


Abstract
Preincubation of the group I intron Ca.LSU from Candida albicans at 37°C in the absence of divalent cations results in partial folding of this intron. This is indicated by increased resistance to T1 ribonuclease cleavage of many G residues in most local helices, including P4-P6, as well as the non-local helix P7, where the G binding site is located. These changes correlate with increased gel mobility and activation of catalysis by precursor RNA containing this intron after preincubation. The presence of divalent cations or spermidine during preincubation results in formation of the predicted helices, as indicated by protection of additional G residues. However, addition of these cations during preincubation of the precursor RNA alters its gel mobility and eliminates the preincubation activation of precursor RNA seen in the absence of cations. These results suggest that, in the presence of divalent cations or spermidine, Ca.LSU folds into a more ordered, stable but misfolded conformation that is less able to convert into the catalytically active form than the ribozyme preincubated without cations. These results indicate that, like the group I intron of Tetrahymena, multiple folding pathways exist for Ca.LSU. However, it appears that the role cations play in the multiple folding pathways leading to the catalytically active form may differ between folding of these two group I introns.
INTRODUCTION
Group I introns are catalytically active RNA molecules that catalyze their own excision from the precursor RNA molecules in which they are inserted (1). Some of these introns can catalyze self-splicing in vitro in a two-step reaction resulting in excision of the intron and ligation of the flanking exons. In the first reaction, the 3′-hydroxyl of an exogenous guanosine or guanosine nucleotide attacks the phosphorus atom at the 5′ splice site to form a 3′,5′-phosphodiester bond to the 5′ nucleotide of the intron (I), leaving a free 3′-hydroxyl group on the 5′ exon (E1). This 3′-hydroxyl group then attacks the phosphorus at the 3′ splice site, resulting in ligation of the two exons (E1–E2) and release of the 5′-guanylylated intron (GI), as indicated in Figure 1. Over 90 group I introns are known, which share elements of secondary structure including at least nine double helices, denoted P1–P9, with the active guanosine-binding site involved in the cleavage and formation of phosphodiester linkages located in helix P7 (2). Despite their overall structural similarity, relatively few nucleotides are conserved among these introns, and they differ in many details of their apparent secondary structure within the general framework of group I intron structure (2).
Figure 1.
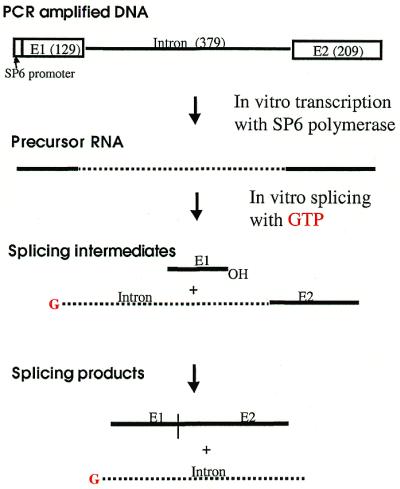
Schemes of in vitro transcription and self-splicing of Ca.LSU. To generate uniformly labeled precursor RNA containing Ca.LSU intron, [α-32P]UTP was included in the in vitro transcription. To generate the 5′-end-labeled Ca.LSU intron, [α-32P]GTP was used in the self-splicing of non-radiolabeled precursor RNA (shown here). See Materials and Methods for details. Note that E1 here is 130 nt long, consisting of 129 nt of the E1 sequence with a 5′-terminal G derived from the SP6 promoter consensus sequence.
Based on the phylogenetic comparisons of the many group I introns and site-specific mutagenesis, a structural model for these molecules has been proposed, in which the ribozyme is organized into three helical domains, P1-P2, P4-P6 and P3-P9, with the latter two domains forming the catalytic core of the ribozyme (2,3). Extensive studies on the structure of the group I intron in the 26S rRNA gene of Tetrahymena and its derivatives have confirmed and added details to this structure (3–7). Metal ions are required for tertiary folding of the catalytic RNA and also participate in catalysis (3–15). Only Mg2+ supports efficient catalysis by group I introns containing natural ribonucleotides, although some other divalent cations are able to support the folding of the Tetrahymena intron and quantitatively reduce its Mg2+ requirement (8,10,12,16). Spermidine was reported not to support the folding of this intron, although, like some divalent cations, it could reduce the Mg2+ requirement (10).
Extensive studies of the kinetics of folding of the Tetrahymena ribozyme have revealed that this RNA molecule folds to its native catalytically active structure through multiple intermediate states and pathways, which include misfolded and inactive states that can function as kinetic traps (10,15,17–22). Although it has been suggested that the intron can form most of its secondary structure in the absence of Mg2+, this cation appears to be required for more extensive folding, which in turn is required for catalytic activity of the ribozyme (10,17,23,24). Folding is believed to initiate with the P5abc domain, followed sequentially by folding of the P4-P6 domain, which then interacts with other parts of the ribozyme to more slowly form the remaining folded domains (5,6,15,17,19–21).
Ca.LSU is a self-splicing group I intron found in the 26S rRNA gene of many isolates of Candida albicans, the most common opportunistic fungal pathogen (25). We have found that splicing of this intron is sensitive to pentamidine in vitro and in vivo, and that inhibition of Ca.LSU splicing correlates with growth inhibition of strains harboring the intron. Furthermore, strains lacking this intron are relatively resistant to inhibition by pentamidine (26). The presence of similar group I introns in other human pathogens including Candida dubliniensis (27), Pneumocystis carinii (28), the mitochondrial genome of Aspergillus nidulans (29), Acanthamoeba (30) and Naegleria (31) suggests that these introns might be new targets for development of antimicrobial chemotherapeutic agents (25,28,32–39). Thus, the process of folding of these introns is of great interest. In the course of studies of the ribozyme activity of Ca.LSU, we noted that preincubation in buffer without divalent cations increased the enzymatic activity of this ribozyme (40). Since preincubation activation of the Tetrahymena ribozyme appears to require the presence of such cations, we further characterized the effect of preincubation on Ca.LSU, assaying the effect of this treatment by nuclease protection assays, gel mobility and ribozyme activity. Despite the great similarity between the structures of various group I introns (2), our results indicate that the different introns may follow different pathways that have different requirements and that generate different intermediates in the folding process.
