
2.6. Culture and In Vitro Stimulation of PCP-R Cells
Porcine choroid plexus epithelial (PCP-R) cells were obtained from H. Schroten (Department of Pediatrics, Heidelberg University, Germany) [18] and cultured in IMDM containing 10% FCS, 0.05% insulin, and 1% PenStrep at 37°C in a 10% CO2-enriched atmosphere. Cultures were split 1 : 10 every 4 days using 0.05% trypsin-EDTA (Gibco).
1.5 × 105 PCP-R cells/well were stimulated with different amounts of porcine IFN-γ (R&D Systems) for 72 h in 200 μl cell culture medium or left untreated as negative control. Samples for subsequent kynurenine detection were supplemented with 100 μg/ml L-tryptophan (Sigma Aldrich, St. Louis, USA). In addition, some samples were treated with 100 μg/ml 1-L-methyl-tryptophan (1-MT; Sigma Aldrich), an IDO-specific inhibitor.
After three days of incubation, supernatants of stimulated or unstimulated PCP-R cells were harvested and the amount of kynurenine was determined using Ehrlich's reagent [19].
2.7. T. gondii Infection of PCP-R Cells
After 72 h of stimulation with or without IFN-γ in the presence or absence of 1-MT or additional L-tryptophan, PCP-R cells were infected with 105 T. gondii ME49 tachyzoites per well. The infected cells were incubated at 37°C in a 10% CO2-enriched atmosphere. After 24 h, 0.012 MBq [3H]-uracil was added to the cells, and after lysis of the cells, the cell culture plates were frozen at −20°C. T. gondii proliferation was determined using liquid scintillation spectrometry (1205 Betaplate, PerkinElmer, Jügesheim, Germany).
2.8. Statistical Analysis
All experiments were performed in triplicate, and data are given as mean ± standard error of a minimum of three independent experiments. For statistical analysis, the two-tailed unpaired t-test was used and significant differences are indicated with asterisks (∗p ≤ 0.05, ∗∗p ≤ 0.01, and ∗∗∗p ≤ 0.0001). Analysis was performed using GraphPad Prism software (GraphPad Software Inc., San Diego, CA).
3. Results
The oral uptake of T. gondii oocysts frequently results in an asymptomatic infection in pigs, which induces a robust production of parasite-specific antibodies. Therefore, seroprevalence studies are usually performed to detect T. gondii infection in pigs. In a first study, we therefore analysed the antibody production against recombinant T. gondii proteins in pigs infected orally with T. gondii (103 or 105 oocysts per animal). We performed Western blot analysis using recombinant T. gondii proteins GRA1, GRA2, GRA7, and GRA9. We found antibodies against GRA2, GRA7, and GRA9 98 dpi in all sera, irrespective of infection dose. However, antibodies against GRA1 were detected only infrequently (Table 1). No antibodies against T. gondii proteins could be detected in sera from uninfected animals (n = 7). This serologic data indicate that oral infection with 103 as well as 105 oocysts is effective.
Table 1.
Immunoreactivity of sera from T. gondii-infected pigs to different recombinant GRA antigens (GRA) based on Western blot analyses.
| Frequency of positive detection (%) | ||||
|---|---|---|---|---|
| GRA1 | GRA2 | GRA7 | GRA9 | |
| Sera of uninfected pigs (n = 7) | 0 | 0 | 0 | 0 |
| Sera of infected pigs 98 dpi with 103 oocysts (n = 7) |
20 | 100 | 100 | 80 |
| Sera of infected pigs 98 dpi with 105 oocysts (n = 7) |
0 | 100 | 100 | 100 |
In the second part of our study, we focused on the cellular immune response during porcine toxoplasmosis. In the early phase of toxoplasmosis, a strong immunosuppression is described in mice [20, 21]. Hence, the ability of the parasite to interfere with the porcine immune system was analysed. To this end, the responsiveness of porcine T cells was determined prior to infection or at early time points postinfection (7 dpi). In these proliferation studies, peripheral blood mononuclear cells (PBMC) from infected and noninfected animals were stimulated with the T cell mitogen concanavalin A (ConA) or with TLA. After five days of incubation, T cell proliferation was determined by 3H-thymidine incorporation. As shown in Figure 1(a) T cells from noninfected and infected pigs respond comparably to the polyclonal stimulus ConA. Interestingly, only cells harvested from infected animals (7 dpi) show a significant proliferation after TLA stimulation. Comparable data were also obtained with cells harvested at 28 dpi shown in Figure 1(b). PBMCs, sampled 28 dpi and later (42, 56, 84, and 159 dpi) demonstrated a strong antigen-specific T cell proliferation already after 3 days of stimulation.
Figure 1.
