

Abstract
Significance: Oxidative stress is thought to account for aberrant redox homeostasis and contribute to aging and disease. However, more often than not, administration of antioxidants is ineffective, suggesting that our current understanding of the underlying regulatory processes is incomplete.
Recent Advances: Similar to reactive oxygen species and reactive nitrogen species, reactive sulfur species are now emerging as important signaling molecules, targeting regulatory cysteine redox switches in proteins, affecting gene regulation, ion transport, intermediary metabolism, and mitochondrial function. To rationalize the complexity of chemical interactions of reactive species with themselves and their targets and help define their role in systemic metabolic control, we here introduce a novel integrative concept defined as the reactive species interactome (RSI). The RSI is a primeval multilevel redox regulatory system whose architecture, together with the physicochemical characteristics of its constituents, allows efficient sensing and rapid adaptation to environmental changes and various other stressors to enhance fitness and resilience at the local and whole-organism level.
Critical Issues: To better characterize the RSI-related processes that determine fluxes through specific pathways and enable integration, it is necessary to disentangle the chemical biology and activity of reactive species (including precursors and reaction products), their targets, communication systems, and effects on cellular, organ, and whole-organism bioenergetics using system-level/network analyses.
Future Directions: Understanding the mechanisms through which the RSI operates will enable a better appreciation of the possibilities to modulate the entire biological system; moreover, unveiling molecular signatures that characterize specific environmental challenges or other forms of stress will provide new prevention/intervention opportunities for personalized medicine. Antioxid. Redox Signal. 27, 684–712.
Keywords: : hydrogen sulfide, polysulfides, nitric oxide, systems biology, microbiome, network medicine
Introduction
Nothing in biology makes sense except in the light of evolution.
—Theodosius Dobzhansky
Life is nothing but an electron looking for a place to rest.
—Albert Szent-Györgyi
We are witnessing an unprecedented paradigmatic change in the practice of medicine whereby the concept of intervention is evolving from treating diseases in a one-organ/one-symptom manner to a systems-based approach that considers a patient's pathophysiological condition, including his/her individual genetic blueprint, microbiome, disease history, and lifestyle. Indeed, diseases presenting with similar clinical phenotypes are often heterogeneous conditions of multifactorial origin, involving a multitude of molecular, cellular, and organ systems. Their multilevel nature and complexity pose a formidable challenge to identifying the molecular causes; finding the most suitable therapy for each specific case demands a thorough understanding of the fundamental principles of biological regulation and a refined interdisciplinary systems approach encompassing medicine, pharmacology, biology, chemistry, and physics. Recent analyses indicate the existence of disease-specific functional modules that are central hubs in the vast network of human diseases, offering additional opportunities by embracing mathematical approaches.
A limited number of risk factors (such as poor quality nutrition or physical inactivity) and chronic conditions (including hypertension, cardiovascular disease, obesity, asthma, diabetes, neurodegenerative diseases, and certain forms of cancer) account for the majority of the global burden of disease (114), overall life expectancy, and all-cause mortality (203). A common feature of many of these conditions is oxidative stress (176), and some have been redefined as redox diseases (26, 206). The term oxidative stress was originally described as an imbalance between oxidants and antioxidants in favor of the oxidants, leading to a disruption of redox signaling and control and/or molecular damage (91, 174); it was initially considered to be triggered by an inflammatory process or mitochondrial dysfunction. However, the use of selective antioxidants for redox diseases has not had the effect anticipated, suggesting that our current understanding of the underlying pathophysiological processes is incomplete (26, 176).
Recently, Jones and Sies proposed that besides the Genetic Code, allowing reproduction and defining heredity, there exists a Redox Codei that identifies the regulatory elements and defines the principles through which biological function is enabled and protected (94). Within this concept, the endogenous production of reactive oxygen species (ROS) is a highly regulated enzymatic process, which serves the purpose of signaling and can lead to modification of cysteine redox switches. Modification of these switches leads to modulation of their functional state, which would result in alterations of protein structure, enzymatic activity, or gene transcription. Modifying responses to match a changed environment creates the opportunity for adaptive changes that enhance an organism's fitness for purpose.
However, there is more to this redox network than ROS. Nitric oxide (NO) is a free radical, which is produced endogenously by NO synthases (NOS) and acts as an effector and messenger, regulating a variety of physiological processes. Chemical interactions of NO with ROS form reactive nitrogen species (RNS) and constitute the basis for the formation of a multitude of additional oxidative signaling elements (65), including the highly reactive and potentially damaging peroxynitrite (ONOO−). Both ROS and RNS may target cysteine thiols leading to oxidative modifications (including formation of sulfinic acid, sulfenic acids, thiyl radicals, and sulfane sulfur-containing molecules, such as persulfides and polysulfides) (131, 132, 140). By analogy to ROS and RNS, these compounds are identified as reactive sulfur species (RSS) (67). Similar to ROS and RNS, RSS were first considered to be produced only under pathological conditions and not recognized as being involved in signaling functions. More recently, hydrogen sulfide (H2S) and its sulfane sulfur derivatives have been shown to participate in fundamental biochemical pathways that control cellular redox homeostasis, signaling, metabolism, and mitochondrial function (145, 204); this is perhaps most intriguingly illustrated by the phenomenon of suspended animation observed after H2S inhalation in small rodents (11). These observations have led to a renewed interest in sulfide chemistry and biology (37, 38, 104, 126, 187) and RSS, together with ROS and RNS, to be considered as important physiological signaling molecules.
The view that placed oxygen at the center of the redox regulatory system has been questioned recently by the realization that much of the evolutionary biology of life evolved in a sulfur-rich atmosphere virtually free of oxygen (135, 139). Indeed, it is considered likely that the interaction of RSS with RNS to form S/N-hybrid species participated in forming the building blocks of life and preceded the advent of aerobic respiration and ROS formation (41). In this model, as the level of atmospheric oxygen rose, enabling the development of larger and more energy-efficient organisms, the ancient mechanisms of sulfur metabolism had to face the new challenge of dealing with rapid oxidation processes superimposed onto those that controlled electron transfer in all life forms until then. This model helps to explain why many regulatory pathways are connected to fundamental sulfur-mediated electron transfer processes. As the levels of complexity increased from unicellular to larger multicellular organisms, the fundamental principles of regulation were conserved.
In this review, we provide an integrative biology concept of redox regulation (Fig. 1). We here define the chemical interaction of RSS, RNS, and ROS among themselves and with downstream biological targets as the reactive species interactome (RSI) (Box 1; Table 1). We propose that the RSI serves an integrative function to sense multiple stressors and adjust bioenergetic/metabolic needs accordingly by activating downstream effector pathways to ensure the organism is able to respond to environmental change and stay fit for purpose (Box 2 and Fig. 1). Within this model, H2S along with other thiols is considered an important source of RSS, which have a critical role in enabling and supporting this complex cell signaling network.
FIG. 1.