MATERIALS AND METHODS
Preparation of the radiolabeled precursor and 5′-end-labeled intron
Genomic DNA extraction from C.albicans and PCR amplification of the DNA fragment containing the Ca.LSU intron sequence, flanking sequences and the bacteriophage SP6 promoter at the 5′-end of the sense strand (Fig. 1) were performed as described (26). This amplified DNA fragment was used to generate uniformly radiolabeled RNA transcripts using bacteriophage SP6 RNA polymerase (Promega) in reactions containing 500 µM rATP, rCTP, rGTP, 200 µM rUTP and 20 µCi [α-32P]UTP (3000 Ci/mmol, NEN-DuPont), following the manufacturer’s instructions. The transcripts were passed through a Sephadex G-50 column (Boehringer Mannheim) and precursor RNA containing Ca.LSU was purified by electrophoresis on a 5% polyacrylamide/8 M urea gel (28,41).
To prepare the 5′-end-labeled guanylylated Ca.LSU intron, non-radiolabeled precursor RNA was synthesized without isotope as described above, except that each rNTP was present at 500 µM. This precursor (1–2 µg) was then self-spliced under ribozyme assay conditions except for the presence of 50 µCi [α-32P]GTP (3000 Ci/mmol, NEN-DuPont) and 2.5 mM MgCl2 (Fig. 1), after which low molecular weight contaminants were removed using G-50 columns and the RNA was ethanol precipitated.
Native gel analysis of Ca.LSU
Purified uniformly radiolabeled precursor RNA (0.3 nM) was preincubated at 37°C or on ice (control) for the indicated period of time in 8 µl of 62.5 mM of Tris–HCl pH 7.5 containing 10 U of RNasin (Promega). Samples were then chilled on ice and analyzed by electrophoresis on 5% polyacrylamide gels in 1× TB buffer (0.045 M Tris–borate) at 4°C with a voltage gradient of 12–15 V/cm. The gels were then dried and exposed to X-ray film, and were quantified using a Bio-Rad GS700 Densitometer.
Ribonuclease T1 protection assay
The 5′-end-labeled Ca.LSU intron was preincubated in the indicated concentrations of Tris–HCl pH 7.5 in the presence or absence of various cations or GTP at 37°C for 20 min. Then ribonuclease T1 (0.006 U) was added to each sample and incubation was continued for 20 min. All samples were then chilled on ice and the cleaved products were separated on an 8% polyacrylamide/urea gel, followed by visualization by autoradiography. RNA ladders were generated by cleaving the 5′-end-labeled denatured Ca.LSU intron with 0.01 U of T1 for G, 0.2 U of U2 for A, 0.2 U of PhyM for A+U in buffers containing urea at 50°C.
Ribozyme assay
The splicing reaction of uniformly 32P-labeled precursor containing Ca.LSU was assayed in 10 µl reactions in the presence of 50 mM Tris–HCl pH 7.5, 1.25 mM MgCl2, 0.4 mM spermidine, 10 µM GTP and 10 U of RNasin (Promega) at 37°C for 20 min unless otherwise indicated. The optimum concentration of MgCl2 for the splicing reaction is 1.25 mM (26). Prior to the reaction, preincubation (where indicated) was performed in 8 µl in the presence of precursor RNA, indicated concentrations of pentamidine, cations or GTP at 37°C for the indicated period of time. After preincubation, 2 µl of reagent mix was added to each reaction so that the final concentration of each splicing component was the same as above, unless otherwise indicated. Then, the splicing reactions were performed at 37°C. The splicing reactions were analyzed by electrophoresis on 5% polyacrylamide/8 M urea gels, which were dried and exposed to X-ray film or scanned and analyzed using a phosphorimager (Bio-Rad GS-525 Molecular Imager). Note that in Figures 4 and 5, concentrations of all components present during preincubation are indicated based on the final 10 µl reaction volume. Thus, the concentrations were actually 25% higher than indicated in the preincubation before ribozyme reactions.
Figure 5.



Presence of oligovalent cations during preincubation prevents activation of Ca.LSU self-splicing. (A) Equal amounts of radiolabeled Ca.LSU precursor were preincubated in Tris–HCl pH 7.5 in the presence of 1.25 mM MgCl2, 0.4 mM spermidine, and 10 µM GTP, or with no additions (control) at 37°C for 5 min. Then the missing splicing components were added to standard concentrations, and MgCl2, spermidine and GTP were brought to standard splicing concentrations. Splicing reactions were run at 37°C for the indicated times. The splicing products were analyzed as in Figure 4B. (B) Equal amounts of radiolabeled precursor RNA were preincubated in Tris–HCl pH 7.5 in the presence of the indicated concentration of each cation at 37°C for 20 min, and then the splicing components were added to each sample to the standard concentrations except for 1.25 mM MgCl2 and 1.25 mM CaCl2 for all CaCl2 samples, and 2.5 mM MgCl2 for MgCl2 and spermidine samples; this change was necessary to maintain the same divalent cation concentration in all splicing reactions in each series of experiments. (C) Equal amounts of radiolabeled Ca.LSU precursor RNA were diluted in 10 µl of 50 mM Tris–HCl pH 7.5 with the indicated concentrations of MgCl2, and were incubated at the indicated temperatures for 20 min before analysis by native polyacrylamide gel electrophoresis.
RESULTS
Folding of Ca.LSU in Tris buffer
Ribonuclease T1 is a small (104 amino acids) globular endonuclease that hydrolyzes single-stranded RNA molecules with high specificity for cleavage on the 3′ side of G residues. G residues participating in base pairing or buried in a large molecule are resistant to cleavage by ribonuclease T1 (42,43). Ca.LSU intron was labeled with [α-32P]GTP at its 5′-terminus by self-splicing of precursor RNA transcribed in vitro in the presence of this nucleotide (Fig. 1). This radiolabeled full-length guanylylated intron was used for ribonuclease T1 protection studies because of its relatively small size (380 nt) and the efficiency and specificity of this labeling method. Furthermore, the lack of self-splicing activity by the excised intron made it easier to study than the larger catalytically active precursor RNA, which would undergo extensive self-splicing under conditions needed to 5′-label with polynucleotide kinase from bacteriophage T4. Ribonuclease T1 cleaved the denatured ribozyme to approximately the same extent at each G residue (Fig. 2A, lane G); similar uniform cleavage was observed (data not shown) under denaturing conditions (4 M urea) using ribonucleases U2 (A-specific) and PhyM (A,U-specific). In the absence of denaturant, ribonuclease T1 failed to cleave the RNA in water, although non-specific hydrolysis was seen at every nucleotide (Fig. 2A, lane 0). Note that this cleavage was not seen in undigested RNA [Fig. 3A, lane (–)]. The presence of 2 mM Tris–HCl pH 7.5 protected the RNA from non-specific hydrolysis, but did not support T1 cleavage. Ribonuclease T1 cleaved the intron RNA at G residues at 10–50 mM Tris–HCl pH 7.5, with a pattern of protection of many G residues in helical regions that was similar at these concentrations (Fig. 2A). This pattern is schematically represented in Figure 2B, where helical regions of secondary structure are designated P (for paired), loops at the end of helical domains are referred to by the number of the helix on which they are located, and unpaired junction regions between helices as J followed by the numbers of the flanking paired regions (2,25).