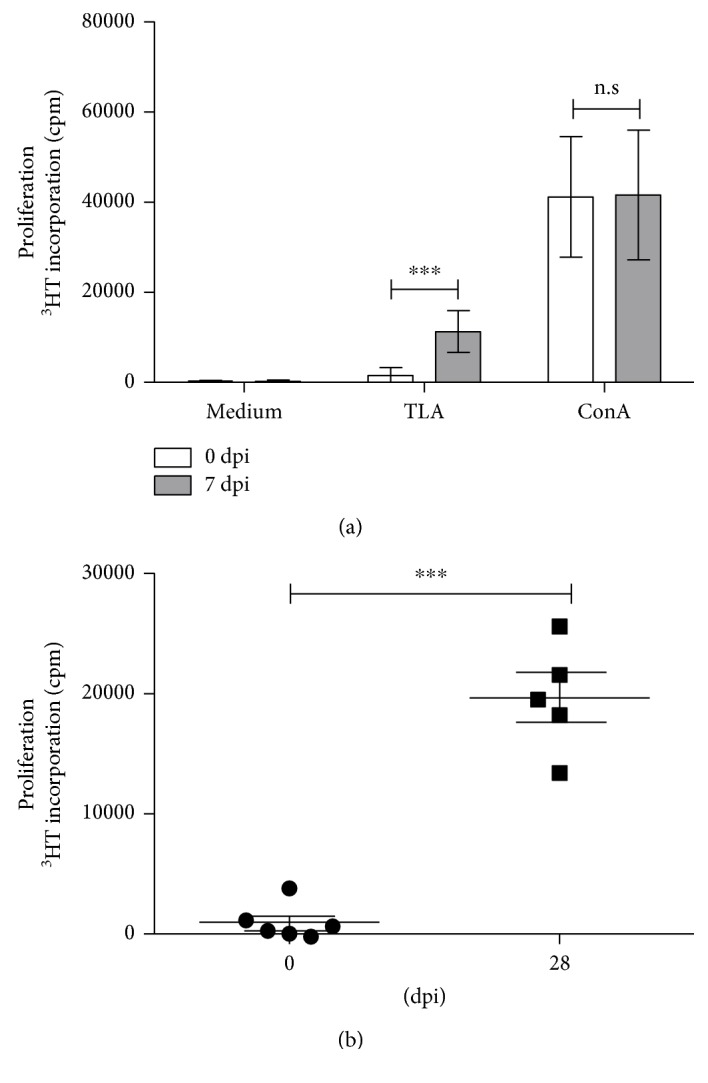
Toxoplasma antigen-specific T cell proliferation in cells from T. gondii-infected animals. (a) 1.5 × 105 PBMC/well from 6 pigs before infection and 7 dpi were stimulated in triplicate with TLA or ConA for 4 days. Thereafter, 3H-thymidine was added for 18–24 h. Data are given as mean cpm ± SD from 6 individuals. (b) TLA-specific cell proliferation from uninfected and infected (28 dpi) animals. Each dot represents the result of one individual. Significant differences are marked with asterisks. Data are given as mean cpm ± SD from 5 individuals; (n.s. = not significant; ∗∗∗p ≤ 0.0001).
In subsequent studies, the T cell response against several recombinant T. gondii antigens was analysed. In these experiments PBMC from infected pigs were stimulated with four different recombinant antigens (GRA1, GRA2, GRA7, and GRA9) at a concentration of 1 μg/ml. In addition, recombinant SAG1 and BAG1 antigens were used since these are typical antigens characterising the tachyzoite and bradyzoite stage of T. gondii, respectively. Both antigens have previously been described as target antigens for murine and human T cells [21, 22]. After five days of stimulation with recombinant T. gondii proteins, T cell growth was monitored by 3H-thymidine incorporation. The data presented in Figure 2 were obtained with PBMC of 8 animals harvested 28 dpi and show that all recombinant T. gondii antigens tested can elicit a T cell response in infected animals, while no proliferative response was observed when PBMC from noninfected animals were stimulated with the same antigens. In this experiment, GRA1 was found to activate cells from five animals, GRA7, GRA9, and SAG1 were recognised by T cells from four animals, and T cells from two animals recognised only GRA2 or BAG1. However, every infected animal had T cells recognising at least one of the tested recombinant antigens. In further experiments, PBMC harvested 7, 28, and 48 dpi and were tested and comparable data were obtained.
Figure 2.

T cell proliferation after stimulation with recombinant antigens from T. gondii. 1.5 × 105 PBMC/well from 8 pigs before infection (0 dpi) and day 28 post infection (28 dpi) were stimulated in triplicate with recombinant GRA1, GRA2, GRA7, GRA9, BAG1, or SAG1 (1 μg/ml) and T cell proliferation was determined via 3H-thymidine incorporation. Each dot represents one individual; black bars indicate mean counts per minute from all individuals tested; significant differences are marked with asterisks (∗p ≤ 0.05, ∗∗p ≤ 0.01, and ∗∗∗p ≤ 0.0001).
Figure 3 depicts the individual in vitro reactivity of T cells from three different pigs after oral infection with oocysts (7 dpi). For example, pig number 1 developed a T cell response to all tested antigens with the strongest response directed against GRA1 and SAG1. Pig number 2 developed a comparable T cell response against all six antigens tested, while cells from pig number 3 showed a weak response to GRA1, GRA2, and SAG1 and no response was observed to GRA7, GRA9, and BAG1.
Figure 3.
