

Abstract
Purpose of review
This review addresses the most recent developments on cockroach allergen research in relation to allergic diseases, especially asthma.
Recent findings
The number of allergens relevant to cockroach allergy has recently expanded considerably up to 12 groups. New X-ray crystal structures of allergens from groups 1, 2 and 5 revealed interesting features with implications for allergen standardization, sensitization, diagnosis and therapy.
Summary
Cockroach allergy is strongly associated with asthma particularly among children and young adults living in inner-city environments, posing challenges for disease control. Environmental interventions targeted at reducing cockroach allergen exposure have provided conflicting results. Immunotherapy may be a way to modify the natural history of cockroach allergy and decrease symptoms and asthma severity among sensitized and exposed individuals. The new information on cockroach allergens is important for the assessment of allergen markers of exposure and disease, and for the design of immunotherapy trials.
Keywords: Cockroach allergy, Asthma, IgE, Allergy Diagnosis, Recombinant Allergens, Allergen Structure
Introduction
Cockroach allergy has been established as an important cause of asthma for over 50 years. Bernton and Brown, in 1964, were the first to report the presence of positive skin tests to cockroach extract among patients living in New York [1]. Subsequent studies by Kang et al. established a causal relationship between cockroach allergy and asthma by demonstrating bronchoconstriction following inhalation of cockroach extract by cockroach allergic asthmatic patients [2].
Environmental exposure to cockroach has been linked to cockroach sensitization and asthma. A large number of cockroach species exist worldwide (approximately 4,000 species), but only few are domiciliary cockroaches, including around 25 species (Order Blattaria, Class Insecta, Phyllum Arthropoda) [3]. Two species, German cockroach (Blattella germanica) and American cockroach (Periplaneta americana) predominate in temperate and tropical areas, respectively. The German cockroach prefers cool and dry climates (i.e. Europe and USA), whereas the larger American cockroach is common in hot and humid conditions (i.e. Taiwan, Brazil and Thailand) [4,5]. These two species are the main focus of allergy research. Sensitization to cockroach allergens is one of the strongest risk factors for the development of asthma in low-income urban populations worldwide. A recent review by Do et al. highlights that the prevalence of cockroach allergy ranges from 17 to 41% in the United States (US), with cockroach allergens being detected in 85% of inner-city US homes. Based on skin prick testing, 60–80% of inner-city children with asthma are sensitized to cockroach [6].
This review describes progress on research about cockroach allergens, especially within the last three years, and implications for diagnosis and therapy.
Cockroach allergy and inner-city asthma
It is well established that inner-city children present a high burden of asthma symptoms and morbidity even when complying with guidelines-directed management [7]●●[8]. Cockroach and mouse represent a significant percentage of allergic sensitization in this population, which is highly exposed to allergens from both sources [9,10]●●[11]. In particular, the combination of exposure and sensitization to cockroach appears to have a greater effect on morbidity and severity of asthma than exposure and sensitization to other allergens including dust mites or pets, among inner-city children and young adults with asthma [12]. Association with increased severity was also described among cockroach allergic adult patients with asthma [6,13]. Likewise, mouse-sensitized urban children are at increased risk for asthma prevalence and severity [9,14,15]. More than 50% of urban children with asthma are sensitized to either mouse or cockroach, and co-exposure is common [9]. Ahluwalia et al. observed that sensitization/exposure to both mouse and cockroach was associated with worse asthma in an inner-city community in Baltimore exposed to high levels of both allergens. However, mouse allergen appeared to be more strongly and consistently associated with poor asthma outcomes than cockroach allergen [16,17].
The Inner-City Asthma Consortium - Asthma Phenotypes in the Inner City (APIC) study contributed to understanding the role of host and environmental factors influencing severity of asthma among inner-city children and adolescents in the US ●●[18,19]. Results of this study revealed that, in spite of good adherence to optimal treatment, inner-city children and adolescents with difficult-to-control asthma had little improvement in symptoms, exacerbations, and pulmonary physiology over a one-year follow-up. Bronchodilator responsiveness, pulmonary physiology measures, atopy and rhinitis severity were associated with a necessity for high-dose asthma controller therapy ●●[8]. In particular, total serum IgE levels, mold sensitization, and the total number of allergen sensitizations at baseline distinguished patients with difficult-to-control asthma, from those with easy-to-control disease. Sensitization to cockroach and mouse was not found to be significantly associated with severity, probably due to the fact that the relevance of these allergens to disease is strongly linked to both sensitization and exposure [12].
A conceptual model was constructed with data from 561 children 6- to 17-year-old with asthma and rhinitis from 9 US inner cities, in the APIC study, who were prospectively evaluated every 2 months for 1 year. Allergen sensitization, allergic inflammation, pulmonary physiology, environmental tobacco smoke (ETS) exposure, and rhinitis severity were significantly related to asthma severity, whereas stress, obesity, and vitamin D showed no significant effect on disease severity ●●[18]. Allergen sensitization was the strongest association detected (linked to allergic inflammation, and subsequently to pulmonary physiology and rhinitis severity) to contribute to asthma severity, suggesting that allergen sensitization could be the originating domain among the multiple pathways involved in severity of the disease ●●[18].
In conjunction, these data highlight the importance of allergen sensitization and its consequences in children with asthma in low-income areas of US inner cities ●●[11]. Personalized care in this population may require emphasis on environmental allergen management, allergen desensitization, and anti-Th2 therapy, which would be appropriate for most asthma phenotypes. In this regard, total IgE measurement and assessment of IgE to environmental allergens, including cockroach and mouse, are among the priorities for identifying inner-city children with difficult-to-control asthma ●●[8].
Environmental interventions and anti-IgE therapy
The findings from studies of the Inner-City Asthma Consortium (ICAC) have shed light on the causes and treatment of asthma exacerbations among underprivileged and minority children. A comprehensive environmental intervention study, including specific measures addressing cockroach allergen exposure, made it possible to reduce allergen levels in the homes of children with asthma, and this decrease was associated with improvement in symptoms. However, lesser improvement in asthma exacerbations was achieved; therefore the intervention was not sufficient to bring the children to optimal asthma control [20]. Other locations, including schools and day care, may be important sources for continued exposure [21].
Portnoy et al. developed a comprehensive practice parameter for assessment of environmental cockroach exposure and methods for allergen reduction and avoidance [22]. In addition to evidence of the presence of cockroaches by visual inspection or sticky trap placement, measurement of cockroach allergen levels could be useful to guide environmental interventions aimed at reducing cockroach exposure. The practice parameter acknowledges that a cutoff of 0.04 µg/g dust for cockroach allergen Bla g 2 has been proposed as a threshold below which sensitization is prevented, and that levels greater than 0.08 µg/g have been associated with development of disease and symptoms [22]. In fact, 2 U/g and 8 U/g of Bla g 1 or Bla g 2 have been considered thresholds of sensitization and asthma morbidity, respectively, for a long time (1 unit of Bla g 1 is equivalent to 0.1 µg [23] and 1 unit of Bla g 2 is equivalent to 0.04 µg [22,24,25]). However, there is evidence supporting a dose-response relationship between exposure and sensitization or disease, rather than the existence of specific thresholds. Bla g 1 levels greater than 0.1 µg/g dust or Bla g 2 levels greater than 0.04 µg/g have indicated an increased likelihood that there are sources of cockroach allergen production; therefore, it has been recommended that Bla g 1 levels ideally should be reduced to less than twice these levels. Elimination of cockroaches carried out through an integrated pest management and removal of reservoirs of cockroach contaminants were the strongest recommendations to decrease cockroach exposure and reduce asthma morbidity [22].
A recent randomized controlled trial was conducted to determine the effect of a multifaceted indoor allergen avoidance intervention for over 40 weeks in reducing asthma controller therapy in adults and children with asthma residing in New York City, who were both sensitized and exposed to at least one indoor allergen, including cockroach ●[26]. The results revealed that the individualized intervention leading to effective reduction of household allergens (cockroach, mouse, dust mite, cat, and dog), did not allow for a significant reduction in therapy or improvement in control of asthma as compared with the non-intervention group ●[26]. On the other hand, a 12-month randomized trial of a single intervention, placement of insecticidal bait, resulted in sustained cockroach elimination and was associated with improved asthma outcomes in a group of 102 children aged 5 to 17 years with moderate to severe asthma living in New Orleans ●[27]. In particular, children in control homes had more asthma symptoms and unscheduled health care utilization, and lower pulmonary function as compared with children from the intervention homes. The beneficial effect was more pronounced in children exposed and sensitized to cockroach [22].
It is becoming evident that, despite excellent control of symptoms by guidelines-based conventional asthma treatment, exacerbations still remained a significant problem that cannot be controlled in many inner-city patients, particularly during the seasonal pattern of fall peaks [19]●[28]. Two studies clearly demonstrated the added benefit of omalizumab to guidelines-based therapy in reducing fall asthma exacerbations and decreasing need for asthma control medications [29]●[30]. These were the Inner-City Anti-IgE Therapy for Asthma (ICATA) trial, which included children and young adults 6 to 20 years of age with moderate-to-severe asthma treated for one year, and the Preventative Omalizumab or Step-up Therapy for Severe Fall Exacerbations (PROSE) trial, which targeted fall exacerbations by beginning the intervention with omalizumab shortly before the start of school and continuing it through the fall season. Strikingly, ICATA patients both sensitized and exposed to cockroach (Bla g 1 levels ≥ 2 U/g) in their homes benefited most from omalizumab treatment, with a 71.2% reduction in exacerbations, greater improvement of symptoms, and greater reduction of dose of corticosteroids, as compared to the other study participants. These data suggested that combination of sensitization and exposure to cockroach could serve as marker for good clinical response to treatment with omalizumab among inner-city children and young adults with allergic asthma [29]. A recent study addressing children and adolescents with rhinovirus-triggered asthma exacerbations has shown that baseline mouse-specific IgE and baseline dust mite-specific IgE levels each were associated with rhinovirus infection to increase severity ●●[31]. Subjects who were rhinovirus positive at the time of asthma exacerbation demonstrated an increase in mouse-specific, dust mite-specific and total IgE levels, compared to levels obtained when they had returned to their symptomatic baseline. This increase was significantly greater than that observed in virus negative subjects. A subset of children who had received treatment with omalizumab within 4 weeks before study enrollment, presented significant reduction of acute severity of rhinovirus-triggered asthma exacerbation, reduction in the time to every-2-hours albuterol, and decrease in hospital length of stay ●[32]. In keeping with these observations, high titers of IgE antibody to dust mite allergen were associated with increased risk for acute wheezing provoked by rhinovirus among asthmatic children [33]. Similar effects are likely to occur among children and young adults living in areas where cockroach exposure and sensitization are predominant [29].
Cockroach exposure and sensitization in relation to development of asthma
Early exposure to cockroach may increase the risk of developing asthma among children from atopic families. Results of a birth cohort study revealed that children exposed to levels of Bla g 1 or Bla g 2 of 2 U/g dust or greater had a relative risk for incident asthma of 35.87 that was ~4-fold higher compared to children with exposure to lower levels. Cockroach allergen exposure was also a significant predictor for recurrent wheezing by multivariate analysis [34].