Intracellular, extracellular, and interorgan/systemic role of the RSI. Precursors of the RSI are organic and inorganic substrates and cofactors, including amino acids (e.g., arginine, methionine), vitamins (B6, B12, C), and xanthine, as well as oxygen, nitrite, polysulfides, thiosulfate, and sulfate, which are transformed by mitochondrial or cytoplasmic enzymes into ROS, RNS, and RSS. The chemical interactions among ROS, RNS, and RSS lead to formation of a number of products with different reactivities, stabilities, half-lives, and therefore different lifetimes defined by their physicochemical properties, covering a wide range of maximal travel distances. A common target of the RSI are cysteine thiols in proteins, acting as redox switches, able to fine-tune activity of signaling molecules, and leading to short-term responses (e.g., protein kinases and phosphatases inducing changes in signaling and glucose metabolism) or long-term adaptation (by modifying redox switches responsible for gene expression regulation, such as the HIF, NFkB, and Keap1/Nrf2 pathways). The RSI serves also as a local and systemic heterocellular communication system mediated by actions of longer-lasting products of the RSI (e.g., nitrite, polysulfides) and circulating thiols. The nutritional and physiological status of the organism affects the RSI by reciprocally regulating precursor availability, metabolism, signaling, and mitochondrial function. RNS, reactive nitrogen species; ROS, reactive oxygen species; RSS, reactive sulfur species.
Box 1. The Reactive Species Interactome.
The reactive species interactome (RSI) is a redox system consisting of chemical interactions of RSS, RNS, and ROS among themselves and with downstream biological targets.
The RSI is characterized by (i) robustness and flexibility; (ii) adaptability; (iii) rapid responsiveness; (iv) ability to sense the environment; and (v) the ability to transduce signals that are required for fine-tuning of biological functions and communication at multiple levels.
The richness of chemical products of the RSI affords the unique redundancy and flexibility of the system. The products of the RSI are continually generated by enzymatic reactions and are as varied as the chemistries of the reactive species themselves. Chemical interactions of the RSI include one- and two-electron oxidation, nitrosation, nitration, and sulfuration/polysulfidation reactions. Each of the species of this interactome has a distinct reactivity and lifetime that are defined by its physicochemical properties (65) and by environmental conditions (such as temperature, pH, and pO2), covering a wide range of maximal travel distances and thus action radii (35, 79, 215).
Being a product of regulated enzymatic transformation of nutrients (amino acids and inorganic substrates such as nitrite and sulfate), the RSI allows rapid adjustment to changes in environmental conditions (e.g., by post-translational modifications) as well as long-term adaptation by regulation of gene expression. It thereby serves an integrative function to sense and transduce multiple stressors, adjust bioenergetic/metabolic needs, and activate downstream effector pathways to ensure the organism stays fit for purpose.
Table 1.
Definition of Terms
| Term | Definition | References |
|---|---|---|
| Stress and adaptation | ||
| Stress/adaptation to stress | The term stress, as used in biomedical literature, was coined by Hans Selye in the 1930s, who defined it as ‘the nonspecific response of the body to any demand for change’ | (166) |
| Cellular stress response | A set of cellular responses (including the downregulation of protein synthesis and upregulation of transcription factors involved in defense and repair mechanisms) common to all eukaryotes aimed at coping with various forms of stress; also known as ‘integrated stress response’ | (141, 150) |
| Hormesis | Beneficial effects of low levels of stress (“what does not kill us makes us stronger”) | (121) |
| Redox chemistry and biology | ||
| Oxidative stress | An imbalance between oxidants and antioxidants in favor of the oxidants, leading to a disruption of redox signaling and control and/or molecular damage | (176) |
| Reactive species | Compounds with high chemical reactivity produced enzymatically or nonenzymatically; the interaction of reactive species may produce other reactive species | (73) |
| Reactive oxygen species (ROS) | Chemically reactive compounds derived from oxygen | |
| Reactive nitrogen species (RNS) | Chemically reactive compounds originating from the reaction of nitric oxide with oxygen or oxygen-derived compounds | |
| Reactive sulfur species (RSS) | According to Jacobs et al. produced by toxification of thiols by reactive oxygen and nitrogen species, including disulfide-S-oxides, sulfenic acids, and thiyl radicals, and predicted to modulate the redox status of biological thiols and disulfides; more recently, RSS were discussed as putative regulatory entities of biological significance | (67, 126) |
| Reactive species interactome (RSI) | Chemical interaction of RSS, RNS, and ROS (possibly including other short-lived species to be discovered in the future) among themselves and with downstream biological targets. The RSI is characterized by (i) high variability/adaptability; (ii) rapid responsiveness; (iii) flexibility, which is required for fine-tuning of biological functions and communication at multiple levels; and (iv) high redundancy (explaining why antioxidants do not work) | This review |
| Redox regulation | A term used to define the control of redox signaling | (176) |
| Redox code | A set of principles that defines the positioning of the nicotinamide adenine dinucleotide (NAD, NADP), thiol/disulfide, and other redox systems, as well as the thiol redox proteome in space and time in biological systems | (94) |
| Personalized and redox medicine | ||
| Redox switches/protein thiol switches | Protein thiols that are specifically and reversibly modified by oxidation, thereby switching the protein between different conformational and functional states | (15,70) |
| Personalized medicine | Healthcare that is tailored to individual condition, needs, and lifestyle (in contrast to one size fits all approach) | |
| Precision medicine | Healthcare that is tailored to the basis of an individual's genes, lifestyle, and environment | (81) |
| Stratified medicine | A therapy that is targeted to a specific patient population on the basis of a clinical characteristic such as a biomarker that predicts treatment response | (194) |
| Redox medicine/redox diseases | Diseases with involvement of aberrant redox regulation/processes ranging from oxidative to reductive stress | (26, 206) |
| P4 medicine | Predictive, personalized, preventive, and participatory (P4) medicine. P4 medicine is fueled by systems approaches to disease, emerging technologies, and analytical tools | (84) |
| Developmental origins of health and disease | A paradigm affirming that environmental influences experienced during early embryonic development may influence the risk of noncommunicable diseases later in life and across generations | (77) |
The table defines the scientific terms used in the Review; the defined terms were marked in italics in the main text.
RNS, reactive nitrogen species; ROS, reactive oxygen species; RSI, reactive species interactome; RSS, reactive sulfur species.
Box 2. The Reactive Species Interactome in Sensing, Signaling, and Adaptation to Stress: Principles of Regulation in the Context of Origins of life, Evolution, and Adaptation.
Organisms observed today represent a snapshot of now—that is, a cross-sectional view encompassing historical experience of preferred life forms that survived past evolutionary stresses, are compatible with the prevailing environment, and fit for purpose. Assuming the overarching biological purpose is reproduction, this requires faithful replication of complex structures and molecular forms, which need to be dynamic yet sufficiently stable to assure structural and functional integrity of the system as a whole.
Biological flexibility in response provides resilience to all sorts of stressors in a constantly changing environment and can be identified at all levels of organization, which can be rationalized in terms of mathematics (networks), (bio)physical, (bio)chemical, and physiological principles, as well as individual behavior and function, group behavior, and social realities on a global scale.
Influenced by Claude Bernard's concept of the milieu interieure and Walter Cannon's notion of homeostasis, the term stress was coined by Hans Selye in the middle of the last century; using experimental animal models, Selye also observed that persistent stress could lead to the development of various diseases (166). Mechanisms enabling to cope with stress are crucial checkpoints for resilience. Adaptation to (perceived or real) environmental, nutritional, lifestyle-related, or mental stresses serves the purpose to improve the fitness of a biological organism to deal with those stresses in the future. The concept of hormesis describes the ability of small stresses to confer protection via activation of the cellular stress response (121), a universal defense reaction of cells to damage to cellular macromolecules (107).