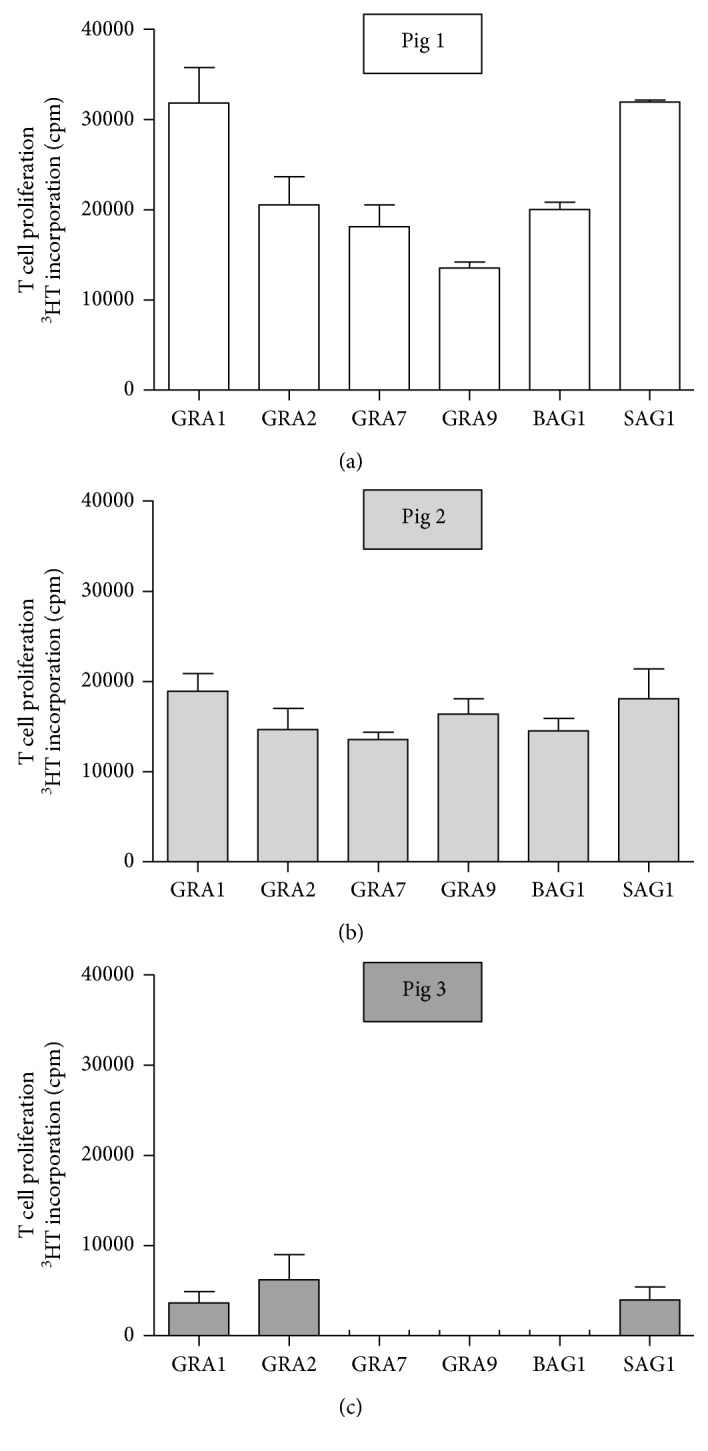
Differential response of PBMC harvested from different animals (pigs 1–3) to recombinant T. gondii antigens. 1.5 × 105 PBMC harvested 7 dpi were stimulated with recombinant T. gondii antigens (1 μg/ml). T cell proliferation was determined on day 5 after in vitro stimulation. Data are given as mean counts per minute (cpm).
In immunocompetent individuals, infection with T. gondii results in a robust stimulation of a TH1 response. Since IFN-γ is the most prominent TH1 cytokine, the IFN-γ production induced by stimulation with TLA (Figure 4(a)) or recombinant antigens (Figure 4(b)) was determined in PBMC from infected animals. IFN-γ is produced in high amounts by T cells harvested from blood after 28 dpi. A comparable IFN-γ production was also found in T cells harvested 159 dpi. Furthermore, T cells stimulated with recombinant antigens (e.g., GRA1) also produce IFN-γ, but the response is less intense as shown in Figure 4(b). Comparable data were also obtained with other recombinant antigens tested (data not shown).
Figure 4.

IFN-γ production by T. gondii-specific T cells. 1.5 × 105 peripheral blood mononuclear cells from infected animals were stimulated with TLA (a) or GRA1 (b) for five days. Thereafter, supernatants were harvested and the IFN-γ concentration was determined by ELISA. Data are given as mean ± SD from two experiments, performed in duplicate (a) or as mean ± SD from four experiments, performed in duplicate (b).
IFN-γ induces a plethora of antimicrobial effector mechanisms. In previous studies, we discovered that native porcine PCP-R cells are able to restrict the growth of Streptococcus suis after IFN-γ stimulation. Furthermore, we showed that IFN-γ induced the expression of indoleamine 2,3-dioxygenase (IDO) and mediated antibacterial effects by tryptophan depletion [23].
Therefore, we tested whether IFN-γ-activated PCP-R cells [18] can restrict the growth of T. gondii. Data of a representative experiment are shown in Figure 5.
Figure 5.

IFN-γ induces IDO activity and antiparasitic effects in porcine PCP-R cells. 3 × 105 PCP-R cells were stimulated with IFN-γ (100 ng/ml). In some groups, the IDO inhibitor 1-methyltryptophan (1-MT) was supplemented (100 μg/ml). Kynurenine concentration was determined in the supernatants using Ehrlich's reagent after three days of stimulation (a). Identical cell cultures were stimulated and afterwards infected with T. gondii (3 × 104 ME49 tachyzoites per well). In the control group, L-tryptophan (L-Trp) (100 μg/ml) was added at the time of infection. After three days, toxoplasma growth was monitored using 3H-uracil incorporation (b). Data are given as mean ± SD of triplicate cultures; significant differences were marked with asterisks (∗p ≤ 0.05, ∗∗p ≤ 0.01, and ∗∗∗p ≤ 0.0001).
As expected, IDO is induced by IFN-γ in PCP-R cells. Interestingly, we found that these activated PCP-R cells can restrict the growth of T. gondii. Furthermore, this antiparasitic effect could be blocked either by tryptophan supplementation or by 1-MT (an IDO-specific inhibitor), indicating that IDO is involved in the defence of PCP-R cells against T. gondii.
The last experimental setting was designed to show whether the amount of IFN-γ produced by T. gondii-stimulated T cells is sufficient to reduce T. gondii proliferation in PCP-R cells. Therefore, T cell supernatants harvested from TLA-stimulated porcine T cells were added to PCP-R cells, and kynurenine production (indicating IDO activity) was determined after three days of culture.
PCP-R cells stimulated with T cell supernatants harvested from culture supernatants from TLA-stimulated T cells (from T. gondii-infected animals) express IDO activity while supernatants harvested from cells from uninfected animals did not (Figure 6(a)). And, even more interestingly, supernatants harvested from TLA-stimulated PBMC from infected animals can induce a parasitostatic state in PCP-R cells which could be reversed by 1-MT or tryptophan supplementation (Figure 6(b)), indicating that the parasitostatic effect is based on IDO-mediated tryptophan degradation.