Figure 2.
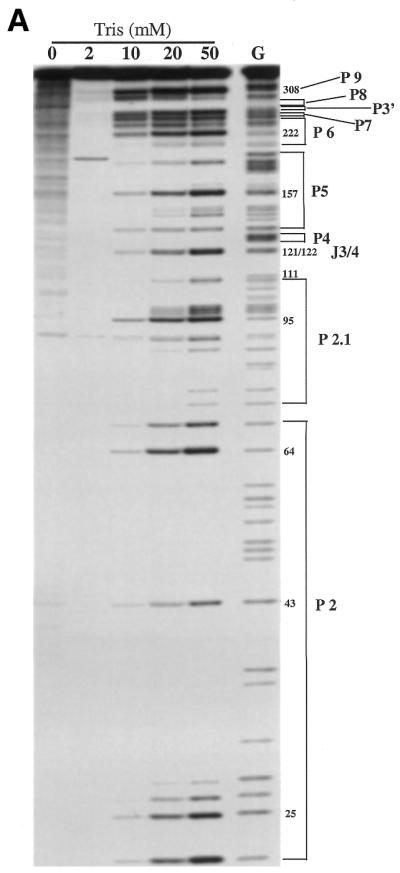

Effect of preincubation in Tris–HCl buffer on ribonuclease T1 sensitivity of Ca.LSU. The 5′-end labeled Ca.LSU intron was preincubated with the indicated concentration of Tris–HCl pH 7.5 and then cleaved by T1 ribonuclease as described in Materials and Methods. The G lane contains denatured Ca.LSU, which was cleaved without preincubation; the RNA in the other lanes was not denatured. The cleaved products were separated on multiple 8% polyacrylamide/urea gels. One of those gels is shown in (A): selected G residues are numbered and the corresponding helices are indicated. (B) T1 cleavage pattern of each G of Ca.LSU in 50 mM Tris is represented on the proposed secondary structure of the intron. Here arrows indicate sites of G cleavage by ribonuclease T1, with extent of cleavage (band intensity) being proportional to the size of the arrows, with the largest arrows representing the most cleavage and dots representing minimal cleavage; G residues without arrows or dots were not cleaved. The RNA sequence (25) has been confirmed by sequence analysis. The proposed guanosine binding site (G247) is circled. The secondary structure is slightly modified here based on our T1 protection results with divalent cations.
Figure 3.



Effect of cations on Ca.LSU folding assayed by ribonuclease T1 cleavage. The 5′-end-labeled Ca.LSU intron was assayed as in Figure 2, except that the indicated concentrations of cations were added to each sample together with 50 mM Tris–HCl pH 7.5. (A, opposite) A representative gel, showing the effects of MgCl2, CaCl2 and spermidine present during incubation prior to ribonuclease T1 digestion. Lane (–), undigested RNA. The 0°C lane represents RNA preincubated at that temperature rather than at 37°C, as were all other samples. (B, above) Effect of addition of MgCl2 or CaCl2 on ribonuclease T1 cleavage. This panel is represented as in Figure 2B, except orange symbols indicate sites at which ribonuclease T1 cleavage is reduced by incubation with these cations. Orange Gs, complete protection; orange arrows and dots, the reduced level of cleavage observed in the presence of these cations; black arrows and letters, no change relative to incubation in buffer alone; blue dots, sites at which ribonuclease T1 did not cleave Ca.LSU incubated in buffer alone, but at which cleavage was induced by incubation with these cations. (C, above) Spermidine effect on ribonuclease T1 cleavage. Green symbols, differences in the T1 cleavage pattern in the presence of spermidine as opposed to the pattern in the presence of MgCl2 or CaCl2 (B); orange letters, arrows and dots, sites at which the effect of spermidine was the same as that of MgCl2 or CaCl2; blue dots, sites at which ribonuclease T1 did not cleave Ca.LSU incubated in buffer alone, but at which cleavage was equivalently induced by incubation with divalent metal ions or spermidine.
Figure 2B shows that most of the protected G residues are located in helical regions of the Ca.LSU secondary structure, indicating that these helical structures can form in the absence of divalent cations. Most of the helical regions of P4-P6 were protected, with the exception of a number of G residues involved in G-U base pairs, which were partially cleaved. Strong cleavage was seen at the bulge between P5b and P5c, and in loop 6. The differential cleavage of the three adjacent Gs in the asymmetric bulge loop between P5b and P5c might indicate that the less cleaved Gs interact with some other structural element or are somehow protected inside some higher order structure resulting from tertiary interactions within the intron. Other helices that showed protection under these conditions included P8, P9.1 and P9.2, and the longest paired regions of P2 and P2.1. The Gs in ‘internal guide sequence’ of the incomplete helix P1 (which cannot form since the 3′-terminus of E1, with which they pair, is absent), were not cleaved under these conditions, perhaps reflecting their pairing with some other portion of the ribozyme. Strong cleavage was seen at most Gs in helices P3 and P9.0, indicating that the long distance interactions required for these structures were not established in Tris buffer. The formation of the short helix P7, where the proposed G binding site (G247) is located also involves long distance pairing. T1 protection was evident in three of the five Gs in this helix, suggesting that most of this helix was base paired, with the G247-C294 and G245-C295 base pairs being less stable. Two of the three helical Gs in the short P9 helix, which does not require long distance pairing, were cleaved by T1. Thus, in helical domain P3-P7 of Ca.LSU, helices P8, P9.1 and P9.2 form in Tris buffer while helix P7 is partially formed and helices P3, P9 and P9.0 may not form. Strong cleavages were observed in some (but not all) regions of P2 and P2.1, suggesting that these helices are not uniformly stable in Tris buffer. Cleavage was also evident in junction regions J3/4, J6/7 and J8/7, and at various predicted loop regions including loops 2, 2.1, 5c, 6, 9 and 9.2. Thus, the pattern of cleavage by ribonuclease T1 of Ca.LSU preincubated in Tris buffer without divalent cations indicates that most of the predicated helices in all three helical domains of the ribozyme had formed, including local secondary structure and some long distance tertiary interactions. On the other hand, P9.0 and portions of P2 and P2.1 did not form in Tris buffer, indicating that not all of the local secondary structure of Ca.LSU forms in the absence of divalent cations.