Providing biological flexibility requires energy, which is derived from the release of chemical energy in food to produce heat, readily available forms of energy (e.g., ATP, creatine phosphate), and reducing equivalents [NAD(P)H]; reacting to specific shortages in energy supply requires metabolic plasticity; and adjusting metabolism appropriate to the prevailing conditions requires a sensing and adaptation system. Resilience against stress also demands coupling of the sensing elements to appropriate protection and repair systems that provide a first line of defense (in the form of e.g., antioxidants/antioxidant enzymes) and a system that can identify, repair, or excise damage to biomolecules or tissues caused by endogenous reactive species or exogenous toxicants. Thus, effective buffering against potentially harmful conditions and/or environmental threats requires all four: sensing, adaptation, defense, and repair systems working in concert to confer protection, offer stability, and offset damage. The RSI is involved in sensing/utilizing available energy resources as efficiently as possible by coordinating a system of complex interwoven pathways that ensure smooth operation while allocating sufficient energy for protection and repair of damage to DNA and other critical cell constituents by environmental stressors. Compromised bioenergetic status comes at the price of increased vulnerability to environmental threats.
The current review discusses how (i) the RSI evolved and contributed to shape evolution; (ii) the chemistry of the RSI is linked to cysteine-based redox switches/relays, enabling sensing and transduction of stress signaling for short- and long-term adaptation; (iii) precursor/cofactor availabilities due to changes in intermediary metabolism affect the RSI; (iv) a system-level analysis of these redox signaling elements can contribute to our understanding of fundamental biology and (patho)physiology; and how (v) embracing redox biology in clinical practice and public health can help explain variability in response and thereby contribute to the development of more appropriate sensitive and specific and preventive or therapeutic interventions.
How It All Began: Evolution of the RSI
Life began nearly 4 billion years ago (bya), and ∼85% of all ensuing evolution occurred under anoxic or extremely hypoxic conditions. Rather than oxygen, two other gases, H2S and NO, were present in the early atmosphere and arguably shaped the bulk of the evolution of life on Earth.
Two decades ago, a case was made that NO production by simple life forms may have provided a crucial survival mechanism against ROS at the time of emergence of aerobic life, offering an opportunity for its further utilization as an early signaling molecule (56). Intriguingly, the story of reactive species may have started much earlier. Indeed, more recently, H2S has been implicated in the origin of life (139). The following section places emphasis on the role of sulfur in the evolution of redox metabolic systems; how, at a later stage, O2 replaced some of the roles of sulfur as donor and acceptor of electrons for energy metabolism and signaling; and how reactive species may have contributed to the evolution of life by enabling environmental sensing, metabolic plasticity, and cell–cell communication. Comparisons with other life origin theories, for example, the hydrogen hypothesis, the RNA world, and panspermia, are beyond the scope of this article. For a more comprehensive treatise of how the emerging field of systems chemistry has shaped our understanding of the origin of life and that of metabolism and the fundamentals of biochemical adaptation, the reader is referred elsewhere (20, 80, 149).
Prebiotic primordial interactions: generating the building blocks of life
Life requires a set of essential molecular building blocks from which to assemble more complex structures, enzymes to direct these processes, membranes to partition simultaneously occurring events, and energy to overcome inherent entropies (Box 3). The building blocks of life comprise inorganic or organic precursors of RNA, DNA, and proteins. These were proposed to be derived from (i) intense electrical discharge in a primordial soup containing basic elements, such as carbon, sulfur, and nitrogen (125); (ii) atmospheric photochemical reactions (158); (iii) through volcanic activity and hydrothermal fissures in the Earth's crust; and (iv) extraterrestrial sources ranging from collisions with massive objects (22, 147) to the fine interstellar dust, which continues to add a large amount of organic compounds to Earth on a daily basis (9). However, as only volcanic activity can provide a constant and reliable source of energy, it is most likely that life emerged here (98, 148).
Box 3. Fundamental Principles Underpinning Molecular Interactions in Biology.
1. Chemical principles
Inorganic interactions. The characteristic aqueous environment and energy (in the form of heat) create conditions wherein random interactions lead to the formation of reactive chemicals, based upon the physicochemical characteristics of the atoms/elements (including S, N, O) they contain, prevailing in gaseous and ionic form, and as solutes.
Transition metal chemistry. Transition metals play a special role in catalysis, Fenton reactions, protection against free radicals, redox status (Fe, Mg, Zn, Cu, W, Tn), and display a highly protected/compartmentalized metabolism.
2. Biophysical principles
Organic compounds formed at some stage from carbon and hydrogen, allowing to form alkane/alkene (CH) chains; oxygen enabled carbohydrate (CHO) and fatty acids formation, whereas N and S enabled amino acid and protein formation with special structural characteristics whereas lipids enable boundary formation and compartmentalization.
The bilayer structure of membranes is dependent upon physical rearrangement of the chains according to their lipophilicity / hydrophilicity, i.e. based on physicochemical properties.
3. Biological ordering principles
Compartmentalization enabled cellular and sub-cellular activities, mitochondria increased energetic efficiency, and protein and CHO buffering capacity to bring some stability to the internal environment (physical, pH, redox), with preferential reactions, structured in both space and time.
4. Bioenergetic principles
All reactions have positive and negative consequences. The goal is to achieve a balance of these to support and enable a suitable structure that enables function – ongoing iteration serves to refine both structure and function to make the organism more fit for purpose and better fit for the current environment, its challenges and opportunities.
All processes aimed at counteracting entropy are energetically costly, and a sensible balance must be struck between costs of living and repair. Under stress, repair processes are placed ‘on hold’ until the acute situation is under control; processes impairing mitochondrial activity more chronically have the potential to lead to cellular bioenergetic failure, accumulation of damage and ultimate system failure.
The Earth's earliest atmosphere must have contained large amounts of H2S (198). Sulfide is an efficient reductant and its chemical nature was fundamental for driving protometabolic reactions with N2 and CO2 to form RNA, amino acids, and lipid precursors (142). Our planet has been likened to a primordial reaction cell (148) where energy in the form of reducing equivalents, namely ferrous iron (Fe2+) and sulfide (H2S, HS−, S2−), traversed the Earth's crust through pores (hydrothermal vents) at a steady and therefore dependable rate. Many of these hydrothermal vents sit on massive sulfide deposits called sulfide lenses (168, 169), where the magmatic flow heats the water to over 400°C. The combination of heat, water, and high pressure drives organic synthesis not possible under other conditions, and as the water rises and cools, the products become stable.
An argument can be made for the primacy of sulfide in the origin of life: in combination with transition metals, especially iron, copper, zinc, molybdenum, and tungsten, allowing one- and two-electron transitions, sulfides formed a variety of catalysts that were prototypical enzymes for organic synthesis and created a platform upon which synthesis could occur (33, 163, 200, 201). These metal sulfide minerals formed primordial membranes, allowing compartmentalization of parallel chemical reactions (122). Then, the constant flux of reducing equivalents into the comparatively oxidizing environment of seawater provided a continually renewed and reliable energy source. In addition, some oxidized sulfur coming from the vents gave rise to the formation of defined redox zones.