Figure 6.

Supernatants of TLA-stimulated PBMC obtained from T. gondii-infected animals induce IDO activity and antiparasitic effects in porcine PCP-R cells. 3 × 105 PCP-R cells were stimulated with serially diluted supernatants from TLA-activated porcine PBMC. (a) After three days of PBMC culture, supernatants were harvested and the kynurenine concentration was determined using Ehrlich's reagent. (b) Identical cell cultures were infected with T. gondii (3 × 104 ME49 tachyzoites per well) after stimulation with T cell supernatants. In some cultures, L-tryptophan (100 μg/ml) or 1 MT (100 μg/ml) was added. After three days, toxoplasma growth was monitored using 3H-uracil incorporation. Data are given as mean ± SD of triplicate cultures; significant differences were marked with asterisks (n.s. = not significant; ∗p ≤ 0.05, ∗∗p ≤ 0.01, and ∗∗∗p ≤ 0.0001).
4. Discussion
The use of porcine cells to study human diseases is not very common. However, in the case of toxoplasmosis, the porcine model is superior to the murine model which is usually employed. For example, infection of pregnant pigs with toxoplasma results in abortion or in the birth of infected symptomatic and asymptomatic piglets [24, 25]. This resembles the human situation where an intrauterine infection might also lead either to abortion or to the birth of an infected child [1, 26]. The risk of mother to child transmission during primary toxoplasmosis depends on the gestation time. The transmission risk in the first trimenon is lower than in the third trimenon, and not all primary infections result in vertical transmission of the parasite [26], suggesting that a mechanism exists which protects the unborn child.
In contrast, abortion during toxoplasmosis in mice is mainly due to the abortogenic effect of the TH1 cytokine IFN-γ and infected pups are usually not found in immunocompetent mice [27, 28]. In addition, in humans and pigs, postnatally acquired toxoplasmosis in immunocompetent individuals usually only causes a mild clinical symptom (e.g., fever) or remains asymptomatic[29], while toxoplasmosis in inbred mice is, dependent on toxoplasma strain virulence and infection dose, a life-threatening disease. In contrast, T. gondii strains which are apathogenic in the murine system might cause clinical disease in humans and in pigs [30].
Usually, T. gondii infection in pigs results in the induction of a strong humoral and cellular immune response. Both are directed against soluble T. gondii proteins as well as surface antigens. The so-called “excreted secreted antigens” (ESA) from T. gondii have been found to be immunodominant antigens and consist of a mixture of GRA proteins from T. gondii including GRA1, GRA2, GRA7, and GRA9 [11].
When pigs are immunized with ESA preparations, they develop a strong cellular and humoral immune response resulting in resistance to infections with virulent type I strains. The porcine immune response has also been shown to be mainly directed against ESA proteins with molecular masses between 34 and 116 kDa. Furthermore, the immunization also results in reduced cyst formation in the muscle which might be beneficial for the consumer due to a reduced risk of infection [31].
We were therefore interested in analysing the humoral immune responses in sera of experimentally infected pigs directed against defined GRA proteins from T. gondii. Our Western blot analysis revealed that all orally infected animals developed a strong humoral immune response against GRA2, GRA7, and GRA9, while no antibodies directed against GRA1 were found. This is in accordance with published data from Verhelst et al. [12] who analysed the antibody production in pigs after oral infection with T. gondii tissue cysts. Using an ELISA technique, they found strong immunoreactivity to GRA7 and also no antibodies directed against GRA1. In addition, Jongert et al. immunized pigs with a GRA1-GRA7 cocktail DNA vaccine [13]. After immunization with this DNA vaccine, they found a strong humoral immune response directed against GRA7 and GRA1, whereas no detectable immune response was elicited in animals vaccinated with tachyzoites of the RH strain. However, RH-vaccinated animals developed high amounts of anti-GRA7 antibodies, but not anti-GRA1 antibodies after reinfection with T. gondii tissue cysts. Altogether, we confirmed previous findings concerning the immunogenicity of GRA1 and GRA7 and additionally demonstrated a high immunogenicity of GRA2 and GRA9.
Antibodies in combination with complement can only kill extracellular parasites; therefore, a cellular immune response is necessary to control intracellular T. gondii. However, the strong antigen-specific T cell response of porcine cells even in the acute phase of toxoplasmosis (7 dpi) stands in striking contrast to data published with murine cells. Murine spleen cells harvested within the first weeks after T. gondii infection are unable to respond to stimulation with ConA or TLA. Detailed analysis showed T cell suppression in spleen cells from T. gondii-infected mice. This suppression is mainly due to an IFN-γ-dependent induction of the inducible nitric oxide synthase (iNOS) enzyme activity [20, 32].
In contrast, peripheral blood lymphocytes from patients with acute, symptomatic toxoplasmosis can mount a proliferative response after stimulation with soluble toxoplasma antigen [33]. We also found strong toxoplasma antigen-specific proliferation after stimulating PBLs from three different donors with acute symptomatic toxoplasmosis (own unpublished observations). Therefore, T cell reactivity during acute toxoplasmosis is comparable between humans and pigs but not mice.
Activated T cells usually do not interact directly with T. gondii, but T cell cytokines can induce antiparasitic mechanisms in different cell types. Several publications confirm that pigs, like many other species including humans, mount a TH1-type immune response during toxoplasmosis. Ex vivo analysis showed enhanced proliferation and expression of activation markers on T cells and increased IFN-γ production by T cells from infected animals [15, 31]. Furthermore, increased amounts of IFN-γ in sera of infected pigs were described as well as increased transcription of IFN-γ and IRF-1 [12, 14].