A prospective study carried out in Brazil revealed that 52% of children seen at the Emergency Department for acute wheezing in the first 2 years of life presented three or more wheezing episodes in the past 12 months (persistent wheezers), after 2 years of follow up. Independent risk factors for persistence of wheezing were allergic sensitization and exposure to cockroach allergen in the kitchen [35].
An interaction between exposure to cockroach and environmental pollution has been uncovered. Perzanowski et al. showed that prenatal exposure to cockroach allergen among Dominican and African-American pregnant women in New York City was associated with a greater risk of allergic sensitization at the ages of 5 to 7 years. Risk was increased only among children who were also exposed to nonvolatile polycyclic aromatic hydrocarbons (PAHs), with children null for the Glutathione-S-transferase µ 1 (GSTM1) mutation being particularly vulnerable. PAHs are ubiquitous environmental pollutants produced during the incomplete combustion of organic materials; urban children are exposed to high levels of lower-molecular-weight semivolatile PAHs. The study highlighted the fact that combustion byproducts can act as adjuvants in the development of cockroach sensitization in urban environments [36]. Jung et al. examined prospectively whether PAH exposure at ages 5 and 9–10 years, would be associated with the development of sensitization to cockroach among inner-city children. Repeatedly high levels of PAH metabolites during childhood were associated with new sensitization to cockroach at age 9 years, which is linked to a well-established risk of development of asthma ●●[37].
Additional factors might also play a role in recurrent wheezing early in life. The Urban Environment and Childhood Asthma (URECA) study, a birth cohort which enrolled 560 inner-city children at high risk for asthma, examined the role of environmental factors in early childhood associated with recurrent wheezing. Cumulative exposure to cockroach, mouse, and dust mite over the first 3 years correlated with sensitization to these allergens at age 3 years, and allergen sensitization was positively associated to recurrent wheeze. However, contrary to expectations, exposure to high levels of cockroach, mouse, and cat allergen in the first-year of life showed a strong inverse correlation with recurrent wheeze at age 3 years. A nested case-control study involving 104 children of this cohort, which analyzed the bacterial content of house dust collected in the first year of life, revealed interesting results. The group of children with neither wheeze nor atopy had the highest first-year exposure to allergens and to bacterial species identified as potentially protective against atopic wheeze, particularly members of the Bacteroidetes and Firmicutes phyla. Interestingly, these results suggested that exposure to high levels of allergens in combination with an environment rich in specific bacteria in early childhood might be beneficial, leading to a protective effect against wheeze and atopy [38].
Only few studies report associations between sensitization to specific allergens and disease. In Taiwan, the number of IgE-binding allergens did not correlate with the clinical severity of airway allergy to cockroaches. However, sensitization to Per a 2 correlated with more severe airway allergy and elevated proinflammatory cytokines: 81% of patients with persistent asthma and rhinitis (AS) (and only 45% of patients with allergic rhinitis only) were sensitized to Per a 2. In contrast, this association was inverted for Per a 9: 80% of rhinitis patients and only 28.5% of AS patients were sensitized to this allergen [39]. In agreement with this finding, Bla g 2 (followed by Bla g 5) was found to be a dominant allergen in a group of 118 US patients from which 70% were asthmatic [40]. Table 1 summarizes the reported prevalences of IgE sensitization to cockroach allergens. Additional information, including tropomyosin as a dominant cockroach allergen in Brazil, is provided in the section Recombinant cockroach allergens for diagnosis.
Table 1.
Cockroach allergens listed in the WHO/IUIS Allergen Nomenclature database
| Properties of cockroach allergens | ||||||
|---|---|---|---|---|---|---|
| Allergen | M.W.* | Function/homology | IgE prevalence (%) |
PDB ID | References | |
| Blattella germanica | ||||||
| Bla g 1 | ~25–90 | Midgut microvilli | 20–40 | 4JRB | [24,40,62,64,65] | |
| Bla g 1.0101 | 46, 21 | protein homolog | [65] | |||
| Bla g 1.0102 | 25–37 | [64] | ||||
| Bla g 1.0201 | 56 | [65] | ||||
| Bla g 2 | 36 | Inactive aspartic protease | 40–70 | 1YG9, 2NR6**, 3LIZ**, 4RLD | [24,62,80,81,82, 85]●●[87] | |
| Bla g 3 | 79 | Arylphorin/hemocyanin | 22 | [91] | ||
| Bla g 4 | 21 | Lipocalin | 17–40, 53 | 3EBK*** | [62,92,93] | |
| Bla g 5 | 23 | Glutathione S- Transferase | 35–68 | 4QR5 | [62,69]●●[95] | |
| Bla g 6 | Bla g 6.0101 | 17 | Troponin C | 14 | [96] | |
| Bla g 6.0201 | [96] | |||||
| Bla g 6.0301 | [96] | |||||
| Bla g 7 | 33 | Tropomyosin | 18–31 | [62,97] | ||
| Bla g 8 | --- | Myosin light chain | 14 | [96] | ||
| Bla g 9 | Bla g 9.0101 | 40 | Arginine kinase | 35–50 | [62] | |
| Bla g 11 | 57 | α-Amylase | 41 | [63] | ||
| Periplaneta americana | ||||||
| Per a 1 | ~25–45 | Midgut microvilli | 5–9, 54, 100 | |||
| Per a 1.0101 | 26 | protein homolog | [70] | |||
| Per a 1.0102 | 26 | [71] | ||||
| Per a 1.0103 | 45 | [72] | ||||
| Per a 1.0104 | 31 | [71] | ||||
| Per a 1.0201 | 51 | [71] | ||||
| Per a 2 | 42 | Inactive aspartic protease | 63 | [39] | ||
| Per a 3 | 46–79 | Arylphorin/hemocyanin | 26–95 | [88,90] | ||
| Per a 3.0101 | 79 | [88] | ||||
| Per a 3.0201 | 75 | [88] | ||||
| Per a 3.0202 | 56 | [90] | ||||
| Per a 3.0203 | 46 | [90] | ||||
| Per a 6 | 17 | Troponin C | n.p. | [96] | ||
| Per a 7 | 33 | Tropomyosin | 13–54 | [40,98,99,115] | ||
| Per a 7.0101 | [98] | |||||
| Per a 7.0102 | [99] | |||||
| Per a 9 | 43 | Arginine kinase | 80–100 | [101] | ||
| Per a 10 | 28 | Serine protease | 82 | [107] | ||
| Per a 11 | 55 | α-Amylase | 83 | [111] | ||
| Per a 12 | 45 | Chitinase | 64 | [111] | ||
PDB ID: Protein Data Bank Accession number; n.p.: Not published
Molecular weight calculated from sequence
X-ray crystal structures of Bla g 2 in complex with monoclonal antibodies that inhibit IgE antibody binding
The structure of a P. americana homolog was reported under the PDB ID 3EBW
Mechanisms leading to cockroach allergy
In addition to genetic factors, different mechanisms of innate and adaptive immunity contribute to the development of cockroach allergy [41] (Figure 1). Effects of cockroach extracts on human airway epithelial cells have been described, that include the induction of cytokines and chemokines (IL-8, IL-25, IL-33, CCL20 and GM-CSF) [42–44]. Some effects are mediated by proteases acting on protease-activated receptors (PAR-2) (see also Group 10 section) [45,46]. Additional mechanisms of action have been reported in mouse models. First, German cockroach frass was shown to directly affect neutrophil cytokine production via TLR2, but not TLR4, suggesting an important link between innate and adaptive immunity [47]. Second, activation of the signaling associated with the aryl hydrocarbon receptor (which responds to environmental stimuli and is involved in the pathogenesis of asthma), protected lungs from cockroach-induced inflammation [48]. Third, neonatal mice immunized with α-1,3 glucan developed IgA-secreting B cells that suppressed the development of cockroach allergy [49]. Most of these studies were performed with cockroach extracts that are known to be very variable in content [50].
Figure 1. Proposed mechanisms of cockroach allergy.
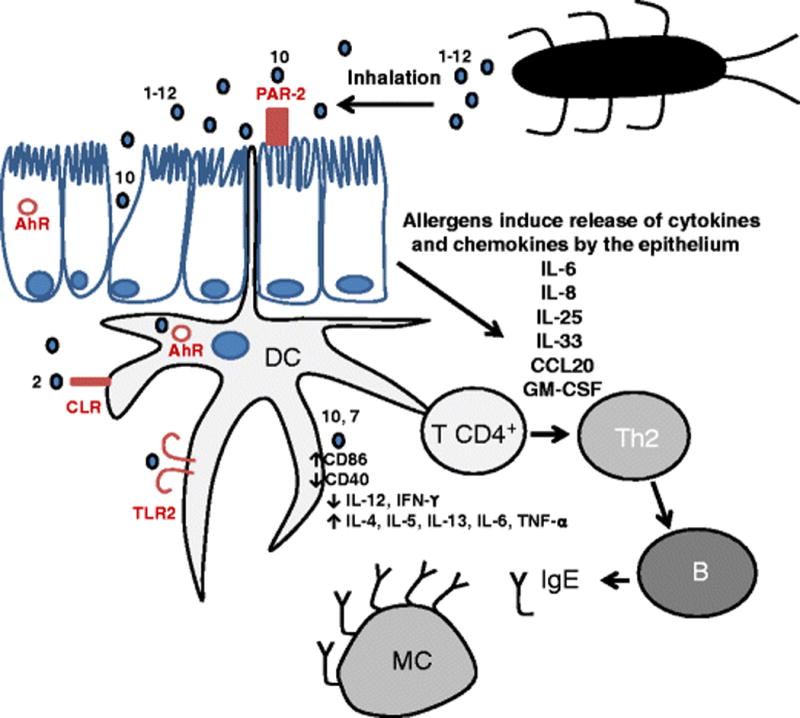
Cockroach allergens, belonging to 12 different groups, are carried by particles that are inhaled to the human lung, where they activate innate and adaptive immune responses. Mechanisms involved in the process include: a) disruption of epithelial integrity by proteases (such as Per a 10) that facilitate allergen penetration, b) activation of release of pro-inflammatory cytokines from the epithelium in a PAR-2 dependent manner by proteases, c) allergen interaction with different receptors (some of which contribute to the uptake of allergens by dendritic cells -TLR, CLR-), and subsequent activation of the adaptive immunity with production of IgE antibodies that bind to the high affinity IgE receptors on mast cells. Numbers indicate the allergen group number. TLR: Toll like receptors, CLR: C-type lectin receptors including mannose receptors, AhR: Aryl hydrocarbon receptor, DC: dendritic cell; T CD4+ and Th2: T cells; B: B cell; MC: mast cell.
In the past 20 years, the identification of twelve groups of cockroach allergens has enabled studies on allergen-specific mechanisms of action. Among these, there is evidence of a role of carbohydrates on the interaction of Bla g 2 with the mannose receptor C-type lectin (CD206) in human circulating fibrocytes. These carbohydrates are predominantly small mannose-terminated glycans with and without fucose and stimulate up-regulation of inflammatory cytokines (TNF-α, IL-6) and activation of signaling molecules such as nuclear factor kappa B (NF-kB) [51]. Effects of group 7 allergens that lead to Th2 polarization include promotion of T cell immunoglobulin mucin domain 4 (TIM4) expression in dendritic cells, down-regulation of toll-like receptor 9 and IL-12 release, induction of T-helper type 2 cytokine release and up-regulation of expression of protease-activated receptors on murine mast cells [52–54].