Evolution of life: from sulfur to oxygen
Life is likely to have begun around hydrothermal vents in a ferrungious (anoxic and Fe2+ rich) ocean ∼3.8 bya (153, 164) and it was chemolithotrophic, completely dependent upon the Earth for energy (Fig. 2). Within a surprisingly short time, 200–400 million years, photosynthesis appeared, which allowed life to become independent from Earth's energy. The earliest light-gathering antennae were not able to harvest enough light to oxidize water and the process was anoxygenic. It has been proposed that an intermediate such as H2O2 was used as the initial electron donor (154). However, H2O2 would have been in short supply, and it is more likely to have been H2S, H2S2, or a related sulfur species (Eq. 1), as seen in modern-day green and purple sulfur bacteria (62).
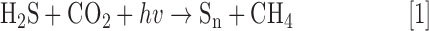 |
FIG. 2.

Evolution of sulfur and oxygen metabolism. The lines indicate fluctuations in concentration of atmospheric oxygen (blue) and oceanic sulfide (orange) over evolutionary times. Atmospheric O2 was essentially absent from the environment at the onset of life ∼3.8 bya. After the great oxidation event (GOE), the concentration of O2 in the atmosphere increased, which was accompanied by a substantial increase in H2S. The first eukaryotes appeared in oceans and developed in anoxic and sulfidic (euxinic) conditions for hundreds of millions of years using sulfur as their energy source, producing RSS. During this time, defense mechanisms against RSS evolved, improving cell survival and minimizing the need for repair of damaged cell constituents. Appearance of oxygenic cyanobacteria and plants led to increases in O2 levels and oxidation of H2S and Fe2+ ∼0.6 bya. Those changes were accompanied by a significant decrease in dissolved H2S and a repurposing of enzymatic systems that originally evolved to protect organisms against RSS to serve additional antioxidative protective functions. Mass extinctions (*, percentage of marine and land life) were often associated with a fall in ambient O2 and increases in H2S, perhaps providing a biological filter for descendants that retained some degree of tolerance to hypoxia and sulfide. Modified with permission from Olson and Straub (139). bya, billion years ago; H2S, hydrogen sulfide.
This reaction is important because H2S was plentiful and the enzymes that evolved to catalyze this reaction could be readily adapted to oxidize H2O once sufficient energy could be extracted from the sun.
Oxygenic photosynthesis likely first appeared in cyanobacteria around 3 bya (Fig. 2). This ultimately led to the great oxidation event around 2.3 bya when atmospheric O2 is thought to have risen to 1%–2%, which is 5%–10% of present atmospheric levels (45, 164). However, apart from small oxygen oases in the shallows, the oceans remained anoxic. Although limited, atmospheric O2 slowly oxidized exposed elemental sulfur and dissolved H2S/HS− to sulfate, which was then carried to the oceans, reduced to H2S by the pervasive Fe2+, and within a hundred million years, vast areas of ocean became euxinic (anoxic and sulfidic). It was in this environment that endosymbiosis, in which a sulfur-reducing Archaea engulfed a sulfide-oxidizing α-protobacterium, produced the mitochondrion around 1.5 bya (111, 165). These early eukaryotes would later incorporate cyanobacteria and thus become the ancestors of modern-day plants. Combined oxygen production by cyanobacteria and primitive plants eventually oxidized all the oceanic iron and sulfide, and around 600 million years ago, atmospheric O2 began to increase to present-day levels (Fig. 2). This oxic environment is generally thought to have had dire consequences due to formation of hydroxyl radicals (HO•), H2O2, and superoxide (O2•−), collectively defined as ROS, (Eq. 2);
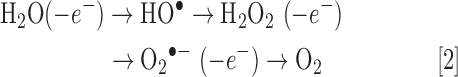 |
According to the OxTox hypothesis, organisms had to develop antioxidant strategies (109), retreat to anoxic niches, or die. However, was it really this bad?
Antioxidant defense or rather sulfur detoxifying strategies?
Before the atmosphere enriched with O2, it is quite likely that the early anoxygenic photosynthesis (Eq. 3) initially evolved as stepwise one-electron oxidation of H2S (Eq. 3);
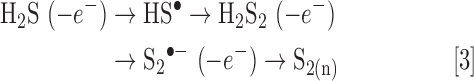 |
Thiyl radicals (HS•), disulfane (H2S2), and persulfide radicals (S2•−, supersulfide) thus generated can indeed be very reactive and are either potent oxidants or reductants (see The RSI: Sensing and Transducing Elements section), collectively known as RSS.
Like ROS, RSS could have had dire consequences: organisms either had to acquire detoxification capability for coping with RSS or die. These capabilities would have to be different from those found in anaerobic organisms, which could escape the oxygen by retreating to anoxic niches. For organisms carrying out anoxygenic photosynthesis, retreating to asulfidic niches was not an option as these did not exist. Therefore, safe disposing of RSS, or their use for signaling or further metabolism, must have enabled the acquisition of appropriate metabolic pathways long before oxygen became prevalent. This would explain why antioxidant systems, such as superoxide dismutase (SOD) (catalyzing reduction of O2•−), catalase and glutathione peroxidase (catalyzing reduction of H2O2), and redox systems governed by thioredoxins, peroxiredoxins, and glutaredoxins, all appeared with the advent of anoxygenic photosynthesis more than 2 bya before they would be called on to deal with ROS (21, 103, 124, 137, 221). Therefore, we suggest that—contrary to common belief (8)—these systems evolved to detoxify RSS and/or to shuttle reducing equivalents for energy utilization or signaling. With the advent of O2, it would have been a relatively trivial matter to switch from dealing with RSS to ROS as their biological chemistries present more similarities than differences. The use of multiple reactive species is in alignment with the need to keep the composition of the internal environment (Claude Bernard's milieu interieure) relatively constant (83).
RSI for sensing and metabolic plasticity
The ready availability of energy in a usable form is a fundamental requirement for survival and reproductive success. In addition to defense and repair systems suitable to cope with harsh environmental conditions, in a world of finite resources, organisms require metabolic flexibility to respond and adapt to changes in environmental conditions (Box 2). The capability of early life forms to adjust their energetic needs and metabolic capability to effectively respond to a variable availability of nutrients/precursors requires an ability to sense and respond to those changes. This would involve the capability to “sniff out” the prevailing conditions in the extracellular environment and adjust metabolic pathways accordingly. The metabolic plasticity required for this responsiveness in living organisms presumes the ability to securely cope with reactive species. Reactive species are formed mainly as enzymatic products from specific organic and inorganic substrates, including amino acids, nitrite, polysulfides, sulfite, sulfate, and O2 (as discussed in detail in the RSI Precursors in the Context of Intermediary Metabolism and Nutrition section). The RSI captures the interaction at the interface between internal and external milieu that enabled metabolic plasticity of early, unicellular life forms and persists in regulating the intersection of co-metabolism and pathogenesis in response to bacterial infection today (155).
From monocellular to multicellular life forms
The development of intercellular communication and the emergence of symbiotic arrangements provided new collaborative opportunities to cope with environmental and infectious threats as well as nutritional shortages. As discussed for NO earlier (Box 4) (56), longer-lived RSI metabolites may later have participated in cell–cell communication and enabled co-metabolic negotiations. As levels of regulatory complexity within those multicellular life forms increased, so did the need for communication. Yet, without appropriate protection, these symbiotic life forms were still vulnerable to threats and dependent on opportunities provided by their local environment. Gaining independence and resilience against external stressors required formation of cell assemblies allowing robust growth and movement. This may have been a driver for the development of larger multicellular organisms with distributed critical functions and enhanced resilience. With redox processes at the heart of global regulation, all organisms larger than perhaps a few hundred cells would require an internal system to communicate metabolic activity status and perceived threat level throughout the entire system, allowing bioenergetic prioritization to survive and reproduce; in other words, an interorgan communication system (see the Interorgan Redox Communication Systems section for further details).