Our results support these findings since we found a strong proliferative T cell response after stimulation of PBMC with TLA in all animals orally infected with T. gondii. In addition, these T cells produced high amounts of IFN-γ. However, only little information is available concerning T cell stimulation with defined T. gondii antigens. For example, Jongert et al. saw that the GRA1-GRA7 DNA vaccine-immunized pigs developed GRA1- and GRA7-specific T cells and that these T cells produced the TH1 cytokine IFN-γ [13]. Here, we prove that all tested recombinant GRA and surface antigens were recognized by T cells of infected animals. However, GRA1, which causes no or only a weak humoral immune response, initiates strong T cell activation. Furthermore, we found that GRA-specific T cells can produce IFN-γ. However, the amount of IFN-γ produced in comparison to that of TLA-stimulated cells was low. This might be due to polyclonal T cell stimulation via the complex toxoplasma lysate antigen in contrast to the mono- or oligoclonal stimulation via defined recombinant antigens.
IFN-γ is well known as potent inducer of antimicrobial effector mechanisms; among them, the inducible nitrite oxide synthase (iNOS) [34], interferon-induced GTPases [35], and tryptophan-degrading enzyme IDO [36] are most important in the defense against T. gondii in several species. For example, iNOS was frequently reported to inhibit the growth of parasites in vitro and in vivo especially in mice. However, despite the observation that nitric oxide is involved in the pathophysiological processes during septic shock and LPS response in pigs [37, 38] and that iNOS is upregulated in the central nervous system of swine following infection with pseudorabies [39], iNOS is not an essential part of the innate immune response in pigs [40].
Interferon-induced GTPases were described to be involved in antimicrobial defence. Among them, Mx proteins, mainly dependent on type 1 interferon induction, mediate antiviral effects in human, murine, and porcine cells [41, 42].
Additionally, p47 GTPases mainly stimulated by IFN-γ were described to mediate antimicrobial effects in murine [42] but not in human cells [43], while data on porcine cells are lacking. The third family of IFN-induced GTPases is the guanylate-binding proteins (GBPs). Different mGBPs can mediate antitoxoplasma effects in murine cells [44, 45]. More recently, human GBP-1 was described to mediate antitoxoplasma effects in human mesenchymal stem cells [46]. No data concerning GBP-mediated antiparasitic effects have been published with porcine cells up to now; however, very recently, GBP1 was found to be a mediator of antiviral effects in a porcine kidney cell line [47].
The IFN-γ induced induction of IDO with subsequent degradation of tryptophan and inhibition of toxoplasma growth was first recognized by Pfefferkorn in 1984 [36]. Meanwhile, many human cells were found to use this mechanism to inhibit the growth of T. gondii tachyzoites and other tryptophan-auxotroph microorganisms [48]. Overall, only few papers have been published showing IDO-mediated effects in porcine cells. For example, porcine IDO was cloned in 2012 and was found to be more similar to human IDO than murine IDO [49]. On a functional level, porcine IDO was linked to protective effects in transplant arteriosclerosis and in xenoreactions [50, 51]. Furthermore, the role of IDO in the porcine lung in inflammation models has been well studied and IDO-mediated effects against Streptococcus suis have been described [23, 52].
In this manuscript, we show for the first time that the induction of IDO is responsible for the inhibition of T. gondii growth in porcine cells. We found that PCP-R cells can restrict the growth of T. gondii after stimulation with IFN-γ or after coincubation with T cell supernatants harvested from TLA-stimulated cells originating from infected animals. This antiparasitic effect could be blocked, at least in part, by addition of the IDO-specific inhibitor 1-MT or by supplementation of large amounts of tryptophan. IDO- and iNOS-mediated antiparasitic effects were found to be species-specific. Furthermore, we found that in porcine and human cells, IDO induction is the most important antiparasitic effector mechanism directed against T. gondii. Thus, antimicrobial effects in human and porcine cells were similar, but different from those in murine cells [53, 54].
IDO and iNOS mediate antimicrobial and immunoregulatory effects and human as well as porcine mesenchymal stem cells can inhibit alloantigen-driven T cell responses [55, 56]. Interestingly, also in respect to immunoregulatory effects, a species-specific difference is described. For example, in human and porcine mesenchymal stem cells, IDO was found to mediate immunosuppressive effects, while iNOS was involved in immunoregulation mediated by murine MSC [57]. Once again, porcine cells act more like human cells than murine cells do.
Acknowledgments
This work was supported by grants from the Federal Ministry of Education and Research (BMBF) within the TOXONET consortium (Grants 01 KI 0764 and 01 KI 1002E to Walter Däubener and 01 KI 0766 and 01 KI 1002B to Uwe Groß).
Conflicts of Interest
The authors declare that there is no conflict of interest regarding the publication of this paper.