Cockroach allergens: a heterogeneous group of proteins
Twelve groups of cockroach allergens are currently listed in the official Allergen Nomenclature database maintained by the World Health Organization/International Union of Immunological Societies (WHO/IUIS) Sub-Committee (www.allergen.org). These groups comprise allergens with a wide variety of structures and functions summarized in Table 1. Allergens from groups 1, 2 and 10 are excreted into the feaces, which facilitates environmental exposure, while others are found predominantly in the bodies [24,39,55,56]. In fact, groups 10, 11 and 12 are digestive enzymes [57]. In contrast, Bla g 4 is expressed only in the adult male reproductive system and transferred to the female within the spermatophore [58]. Other allergens have functions associated with muscle contraction (groups 6, 7 and 8) or metabolism (groups 3, 5 and 9), and most of these allergens are expected to be released to the environment upon death of the cockroach. Since reviewed in 2014 [5,59], molecular studies have unveiled new structural features of allergens in groups 1, 2 and 5, and two new groups (11 and 12) have been added to the list. In addition to these officially recognized cockroach allergens, other potential allergens have also been identified that need further study [60–63]. This section will describe the twelve official groups of cockroach allergens with emphasis on new studies performed in the last 5 years.
Group 1: Tandem repeated structures
The group 1 cockroach allergens have been a challenge to characterize. The genetic structure is composed of multiple sequence repeats that are likely the result of multiple gene duplication events, at different times in evolution [64–67]. This confounded early studies to clone the allergen. Adding to the difficulty was the observation that the protein would degrade in a ladder-like pattern when separated on protein gels, with most of rungs binding IgE antibodies [68]. This molecular heterogeneity created a problem in standardizing the allergenic unit, since the size of proteins binding IgE was variable. Many of these obstacles were overcome with the determination of the crystal structure [23]. The structure revealed that the repeated unit contained six helices that assemble with another repeated unit to form a spherical capsule (Figure 2). The allergen structure can be viewed as several of these capsules on a string, tethered by flexible regions susceptible to proteolysis, which results in the ladder-like effect on protein gels. By comparison of the spherical structural unit with natural Bla g 1 by immunoassay, a relative unit could be calibrated to 104 ng of allergen [23]. This equivalence in absolute mass units is useful for standardization of allergen measurements for the assessment of environmental exposure and allergen content in extracts for immunotherapy [50].
Figure 2. Structural analyses of Bla g 1 and Bla g 5.
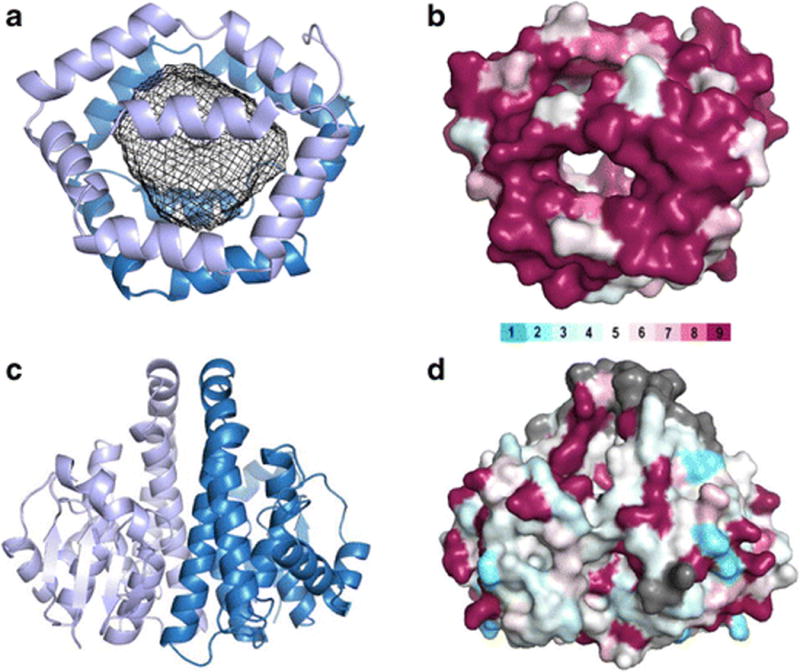
Bla g 1 (4JRB, panels A and B) and Bla g 5 (4QR5, panels C and D) are presented as ribbon diagrams (panels A and C) and surface representations (panels B and D). Bla g 1 in panel A is colored blue and light blue to differentiate the two hemispheres of 6 helices (all from one polypeptide chain) that encapsulate a large hydrophobic cavity, shown with a mesh rendering. Bla g 5 in panel C is colored blue and light blue to show the two polypeptide chains that come together to form a typical GST dimer. In panel B, Bla g 1 is colored based on similarity to Per a 1, and in panel D, Bla g 5 is colored with respect to similarity to Der p 8. The color bar represents residue similarity from low (light blue) to high (maroon) [95]. Gray represents gaps or insertions in the sequence alignment. Visual analysis of the coloring shows the structural basis of cross-reactivity for Bla g 1 and Per a 1 and the lack of significant cross-reactivity between Bla g 5 and Der p 8, which has been experimentally confirmed.
Bla g 1 and the Periplaneta americana homolog Per a 1 share 70–72% sequence identity and show antigenic cross-reactivity [65,70–72]. An anti-Bla g 1 antibody developed at the Food and Drug Administrations (FDA) is species specific, which could be useful in differentiating exposure to the two species [73]. This epitope was mapped by scanning alanine mutagenesis to a region of substantial residue differences between Bla g 1 and Per a 1.
The structure of Bla g 1 revealed a unique protein fold with a large hydrophobic interior cavity. Incomplete electron density suggested that phospholipids were present; this was subsequently confirmed by mass spectrometry and 31P Nuclear Magnetic Resonance (NMR) [23]. However, in Bla g 1 purified from cockroach frass only long chain fatty acids were found: oleate, sterate, and palmitate. This suggested that possibly Bla g 1 was involved in the dietary uptake of phospholipids and the excretion of less nutritive lipids. Indirect support for this hypothesis was obtained from: 1) the observation that cockroaches injected with RNAi to suppress Bla g 1 production effectively starve [74], and 2) the existence of a related protein in the mosquito Anopheles gambiae that stains to microvilli in the gut [75]. Bla g 1 has been localized specifically to the midgut in Blattella germanica [55]. In the mosquito Aedes aegypti, related proteins (19–43% identity) are upregulated in response to a blood meal and not a sugar feeding, further supporting the contention that lipids (or foreign proteins) are important for the expression pattern [76]. Other recent data suggest that the same A. aegypti proteins are up- and down-regulated in response to flavivirus and parasite infections, connecting expression of these proteins to innate immune pathways like Toll, immune deficiency, and Jak/stat in the insect [77,78].
In terms of the relation to allergic disease, more than half of known allergens bind to various hydrophobic ligands [79]. The saturated lipids found with Bla g 1 in cockroach waste have been suggested to stimulate TLR pathways in humans [80]. Possibly, the delivery of these hydrophobic compounds with Bla g 1 or other allergens skews the immune response towards Th2. Deciphering the exact mechanisms of how this happens is an active area of research.
Group 2: Inactive aspartic proteases
Allergens from group 2 are inactive aspartic proteases due to amino acid substitutions and distortions, compared to pepsin, in the area corresponding to the catalytic site [81,82]. Bla g 2, Per a 2 and the homologous protein from Leucophaea maderae are zinc binding proteins and share a 4-residue zinc-binding site [81]. Bla g 2 is one of the most important cockroach allergens in terms of IgE antibody prevalence (42–70%) in temperate areas like the US [24]. In Taiwan, airway allergic patients sensitized to Per a 2 had more severe allergy and elevated proinflammatory chemokines [39]. Secretion of group 2 allergens into the gut has been demonstrated for Bla g 2 by immunoassays of dissected cockroach body parts and tissues, and for Per a 2 by immunohistochemical staining [24,83]. Eventually, allergens from group 2 (and group 1) are excreted to the environment, a fact that, combined with their molecular stability, makes them excellent markers of cockroach allergen exposure. Per a 2, together with Per a 9 and Per a 10, were found to be the most abundant allergens in patient houses in Taiwan [56].
The X-ray crystal structures of Bla g 2 in complex with murine monoclonal antibodies that interfere with IgE antibody binding led to the identification of amino acids involved in allergen-antibody interactions and interesting mechanisms of antibody recognition, including the contribution of glycans from the allergen to antibody binding [84–86]. Recently, a site-directed mutagenesis analysis of B cell epitopes in Bla g 2 led to the design and expression of a triple mutant with reduced capacity to bind IgE that displayed immunomodulatory properties while retaining the native allergen fold ●●[87].
Group 3: Hemolymph homologs
Allergens from group 3 are hexameric proteins homologous to arylphorins or insect storage proteins (20–34%), insect juvenile hormone-suppressible proteins (31–36%) and arthropod hemocyanins (30–35%) [88,89]. Per a 3 comprises different variants or isoallergens, with a wide range of skin test reactivity (26–95%) [88,90]. A B. germanica homolog, Bla g 3, was originally reported in the Genbank (GU086323). Recently, a Bla g 3 protein was immunoprecipitated using a cockroach-specific avian single-chain variable antibody and was recognized as a minor allergen by an US population with a 22% IgE prevalence [91].
Group 4: Insect lipocalins involved in reproduction
Group 4 comprises small extra-cellular allergens that belong to the lipocalin family of proteins. A large group of homologous lipocalins are mammalian allergens (in cow, dog, cat, horse, mouse, rat and guinea pig), with a low degree of amino acid identity with the group 4 cockroach allergens (10–20%). Nevertheless, all have a conserved fold that consists of a C-terminal α-helix and an antiparallel β-barrel with an internal hydrophobic ligand binding cavity [92]. Lipocalins are secreted or excreted molecules with functions associated with their capacity to bind small hydrophobic ligands such as pheromones, retinoids, steroids and arachidonic acid. Bla g 4 is an 18 kDa protein, expressed exclusively in the adult male reproductive system, and involved in reproductive function [58,93]. A structural analysis of native Bla g 4 (combined with nuclear magnetic resonance and isothermal titration calorimetry) revealed that tyramine and octopamine are ligands that might control pheromone production [94].
Group 5: Glutathione S-transferases
Bla g 5 is one of the most prevalent cockroach allergens in the US population [95]. Cockroach group 5 allergens are glutathione S-transferase enzymes (GSTs), similar to mite group 8, and Ascaris group 13. It was recently shown that there is no significant IgE cross-reactivity among Bla g 5, Blo t 8, Der p 8 and Asc s 13 in North American cockroach allergic patients, in agreement with the low amino acid identity at the level of the molecular surface (Figure 2) ●●[69]. This implies that recombinant forms of these allergens will be very useful for accurate molecular diagnoses of the sensitizing species in areas where exposure to multiple species is common.