Effect of oligovalent cations on Ca.LSU folding
Folding of the Tetrahymena ribozyme is dependent on Mg2+ or other divalent cations. As expected from results with the Tetrahymena ribozyme, addition of MgCl2, CaCl2 or spermidine during preincubation caused the folding of Ca.LSU to a more ordered state, as indicated by reduced sensitivity to ribonuclease T1 at many G residues (Fig. 3A–C). A representative gel showing the effects of these divalent cations and of spermidine is shown in Figure 3A. This panel shows that both MgCl2 and CaCl2 altered the protection pattern at concentrations of 0.25–1.25 mM, with the protection being similar at all concentrations tested at most sites. On the other hand, spermidine showed dose-dependent protection at many sites in the concentration range of 0.05–0.4 mM, with 0.4 mM spermidine showing a similar extent of protection at most sites to that seen with 0.25 mM MgCl2 or CaCl2. Figure 3B schematically indicates the effects of divalent cations on the protection pattern. As indicated by the orange symbols, Mg2+ or Ca2+ reduced cleavage of many of the Gs in both helical and unpaired regions which were cleaved in its absence. The protection at helical Gs suggested that all predicted helices formed in the presence of divalent cations. The reduced cleavages in the junction regions J3/4, J6/7 and J8/7 suggest that the catalytic domains P4-P6 and P3-P9 fold to form a more ordered ‘core’ structure under these conditions. In addition, protection of many bulge and loop Gs suggests a more folded state of ribozyme Ca.LSU in the presence of these two divalent cations. This protection should result from tertiary interactions, including tetraloop interactions with tetraloop receptors, helix–helix interactions and phylogenetically conserved base pairs between otherwise unpaired bases in various regions of the ribozyme. In helix P7, where the predicted guanosine binding site G247 is located, the weakly cleaved G247 was completely protected and the moderately cleaved G245 was partially protected in the presence of divalent cations. Therefore, helix P7 is more ordered in the presence of Mg2+ or Ca2+. Increased protection at G256 and G257 suggests that P3 is also more ordered in the presence of divalent cations. Furthermore, divalent cations appear to induce complete base pairing of helices P9 and P9.0, which showed high ribonuclease T1 sensitivity in Tris buffer. Thus, the P3-P9 domain is much more ordered in the presence of Mg2+ or Ca2+. Some nucleotides were weakly sensitized to ribonuclease T1 cleavage by Mg2+ or Ca2+, including the five G residues in the internal guide sequence (G12–G16). It appears likely that the more ordered Ca.LSU conformation does not favor the pairing of these G residues with a C-rich region, thus making them accessible for cleavage. Cleavage of G111, located in J2.1/3, was induced by Mg2+, and more weakly by Ca2+ in a dose-dependent fashion; no such cleavage was seen with spermidine (Fig. 3A–C). Surprisingly, U273 in loop 8 was partially cleaved in the presence of the divalent cations; this cleavage was confirmed on other gels run in parallel with ladders generated under denaturing conditions with ribonucleases PhyM (A+U-specific) and U2 (A-specific, data not shown). Note that all of the regions sensitized to ribonuclease T1 are predicted not to be involved in secondary or tertiary structure interactions if Ca.LSU has similar structure to the Tetrahymena ribozyme.
As indicated in Figure 3C, 0.4 mM spermidine had effects similar to those of 0.25–1.25 mM MgCl2 and CaCl2. In this panel orange symbols indicate those nucleotides whose protection by spermidine is the same as by the divalent cations, and green symbols indicate sites where spermidine protection differed from that due to divalent cations. It can be seen that in P2, spermidine protected G25 and G26, and it also protected G146 in P5b, G144 and G184 in the bulge between P5a and P5b, in addition to the nucleotides that it protected as did the divalent cations. Spermidine was less protective than the divalent cations for G222 in L6 and G361 and G363 in L9.2. Thus, spermidine supports the secondary and tertiary folding of Ca.LSU although the structure induced by spermidine may differ from that formed in the presence of the divalent metal cations. Spermidine also sensitized cleavage at U23 near the base of P2 and U115 in P3 (Fig. 3C, green dots), in addition to cleavage at U273. Since ribonuclease T1 is highly specific for cleavage 3′ to G residues, we have no explanation for the few cleavages at U residues stimulated by incubation of Ca.LSU with divalent cations or spermidine, although we speculate that the folded state induced by these ionic species may make the phosphodiester bonds 3′ to these specific U residues susceptible to spontaneous cleavage, that seems a more probable explanation than alteration of the specificity of ribonuclease T1.
Preincubation of Ca.LSU precursor RNA in Tris buffer increases ribozyme activity
Preincubation in the presence of Mg2+, Ca2+ or spermidine protected many nucleotides of Ca.LSU from ribonuclease T1 in a pattern largely consistent with the phylogenetically conserved folding model for group I introns, with greater protection overall than was seen with preincubation in Tris buffer alone. Therefore, we tested the effect of preincubation with and without cations on ribozyme activity of precursor RNA. Note that although radioactive guanylylated intron and precursor RNA were prepared by different methods, the last step of both was alcohol precipitation in the presence of sodium acetate, and the precipitated RNA was diluted extensively (500–1000-fold) in each reaction. Thus divalent cation concentrations should be negligible. Preincubation in Tris buffer alone resulted in an increase in ribozyme activity of 26-fold (at 2 min assay time) or 6-fold (at 4 min assay time), with an approximate doubling of the fraction of RNA molecules capable of splicing relative to precursor preincubated on ice (Fig. 4A and B). The effect of preincubation was temperature dependent, and was seen after only 1 min at 37°C. Thus, unlike the Tetrahymena ribozyme, the changes induced in precursor RNA by incubation in the absence of divalent cations result in activation of the ribozyme. These results are consistent with the T1 protection results suggesting folding of the Ca.LSU intron in Tris buffer.
Figure 4.