References
- 1.Schlüter D., Däubener W., Schares G., Groß U., Pleyer U., Lüder C. Animals are key to human toxoplasmosis. International Journal of Medical Microbiology. 2014;304(7):917–929. doi: 10.1016/j.ijmm.2014.09.002. [DOI] [PubMed] [Google Scholar]
- 2.Tenter A. M., Heckeroth A. R., Weiss L. M. Toxoplasma gondii: from animals to humans. International Journal of Parasitology. 2000;30(12-13):1217–1258. doi: 10.1016/s0020-7519(00)00124-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Tenter A. M. Toxoplasma gondii in animals used for human consumption. Memórias do Instituto Oswaldo Cruz. 2009;104(2):364–369. doi: 10.1590/s0074-02762009000200033. [DOI] [PubMed] [Google Scholar]
- 4.Alvarado-Esquivel C., Romero-Salas D., García-Vázquez Z., et al. Seroprevalence and correlates of Toxoplasma gondii infection in domestic pigs in Veracruz State, Mexico. Tropical Animal Health and Production. 2014;46(4):705–709. doi: 10.1007/s11250-014-0551-3. [DOI] [PubMed] [Google Scholar]
- 5.Steinparzer R., Reisp K., Grünberger B., Köfer J., Schmoll F., Sattler T. Comparison of different commercial serological tests for the detection of Toxoplasma gondii antibodies in serum of naturally exposed pigs. Zoonoses and Public Health. 2015;62(2):119–124. doi: 10.1111/zph.12122. [DOI] [PubMed] [Google Scholar]
- 6.Herrero L., Gracia M. J., Pérez-Arquillué C., et al. Toxoplasma gondii: pig seroprevalence, associated risk factors and viability in fresh pork meat. Veterinary Parasitology. 2016;224(5):52–59. doi: 10.1016/j.vetpar.2016.05.010. [DOI] [PubMed] [Google Scholar]
- 7.Damriyasa I. M., Bauer C., Edelhofer R., et al. Cross-sectional survey in pig breeding farms in Hessen, Germany: seroprevalence and risk factors of infections with Toxoplasma gondii, Sarcocystis spp. and Neospora caninum in sows. Veterinary Parasitology. 2004;126(3):271–286. doi: 10.1016/j.vetpar.2004.07.016. [DOI] [PubMed] [Google Scholar]
- 8.Meemken D., Tangemann A. H., Meermeier D., et al. Establishment of serological herd profiles for zoonoses and production diseases in pigs by “meat juice multi-serology”. Preventive Veterinary Medicine. 2014;113(4):589–598. doi: 10.1016/j.prevetmed.2013.12.006. [DOI] [PubMed] [Google Scholar]
- 9.Djokic V., Blaga R., Aubert D., et al. Toxoplasma gondii infection in pork produced in France. Parasitology. 2016;143(5):557–567. doi: 10.1017/S0031182015001870. [DOI] [PubMed] [Google Scholar]
- 10.Fang R., Feng H., Nie H., et al. Construction and immunogenicity of pseudotype baculovirus expressing Toxoplasma gondii SAG1 protein in BALB/c mice model. Vaccine. 2009;28(7):1803–1807. doi: 10.1016/j.vaccine.2009.12.005. [DOI] [PubMed] [Google Scholar]
- 11.Mercier C., Adjogble K. D., Däubener W., Delauw M. F. Dense granules: are they key organelles to help understand the parasitophorous vacuole of all apicomplexa parasites? International Journal of Parasitology. 2005;35(8):829–849. doi: 10.1016/j.ijpara.2005.03.011. [DOI] [PubMed] [Google Scholar]
- 12.Verhelst D., De Craeye S., Dorny P., et al. IFN-γ expression and infectivity of toxoplasma infected tissues are associated with an antibody response against GRA7 in experimentally infected pigs. Veterinary Parasitology. 2011;179(1–3):14–21. doi: 10.1016/j.vetpar.2011.02.015. [DOI] [PubMed] [Google Scholar]
- 13.Jongert E., Melkebeek V., De Craeye S., Dewit J., Verhelst D., Cox E. An enhanced GRA1-GRA7 cocktail DNA vaccine primes anti-toxoplasma immune responses in pigs. Vaccine. 2008;26(8):1025–1031. doi: 10.1016/j.vaccine.2007.11.058. [DOI] [PubMed] [Google Scholar]
- 14.Solano Aguilar G. I., Beshah E., Vengroski K. G., et al. Cytokine and lymphocyte profiles in miniature swine after oral infection with Toxoplasma gondii oocysts. International Journal of Parasitology. 2001;31(2):187–195. doi: 10.1016/S0020-7519(00)00159-4. [DOI] [PubMed] [Google Scholar]
- 15.Dawson H. D., Royaee A. R., Nishi S., et al. Identification of key immune mediators regulating T helper 1 responses in swine. Veterinary Immunology and Immunopathology. 2004;100(1-2):105–111. doi: 10.1016/j.vetimm.2004.03.006. [DOI] [PubMed] [Google Scholar]
- 16.Bokken G. C. A. M., van Eerden E., Opsteegh M., et al. Specific serum antibody responses following a Toxoplasma gondii and Trichinella spiralis co-infection in swine. Veterinary Parasitology. 2012;184(2–4):126–132. doi: 10.1016/j.vetpar.2011.08.028. [DOI] [PubMed] [Google Scholar]
- 17.Hotop A., Buschtöns S., Bangouras B., et al. Humoral immune responses in chickens and turkeys after infection with Toxoplasma gondii by using recombinant antigens. Parasitology Research. 2014;113(4):1473–1480. doi: 10.1007/s00436-014-3788-x. [DOI] [PubMed] [Google Scholar]
- 18.Schroten M., Hanisch F. G., Quednau N., et al. A novel porcine in vitro model of the blood-cerebrospinal fluid barrier with strong barrier function. PLoS One. 2012;7(6, article e39835) doi: 10.1371/journal.pone.0039835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Däubener W., Wanagat N., Pilz K., Seghrouchni S., Fischer H. G., Hadding U. A new, simple, bioassay for human IFN-gamma. Journal of Immunological Methods. 1994;168(1):39–47. doi: 10.1016/0022-1759(94)90207-0. [DOI] [PubMed] [Google Scholar]