Bla g 5 belongs to the sigma class of GST while a P. americana homolog (not yet listed in the official WHO/IUIS database) is a delta class GST. The two proteins share 17% identity so it is unlikely there is any cross-reactivity. The genomes and/or transcriptomes of B. germanica and P. americana have recently been determined. A search of the predicted proteomes reveals that P. americana has a sigma class GST with 74% identity to Bla g 5, and reciprocally, B. germanica has a delta class GST with 81% identity to Per a 5 (NCBI Bioproject PRJDB1997 for Per a 5, and NCBI SRA ID SRX682022 and the genome from Baylor College of Medicine for Bla g 5). Future investigations are warranted to examine if these new proteins are allergens and if they could be a source of cross-reactivity.
Groups 6, 7 and 8: Allergen homologs to proteins involved in contraction
Allergens from groups 6 and 8 share homology to troponin C and myosin light chain, respectively. These molecules, by binding calcium to EF-hand motifs, undergo conformational changes that are involved in their function, and affect IgE antibody binding recognition [96]. The group 7 includes tropomyosins such as Bla g 7 and Per a 7, also involved in muscle contraction [97–99]. Tropomyosin is a pan-allergen present in muscle of many animal species. Originally identified as a major shrimp allergen, it is also present in a number of mollusks (i.e. squid), arthropods (arachnids and insects) and parasites, and leads to IgE cross-reactivity [100].
Group 9: Arginine kinases
Group 9 allergens are arginine kinases involved in the metabolism of ATP by catalyzing the reversible transfer of the high energy phosphoryl group from ATP to arginine [62,101,102]. These enzymes were first described as allergens in the Indian meal moth (Plodia interpunctella; Plo i 1) which produces an arginine kinase that cross-reacts with a cockroach homolog and in black tiger shrimp (Penaeus monodon; Pen m 2) [102,103]. Recently, arginine kinase (together with hemocyanin) has been reported to be a cross-reacting allergen between shrimp and arthropods [104]. New evidence supports that group 9 is relevant for cockroach allergy. Per a 9, despite its low stability, is one of the most abundant cockroach allergens in Taiwanese patient’s homes, indicating current status of cockroach control [56]. Intranasal, liposome-adjuvanted allergy vaccines made of Per a 9 resulted in better attenuation of allergic airway inflammation in a mouse model than the equivalent vaccine made with whole-body extract from P. americana [105]. Recently, Bla g 9 has been found to be an immunodominant allergen regarding T cell responses in asthmatic subjects ●[106].
Group 10: Serine proteases
Per a 10 is a 28 kDa serine protease reported in India as a major allergen recognized by 82% (37/45) of cockroach sensitized patients [107]. Recent studies by the same group showed that active Per a 10 induces a dendritic cell type 2 polarization depending on the allergic status of the individual and the protease activity of this allergen [108,109] (Figure 2). This effect was mediated by an upregulation of CD86 and a reduction in CD40 expression on the surface of dendritic cells from cockroach allergic patients, while soluble CD40 levels were enhanced, presumably due to the cleavage of this receptor. This effect was associated with reduced IL-12 and IFN-γ secretion by inhibition of the nuclear factor-kappa B pathway, and with increased secretions of IL-4, IL-5, IL-6 and TNF-α [108]. Per a 10 was also able to activate a human derived epithelial cell line to secrete proinflammatory cytokines (IL-6, IL-8 and GM-CSF) in a protease dependent manner via protease-activated receptor 2 (PAR-2) [110].
Group 11: α-Amylases
Originally identified in German cockroach fecal extracts by a proteomic approach, group 11 comprises α-amylases. These are ubiquitous endoglycosidases that hydrolize α-linked polysaccharides to yield glucose and maltose [111]. Bla g 11 shares the highest identity with α-amylase from pig (55.8%) and mite group 4 allergens (Blo t 4, 50.4%; Der p 4, 49.8%; Eur m 4, 47.4%). However, the cross-reactivity between α-amylases from cockroach and allergenic α-amylases from other species needs to be further investigated. Recently, a homolog in P. americana, Per a 11, that shares 77% amino acid identity with Bla g 11, has also been identified in China [111]. Several indications suggest that group 11 cockroach allergens may play an important role in allergic disease. First, recombinant Bla g 11 inhibited 55% of specific IgE to German cockroach whole-body extract among patients in Korea [63]. Second, the prevalence of IgE reactivity to group 11 allergens is high for certain populations: 1) Bla g 11 was recognized by 41.4% (12/29) of German cockroach sensitized subjects in Korea [63], and 2) a large proportion of Chinese cockroach allergic patients recognized natural Per a 11 purified from midgut of cockroaches using immunoblots (83%; 39/47) and skin prick test (80%; 12/15) [111]. Finally, Bla g 11 was immunodominant regarding T cell responses in asthmatic subjects, together with Bla g 5 and Bla g 9 ●[106].
Group 12: Chitinases
Group 12 cockroach allergens are chitinases. Per a 12 shows 33% amino acid sequence identity to group 15 mite allergens. Although their presence in the gastrointestinal tract of cockroaches has been known for a long time, chitinases have only recently been recognized as cockroach allergens [111,112]. These enzymes hydrolyze the N-acetyl-D-glucosamine 1,4-β-linkages of chitin polymers and are essential for digestion of chitin [63,111]. Chitinases, together with trypsins (group 10) and α-amylases (group 11), are part of the major digestive enzymes in cockroach [57].
Per a 12 was a major allergen in the Chinese population where it was first described, since sera from 63.8% of patients (30/47) reacted to Per a 12 on immunoblots and 60% (9/15) had a positive skin prick test. In addition, ELISA inhibition of serum IgE reactivity to the cockroach midgut extract (CME) by purified Per a 12 was significant (40%) [111].
Recombinant cockroach allergens for diagnosis
In clinical practice, diagnosis of cockroach allergy is performed using crude extracts by in vivo skin testing and/or in vitro measurement of IgE to cockroach. Cockroach extracts available in the US for allergy diagnosis are non-standardized, highly variable in allergen content and show low potency as compared to standardized mite, cat or grass extracts [113,114]. Attempts to standardize cockroach extracts have been hampered by the fact that no single cockroach allergen is immunodominant, in a way that it could be measured as a marker for standardization purposes [22].
Recombinant cockroach allergens have been successfully used for in vivo and in vitro assessment of sensitization [59]. Cockroach allergic patients present variable allergen sensitization profiles, without a major allergen accounting for most of the IgE reactivity to cockroach within a given population. The importance of individual allergens in causing sensitization varies in different areas of the world, possibly due to the influence of sensitization to cross-reactive antigens.
In studies carried out in the US, sensitization to Bla g 2, Bla g 4 and Bla g 5 presented the highest prevalence among cockroach allergic patients with asthma, and a panel of 5 recombinant allergens (Bla g 1, Bla g 2, Bla g 4, Bla g 5, and Per a 7) could identify 64% of cockroach allergic US patients [40]. Using recombinant Bla g 1, Bla g 2, Bla g 4, Bla g 5, Bla g 7, and the newly identified B. germanica enolase, arginine kinase, and vitellogenin, Chuang et al. showed that all cockroach allergic patients reacted to at least one allergen on an IgE dot-blot immunoassay, in a study carried out in Taiwan [62]. The prevalence of IgE recognition was highest for Bla g 2 (63%), followed by Bla g 4 (53%), vitellogenin (47%), Bla g 1 and arginine kinase (34%), Bla g 5 and Bla g 7 (31%), and enolase (25.0%).
In contrast, data from Brazil revealed striking differences from the results observed in US and Taiwan. Using a panel of 5 recombinant allergens (Per a 1, Per a 7, Bla g 2, Bla g 4 and Bla g 5), 24/57 (42%) cockroach allergic patients had positive skin tests to Per a 7, but the reactivity to the other allergens tested was remarkably low (≤ 7%) [59,115]. The high prevalence of IgE reactivity to tropomyosin in Brazil could reflect cross-reactivity to mite tropomyosin, which shares 80% sequence identity to the cockroach homolog. Cross-reactivity with tropomyosin from intestinal parasites, particularly Ascaris lumbricoides, could account for the high frequency of sensitization to tropomyosins in Brazil and Africa [59,116].
As mentioned above, in Taiwan, IgE-binding to Per a 2 was more frequently found among patients with persistent asthma, as compared to patients with rhinitis only (81% versus 45%). The results suggested that sensitization to Per a 2 could be a marker of more severe airway disease [39].
The availability of individual recombinant and natural cockroach allergens will facilitate the diagnosis of individual profiles of IgE reactivity and the identification of specific allergen/s affecting each cockroach allergic patient.
Cockroach Immunotherapy
Cockroach immunotherapy has the potential of modifying the course of asthma and providing sustained clinical benefit. A limited number of clinical trials using cockroach allergen extracts showed improvement in both immunological and clinical parameters [59,117] (Table 2). Kang et al. in 1988 reported in a small number of patients (n = 11) that immunotherapy with cockroach extract resulted in improvement of immunological and clinical parameters after five years of treatment [118]. Another study described decrease in nasal symptoms and increase in cockroach specific IgG levels, accompanied by decrease in serum levels of IL-2, IL-4 and IL-4 receptor, after 3 years of immunotherapy with cockroach extract [119]. A double-blind, placebo controlled cockroach immunotherapy trial from India reported, at one year, a significant improvement in clinical scores and bronchial hyper-reactivity, and increase in cockroach specific IgG4 [120]. These small studies suggested that cockroach immunotherapy could be effective.
Table 2.
Clinical trials of immunotherapy with cockroach extracts for cockroach allergic patients.