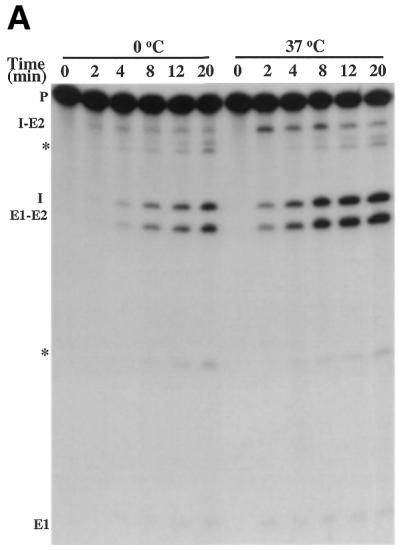

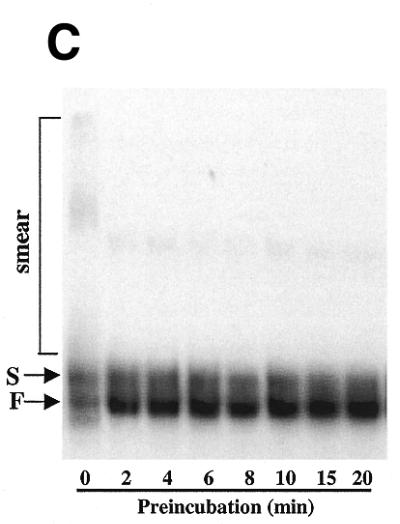
Effect of the preincubation on ribozyme folding and activity. (A) Equal quantities of uniformly labeled precursor RNA containing intron Ca.LSU were preincubated on ice or at 37°C for 1 min. Then the ribozyme reaction was initiated by addition of splicing components to the standard concentrations, and the reaction was run at 37°C for the indicated times. Self-splicing products were resolved on 5% polyacrylamide/8 M urea gels. RNA species are indicated on the left of the gel, with * indicating the products possibly generated by Mg2+-induced hydrolysis of the precursor RNA, E1 indicating 5′ exon, E2 indicating 3′ exon and I indicating intron. (B) Data from gels analyzed by a phosphorimager, where each curve represents the splicing of precursor RNA that has been preincubated as indicated. Splicing fraction of each reaction is the ratio of the splicing products (E1-E2 and GI) to total radioactivity (E1-E2, GI and precursor) in that lane. Preincubation without cations resulted in an increase in splicing rate of 26.8-fold at 2 min and 6.0-fold at 4 min. The splicing rates varied by 2% (2 min) or 8% (4min) in experiments carried out on different days using the same RNA preparation. (C) Preincubation of precursor RNA alters gel mobility. Equal amounts of radiolabeled precursor RNA were incubated in Tris–HCl pH 7.5 at 37°C for the indicated time (0 time was held on ice), and then subjected to native polyacrylamide gel electrophoresis (see Materials and Methods). The smear (seen more clearly on longer exposures) and the S and F bands are indicated.
Non-denaturing polyacrylamide gel electrophoresis was used to determine whether preincubation alters the conformation of the precursor RNA containing Ca.LSU. As can be seen in Figure 4C, preincubation of this RNA in Tris buffer at 37°C resulted in a rapid redistribution of RNA among forms having different gel mobilities. The precursor RNA transcript without preincubation existed in two bands, denoted S for slow and F for fast, as well as a smear of more slowly migrating forms. This polydisperse slowly migrating RNA, which can be seen more clearly on longer exposures, presumably represents a population of extended randomly folded and/or non-folded conformations. Within 2 min of preincubation, the earliest determination made, the amount of RNA in the F band increased 5.4-fold; this increase occurred by depletion of RNA in the smear that migrates more slowly than the S band. Note that in Figure 4C the same amount of RNA was loaded onto each lane. Thus, the increase in F band during preincubation in Tris buffer correlates with the increase in ribozyme activity, indicating that F band RNA is more active in ribozyme catalysis. Note that similar preincubation in Tris buffer increased ribonuclease T1 protection of multiple G residues in the guanylylated intron. Conversion of the precursor RNA to the fast migrating F form does not occur during preincubation at 0°C, indicating that the change occurring during preincubation is endothermic, as would be expected for a conformational change. Modest variation in amount of RNA loaded onto the gel and use of different RNA preparations did not significantly alter these results.
Preincubation of Ca.LSU precursor RNA with cations decreases ribozyme activation
Since the cationic species Mg2+, Ca2+ and spermidine each caused guanylylated Ca.LSU intron to fold into more ordered states, we tested the effect of adding these molecules or GTP during preincubation of precursor RNA containing Ca.LSU. Preincubation of precursor RNA for 5 min in the presence of Tris–HCl pH 7.5 with or without 12.5 µM GTP resulted in similar increases in ribozyme activity. However, the presence of 1.5 mM MgCl2 or 0.4 mM spermidine during preincubation eliminated ribozyme activation (Fig. 5A). As shown in Figure 5B, the effect of MgCl2, CaCl2 or spermidine added to the preincubation was dose-dependent, with Mg2+ or Ca2+ reducing splicing activity to about the basal level at 0.2 mM, while 0.05 mM spermidine reduced splicing activity to about the same level. Preincubation of precursor RNA in the presence of varying concentrations of MgCl2 (0.2–1.25 mM) resulted in an altered gel mobility pattern showing an approximate equal distribution of RNA between the F form and a species comigrating with the S form, accompanied by disappearance of the diffuse RNA smear and increased intensity of both the F and S bands (Fig. 5C). The arrow in Figure 5C indicates another more rapidly migrating RNA species only seen after preincubation with 1.25 mM MgCl2 (which was also seen at higher concentrations, data not shown). A similar increase in a band at the S position at the expense of the band at the F position was seen after preincubation of precursor RNA in the presence of CaCl2 or spermidine, and the change occurred within 2 min of preincubation (data not shown). Thus, preincubation in the presence of any of the three cationic species altered the conformation of the guanylylated intron in similar ways, based on ribonuclease T1 protection assays; similarly all three cations prevented preincubation activation of precursor RNA containing Ca.LSU and altered the electrophoretic mobility of that RNA on native polyacrylamide gels. These results suggest that the more ordered Ca.LSU conformations formed by preincubation in the presence of the cations tested are metastable misfolded structures that may refold into the active conformation slowly, as suggested for the Tetrahymena intron (18).
DISCUSSION
The folding of group I intron ribozymes has been extensively studied using various derivatives of the Tt.LSU ribozyme from Tetrahymena. It has been suggested that, while much of the secondary structure of the ribozyme can form during preincubation in the absence of divalent cations, complete folding to the catalytically active form of the ribozyme requires these cations (10,23–24). Folding of the Tetrahymena ribozyme appears to initiate with folding of the P5abc secondary structure, and is followed sequentially by folding of the P4-6 domain, and then the other portions of the ribozyme (5,15,17,19–21). Specific regions of the ribozyme have been suggested to play critical roles in this folding process. These include the branched helical structure of P5abc, which includes a GAAA tetraloop at loop 5b. This tetraloop binds to an A-rich bulge of helix P6 (6). Similarly, the GAAA tetraloop at loop 9 docks in the minor groove of P5 (2,3). Adding to the complexity of the folding problem, this ribozyme folds to its native catalytically active structure through multiple intermediate states and pathways, which include misfolded and inactive states that can function as kinetic traps (10,14–15,17–22).