Box 4. Evolution of the RSI – From Sulfur and Nitrogen to Oxygen.
Life is believed to have evolved in a sulfur-rich atmosphere essentially devoid of oxygen, developing bottom-up, first by simple chemical interaction (protometabolism) between atmospheric constituents and reactive materials present on/transported to the Earth's surface comprising redox-active metals, H2S, CO, H2, CH4 and NH3 (not discussed in the present article due to space constraints), nitrogen oxides (NO, NO2. N2O3, NO2−, NO3−) and sources of carbon. Lipid-like membranes would have allowed compartmentalization and formation of primitive single-celled self-replicating organisms feeding on finite local substrates to fulfill their energetic demands. According to this view, the chemical biology of RSS, and their interaction with RNS and later on with ROS governed the emergence and evolution of Life. Furthermore, to cope with reactive species cells had to develop detoxifying systems early on. Interestingly, the detoxifying systems are also based on sulfur-containing elements (i.e. thiols), and NO may have played a key role in intercepting other free radicals. The dynamic and rapid equilibria among RSS and detoxifying systems have probably been one of the most powerful driving forces connecting cellular metabolic capacity with the extracellular milieu, allowing cells to find multiple ways to survive and increase their robustness; this may have included adaptation to changes in the environment and communication to other cells, driving the emergence of symbiotic niches and the development of multicellular organisms.
NO was the first among the reactive species to be proposed to have played a key role in intercellular communication and development of Life on Earth (56) well before the need to deal with the adverse consequences of rising atmospheric O2 levels in the form of unintentional cellular ROS generation (56). We do not know with any certainty where, when and how Life on Earth began – similarly little is known about the onset of biogenic NO formation. Geochemically, NO/NOx would have been formed as a result of lightning and volcanic activity. Since contemporary eukaryotic NO synthases require O2 to generate NO from L-arginine this process would not have worked reliably under hypoxic conditions; biological NO production must therefore have originated from simpler prokaryotic processes such as denitrification (the process of nitrate reduction to dinitrogen) and/or ammonia oxidation (56) where both NO2− and NO are reaction intermediates.
There is an astonishing redundancy and abundance of nitrite reductases in contemporary cells of all kingdoms. One characteristic of eukaryotic nitrite reductases is their susceptibility to inhibition by O2 (55). It is possible that multiple NO producing systems coexisted side-by-side, depending on whether the environment was reducing or oxidizing in nature, what substrates were available and what other biochemical processes this could be linked to. Several atypical NO synthases have meanwhile been discovered in fungi and bacteria (44), and it is likely that other non-classical NO producing enzymes will be identified in other life forms in the future. The nitrogen cycle is probably of similar antiquity as the sulfur cycle since both N and S are essential for Life. Nitrogenase is arguably the most important enzyme in the process of nitrogen fixation by allowing the reduction of N2 to NH3, a key step that enables the incorporation of nitrogen into amino acids, nucleotides and other essential biomolecules. Nitrogenase is produced by cyanobacteria (blue-green algae), green sulfur bacteria and several symbiotic bacteria such as those living on the roots of leguminous plants. The enzyme is oxygen-sensitive and comprises an iron-containing protein that supplies electrons to the FeMo protein, which uses those electrons to reduce N2 to NH3, forming H2 as a by-product. The FeMO protein contains a 4Fe-4S and a Mo-3Fe-3S cluster held in place by a cysteine and a histidine on either end. The presence of iron, molybdenum and sulfur suggests a hydrothermal heritage (135). Thus, there is precedence for early interaction between sulfur and nitrogen metabolism long before the emergence of oxygen.
The RSI: Sensing and Transducing Elements
The rich chemistry of the RSI offers a unique opportunity to fine-tune biological reactions, taking advantage of the diverse chemical nature and lifetimes of the intermediary products formed. The interaction of RNS with ROS, exemplified by the formation of ONOO−, from O2•− and NO has been conceptualized in the form of the chemical biology of NO previously (65, 190). ROS/RSS interaction leads to production of oxidized sulfur species, some of which can further react with biological targets, including cysteine thiolates, to generate, for example, persulfides. Much less is known about the interaction of RSS with RNS to generate S/N hybrid species. However, HSNO/SNO− and SSNO− are prominent examples (37–39) that along with persulfides/polysulfides (37–39, 41) garnered significant interest lately. Persulfides and S/N-hybrid species have a chemical biology with unique characteristics (37). The following section provides a brief overview of known interaction products of biological significance and discusses how their fundamental chemistry dictates their kinetics of formation, action radius, and biological reactivity and how they are particularly fit for purpose in regulated biological systems (Box 3).
The chemical biology of the RSI: interaction of NO, H2S, and O2 and derived species
It is becoming increasingly evident that the signaling and physiological functions of NO, H2S, and O2 should be viewed as components of an integrated whole (219) since they have the potential to interact with each other and affect common biological targets. A comprehensive treatment of all the possible chemical/biochemical interactions between NO, H2S, and O2 (and derived species) and their potential interactions at common biological targets is an enormous undertaking and beyond the scope of this review; other more comprehensive treatments are available (5, 65). Therefore, the possible interactive nature between NO, H2S, and O2 will be discussed in very general terms. However, a more detailed emphasis on sulfide species will be given because this is an area of significant current activity with understanding of much of the chemical biology of these functional groups coming to light only recently.
Of all the small molecule bioregulators, the chemical biology of O2 is clearly the most studied and established. As the ultimate electron acceptor for aerobic life, reduced O2 species such as superoxide (O2•−), hydrogen peroxide (H2O2), and hydroxyl radical (HO•) are thought to be generated enzymatically and nonenzymatically and possess biological relevance. Indeed, all have been proposed to serve as cell signaling agents and/or have pathophysiological consequences. All of these species have been grouped together under the somewhat misleading term ROS even though their reactivities are distinct, highly dependent on the cellular environment, and potentially opposing. For the sake of brevity, it is probably best to categorize the different entities according to their predominant chemical attributes in tabular form (Table 2).
Table 2.