| Authors (reference) |
Study design | Study characteristics | Clinical outcomes | Immunological outcomes |
|---|---|---|---|---|
|
| ||||
| Kang et al 1988 USA (118) | “Active treatment”: IT with CR extract plus other allergens to which patients were sensitized; “control”: IT with all relevant allergens except CR | 28 patients with asthma; SCIT with mixed CR extracts (German, American and Oriental) for 5 years. 11/15 and 2/13 “active treatment” and “control” completed the study, respectively | Reduction in symptom and medication scores | Increase in CR- specific blocking antibody and blunting of in vitro basophil histamine release |
|
| ||||
| Alonso et al 1999 Argentina (119) | Open label | SCIT with American CR extract for 3 years | Decrease in nasal symptoms | Increase in CR- specific IgG, decrease in serum IL- 2, IL-4 and IL-4R. |
|
| ||||
| Srivasta et al 2011 India (120) | Double-blind, placebo controlled trial | 42 adult patients with asthma and/or rhinitis (28 active treatment, 18 placebo), SCIT with American CR extract for one year. 12 active treatment and no placebo after 2 years | Improvement in clinical scores and in bronchial hyper-reactivity after one year. Reduction in symptoms and medication use after 2 years | Increase in CR- specific IgG4 after one and two years |
|
| ||||
| Wood et al 2014 USA, ICAC phase I/II studies (121) | ||||
| SCSS | Open label, single site. Designed to study safety of the therapy | 27 patients, with perennial rhinitis with or without asthma (adults, 8- 17 and 5–7 years-old children). SLIT with German CR extract, dose escalation on day 1, maintenance for 14 days. CR allergen dose per day - Bla g 2: 4.2 µg, Bla g 1: 50 µg | Mild to moderate events: oral or throat pruritus, skin rash, nausea. One subject discontinued due to oral symptoms and vomiting. One patient had throat irritation and cough, graded as severe. No SAE. | Not applicable |
|
| ||||
| BioCSI | Double-blind, placebo controlled, multicenter. Designed to determine whether the dose for CR SLIT was immunologically active | 54 adult patients (18–54 years- old), with perennial allergic rhinitis, asthma, or both. SLIT with German CR extract, dose escalation on day 1 (day 2 if needed), maintenance for 6 months. CR allergen dose per day - Bla g 2: 4.2 µg, Bla g 1: 50 µg | Not applicable | Increase in CR- specific IgE levels. No significant increase in CR-specific IgG4 |
|
| ||||
| BioCSI2 | Double-blind, placebo controlled, low and higher dose, multicenter | 89 children (4–17 years-old) SLIT with German CR extract, dose escalation for low dose on day 1 (day 2 if needed); for high dose, over 2 to 4 weeks; maintenance for 3 months. CR allergen doses per day - Low dose: Bla g 2: 4.2 µg, Bla g 1: 50 µg; High dose: Bla g 2: 16.8 µg, Bla g 1: 202 µg | Not applicable | Increase in CR- specific IgE in low and high dose. No increase in CR- specific IgG4 in low dose. Increase in CR- specific IgG and IgG4 in high dose. Increase in FAB activity in the low- dose group. |
|
| ||||
| SCITCO | Open label, single site. Designed to determine safety and as a proof of concept of the immunologic effect of German CR extract | 10 adult patients with perennial allergic rhinitis, asthma, or both. SCIT with German CR extracts, escalation over 11 weeks to maintenance with 0.6 mL of a 1:20 concentration of extract for 15 weeks. | Mild reactions common, not affecting dosing. No SAE | Increase in CR- specific IgE (similar to SLIT); reduction in FAB activity and increase in IgG4 (more vigorous and consistent than with SLIT) |
IT: Immunotherapy; CR: cockroach; SCIT: Subcutaneous immunotherapy; SLIT: Sublingual immunotherapy; SAE: serious adverse event; ICAC: Inner City Asthma Consortium
Wood et al. summarized the results of four pilot clinical trials using sublingual immunotherapy (SLIT) and subcutaneous immunotherapy (SCIT) with German cockroach extract to treat cockroach allergic adults and children ●●[121] (Table 2). Both modalities of cockroach immunotherapy were safe, with no severe reactions seen that could bring concerns of continuation of research on cockroach immunotherapy. SCIT was immunologically more active than SLIT, particularly with regard to IgG4 levels and blocking antibody responses. The adult SLIT study (n = 54) showed a greater increase in cockroach-specific IgE levels between the active and placebo groups and a trend toward increased cockroach-specific IgG4 levels in actively treated subjects, but no evidence of functional blocking antibody response. The pediatric SLIT study (n = 99) revealed significant differences in IgE, IgG, and IgG4 responses between both active groups and the placebo group but no consistent differences between the high- and low-dose groups. The SCIT adult study (n = 10) found that the treatment resulted in significant changes from baseline in cockroach IgE, IgG4, and blocking antibody levels [117]. No further clinical trials conducted with cockroach extracts have been reported.
Lack of standardized extracts and complex patterns of IgE responses to cockroach allergens may represent barriers to obtaining full efficacy of cockroach immunotherapy. Prior to the ICAS immunotherapy trials, an initial analysis of the relative potencies of 3 commercially available German cockroach extracts revealed that these extracts were all of relatively low potency ●●[121]. The first two ICAC trials of SLIT were conducted with a maintenance dose of 0.42 ml daily (approximately 3,685 Bioequivalent allergy units, containing 4.2/50 mcg of Bla g 2/1), and in a third arm of the last trial, this intermediate dose was tested against placebo and a 4-fold higher dose (0.84 ml b.i.d.) [122]. Dose finding studies conducted with standardized extracts for SLIT have led to the identification of optimal maintenance dose for some products, ranging from 12 µg to 128 µg of major allergens per day [123].
Khurana et al. have recently reported the development of a multiple allergen extract potency assay (MAEPA) for simultaneous measurement of identified and unidentified allergens in German cockroach extracts, and for assessing the overall potency of these extracts [114]. The role of MAEPA as a regulatory assay for complex allergen extracts remains to be established, but the study points out that accurate comparisons of extracts are crucial for comparing the results of multiple studies [114].
The use of purified natural or recombinant allergens or of hypoallergenic allergen derivatives has the potential to consistently improve efficacy of cockroach immunotherapy, and may overcome the limitations of using crude cockroach natural extracts. However, this approach has not reached clinical practice yet.
CONCLUSIONS
Cockroach allergy is associated with high morbidity and increased severity of disease among asthmatic patients, particularly children, adolescent and young adults living in low-income inner-city environments. Continued indoor exposure to low levels of cockroach allergens may lead to sensitization and subsequent development of symptoms. Strategies effective at decreasing environmental exposure to cockroach allergens have provided conflicting results in clinical outcomes. Immunotherapy using cockroach crude non-standardized, low potency extracts has been attempted, with promising results but limited reports of outcomes. Both sublingual and subcutaneous cockroach immunotherapy were shown to be safe in adults and children. Progress in molecular cloning and expression of recombinant allergens has led to improved knowledge of the structure and function of cockroach allergens, which is fundamental for developing novel strategies for diagnosis and therapy of cockroach allergic patients. Currently, twelve Groups of well-characterized cockroach allergens from both B. germanica and P. americana are listed with the WHO/IUIS Allergen Nomenclature database. The use of recombinant allergens for diagnostic purposes has been investigated by skin testing or measurements of serum specific IgE. Results have demonstrated heterogeneous IgE reactivity profiles, without a dominant cockroach allergen. Further studies will be necessary to characterize the relative importance of different cockroach allergens in a larger number of patients, which will provide information on cockroach sensitization in relation to allergen exposure, geographic location and genetic background. These studies would help selecting the most appropriate cockroach allergens to be used for diagnosis and therapy in a given area. Ultimately, recombinant allergens could be used in clinical trials for immunotherapy for cockroach allergic patients, particularly those at a higher risk for more severe disease including children and young adults living in inner-city environments.
Acknowledgments
Drs. Chapman and Pomés declare a grant from NIAID and are employed by Indoor Biotechnologies. Dr. Arruda reports grants from São Paulo State Research Funding Agency (FAPESP), grants from Brazilian National Research Council – National Institutes of Science and Technology and Institute of Investigation in Immunology (CNPq–INCT–iii). In addition, Drs. Arruda and Chapman have a United States Patent, number 5,869,288. February 1999 issued.
Part of the research reported in this publication was supported by the National Institute of Allergy and Infectious Diseases of the National Institutes of Health under Award Number R01AI077653 (PI: AP and MDC). The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health. Research carried out in Brazil by LKA has been supported by São Paulo State Research Funding Agency (FAPESP) and Brazilian National Research Council – National Institutes of Science and Technology, Institute of Investigation in Immunology (CNPq–INCT–iii). This study was supported in part by the Intramural Research Program of the National Institute of Environmental Health
Footnotes
Compliance with Ethics Guidelines
Conflict of Interest
Drs. Randall and Mueller declare no conflicts of interest relevant to this manuscript.
Human and Animal Rights and Informed Consent
This article does not contain any studies with human or animal subjects performed by any of the authors.
Contributor Information
Anna Pomés, Research Director, Indoor Biotechnologies, Inc. 700 Harris Street, Charlottesville, VA, USA 22903. Phone: 434 984 2304; Fax 434 984 2709; apomes@inbio.com.
Geoffrey A. Mueller, Genome Integrity and Structural Biology Laboratory, Intramural Program, National Institute of Environmental Health Sciences, National Institutes of Health, 111 T.W. Alexander Drive, MD-MR01, Research Triangle Park, NC, 27709 USA; mueller3@niehs.nih.gov.
Thomas A. Randall, Integrative Bioinformatics, National Institute of Environmental Health Sciences, National Institutes of Health, 111 T.W. Alexander Drive, MD-MR01, Research Triangle Park, NC, 27709 USA; randallta2@niehs.nih.gov
Martin D. Chapman, President, Indoor Biotechnologies, Inc. 700 Harris Street, Charlottesville, VA, USA 22903. Phone: 434 984 2304; Fax 434 984 2709; mdc@inbio.com.
L. Karla Arruda, Department of Medicine, Ribeirao Preto Medical School, University of Sao Paulo. Av. Bandeirantes 3900, Ribeirao Preto, SP, Brazil, 14049-900. Tel.: 55-16-3315 0218; Fax: 55-16-3315 0229; karla@fmrp.usp.br.