Ca.LSU and Tt.LSU are both members of the IC class of group I introns, based on their structural similarities. Despite these similarities, many differences in sequence exist between these two ribozymes, as is generally observed for members of the entire group I intron family (2). Comparison of the sequence (as depicted in Fig. 2B) of Ca.LSU (25) and that of the L-21 ScaI derivative of the Tetrahymena ribozyme (44) reveals that there are a number of differences relevant to the folding problem. The P5abc domain, which appears to be the first region of the Tetrahymena ribozyme to fold, has a branched helical structure in that ribozyme but not in Ca.LSU. Furthermore, the GAAA tetraloop of the Tetrahymena ribozyme is not present on loop 5b of Ca.LSU, and the sequence of the tetraloop docking site in helix P6 is also not conserved. Ca.LSU has a GAAA tetraloop at loop 2, unlike the Tetrahymena ribozyme. The structure of the helices P9-P9.2 in Ca.LSU shows many differences from the Tetrahymena ribozyme, with GAAA tetraloops at loop 9.2 in Tetrahymena and loop 9 in Ca.LSU. Furthermore, the sequences of the relatively conserved elements P4, P6 and P7 of Ca.LSU are among the most divergent from the consensus among the group I introns (25). These and other differences between these RNA molecules suggest the possibility that there should be differences in their secondary and tertiary structures, as well as in the folding pathways and intermediates by which these structures are formed.
It previously has been suggested that local base pairings of the Tetrahymena ribozyme can form in Tris buffer, and that long distance pairing (P3 and P7) that involves tertiary folding only occurs in the presence of Mg2+ (17). Ribonuclease T1 protection shows that many local helical regions of Ca.LSU, especially in the core P4-P6 domain, become protected when guanylylated intron is preincubated in Tris buffer (Fig. 2), although some long distance pairings in P7 also appear to occur. Under these preincubation conditions, precursor RNA containing Ca.LSU is activated (Fig. 4A and B), and its electrophoretic mobility on a native gel is changed from a pattern with a broad slowly migrating smear and two bands, denoted S and F, to a pattern in which the F band predominates (Fig. 4C). These data support the idea that preincubation in Tris buffer causes an altered conformation of Ca.LSU, and this conformation is along a pathway to the catalytically active form of the ribozyme. We cannot exclude the possibility that divalent cations act by slowing the exchange among base paired structures, so that refolding cannot occur to the active conformation within 20 min. However, prolonged incubation of precursor RNA in the presence of Mg2+ results in RNA hydrolysis (data not shown), that has prevented further testing of this alternative explanation.
In the case of the Tetrahymena ribozyme and its derivatives, the addition of divalent metal ions stabilizes the tertiary structure by strengthening base pairing interactions (7,10,15). Based on ribonuclease T1 protection assays, Mg2+ and Ca2+ have a similar effect on Ca.LSU, with the effects of these two cations being virtually identical. In the case of Ca.LSU, spermidine has similar but not identical effects to the two metallic cations. The apparent conformational change exerted by these three cationic species on Ca.LSU is in agreement with the alteration in mobility of precursor containing Ca.LSU to a slower migrating band upon pre-incubation in their presence (Fig. 5C). However, unlike the situation for the Tetrahymena ribozyme, this cation-induced change results in failure of preincubation to activate Ca.LSU (Fig. 5A and B). Note that the same dose of Mg2+ or Ca2+ is required to alter ribonuclease T1 protection of the guanylylated intron, to alter the mobility of precursor RNA on a native electrophoresis gel and to decrease preincubation activation of the ribozyme.
Previously it has been noted that precursor RNA containing Tt.LSU also migrates as multiple bands on a native electrophoretic gel, and incubation at 45°C in the presence of magnesium promotes conversion of other forms into rapidly migrating ‘band 4’, that is more active than the slower ‘band 1’ in reacting with GTP, although both forms can participate in exon ligation (45). Both ribozymes Tt.LSU and Ca.LSU can fold to generate much of the correct secondary structure of a group I intron in Tris buffer without divalent cations or GTP. The presence of divalent cations during preincubation results in increased base pairing interactions in both ribozymes. However, this divalent cation-dependent change in conformation appears to be on the pathway to correct folding of the Tt.LSU ribozyme and its derivatives, while in the case of Ca.LSU this change results in folding which is not on the correct folding pathway. The overall folded structure of the group I introns appears to be highly conserved, based on the stronger conservation of paired elements which generate the folded structure than of primary sequence of the introns (2). However, not surprisingly, the divergence between the sequences and some structural elements of these introns makes some differences in their folding pathways, such as those suggested by the results of this work.
Recently, it has been found that helices P5a, P5b and P5c of the Tetrahymena ribozyme, which define a tertiary structure subdomain, can fold at least 25 times more rapidly in isolation than when allowed to fold when interaction between this subdomain and the rest of the P4-P6 domain is possible (46). These results suggest that, in the process of folding of this group I intron, non-native compact conformations can form that compete with the native (active) structure. A model is presented (46) in which unfolded RNA can collapse into native or non-native compact conformations. Based on sedimentation experiments, Mg2+ appears to be required for collapse of unfolded P4-P6 RNA into the non-native compact state. Mg2+ dependent misfolding is consistent with the ability of high Mg2+ concentrations to stabilize misfolded intermediates and to lower the rate of folding of Tetrahymena pre-rRNA (47–49).
Group I introns are found in the essential chromosomal rRNA genes of a number of eukaryotic microbial pathogens (27–31), including ∼40% of isolates of C.albicans (25). In addition, fungal pathogens may have group I introns in their mitochondrial genomes, as does the yeast Saccharomyces cerevisiae (50). It has been suggested that the group I introns, which are not found in the human genome, might be a new target for antimicrobial chemotherapy (25,28,32–39,51). We have demonstrated that pentamidine inhibits the splicing of group I introns in living cells of C.albicans, and that this inhibition correlates with inhibition of cellular growth (26), supporting the validity of this new target. Understanding the structure and folding pathways of Ca.LSU and other ribozymes might help to predict their sensitivity to different inhibitors. Although it is not known how much sensitivity to various inhibitors might vary between group I introns, we have found that while both Pc1.LSU and Ca.LSU show similar in vitro sensitivity to pentamidine and its derivatives, Ca.LSU is completely resistant to inhibition by tetracycline at doses that completely block catalysis by Pc1.LSU (26,35). We look forward to learning more about the structures and folding pathways of different group I introns, especially those that might represent new antimicrobial targets in pathogenic organisms.