Chemical Attributes of Reactive Oxygen Species, Reactive Nitrogen Species, and Reactive Sulfur Species (Interactions with Metals and Metalloproteins Not Included in This Overview)
| Species | Occurrence/formation | Chemistry |
|---|---|---|
| Dioxygen, O2 | Aerobic life | Has unpaired electrons—reacts readily with other radicals, poor 1e− oxidant, but otherwise easily reduced by 2, 3, and 4e−. |
| Superoxide, O2− | 1e− reduction of O2 | Has one unpaired electron. Good reductant. Under acidic conditions, it can be an oxidant. Reacts with other radicals. |
| Hydrogen peroxide, H2O2 | 1e− reduction of O2− | Two-electron oxidant. Electrophilic. Not a radical species. Can modify RSH (below). |
| Hydroxyl radical, HO• | 1e− reduction of H2O2 | Potent 1e− oxidant. A short-lived radical species capable of abstracting an e− or hydrogen atom from most biological molecules. |
| Nitric oxide, NO | Enzymatic, NO2− reduction | Has unpaired e−. Poor oxidant. Reacts with other radicals (O2 and O2− and other organic radicals [R•]). Can quench radical chemistry. |
| Nitrogen dioxide, NO2 | Oxidation of NO | Good 1e− oxidant. Radical species. Reacts with other radicals (can make R-NO2 when reacted with R•). |
| Dinitrogen trioxide, N2O3 | Oxidation of NO by O2 | Not a radical. Electrophilic and can nitrosate nucleophiles (add equivalent of NO+). Synthesis only relevant at high concentrations of NO. |
| Peroxynitrite, ONOO− | Reaction of NO and O2− | Not a radical, but can generate both HO• and NO2. Rearranges to give nitrate (NO3−). Can oxidize by 2e− (via peroxide-like chemistry). |
| Nitroxyl, HNO | S-nitrosothiol reduction | Reacts readily with thiols. A good hydrogen atom donor. Can act as an antioxidant by quenching radical reactions. |
| Nitrite, NO2− | Oxidation of NO, dietary | Unreactive at neutral pH. Not a radical. Nitrosating agent under acidic conditions. Can be reduced to NO (under acidic conditions). |
| Hydrogen sulfide, H2S | Geochemical, enzymatic | Good metal ligand. Not a radical. Can react with other biological electrophilic sulfur species (e.g., RSSR, RSOH). |
| Thiol, RSH | Endogenous (e.g., cysteine) | Good metal ligand. Not a radical. Can be oxidized to give other biologically relevant sulfur species. |
| Thiyl radical, RS· | 1e− oxidation of RSH | Radical species. Good 1e− oxidant. Will react with other radical species such as NO. |
| Disulfide, RSSR | Oxidation of RSH | Not a radical. Electrophilic. Can be reduced back to RSH under biological conditions. |
| S-Nitrosothiols, RSNO | Nitrosation of RSH | Not a radical. Can be reduced to RSH and HNO. Can transfer NO+ to another thiol (transnitrosation). |
| Sulfenic acid, RSOH | 2e− oxidation of RSH | Not a radical. Electrophilic (reacts with other thiols to give disulfides, RSSR). |
| Hydropersulfide, RSSH | Enzymatic | Not a radical. Superior nucleophile and can be electrophilic (akin to RSSR). Good 1e− reductant. Readily reduced back to RSH. |
| Hydropolysulfide, RSSnH | Oxidation of RSH | Properties similar to RSSH (although enhanced in all aspects). |
| Dialkyl polysulfide, RSSnR | Oxidation of RSH | Properties similar to RSSR (although enhanced in all aspects). Can also be nucleophilic. |
| SSNO− | Unknown, to be determined | Fairly stable. Appears to be a source of NO. Chemical properties pending. |
| SNO− | Unknown | Fleeting. Chemical properties pending. |
Akin to the term ROS, the equivalent terms RNS and RSS denote NO-derived and H2S/RSH-derived species. Undoubtedly, these terms can be equally misleading since the chemical reactivities of RNS and RSS are widely varying and distinct. Regardless, the generation and predominant chemical properties of the RNS and RSS are also listed in Table 2. It is especially noteworthy that the interaction between ROS, RNS, and RSS can lead to products with distinct (and even opposite) chemistry from that of the precursors. For example, the reaction of NO with O2•− to make peroxynitrite (ONOO−) takes two poor oxidants (NO and O2•−) and generates the potentially potent oxidant, ONOO−. As shown in Table 2, ROS, RNS, and RSS taken together cover a wide array of chemical properties ranging from highly reducing (RSSH, O2•−) to highly oxidizing (HO•, NO2), from highly electrophilic (RSOH, H2O2, HNO) to highly nucleophilic (RSSH), and from good hydrogen atom donors (HNO, RSSH) to potent hydrogen atom abstractors (HO•, NO2). This chemical diversity allows Nature to take advantage of widely varying interactive chemistries provided by a limited number of biochemical precursors (namely O2, NO, H2S, and derived species). The cellular conditions conducive to formation of these species imply a selective pressure toward systems that enable a high level of control together with the regulation of cellular function with appropriate biochemical transformations. For sensing/signaling purposes, the biochemical syntheses of ROS, RNS, and RSS must be tightly controlled kinetically, temporally, and spatially. For example, NO biosynthesis can occur via three primary pathways involving NOS enzymes that are distinct with regard to their regulation and location (40, 58). The generation of NO2 from NO is kinetically second order in NO and first order in O2, indicating that significant NO2 levels (at least made via NO/O2 chemistry) can only be produced in compartments possessing high levels of both precursors such as lipid membranes (115). Of note, nitrogen oxide-modified lipids (containing nitrated fatty acids) possess potent biological activities (156). Moreover, significant generation of ONOO− requires that both NO and O2•− be made at the same place, rate, and time (96). This requirement makes ONOO− generation rather difficult, possibly protecting cells from inadvertent formation and narrowing its action radius.
S-nitrosothiol formation can occur in several ways (37, 65). One possibility is the reaction of a free thiol with a nitrosating species, that is, an entity that donates the equivalent of NO+ such as N2O3. Another possibility is the reaction of a thiyl radical with NO. Importantly, N2O3 generation is kinetically restricted (214) (for similar reasons as NO2 generation); as a one-electron oxidant, thiyl radical is very reactive and its formation can only take place under very specific conditions, such as at the active site of enzymes such as ribonucleotide reductase (184). These strict chemical requirements offer a selective advantage by limiting the generation of unwanted reactive and/or deleterious species, which ensures that they are only formed under specific conditions for a particular purpose. By contrast, inadvertent or aberrant generation of any ROS, RNS, or RSS carries the risk of pathophysiological consequences.
Finally, RSS receiving considerable recent attention are hydropersulfides (RSSH). Generation of hydropersulfides from the corresponding thiol represents an oxidation (an RSSH species is at the same oxidation state as a disulfide, RSSR) and can be mediated by several of the oxidants listed in Table 2 (e.g., H2O2 in the presence of H2S). Interestingly, a hydropersulfide is a superior reductant compared with the corresponding thiol. Thus, an extremely potent reductant (RSSH) is made primarily under oxidizing conditions, a fact that seems to have been taken advantage of in nature as it has been proposed that RSSH formation can be protective against oxidative stress (140).
Since RSS are relatively new players in the RSI, some more space is allotted here to the discussion of these species.
The chemistry of persulfides/polysulfides
One of the most well-established reactions of H2S in biochemistry is that with disulfides (10, 60, 140). Reaction of H2S with RSSR yields an equilibrated system involving the corresponding persulfide (RSSH) and thiol (RSH) species (Eq. 4).
 |
RSSH species display a unique chemistry that differs from that of RSH and H2S, conferring potential advantages in biology.
In comparison with RSH, RSSH is both more nucleophilic and reducing. The greater nucleophilicity of RSSH can be explained by (i) the α-affect, in which the electrons of the internal sulfur atom repel those of the external sulfur atom, thus enhancing nucleophilic reactivity—a characteristic lacking in RSH, and (ii) the pKa of RSSH typically being 1–2 units lower than analogous RSH species, making the anionic RSS− present in greater concentrations than RS− at physiological pH.