References
Selected references published in the last three years (by first author chronological order)
Very important ●●
Important ●
- 1.Bernton HS, Brown H. Insect allergy--Preliminary studies of the cockroach. J Allergy Clin Immunol. 1964;35:506–13. doi: 10.1016/0021-8707(64)90082-6. [DOI] [PubMed] [Google Scholar]
- 2.Kang B, Vellody D, Homburger H, Yunginger JW. Cockroach cause of allergic asthma. Its specificity and immunologic profile. J Allergy Clin Immunol. 1979;63:80–6. doi: 10.1016/0091-6749(79)90196-9. [DOI] [PubMed] [Google Scholar]
- 3.Cornwell PB. The cockroach. London: Hutchinson; 1968. [Google Scholar]
- 4.Pomés A. Cockroach and other inhalant insect allergens. Clin Allergy Immunol. 2008;21:183–200. [PubMed] [Google Scholar]
- 5.Pomés A, Arruda LK. Investigating cockroach allergens: Aiming to improve diagnosis and treatment of cockroach allergic patients. Methods. 2014;66:75–85. doi: 10.1016/j.ymeth.2013.07.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Do DC, Zhao Y, Gao P. Cockroach allergen exposure and risk of asthma. Allergy. 2016;71:463–74. doi: 10.1111/all.12827. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Lombardi C, Savi E, Ridolo E, Passalacqua G, Canonica GW. Is allergic sensitization relevant in severe asthma? Which allergens may be culprit? World Allergy Organ J. 2017;10:2. doi: 10.1186/s40413-016-0138-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8●●.Pongracic JA, et al. Distinguishing characteristics of difficult-to-control asthma in inner-city children and adolescents. J Allergy Clin Immunol. 2016;138:1030–41. doi: 10.1016/j.jaci.2016.06.059. A large population of inner-city children with asthma, 6 to 17 years old, was followed prospectively while receiving guidelines-based management for asthma and rhinitis. FEV1 bronchodilator responsiveness, severity of rhinitis and markers of atopy, particularly total serum IgE levels, mold sensitization, and the total number of allergen sensitizations, were significant factors which distinguished difficult-to control from easy-to-control asthma. The study highlights the importance of phenotyping inner-city children with asthma for identifying those who require high-dose asthma controller therapy. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Milligan KL, Matsui E, Sharma H. Asthma in urban children: epidemiology, environmental risk factors, and the public health domain. Curr Allergy Asthma Rep. 2016;16:33. doi: 10.1007/s11882-016-0609-6. [DOI] [PubMed] [Google Scholar]
- 10.Gergen PJ, Togias A. Inner city asthma. Immunol Allergy Clin North Am. 2015;35:101–14. doi: 10.1016/j.iac.2014.09.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11●●.Zoratti EM, et al. Asthma phenotypes in inner-city children. J Allergy Clin Immunol. 2016;138:1016–29. doi: 10.1016/j.jaci.2016.06.061. Cluster analysis using baseline and longitudinal variables was performed in 616 inner-city children with asthma followed prospectively for a year. Five clusters were identified by indicators of asthma and rhinitis severity, pulmonary physiology, allergy (sensitization and total serum IgE), and allergic inflammation. It was shown that allergy distinguished asthma phenotypes in urban children. Severe asthma often coclustered with highly allergic children. However, a symptomatic phenotype with little allergy or allergic inflammation was also identified. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Rosenstreich DL, et al. The role of cockroach allergy and exposure to cockroach allergen in causing morbidity among inner-city children with asthma. N Engl J Med. 1997;336:1356–63. doi: 10.1056/NEJM199705083361904. [DOI] [PubMed] [Google Scholar]
- 13.Sheehan WJ, Phipatanakul W. Indoor allergen exposure and asthma outcomes. Curr Opin Pediatr. 2016 doi: 10.1097/MOP.0000000000000421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Phipatanakul W, Eggleston PA, Wright EC, Wood RA. Mouse allergen. I. The prevalence of mouse allergen in inner-city homes. The National Cooperative Inner-City Asthma Study. J Allergy Clin Immunol. 2000;106:1070–4. doi: 10.1067/mai.2000.110796. [DOI] [PubMed] [Google Scholar]
- 15.Grant T, et al. Mouse sensitization and exposure are associated with asthma severity in urban children. J Allergy Clin Immunol Pract. 2016 doi: 10.1016/j.jaip.2016.10.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Ahluwalia SK, et al. Mouse allergen is the major allergen of public health relevance in Baltimore City. J Allergy Clin Immunol. 2013;132:830–5. doi: 10.1016/j.jaci.2013.05.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Sheehan WJ, Phipatanakul W. Difficult-to-control asthma: epidemiology and its link with environmental factors. Curr Opin Allergy Clin Immunol. 2015;15:397–401. doi: 10.1097/ACI.0000000000000195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18●●.Liu AH, et al. Pathways through which asthma risk factors contribute to asthma severity in inner-city children. J Allergy Clin Immunol. 2016;138:1042–50. doi: 10.1016/j.jaci.2016.06.060. Based on medical evidence in the published literature, the authors developed a conceptual model to describe how 8 risk-factor domains are linked to asthma severity. Among the domains, allergen sensitization (linked sequentially to allergic inflammation, pulmonary physiology and rhinitis severity), and environmental tobacco smoke exposure had the largest effects on asthma severity, whereas vitamin D, stress and obesity showed no significant association. The model supported the concept that multiple pathways contribute significantly to asthma severity. However, allergen sensitization appears to be the originating domain in the pathophysiologic chain. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Gergen PJ, Teach SJ, Togias A, Busse WW. Reducing exacerbations in the inner city: lessons from the Inner-City Asthma Consortium (ICAC) J Allergy Clin Immunol Pract. 2016;4:22–6. doi: 10.1016/j.jaip.2015.07.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Morgan WJ, et al. Results of a home-based environmental intervention among urban children with asthma. N Engl J Med. 2004;351:1068–80. doi: 10.1056/NEJMoa032097. [DOI] [PubMed] [Google Scholar]
- 21.Rullo VE, Rizzo MC, Arruda LK, Sole D, Naspitz CK. Daycare centers and schools as sources of exposure to mites, cockroach, and endotoxin in the city of Sao Paulo, Brazil. J Allergy Clin Immunol. 2002;110:582–8. doi: 10.1067/mai.2002.127511. [DOI] [PubMed] [Google Scholar]
- 22.Portnoy J, et al. Environmental assessment and exposure reduction of cockroaches: a practice parameter. J Allergy Clin Immunol. 2013;132:802–8. doi: 10.1016/j.jaci.2013.04.061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Mueller GA, et al. The novel structure of the cockroach allergen Bla g 1 has implications for allergenicity and exposure assessment. J Allergy Clin Immunol. 2013;132:1420–6. doi: 10.1016/j.jaci.2013.06.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Arruda LK, et al. Molecular cloning of a major cockroach (Blattella germanica) allergen, Bla g 2. Sequence homology to the aspartic proteases. J Biol Chem. 1995;270:19563–8. doi: 10.1074/jbc.270.33.19563. [DOI] [PubMed] [Google Scholar]
- 25.Sporik R, et al. Mite, cat, and cockroach exposure, allergen sensitisation, and asthma in children: a case-control study of three schools. Thorax. 1999;54:675–80. doi: 10.1136/thx.54.8.675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26●.DiMango E, et al. Individualized household allergen intervention lowers allergen level but not asthma medication use: a randomized controlled trial. J Allergy Clin Immunol Pract. 2016;4:671–9. doi: 10.1016/j.jaip.2016.01.016. Individually targeted environmental intervention leading to decrease of indoor allergen levels has not allowed for decrease necessity of asthma medications in adults and children, compared with usual care, in patients already receiving optimal controller therapy. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27●.Rabito FA, Carlson JC, He H, Werthmann D, Schal C. A single intervention for cockroach control reduces cockroach exposure and asthma morbidity in children. J Allergy Clin Immunol. 2017 doi: 10.1016/j.jaci.2016.10.019. A 12-month randomized trial of a single intervention, consisting of placement of insecticidal bait, resulted in sustained cockroach elimination and was associated with improved asthma outcomes in a group of 102 children aged 5 to 17 years with moderate to severe asthma living in New Orleans. This single intervention could be an alternative to multifaceted interventions currently recommended to improve asthma morbidity. [DOI] [PubMed] [Google Scholar]
- 28●.Wisniewski JA, et al. A comparison of seasonal trends in asthma exacerbations among children from geographic regions with different climates. Allergy Asthma Proc. 2016;37:475–81. doi: 10.2500/aap.2016.37.3994. An interesting analysis showing that seasonal peaks for asthma exacerbations varied among children who lived in geographic locations with different climates in the United States, and were not restricted to the beginning of the school year. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Busse WW, et al. Randomized trial of omalizumab (anti-IgE) for asthma in inner-city children. N Engl J Med. 2011;364:1005–15. doi: 10.1056/NEJMoa1009705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30●.Teach SJ, et al. Preseasonal treatment with either omalizumab or an inhaled corticosteroid boost to prevent fall asthma exacerbations. J Allergy Clin Immunol. 2015;136:1476–85. doi: 10.1016/j.jaci.2015.09.008. A double-blind, double placebo-controlled, multicenter clinical trial conducted among over 400 inner-city asthmatic children aged 6 to 17 years with 1 or more recent exacerbations. Anti-IgE therapy with omalizumab before return to school, added to ongoing guidelines-based care, among inner-city youth reduced fall asthma exacerbations, particularly among those with a recent exacerbation. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31●●.Kantor DB, et al. Rhinovirus and serum IgE are associated with acute asthma exacerbation severity in children. J Allergy Clin Immunol. 2016;138:1467–71. doi: 10.1016/j.jaci.2016.04.044. Relevant study showing interactions of rhinovirus infection, IgE sensitization and severity of asthma exacerbations. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32●.Kantor DB, et al. Omalizumab is associated with reduced acute severity of rhinovirus-triggered asthma exacerbation. Am J Respir Crit Care Med. 2016;194:1552–5. doi: 10.1164/rccm.201606-1145LE. Decreasing IgE response with omalizumab reduced severity of rhinovirus-triggered asthma exacerbation, providing a proof-of-concept of the relevance of this association. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Soto-Quiros M, et al. High titers of IgE antibody to dust mite allergen and risk for wheezing among asthmatic children infected with rhinovirus. J Allergy Clin Immunol. 2012;129:1499–505. doi: 10.1016/j.jaci.2012.03.040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Litonjua AA, Carey VJ, Burge HA, Weiss ST, Gold DR. Exposure to cockroach allergen in the home is associated with incident doctor-diagnosed asthma and recurrent wheezing. J Allergy Clin Immunol. 2001;107:41–7. doi: 10.1067/mai.2001.111143. [DOI] [PubMed] [Google Scholar]