Acknowledgments
ACKNOWLEDGEMENTS
We thank G.Lemay (University of Montreal, Montreal, Canada) for the C.albicans strain harboring Ca.LSU, and D.E.Georgopoulos and K.E.Miletti for their assistance. This work was supported by NIH grant 1RO1 GM53815 and a grant from the Foundation for UMDNJ.
References
- 1.Cech T.R. (1990) Self-splicing of group I introns. Annu. Rev. Biochem., 59, 543–568. [DOI] [PubMed] [Google Scholar]
- 2.Michel F. and Westhof,E. (1990) Modelling of the three-dimensional architecture of group I catalytic introns based on comparative sequence analysis. J. Mol. Biol., 216, 585–610. [DOI] [PubMed] [Google Scholar]
- 3.Golden B.L., Gooding,A.R., Podell,E.R. and Cech,T.R. (1998) A preorganized active site in the crystal structure of the Tetrahymena ribozyme. Science, 282, 259–264. [DOI] [PubMed] [Google Scholar]
- 4.Cate J.H., Gooding,A.R., Podell,E., Zhou,K., Golden,B.L., Kundrot,C.E., Cech,T.R. and Doudna,J.A. (1996) Crystal structure of a group I ribozyme domain: principles of RNA packing. Science, 273, 1678–1685. [DOI] [PubMed] [Google Scholar]
- 5.Cate J.H., Hanna,R.L. and Doudna,J.A. (1997) A magnesium ion core at the heart of a ribozyme domain. Nat. Struct. Biol., 4, 553–558. [DOI] [PubMed] [Google Scholar]
- 6.Cate J.H., Gooding,A.R., Podell,E., Zhou,K., Golden,B.L., Szewczak,A.A., Kundrot,C.E., Cech,T.R. and Doudna,J.A. (1996) RNA tertiary structure mediation by adenosine platforms. Science, 273, 1696–1699. [DOI] [PubMed] [Google Scholar]
- 7.Wu M. and Tinoco,I.,Jr (1998) RNA folding causes secondary structure rearrangement. Proc. Natl Acad. Sci. USA, 95, 11555–11560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Piccirilli J.A., Vyle,J.S., Caruthers,M.H. and Cech,T.R. (1993) Metal ion catalysis in the Tetrahymena ribozyme reaction. Nature, 361, 85–88. [DOI] [PubMed] [Google Scholar]
- 9.Pyle A.M. (1996) Role of metal ions in ribozymes. Met. Ions Biol. Syst., 32, 479–520. [PubMed] [Google Scholar]
- 10.Celander D.W. and Cech,T.R. (1991) Visualizing the higher order folding of a catalytic RNA molecule. Science, 251, 401–407. [DOI] [PubMed] [Google Scholar]
- 11.Streicher B., Westhof,E. and Schroeder,R. (1996) The environment of two metal ions surrounding the splice site of a group I intron. EMBO J., 15, 2556–2564. [PMC free article] [PubMed] [Google Scholar]
- 12.Weinstein L.B., Jones,B.C., Cosstick,R. and Cech,T.R. (1997) A second catalytic metal ion in group I ribozyme. Nature, 388, 805–808. [DOI] [PubMed] [Google Scholar]
- 13.McConnell T.S., Herschlag,D. and Cech,T.R. (1997) Effects of divalent metal ions on individual steps of the Tetrahymena ribozyme reaction. Biochemistry, 36, 8293–8303. [DOI] [PubMed] [Google Scholar]
- 14.Shan S.O. and Herschlag,D. (1999) Probing the role of metal ions in RNA catalysis: kinetic and thermodynamic characterization of a metal ion interaction with the 2′-moiety of the guanosine nucleophile in the Tetrahymena group I ribozyme. Biochemistry, 38, 10958–10975. [DOI] [PubMed] [Google Scholar]
- 15.Chaulk S.G. and MacMillan,A.M. (2000) Characterization of the Tetrahymena ribozyme folding pathway using the kinetic footprinting reagent peroxynitrous acid. Biochemistry, 39, 2–8. [DOI] [PubMed] [Google Scholar]
- 16.Christian E.L. and Yarus,M. (1993) Metal coordination sites that contribute to structure and catalysis in the group I intron from Tetrahymena. Biochemistry, 32, 4475–4480. [DOI] [PubMed] [Google Scholar]
- 17.Zarrinkar P.P. and Williamson,J.R. (1994) Kinetic intermediates in RNA folding. Science, 265, 918–924. [DOI] [PubMed] [Google Scholar]
- 18.Pan J., Thirumalai,D. and Woodson,S.A. (1997) Folding of RNA involves parallel pathways. J. Mol. Biol., 273, 7–13. [DOI] [PubMed] [Google Scholar]
- 19.Sclavi B., Sullivan,M., Chance,M.R., Brenowitz,M. and Woodson,S.A. (1998) RNA folding at millisecond intervals by synchrotron hydroxyl radical footprinting. Science, 279, 1940–1943. [DOI] [PubMed] [Google Scholar]
- 20.Treiber D.K., Rook,M.S., Zarrinkar,P.P. and Williamson,J.R. (1998) Kinetic intermediates trapped by native interactions in RNA folding. Science, 279, 1943–1946. [DOI] [PubMed] [Google Scholar]
- 21.Russell R. and Herschlag,D. (1999) New pathways in folding of the Tetrahymena group I RNA enzyme. J. Mol. Biol., 291, 1155–1167. [DOI] [PubMed] [Google Scholar]
- 22.Zhuang X., Bartley,L.E., Babcock,H.P., Russell,R., Ha,T., Herschlag,D. and Chu,S. (2000) A single-molecule study of RNA catalysis and folding. Science, 288, 2048–2051. [DOI] [PubMed] [Google Scholar]
- 23.Jaeger J.A., Zuker,M. and Turner,D.H. (1990) Melting and chemical modification of a cyclized self-splicing group I intron: similarity of structures in 1 M Na+, in 10 mM Mg2+, and in the presence of substrate. Biochemistry, 29, 10147–10158. [DOI] [PubMed] [Google Scholar]
- 24.Banerjee A.R., Jaeger,J.A. and Turner,D.H. (1993) Thermal unfolding of a group I ribozyme: the low-temperature transition is primarily disruption of tertiary structure. Biochemistry, 32, 153–163. [DOI] [PubMed] [Google Scholar]
- 25.Mercure S., Montplaisir,S. and Lemay,G. (1993) Correlation between the presence of a self-splicing intron in the 25S rDNA of C.albicans and strains susceptibility to 5-fluorocytosine. Nucleic Acids Res., 21, 6020–6027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Miletti K.E. and Leibowitz,M.J. (2000) Pentamidine inhibition of group I intron splicing in Candida albicans correlates with growth inhibition. Antimicrob. Agents Chemother., 44, 958–966. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Boucher H., Mercure,S., Montplaisir,S. and Lemay,G. (1996) A novel group I intron in Candida dubliniensis is homologous to a Candida albicans intron. Gene, 180, 189–196. [DOI] [PubMed] [Google Scholar]