RSSH is also a more potent one- and two-electron reductant than RSH. The greater two-electron reducing ability of RSSH can be explained by its greater nucleophilic character. The fact that RSSH is a better one-electron reductant than RSH is explained by the stability of the corresponding perthiyl radical (RSS•) over that of the thiyl radical (RS•). Formation of RSS• leads to a resonance-stabilized unpaired electron (shown below) that does not exist for RS• (Eq. 5)
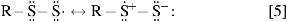
Last and perhaps most intriguing, RSSH are also electrophilic species, whereas RSH are not. Consideration of the oxidation state of sulfur atoms in RSSH reveals that both are in a −1 oxidation state. By comparison, the oxidation state of the sulfur atom of RSH is −2 and therefore RSSH is oxidized with respect to RSH. In this light, RSSH are similar to RSSR (in which both sulfur atoms are also in the −1 oxidation state) and thus are also able to act as an electrophile. Indeed, the electrophilic ability of RSSH is expected to be a function of pH as the deprotonated RSS− species is considered to be less electrophilic (and more nucleophilic) than the protonated RSSH. Electrophilic reactivity of RSSH can yield H2S (via nucleophilic attack on the internal sulfur) or result in a transsulfuration process (via nucleophilic attack at the terminal sulfane sulfur), yielding another RSSH species (Eq. 6) or inorganic polysulfides (i.e., HSSH; Eq. 7).
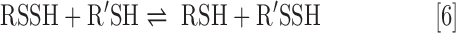 |
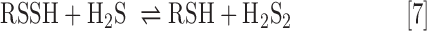 |
S/N hybrid species
Although several groups have investigated the interaction of RSH and nitrogen oxides, specifically NO, mechanisms for the formation of resulting species in biological systems are still controversial. Therefore, the possible reactions of RSH and other related species with NO will be discussed here from a chemical standpoint and implications for biological relevance will be given based on this. As alluded to above, S-nitrosothiols have been reported to have important biological function and serve as biological signaling molecules (17, 59, 179). However, no direct reaction between RSH and NO should be expected to produce such RSNO species because NO has an unpaired electron that occupies an NO antibonding orbital, preventing nucleophilic attack by RSH. However, oxidation of RSH to the corresponding RS• allows for reaction with NO, yielding RSNO (Eq. 8).
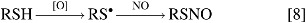
Other pathways leading to RSNO formation include RSH reaction with products from the reaction of O2 and NO (i.e., N2O3, Eq. 9) or by reaction with metal nitrosyl complexes in which the NO ligand acts as a nitrosonium ion (i.e., Fe2+-NO+, Eq. 10). For example, coordination of NO to a ferric iron species (Fe3+) can generate a ferric nitrosyl complex [Fe3+-NO; also described as {Fe(NO)]6 using the Enemark–Feltham notation for metal nitrosyls). This species can be viewed as having significant ferrous nitrosonium character (Fe2+-NO+) and thus can serve as a source of NO+ when reacting with appropriate nucleophiles.
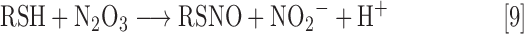 |
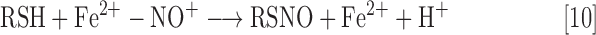 |
Likewise, similar reactivity is predicted for HS− (in comparison with RSH), theoretically leading to formation of HSNO. For the same reasons as outlined for RSH above, no direct reaction between HS− and NO should occur to any significant extent.
Like RSH, RSSH is not expected to react directly with NO. Although one-electron oxidation of RSSH and coworkers to RSS• might be expected to yield the corresponding alkyl-S-nitrosopersulfide (RSSNO) via reaction with NO, recent studies indicate this either does not occur to any great extent (10) or the product has a short lifetime (2). For this reason, RSSNO may not be expected to serve as a biological signaling molecule or NO transporter. Curiously (and unlike thiyl radicals), RSS• is rather stable even in the presence of O2 (10), offering potential opportunities for electron transfer reactions under aerobic conditions (see the Intracellular Redox Regulation, Bioenergetics, and Intermediary Metabolism section).
Contrary to the presumed instability/reversibility of RSSNO, SSNO−ii (37) appears to be relatively stable [a result of the resonance-stabilized anion (10, 120)], existing for extended periods of time even in the presence of other RSH species (39, 41). Although SSNO− has been observed to form under various conditions, including reaction of NO with H2S and polysulfides (HSSnH, n ≥ 2) (41, 120), the exact mechanism for SSNO− formation is unknown. However, it is reasonable to consider that SSNO− is made via reaction of NO with trace polysulfide contaminants present in H2S sources. For example, the presence of trace S2•− (a possible result of one-electron oxidation of S22− or homolytic cleavage of S42−), which is a species well recognized by sulfur chemists to exist in salt melts and heated nonaqueous solutions of sulfur (181), could be expected to react directly with NO, yielding SSNO− (37) (Eq. 10).
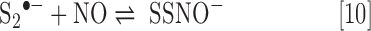 |
It should be noted, however, that to date, SSNO− has yet to be observed in a biological system, leaving its relevance and biological formation still uncertain. Nevertheless, pharmacological SSNO− has been shown to release NO, dilate blood vessels, and activate the prototypical Nrf2 stress response pathway (39, 41, 42).
Cysteine-based redox switches and redox relays
Free sulfhydryl (-SH) groups in low-molecular-weight thiols such as cysteine, peptides (such as GSH), and proteins (e.g., albumin) are predominant targets of RSI signal transduction; others include methionine, tryptophan, tyrosine, and histidine moieties, but their functional significance is not fully understood.
Cysteines may serve structural, catalytic, and regulatory functions in proteins and are considered redox switches as they are targeted for oxidation, nitrosation, thiolation, and sulfidation (also termed sulfhydration). Therefore, rather than on/off switches, protein cysteines may act as multistage cysteine relays (Fig. 3), allowing cells to dynamically adjust protein structure and enzymatic function according to the local redox state (100, 216). In addition to protein thiols, low-molecular-weight thiols, including cysteine and glutathione, are important contributors to intracellular and, via mixed disulfide formation, possibly also extracellular redox status (see section “Interorgan Redox Communication Systems” and corresponding Fig. 5). To function as regulatory elements, those thiol-based post-translational modifications must be also under kinetic control. This is achieved by coupling cysteine-based modifications to a battery of target-specific reductases, denitrosylases, and desulfurases, which together are able to maintain low steady-state concentrations of thiol modifications; these include thioredoxin/thioredoxin reductase, glutaredoxin, peroxiredoxins, and other enzymes (68). Both thiol modifications and their regeneration are dynamically linked to global redox and nutritional status (see the RSI Precursors in the Context of Intermediary Metabolism and Nutrition section and Fig. 1).
FIG. 3.

Cysteine modifications induced by interaction with ROS, RNS, and RSS. The reactive species interactome consists of the interaction of reactive species (ROS, RNS, and RSS) with one another and with cysteine thiols as redox switches (reactions with other functional groups omitted here for the sake of simplicity). The outcome of these interactions depends on the chemical characteristics of the species inducing the modification (e.g., O2•−, H2O2, NO, ONOO−) and their fluxes and the environmental conditions (e.g., pO2, pH), as well as on the reactivity and localization of the targeted cysteines. The lines indicate the outcome of protein cysteine modifications induced by RNS (blue), ROS (red), or RSS (orange).
FIG. 5.