- 35.Silva JM, et al. A prospective study of wheezing in young children: the independent effects of cockroach exposure, breast-feeding and allergic sensitization. Pediatr Allergy Immunol. 2005;16:393–401. doi: 10.1111/j.1399-3038.2005.00308.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Perzanowski MS, et al. Early-life cockroach allergen and polycyclic aromatic hydrocarbon exposures predict cockroach sensitization among inner-city children. J Allergy Clin Immunol. 2013;131:886–93. doi: 10.1016/j.jaci.2012.12.666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37●●.Jung KH, et al. Repeatedly high polycyclic aromatic hydrocarbon exposure and cockroach sensitization among inner-city children. Environ Res. 2015;140:649–56. doi: 10.1016/j.envres.2015.05.027. An interesting study uncovering association of inner-city pollution to development of cockroach sensitization. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Lynch SV, et al. Effects of early-life exposure to allergens and bacteria on recurrent wheeze and atopy in urban children. J Allergy Clin Immunol. 2014;134:593–601. doi: 10.1016/j.jaci.2014.04.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Lee MF, Song PP, Hwang GY, Lin SJ, Chen YH. Sensitization to Per a 2 of the American cockroach correlates with more clinical severity among airway allergic patients in Taiwan. Ann Allergy Asthma Immunol. 2012;108:243–8. doi: 10.1016/j.anai.2012.01.014. [DOI] [PubMed] [Google Scholar]
- 40.Satinover SM, et al. Specific IgE and IgG antibody-binding patterns to recombinant cockroach allergens. J Allergy Clin Immunol. 2005;115:803–9. doi: 10.1016/j.jaci.2005.01.018. [DOI] [PubMed] [Google Scholar]
- 41.Gao P, et al. CD14, a key candidate gene associated with a specific immune response to cockroach. Clin Exp Allergy. 2010;40:1353–64. doi: 10.1111/j.1365-2222.2010.03561.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Kouzaki H, Tojima I, Kita H, Shimizu T. Transcription of interleukin-25 and extracellular release of the protein is regulated by allergen proteases in airway epithelial cells. Am J Respir Cell Mol Biol. 2013;49:741–50. doi: 10.1165/rcmb.2012-0304OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Kouzaki H, Iijima K, Kobayashi T, O’Grady SM, Kita H. The danger signal, extracellular ATP, is a sensor for an airborne allergen and triggers IL-33 release and innate Th2-type responses. J Immunol. 2011;186:4375–87. doi: 10.4049/jimmunol.1003020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Gandhi VD, Vliagoftis H. Airway epithelium interactions with aeroallergens: role of secreted cytokines and chemokines in innate immunity. Front Immunol. 2015;6:147. doi: 10.3389/fimmu.2015.00147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Hong JH, et al. German cockroach extract activates protease-activated receptor 2 in human airway epithelial cells. J Allergy Clin Immunol. 2004;113:315–9. doi: 10.1016/j.jaci.2003.11.026. [DOI] [PubMed] [Google Scholar]
- 46.Page K, Hughes VS, Odoms KK, Dunsmore KE, Hershenson MB. German cockroach proteases regulate interleukin-8 expression via nuclear factor for interleukin-6 in human bronchial epithelial cells. Am J Respir Cell Mol Biol. 2005;32:225–31. doi: 10.1165/rcmb.2004-0231OC. [DOI] [PubMed] [Google Scholar]
- 47.Page K, et al. TLR2-mediated activation of neutrophils in response to German cockroach frass. J Immunol. 2008;180:6317–24. doi: 10.4049/jimmunol.180.9.6317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Xu T, et al. Aryl hydrocarbon receptor protects lungs from cockroach allergen-induced inflammation by modulating mesenchymal stem cells. J Immunol. 2015;195:5539–50. doi: 10.4049/jimmunol.1501198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Patel PS, King RG, Kearney JF. Pulmonary alpha-1,3-Glucan-Specific IgA-Secreting B Cells Suppress the Development of Cockroach Allergy. J Immunol. 2016;197:3175–87. doi: 10.4049/jimmunol.1601039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Patterson ML, Slater JE. Characterization and comparison of commercially available German and American cockroach allergen extracts. Clin Exp Allergy. 2002;32:721–7. doi: 10.1046/j.1365-2222.2002.01397.x. [DOI] [PubMed] [Google Scholar]
- 51.Tsai YM, et al. Functional interaction of cockroach allergens and mannose receptor (CD206) in human circulating fibrocytes. PLoS ONE. 2013;8:e64105. doi: 10.1371/journal.pone.0064105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Xu L, et al. Cockroach allergen Bla g 7 promotes TIM4 expression in dendritic cells leading to Th2 polarization. Mediators Inflamm. 2013;2013:983149. doi: 10.1155/2013/983149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Zhang Z, et al. Induction of T-helper type 2 cytokine release and up-regulated expression of protease-activated receptors on mast cells by recombinant American cockroach allergen Per a 7. Clin Exp Allergy. 2008;38:1160–7. doi: 10.1111/j.1365-2222.2008.02991.x. [DOI] [PubMed] [Google Scholar]
- 54.Yang H, et al. Cockroach allergen Per a 7 down-regulates expression of Toll-like receptor 9 and IL-12 release from P815 cells through PI3K and MAPK signaling pathways. Cell Physiol Biochem. 2012;29:561–70. doi: 10.1159/000338510. [DOI] [PubMed] [Google Scholar]
- 55.Gore JC, Schal C. Gene expression tissue distribution of the major human allergen Bla g 1 in the German cockroach Blattella germanica L. (Dictyoptera: Blattellidae) J Med Entomol. 2004;41:953–60. doi: 10.1603/0022-2585-41.5.953. [DOI] [PubMed] [Google Scholar]
- 56.Lee MF, Chen YH, Chiang CH, Lin SJ, Song PP. Analysis of 10 environmental allergen components of the American cockroach in Taiwan. Ann Allergy Asthma Immunol. 2016;117:535–41. doi: 10.1016/j.anai.2016.09.432. [DOI] [PubMed] [Google Scholar]
- 57.Tamaki FK, et al. Physiology of digestion and the molecular characterization of the major digestive enzymes from Periplaneta americana. J Insect Physiol. 2014;70:22–35. doi: 10.1016/j.jinsphys.2014.08.007. [DOI] [PubMed] [Google Scholar]
- 58.Fan Y, et al. Tissue localization and regulation by juvenile hormone of human allergen Bla g 4 from the German cockroach, Blattella germanica (L.) Insect Mol Biol. 2005;14:45–53. doi: 10.1111/j.1365-2583.2004.00530.x. [DOI] [PubMed] [Google Scholar]
- 59.Arruda LK, et al. Recombinant allergens for diagnosis of cockroach allergy. Curr Allergy Asthma Rep. 2014;14:428. doi: 10.1007/s11882-014-0428-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Khantisitthiporn O, et al. Native troponin-T of the American cockroach (CR), Periplaneta americana, binds to IgE in sera of CR allergic Thais. Asian Pac J Allergy Immunol. 2007;25:189–97. [PubMed] [Google Scholar]
- 61.Jeong KY, Son M, Lee JH, Hong CS, Park JW. Allergenic characterization of a novel allergen, homologous to chymotrypsin, from German cockroach. Allergy Asthma Immunol Res. 2015;7:283–9. doi: 10.4168/aair.2015.7.3.283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Chuang JG, Su SN, Chiang BL, Lee HJ, Chow LP. Proteome mining for novel IgE-binding proteins from the German cockroach (Blattella germanica) and allergen profiling of patients. Proteomics. 2010;10:3854–67. doi: 10.1002/pmic.201000348. [DOI] [PubMed] [Google Scholar]
- 63.Jeong KY, et al. Identification of novel allergenic components from German cockroach fecal extract by a proteomic approach. Int Arch Allergy Immunol. 2013;161:315–24. doi: 10.1159/000347034. [DOI] [PubMed] [Google Scholar]
- 64.Helm R, et al. Isolation and characterization of a clone encoding a major allergen (Bla g Bd90K) involved in IgE-mediated cockroach hypersensitivity. J Allergy Clin Immunol. 1996;98:172–80. doi: 10.1016/s0091-6749(96)70240-3. [DOI] [PubMed] [Google Scholar]
- 65.Pomés A, et al. Novel allergen structures with tandem amino acid repeats derived from German and American cockroach. J Biol Chem. 1998;273:30801–7. doi: 10.1074/jbc.273.46.30801. [DOI] [PubMed] [Google Scholar]
- 66.Fischer HM, Wheat CW, Heckel DG, Vogel H. Evolutionary origins of a novel host plant detoxification gene in butterflies. Mol Biol Evol. 2008;25:809–20. doi: 10.1093/molbev/msn014. [DOI] [PubMed] [Google Scholar]
- 67.Randall TA, Perera L, London RE, Mueller GA. Genomic, RNAseq, and molecular modeling evidence suggests that the major allergen domain in insects evolved from a homodimeric origin. Genome Biol Evol. 2013;5:2344–58. doi: 10.1093/gbe/evt182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Pomés A, Vailes LD, Helm RM, Chapman MD. IgE reactivity of tandem repeats derived from cockroach allergen, Bla g 1. Eur J Biochem. 2002;269:3086–92. doi: 10.1046/j.1432-1033.2002.02990.x. [DOI] [PubMed] [Google Scholar]
- 69●●.Mueller GA, et al. Analysis of glutathione S-transferase allergen cross-reactivity in a North American population: Relevance for molecular diagnosis. J Allergy Clin Immunol. 2015;136:1369–77. doi: 10.1016/j.jaci.2015.03.015. A comparative analysis of the X-ray crystal structures of the glutathione S-transferase Bla g 5 and the homolog allergens from mite (Der p 8 and Blo t 8) and Ascaris (Asp s 13), revealed the structural basis for a lack of significant cross-reactivity among these allergens in a North American population, which has implications for their use in allergy diagnosis. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Melén E, Pomés A, Vailes LD, Arruda LK, Chapman MD. Molecular cloning of Per a 1 and definition of the cross-reactive Group 1 cockroach allergens. J Allergy Clin Immunol. 1999;103:859–64. doi: 10.1016/s0091-6749(99)70430-6. [DOI] [PubMed] [Google Scholar]
- 71.Wu CH, Wang NM, Lee MF, Kao CY, Luo SF. Cloning of the American cockroach Cr-PII allergens: evidence for the existence of cross-reactive allergens between species. J Allergy Clin Immunol. 1998;101:832–40. doi: 10.1016/S0091-6749(98)70312-4. [DOI] [PubMed] [Google Scholar]
- 72.Yang CY, Wu JD, Wu CH. Sequence analysis of the first complete cDNA clone encoding an American cockroach Per a 1 allergen. Biochim Biophys Acta. 2000;1517:153–8. doi: 10.1016/s0167-4781(00)00235-9. [DOI] [PubMed] [Google Scholar]
- 73.Mueller GA, et al. Characterization of an anti-Bla g 1 scFv: epitope mapping and cross-reactivity. Mol Immunol. 2014;59:200–7. doi: 10.1016/j.molimm.2014.02.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Suazo A, Gore C, Schal C. RNA interference-mediated knock-down of Bla g 1 in the German cockroach, Blattella germanica L., implicates this allergen-encoding gene in digestion and nutrient absorption. Insect Mol Biol. 2009;18:727–36. doi: 10.1111/j.1365-2583.2009.00912.x. [DOI] [PubMed] [Google Scholar]
- 75.Nolan T, et al. Analysis of two novel midgut-specific promoters driving transgene expression in Anopheles stephensi mosquitoes. PLoS ONE. 2011;6:e16471. doi: 10.1371/journal.pone.0016471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Shao L, Devenport M, Fujioka H, Ghosh A, Jacobs-Lorena M. Identification and characterization of a novel peritrophic matrix protein, Ae-Aper50, and the microvillar membrane protein, AEG12, from the mosquito, Aedes aegypti. Insect Biochem Mol Biol. 2005;35:947–59. doi: 10.1016/j.ibmb.2005.03.012. [DOI] [PubMed] [Google Scholar]