- 28.Liu Y. and Leibowitz,M.J. (1993) Variation and in vitro splicing of group I introns in rRNA genes of Pneumocystis carinii. Nucleic Acids Res., 21, 2415–2421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Netzker R., Kochel,H.G., Basak,N. and Kuntzel,H. (1982) Nucleotide sequence of Aspergillus nidulans mitochondrial genes coding for ATPase subunit 6, cytochrome oxidase subunit 3, seven unidentified proteins, four tRNAs and L-rRNA. Nucleic Acids Res., 15, 4783–4794. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Gast R.J., Fuerst,P.A. and Byers,T.J. (1994) Discovery of group I introns in the nuclear small subunit ribosomal RNA genes of Acanthamoeba. Nucleic Acids Res., 22, 592–596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Embley T.M., Dyal,P. and Kilvington,S. (1992) A group I intron in the small subunit ribosomal RNA gene from Naegleria andersoni ssp. andersoni strain PPMFB-6. Nucleic Acids Res., 20, 6411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.von Ahsen U., Davies,J. and Schroeder,R. (1991) Antibiotic inhibition of group I ribozyme function. Nature, 353, 368–370. [DOI] [PubMed] [Google Scholar]
- 33.von Ahsen U., Davies,J. and Schroeder,R. (1992) Non-competitive inhibition of group I intron RNA self-splicing by aminoglycoside antibiotics. J. Mol. Biol., 226, 935–941. [DOI] [PubMed] [Google Scholar]
- 34.Liu Y. and Leibowitz,M.J. (1995) Bidirectional effectors of a group I intron ribozyme. Nucleic Acids Res., 23, 1284–1291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Liu Y., Tidwell,R.R. and Leibowitz,M.J. (1994) Inhibition of in vitro splicing of a group I intron of Pneumocystis carinii. J. Eukaryot. Microbiol., 41, 31–38. [DOI] [PubMed] [Google Scholar]
- 36.Mercure S., Cousineau,L., Montplaisir,S., Belhumeur,P. and Lemay,G. (1997) Expression of a reporter gene interrupted by the Candida albicans group I intron is inhibited by base analogs. Nucleic Acids Res., 25, 431–437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Wank H., Rogers,J., Davies,J. and Schroeder,R. (1994) Peptide antibiotics of the tuberactinomycin family as inhibitors of group I intron RNA splicing. J. Mol. Biol., 236, 1001–1010. [DOI] [PubMed] [Google Scholar]
- 38.Rogers J. and Davies,J. (1994) The pseudodisaccharides: a novel class of group I intron splicing inhibitors. Nucleic Acids Res., 22, 4983–4988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Mei H.Y., Cui,M., Sutton,S.T., Truong,H.N., Chung,F.Z. and Czarnik,A.W. (1996) Inhibition of self-splicing group I intron RNA: high-throughput screening assays. Nucleic Acids Res., 24, 5051–5053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Zhang Y. (1999) Group I introns: potential antimicrobial targets. PhD dissertation. UMDNJ Graduate School of Biomedical Sciences, Piscataway, NJ.
- 41.Zhang Y., Bell,A., Perlman,P.S. and Leibowitz,M.J. (2000) Pentamidine inhibits mitochondrial intron splicing and translation in Saccharomyces cerevisiae. RNA, 6, 937–951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Milan S., D’Ari,L. and Chamberlin,M.J. (1999) Structural analysis of ternary complexes of Escherichia coli RNA polymerase: ribonuclease footprinting of the nascent RNA in complexes. Biochemistry, 38, 218–225. [DOI] [PubMed] [Google Scholar]
- 43.Benito Y., Kolb,F.A., Romby,P., Lina,G., Etienne,J. and Vandenesch,F. (2000) Probing the structure of RNAIII, the Staphylococcus aureus agr regulatory RNA, and identification of the RNA domain involved in repression of protein A expression. RNA, 6, 668–679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Zaug A.J., Grosshans,C.A. and Cech,T.R. (1988) Sequence-specific endoribonuclease activity of the Tetrahymena ribozyme: enhanced cleavage of certain oligonucleotide substrates that form mismatched ribozyme-substrate complexes. Biochemistry, 27, 8924–8931. [DOI] [PubMed] [Google Scholar]
- 45.Emerick V.L. and Woodson,S.A. (1994) Fingerprinting the folding of a group I precursor RNA. Proc. Natl Acad. Sci. USA, 91, 9675–9679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Deras M.L., Brenowitz,M., Ralston,C.Y., Chance,M.R. and Woodson,S.A. (2000) Folding mechanism of the Tetrahymena ribozyme P4-P6 domain. Biochemistry, 39, 10975–10985. [DOI] [PubMed] [Google Scholar]
- 47.Fang X.W., Pan,T. and Sosnick,T.R. (1999) Mg2+-dependent folding of a large ribozyme without kinetic traps. Nat. Struct. Biol., 6, 1091–1095. [DOI] [PubMed] [Google Scholar]
- 48.Pan J., Thirumalai,D. and Woodson,S.A. (1999) Magnesium-dependent folding of self-splicing RNA: exploring the link between cooperativity, thermodynamics, and kinetics. Proc. Natl Acad. Sci. USA, 96, 6149–6154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Rook M.S., Treiber,D.K. and Williamson,J.R. (1999) An optimal Mg(2+) concentration for kinetic folding of the Tetrahymena ribozyme. Proc. Natl Acad. Sci. USA, 96, 12471–12476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Dujon B. (1989) Group I introns as mobile genetic elements: facts and mechanistic speculations—a review. Gene, 82, 91–114. [DOI] [PubMed] [Google Scholar]
- 51.von Ahsen U. and Schroeder,R. (1991) Streptomycin inhibits splicing of group I introns by competition with the guanosine substrate. Nucleic Acids Res., 19, 2261–2265. [DOI] [PMC free article] [PubMed] [Google Scholar]