- 20.Candolfi E., Hunter C. A., Remington J. S. Mitogen- and antigen-specific proliferation of T cells in murine toxoplasmosis is inhibited by reactive nitrogen intermediates. Infection and Immunity. 1994;62(5):1995–2001. doi: 10.1128/iai.62.5.1995-2001.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Laguía-Becher M., Martín V., Kraemer M., et al. Effect of codon optimization and subcellular targeting on Toxoplasma gondii antigen SAG1 expression in tobacco leaves to use in subcutaneous and oral immunization in mice. BMC Biotechnology. 2010;10(52) doi: 10.1186/1472-6750-10-52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Di Cristina M., Del Porto P., Buffolano W., et al. The Toxoplasma gondii bradyzoite antigens BAG1 and MAG1 induce early humoral and cell-mediated immune responses upon human infection. Microbes and Infection. 2004;6(2):164–171. doi: 10.1016/j.micinf.2003.11.009. [DOI] [PubMed] [Google Scholar]
- 23.Adam R. A., Tenenbaum T., Valentin-Weigand P., et al. Porcine choroid plexus epithelial cells induce Streptococcus suis bacteriostasis in vitro. Infection and Immunity. 2004;72(5):3084–3087. doi: 10.1128/IAI.72.5.3084-3087.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Jungersen G., Bille-Hansen V., Jensen L., Lind P. Transplacental transmission of Toxoplasma gondii in minipigs infected with strains of different virulence. Journal of Parasitology. 2001;87(1):108–113. doi: 10.1645/0022-3395(2001)087[0108:TTOTGI]2.0.CO;2. [DOI] [PubMed] [Google Scholar]
- 25.Dubey J. P., Urban J. F. Diagnosis of transplacentally induced toxoplasmosis in pigs. American Journal of Veterinary Research. 1990;51(8):1295–1299. [PubMed] [Google Scholar]
- 26.Wallon M., Peyron F., Cornu C., et al. Congenital toxoplasma infection: monthly prenatal screening decreases transmission rate and improves clinical outcome at age 3 years. Clinical Infectious Diseases. 2013;56(9):1223–1231. doi: 10.1093/cid/cit032. [DOI] [PubMed] [Google Scholar]
- 27.Shiono Y., Mun H. S., He N., et al. Maternal-fetal transmission of Toxoplasma gondii in interferon-gamma deficient pregnant mice. Parasitology International. 2007;56(2):141–148. doi: 10.1016/j.parint.2007.01.008. [DOI] [PubMed] [Google Scholar]
- 28.Senegas A., Villard O., Neuville A., et al. Toxoplasma gondii-induced foetal resorption in mice involves interferon-gamma-induced apoptosis and spiral artery dilation at the maternofoetal interface. International Journal of Parasitology. 2009;39(4):481–487. doi: 10.1016/j.ijpara.2008.08.009. [DOI] [PubMed] [Google Scholar]
- 29.Dubey J. P. A review of toxoplasmosis in pigs. Veterinary Parasitology. 1986;19(3-4):181–223. doi: 10.1016/0304-4017(86)90070-1. [DOI] [PubMed] [Google Scholar]
- 30.Jungersen G., Jensen L., Riber U., et al. Pathogenicity of selected Toxoplasma gondii isolates in young pigs. International Journal of Parasitology. 1999;29(8):1307–1319. doi: 10.1016/S0020-7519(99)00078-8. [DOI] [PubMed] [Google Scholar]
- 31.Wang Y., Zhang D., Wang G., Yin H., Wang M. Immunization with excreted-secreted antigens reduces tissue cyst formation in pigs. Parasitology Research. 2013;112(11):3835–3842. doi: 10.1007/s00436-013-3571-4. [DOI] [PubMed] [Google Scholar]
- 32.Candolfi E., Hunter C. A., Remington J. S. Roles of gamma interferon and other cytokines in suppression of the spleen cell proliferative response to concanavalin A and toxoplasma antigen during acute toxoplasmosis. Infection and Immunity. 1995;63(3):751–756. doi: 10.1128/iai.63.3.751-756.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Sklenar I., Jones T. C., Alkan S., Erb P. Association of symptomatic human infection with Toxoplasma gondii with imbalance of monocytes and antigen-specific T cell subsets. The Journal of Infectious Diseases. 1986;153(2):315–324. doi: 10.1093/infdis/153.2.315. [DOI] [PubMed] [Google Scholar]
- 34.Scharton-Kersten T. M., Yap G., Magram J., Sher A. Inducible nitric oxide is essential for host control of persistent but not acute infection with the intracellular pathogen Toxoplasma gondii. Journal of Experimental Medicine. 1997;185(7):1261–1273. doi: 10.1084/jem.185.7.1261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Taylor G. A., Collazo C. M., Yap G. S., et al. Pathogen-specific loss of host resistance in mice lacking the IFN-gamma-inducible gene IGTP. Proceedings of the National Academy of Sciences of the United States of America. 2000;97(2):751–755. doi: 10.1073/pnas.97.2.751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Pfefferkorn E. R. Interferon gamma blocks the growth of Toxoplasma gondii in human fibroblasts by inducing the host cells to degrade tryptophan. Proceedings of the National Academy of Sciences of the United States of America. 1984;81(3):908–912. doi: 10.1073/pnas.81.3.908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Doursout M. F., Oguchi T., Fischer U. M., et al. Distribution of NOS isoforms in a porcine endotoxin shock model. Shock. 2008;29(6):692–670. doi: 10.1097/shk.0b013e3181598b77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Lange M., Enkhbaatar P., Nakano Y., Traber D. L. Role of nitric oxide in shock: the large animal perspective. Frontiers in Bioscience (Landmark Edition) 2009;14:1979–1989. doi: 10.2741/3357. [DOI] [PubMed] [Google Scholar]
- 39.Marcaccini A., López-Peña M., Bermúdez R., et al. Pseudorabies virus induces a rapid up-regulation of nitric oxide synthases in the nervous system of swine. Veterinary Microbiology. 2007;125(3-4):232–243. doi: 10.1016/j.vetmic.2007.05.027. [DOI] [PubMed] [Google Scholar]