- 77.Colpitts TM, et al. Alterations in the Aedes aegypti transcriptome during infection with West Nile, dengue and yellow fever viruses. PLoS Pathog. 2011;7:e1002189. doi: 10.1371/journal.ppat.1002189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Juneja P, et al. Exome and transcriptome sequencing of Aedes aegypti identifies a locus that confers resistance to Brugia malayi and alters the immune response. PLoS Pathog. 2015;11:e1004765. doi: 10.1371/journal.ppat.1004765. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Karp CL. Guilt by intimate association: what makes an allergen an allergen? J Allergy Clin Immunol. 2010;125:955–60. doi: 10.1016/j.jaci.2010.03.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Huang S, et al. Saturated fatty acids activate TLR-mediated proinflammatory signaling pathways. J Lipid Res. 2012;53:2002–13. doi: 10.1194/jlr.D029546. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Gustchina A, et al. Crystal structure of cockroach allergen Bla g 2, an unusual zinc binding aspartic protease with a novel mode of self-inhibition. J Mol Biol. 2005;348:433–44. doi: 10.1016/j.jmb.2005.02.062. [DOI] [PubMed] [Google Scholar]
- 82.Wünschmann S, Gustchina A, Chapman MD, Pomés A. Cockroach allergen Bla g 2: an unusual aspartic proteinase. J Allergy Clin Immunol. 2005;116:140–5. doi: 10.1016/j.jaci.2005.04.024. [DOI] [PubMed] [Google Scholar]
- 83.Lee MF, et al. IgE-binding epitope mapping and tissue localization of the major American cockroach allergen Per a 2. Allergy Asthma Immunol Res. 2015;7:376–83. doi: 10.4168/aair.2015.7.4.376. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Li M, et al. Crystal structure of a dimerized cockroach allergen Bla g 2 complexed with a monoclonal antibody. J Biol Chem. 2008;283:22806–14. doi: 10.1074/jbc.M800937200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Li M, et al. Carbohydrates contribute to the interactions between cockroach allergen Bla g 2 and a monoclonal antibody. J Immunol. 2011;186:333–40. doi: 10.4049/jimmunol.1002318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Glesner J, et al. Mechanisms of allergen-antibody interaction of cockroach allergen Bla g 2 with monoclonal antibodies that inhibit IgE antibody binding. PLoS ONE. 2011;6:e22223. doi: 10.1371/journal.pone.0022223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87●●.Woodfolk JA, et al. Antigenic determinants of the bilobal cockroach allergen Bla g 2. J Biol Chem. 2016;291:2288–301. doi: 10.1074/jbc.M115.702324. This study generated a Bla g 2 triple mutant with reduced capacity to bind IgE (while preserving the fold of the native allergen) and with T cell modulatory properties, which could potentially be used for immunotherapy. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Wu CH, Lee MF, Liao SC, Luo SF. Sequencing analysis of cDNA clones encoding the American cockroach Cr-PI allergens. Homology with insect hemolymph proteins. J Biol Chem. 1996;271:17937–43. doi: 10.1074/jbc.271.30.17937. [DOI] [PubMed] [Google Scholar]
- 89.Mindykowski B, et al. Cockroach allergens Per a 3 are oligomers. Dev Comp Immunol. 2010;34:722–33. doi: 10.1016/j.dci.2010.01.011. [DOI] [PubMed] [Google Scholar]
- 90.Wu CH, Lee MF, Wang NM, Luo SF. Sequencing and immunochemical characterization of the American cockroach Per a 3 (Cr-PI) isoallergenic variants. Mol Immunol. 1997;34:1–8. doi: 10.1016/s0161-5890(97)00009-6. [DOI] [PubMed] [Google Scholar]
- 91.Khurana T, Collison M, Chew FT, Slater JE. Bla g 3: a novel allergen of German cockroach identified using cockroach-specific avian single-chain variable fragment antibody. Ann Allergy Asthma Immunol. 2014;112:140–5. doi: 10.1016/j.anai.2013.11.007. [DOI] [PubMed] [Google Scholar]
- 92.Tan YW, et al. Structures of two major allergens, Bla g 4 and Per a 4, from cockroaches and their IgE binding epitopes. J Biol Chem. 2009;284:3148–57. doi: 10.1074/jbc.M807209200. [DOI] [PubMed] [Google Scholar]
- 93.Arruda LK, Vailes LD, Hayden ML, Benjamin DC, Chapman MD. Cloning of cockroach allergen, Bla g 4, identifies ligand binding proteins (or calycins) as a cause of IgE antibody responses. J Biol Chem. 1995;270:31196–201. doi: 10.1074/jbc.270.52.31196. [DOI] [PubMed] [Google Scholar]
- 94.Offermann LR, et al. The major cockroach allergen Bla g 4 binds tyramine and octopamine. Mol Immunol. 2014;60:86–94. doi: 10.1016/j.molimm.2014.03.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Arruda LK, Vailes LD, Platts-Mills TA, Hayden ML, Chapman MD. Induction of IgE antibody responses by glutathione S-transferase from the German cockroach (Blattella germanica) J Biol Chem. 1997;272:20907–12. doi: 10.1074/jbc.272.33.20907. [DOI] [PubMed] [Google Scholar]
- 96.Hindley J, et al. Bla g 6: a troponin C allergen from Blattella germanica with IgE binding calcium dependence. J Allergy Clin Immunol. 2006;117:1389–95. doi: 10.1016/j.jaci.2006.02.017. [DOI] [PubMed] [Google Scholar]
- 97.Jeong KY, et al. Allergenicity of recombinant Bla g 7, German cockroach tropomyosin. Allergy. 2003;58:1059–63. doi: 10.1034/j.1398-9995.2003.00167.x. [DOI] [PubMed] [Google Scholar]
- 98.Asturias JA, et al. Molecular characterization of American cockroach tropomyosin (Periplaneta americana allergen 7), a cross-reactive allergen. J Immunol. 1999;162:4342–8. [PubMed] [Google Scholar]
- 99.Santos AB, et al. Cockroach allergens and asthma in Brazil: identification of tropomyosin as a major allergen with potential cross-reactivity with mite and shrimp allergens. J Allergy Clin Immunol. 1999;104:329–37. doi: 10.1016/s0091-6749(99)70375-1. [DOI] [PubMed] [Google Scholar]
- 100.Ayuso R, Reese G, Leong-Kee S, Plante M, Lehrer SB. Molecular basis of arthropod cross-reactivity: IgE-binding cross-reactive epitopes of shrimp, house dust mite and cockroach tropomyosins. Int Arch Allergy Immunol. 2002;129:38–48. doi: 10.1159/000065172. [DOI] [PubMed] [Google Scholar]
- 101.Sookrung N, et al. Periplaneta americana arginine kinase as a major cockroach allergen among Thai patients with major cockroach allergies. Environ Health Perspect. 2006;114:875–80. doi: 10.1289/ehp.8650. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Yu CJ, Lin YF, Chiang BL, Chow LP. Proteomics and immunological analysis of a novel shrimp allergen, Pen m 2. J Immunol. 2003;170:445–53. doi: 10.4049/jimmunol.170.1.445. [DOI] [PubMed] [Google Scholar]
- 103.Binder M, et al. Molecular and immunological characterization of arginine kinase from the Indianmeal moth, Plodia interpunctella, a novel cross-reactive invertebrate pan-allergen. J Immunol. 2001;167:5470–7. doi: 10.4049/jimmunol.167.9.5470. [DOI] [PubMed] [Google Scholar]
- 104.Pascal M, et al. Molecular diagnosis of shrimp allergy: efficiency of several allergens to predict clinical reactivity. J Allergy Clin Immunol Pract. 2015;3:521–9. doi: 10.1016/j.jaip.2015.02.001. [DOI] [PubMed] [Google Scholar]
- 105.Meechan P, et al. Intranasal, liposome-adjuvanted cockroach allergy vaccines made of refined major allergen and whole-body extract of Periplaneta americana. Int Arch Allergy Immunol. 2013;161:351–62. doi: 10.1159/000348314. [DOI] [PubMed] [Google Scholar]
- 106●.Dillon MB, et al. Different Bla-g T cell antigens dominate responses in asthma versus rhinitis subjects. Clin Exp Allergy. 2015;45:1856–67. doi: 10.1111/cea.12643. An analysis of German cockroach antigens involved in T cell responses revealed differences in Bla-g T cell antigens dominating responses in asthma (Bla g 5, Bla g 9 and Bla g 11) versus rhinitis subjects (Bla g 5, Bla g 4). Bla g 2, which is a dominant target of IgE response, accounted for a minor fraction of T cell reactivity. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Sudha VT, Arora N, Gaur SN, Pasha S, Singh BP. Identification of a serine protease as a major allergen (Per a 10) of Periplaneta americana. Allergy. 2008;63:768–76. doi: 10.1111/j.1398-9995.2007.01602.x. [DOI] [PubMed] [Google Scholar]
- 108.Goel C, Kalra N, Dwarakanath BS, Gaur SN, Arora N. Per a 10 protease activity modulates CD40 expression on dendritic cell surface by nuclear factor-kappaB pathway. Clin Exp Immunol. 2015;180:341–51. doi: 10.1111/cei.12569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Goel C, Gaur SN, Bhati G, Arora N. DC type 2 polarization depends on both the allergic status of the individual and protease activity of Per a 10. Immunobiology. 2015;220:1113–21. doi: 10.1016/j.imbio.2015.05.010. [DOI] [PubMed] [Google Scholar]
- 110.Kale SL, Arora N. Per a 10 activates human derived epithelial cell line in a protease dependent manner via PAR-2. Immunobiology. 2015;220:525–32. doi: 10.1016/j.imbio.2014.10.018. [DOI] [PubMed] [Google Scholar]
- 111.Fang Y, et al. Two new types of allergens from the cockroach, Periplaneta americana. Allergy. 2015;70:1674–8. doi: 10.1111/all.12766. [DOI] [PubMed] [Google Scholar]
- 112.Powning RF, Irzykiewicz H. A chitinase from the gut of the cockroach Periplaneta americana. Nature. 1963;200:1128. doi: 10.1038/2001128a0. [DOI] [PubMed] [Google Scholar]
- 113.Slater JE, et al. Biological potency of German cockroach allergen extracts determined in an inner city population. Clin Exp Allergy. 2007;37:1033–9. doi: 10.1111/j.1365-2222.2007.02751.x. [DOI] [PubMed] [Google Scholar]
- 114.Khurana T, Dobrovolskaia E, Shartouny JR, Slater JE. Multiplex assay for protein profiling and potency measurement of German cockroach allergen extracts. PLoS ONE. 2015;10:e0140225. doi: 10.1371/journal.pone.0140225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Barbosa MC, et al. Efficacy of recombinant allergens for diagnosis of cockroach allergy in patients with asthma and/or rhinitis. Int Arch Allergy Immunol. 2013;161:213–9. doi: 10.1159/000346318. [DOI] [PubMed] [Google Scholar]
- 116.Santos AB, et al. Cross-reactive IgE antibody responses to tropomyosins from Ascaris lumbricoides and cockroach. J Allergy Clin Immunol. 2008;121:1040–6. doi: 10.1016/j.jaci.2007.12.1147. [DOI] [PubMed] [Google Scholar]
- 117.Bassirpour G, Zoratti E. Cockroach allergy and allergen-specific immunotherapy in asthma: potential and pitfalls. Curr Opin Allergy Clin Immunol. 2014;14:535–41. doi: 10.1097/ACI.0000000000000106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Kang BC, Johnson J, Morgan C, Chang JL. The role of immunotherapy in cockroach asthma. J Asthma. 1988;25:205–18. doi: 10.3109/02770908809071367. [DOI] [PubMed] [Google Scholar]
- 119.Alonso A, et al. Immunological changes during cockroach immunotherapy. J Investig Allergol Clin Immunol. 1999;9:299–304. [PubMed] [Google Scholar]
- 120.Srivastava D, Gaur SN, Arora N, Singh BP. Clinico-immunological changes post-immunotherapy with Periplaneta americana. Eur J Clin Invest. 2011;41:879–88. doi: 10.1111/j.1365-2362.2011.02480.x. [DOI] [PubMed] [Google Scholar]
- 121●●.Wood RA, et al. Development of cockroach immunotherapy by the Inner-City Asthma Consortium. J Allergy Clin Immunol. 2014;133:846–52. doi: 10.1016/j.jaci.2013.08.047. A detailed description of the ICAC first stage studies of developing immunotherapy for cockroach allergy. Analysis of safety and of immunological biomarkers of three pilot studies of sublingual immunotherapy (SLIT) and one of subcutaneous immunotherapy (SCIT) with German cockroach extract are reported, which revealed that SCIT resulted in more vigorous and consistent immunological responses while using the same extract, as compared to SLIT. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Larenas-Linnemann D. Allergen immunotherapy: an update on protocols of administration. Curr Opin Allergy Clin Immunol. 2015;15:556–67. doi: 10.1097/ACI.0000000000000220. [DOI] [PubMed] [Google Scholar]
- 123.Passalacqua G, Canonica GW, Bagnasco D. Benefit of SLIT and SCIT for allergic rhinitis and asthma. Curr Allergy Asthma Rep. 2016;16:88. doi: 10.1007/s11882-016-0666-x. [DOI] [PubMed] [Google Scholar]