- 40.Pampusch M. S., Bennaars A. M., Harsch S., Murtaugh M. P. Inducible nitric oxide synthase expression in porcine immune cells. Veterinary Immunology and Immunopathology. 1998;61(1-2):279–289. doi: 10.1016/S0165-2427(97)00139-6. [DOI] [PubMed] [Google Scholar]
- 41.He D. N., Zhang X. M., Liu K., et al. In vitro inhibition of the replication of classical swine fever virus by porcine Mx1 protein. Antiviral Research. 2014;104:128–135. doi: 10.1016/j.antiviral.2014.01.020. [DOI] [PubMed] [Google Scholar]
- 42.Taylor G. A., Feng C. G., Sher A. Control of IFN-gamma-mediated host resistance to intracellular pathogens by immunity-related GTPases (p47 GTPases) Microbes and Infection. 2007;9(14-15):1644–1651. doi: 10.1016/j.micinf.2007.09.004. [DOI] [PubMed] [Google Scholar]
- 43.Hunn J. P., Feng C. G., Sher A., Howard J. C. The immunity-related GTPases in mammals: a fast-evolving cell-autonomous resistance system against intracellular pathogens. Mammalian Genome. 2011;22(1-2):43–54. doi: 10.1007/s00335-010-9293-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Degrandi D., Konermann C., Beuter-Gunia C., et al. Extensive characterization of IFN-induced GTPases mGBP1 to mGBP10 involved in host defense. Journal of Immunology. 2007;179(11):7729–7740. doi: 10.4049/jimmunol.179.11.7729. [DOI] [PubMed] [Google Scholar]
- 45.Degrandi D., Kravets E., Konermann C., et al. Murine guanylate binding protein 2 (mGBP2) controls Toxoplasma gondii replication. Proceedings of the National Academy of Sciences of the United States of America. 2013;110(1):294–299. doi: 10.1073/pnas.1205635110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Qin A., Lai D. H., Liu Q., et al. Guanylate-binding protein 1 (GBP1) contributes to the immunity of human mesenchymal stromal cells against Toxoplasma gondii. Proceedings of the National Academy of Sciences of the United States of America. 2017;114(4):1365–1370. doi: 10.1073/pnas.1619665114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Li L. F., Yu J., Li Y., et al. Guanylate-binding protein 1, an interferon-induced GTPase, exerts an antiviral activity against classical swine fever virus depending on its GTPase activity. Journal of Virology. 2016;90(8):4412–4426. doi: 10.1128/JVI.02718-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.MacKenzie C. R., Heseler K., Müller A., Däubener W. Role of indoleamine 2,3-dioxygenase in antimicrobial defence and immuno-regulation: tryptophan depletion versus production of toxic kynurenines. Current Drug Metabolism. 2007;8(3):237–244. doi: 10.2174/138920007780362518. [DOI] [PubMed] [Google Scholar]
- 49.Chen C., Wie M., Wang L., Xiang Y., Fu X., Zhu M. Molecular cloning and characterization of porcine indoleamine 2, 3-dioxygenase and its expression in various tissues. Journal of Huazhong University of Science and Technology [Medical Sciences] 2012;32(4):473–479. doi: 10.1007/s11596-012-0082-y. [DOI] [PubMed] [Google Scholar]
- 50.Wee J. L., Christiansen D., Li Y. Q., Boyle W., Sandrin M. S. Suppression of cytotoxic and proliferative xenogeneic T-cell responses by transgenic expression of indoleamine 2,3-dioxygenase. Immunology and Cell Biology. 2008;86(5):460–465. doi: 10.1038/icb.2008.8. [DOI] [PubMed] [Google Scholar]
- 51.Mulley W. R., Li L.- Q., Wee J. L., et al. Local expression of IDO, either alone or in combination with CD40Ig, IL10 or CTLA4Ig, inhibits indirect xenorejection responses. Xenotransplantation. 2008;15(3):174–183. doi: 10.1111/j.1399-3089.2008.00472.x. [DOI] [PubMed] [Google Scholar]
- 52.Melchior D., Mézière N., Sève B., Le Floc'h N. Is tryptophan catabolism increased under indoleamine 2,3 dioxygenase activity during chronic lung inflammation in pigs? Reproduction Nutrition Development. 2005;45(2):175–183. doi: 10.1051/rnd:2005013. [DOI] [PubMed] [Google Scholar]
- 53.Meisel R., Brockers S., Heseler K., et al. Human but not murine multipotent mesenchymal stromal cells exhibit broad-spectrum antimicrobial effector function mediated by indoleamine 2,3-dioxygenase. Leukemia. 2011;25(4):648–654. doi: 10.1038/leu.2010.310. [DOI] [PubMed] [Google Scholar]
- 54.Däubener W., Schmidt S. K., Heseler K., Spekker K. H., MacKenzie C. R. Antimicrobial and immunoregulatory effector mechanisms in human endothelial cells. Indoleamine 2,3-dioxygenase versus inducible nitric oxide synthase. Thrombosis and Haemostasis. 2009;102(6):1110–1116. doi: 10.1160/TH09-04-0250. [DOI] [PubMed] [Google Scholar]
- 55.Meisel R., Zibert A., Laryea M., Göbel U., Däubener W., Dilloo D. Human bone marrow stromal cells inhibit allogeneic T-cell responses by indoleamine 2,3-dioxygenase-mediated tryptophan degradation. Blood. 2004;103(12):4619–4621. doi: 10.1182/blood-2003-11-3909. [DOI] [PubMed] [Google Scholar]
- 56.Jui H. J., Lin C. H., Hsu W. T., et al. Autologous mesenchymal stem cells prevent transplant arteriosclerosis by enhancing local expression of interleukin-10, interferon-γ, and indoleamine 2,3-dioxygenase. Cell Transplantation. 2012;21(5):971–984. doi: 10.3727/096368911X627525. [DOI] [PubMed] [Google Scholar]
- 57.Su J., Chen X., Huang Y., et al. Phylogenetic distinction of iNOS and IDO function in mesenchymal stem cell-mediated immunosuppression in mammalian species. Cell Death and Differentiation. 2014;21(3):388–396. doi: 10.1038/cdd.2013.149. [DOI] [PMC free article] [PubMed] [Google Scholar]