Thiol transport between cell organelles and exchange between the intra- and extracellular compartments. In human plasma, aminothiols such as cysteine, homocysteine, and glutathione exist in free (reduced and oxidized) and protein-bound form, but little is known about the dynamics of their regulation and relationship with each other. As documented for cysteine and glutathione, aminothiols are transported across cell membranes and exchanged between cell organelles, with specific transporters such as the cystine (CysSSCys)/glutamate (Glu) antiporter (Xc), which plays an important role in the regulation of cell surface redox. Continuous arrows indicate known relationships, interrupted arrows represent unknown relationships. In both the intracellular and extracellular compartments, protein thiols represent the main pool of sulfhydryl (-SH) groups. (Note that different font sizes in the figure denote relative concentrations and that mixed disulfides of low-molecular-weight thiols and post-translational thiol modifications are omitted here for the sake of simplicity.) The cytosol is considerably more reduced compared with the extracellular space or the endoplasmic reticulum (where proper protein folding requires more oxidizing conditions). The redox couples, cysteine/cystine, GSH/GSSG, and protein-bound thiols, are not in equilibrium with each other, which suggests the involvement of specific enzyme systems that determine the steady-state levels of these species. Maintaining disequilibria requires energy, and energy tends to be allocated according to criteria that confer robustness of organisms along the evolutionary selection process. First steps into the direction of decoding what determines ATP utilization hierarchies at a systems level are being taken, but the mechanisms of regulation of systemic thiol/disulfide status remain largely obscure. Considering the inverse association of free thiols with risk of death, a further assessment of these relationships and their significance for the cellular stress responses, DNA repair processes, and other hard clinical endpoints seems to be justified. However, no number of observational studies will ever be able to establish causality; this will require prospective and interventional studies.
Biological targets of the RSI
The net biological effects of the reactive species are determined by the nature, level of expression, and function of the biological targets carrying functional cysteine redox switches (Box 1 and Fig. 1). Examples include protein kinases and phosphatases, ion channels, transporters, and enzymes (e.g., those involved in intermediary metabolism), allowing rapid short-term adjustments (Fig. 1). In addition, longer-term regulation is achieved by interaction with redox-sensitive transcription factors, for example, Nrf2/Keap1, NFkB, and HIF (54). Even longer-persisting effects are achieved by redox regulation of gene expression under epigenetic control, making redox effects transmissible to the progeny. This notion is consistent with the developmental origin of health and disease (DOHaD) paradigm, which provides a mechanistic explanation for the pathophysiological basis of how environmental influences experienced during early embryonic development may influence the risk of noncommunicable diseases later in life and across generations (71, 77, 217) (see also the section “Perspective: How Redox Biology and Insights into the Regulation of the RSI May Transform Personalized Medicine”).
Functional significance of the RSI
A corollary of the RSI concept is that reactive species can no longer be regarded as mere stressors (Box 1). Rather, they should be considered controlled reaction products, which serve to sense and transduce information about any changes of internal and/or external conditions; as such, they may be considered as elements of a regulatory system (the RSI) that enable an integrated response to various forms of stress, for example, changes in metabolic, nutritional, and redox status, and environmental conditions (Box 5). See also Boxes 1, 2, and 5.
Box 5. Chemical Biology and Functional Significance of the Reactive Species Interactome.
ROS were initially viewed as mere by-products of redox reactions, especially mitochondrial respiration and certain pathological conditions, leading to oxidative damage of biological targets (protein, lipids, DNA). Furthermore, oxidative reactions were believed to be mediated by RNS, mainly produced by the oxidation of NO. Similarly, cysteine oxidative modification and formation of RSS were first considered only as a consequence of pathological conditions (67). Today, reactive species are considered part of a complex redox signaling network that interacts with protein thiol targets, which act as redox switches to control protein structure and function in dependence of local and global redox and environmental/nutritional status. The analysis of the chemical biology of H2S and related sulfane sulfur species and their interaction with NO and ROS indicate that the RSI is a tightly intertwined redox network that enables rapid sensing and adaptation of the internal cellular milieu to a changing environment. As indicated in Box 1, it is noteworthy that this redox metabolic network appears to have evolved in a world dominated by sulfur and only later incorporating the wider range of options involving nitrogen and oxygen species. This is exactly the opposite order as to how these species were discovered and discussed as contributors to redox biology in the literature. The basis of the interactions among the reactive species in the RSI is defined by the fundamental chemistries of their atomic constituents. However, to be able to operate effectively and enable regulatory control, the system also requires an ability to sense, respond, and adapt to the prevailing state. This is achieved by subjecting both formation and elimination of reactive species and their downstream metabolites to kinetic control through the activity of specific enzymes.
RSI Precursors in the Context of Intermediary Metabolism and Nutrition
The RSI is driven by specific substrates for enzymatic production of individual ROS, RNS, and RSS. Local production of reactive species depends on the availability of O2, certain amino acids, and cofactors, as well as the activity of specific enzymes, which together are embedded within an intricate system of intermediary metabolism that determines the pattern and rate of fluxes according to synthetic and energetic needs (Fig. 4). Given the fundamental role redox regulation plays in cellular defense, repair, and survival, the balance of metabolic fluxes must be prioritized to first support an adequate redox status before fulfilling local metabolic needs. The following section provides a short overview of the precursors needed for ROS, RNS, and RSS synthesis in the context of dietary intake and human nutrition (Fig. 4).
FIG. 4.

Metabolic pathways fueling the RSI. In mammals, L-arginine is formed from citrulline, derived either from dietary glutamine or proline via ornithine and carbamoylphosphate in the mitochondria or from bicarbonate (HCO3−) and ammonia (NH3) via the hepatic urea cycle. Citrulline is then transported via the blood to the kidney where it is converted into arginine. Arginine used for protein formation and (O2-dependent) NO synthesis can be recycled via the arginine/citrulline cycle. NOS activity is inhibited by different methylated arginine residues released by proteolysis (e.g., ADMA). A key interaction between nitrogen and sulfur metabolism is the methylation of arginine using SAM-dependent methyltransferases. SAM is a cofactor produced from methionine and used for methylation of a large number of biomolecules; in the methionine recycling pathway, the removal of one methyl group (-CH3) results in the formation of homocysteine. Depending on the availability of methionine, homocysteine is either recycled to methionine with the help of vitamin B12 and folic acid or is degraded to cystathionine and cysteine. While the latter also serves as precursor of cellular glutathione production, both compounds can generate H2S in the transsulfuration pathway. Not shown here is the formation of ROS via NADPH oxidases, the mitochondrial respiratory chain, and other sources. See Supplementary Figure S1 for more details. ADMA, asymmetric dimethylarginine; NOS, nitric oxide synthases; SAH, S-adenosyl-homocysteine; SAM, S-adenosyl-methionine.
Precursors of ROS, RNS, and RSS: the oxygen–arginine–methionine metabolome
Unsurprisingly, in aerobic organisms, oxygen sensing is intimately linked to intermediary metabolism (1). ROS production involves a variety of different enzymes and organelles utilizing oxygen, but the relationship is not straightforward; counterintuitively, more mitochondrial O2•− is produced in hypoxia than under normoxia (207). O2 is also the substrate of various NADPH oxidases producing either O2− or H2O2 (16, 199). Other sources include xanthine oxidoreductase, 5-lipoxygenase, and cytochrome P450. Nonenzymatic ROS production may also be generated in an unregulated manner through the metal-driven Haber–Weiss reaction leading to the formation of OH•. Enzymatic generation of NO requires both a source of N and O, specifically in the form of arginine (and possibly homoarginine) and O2 (72, 127, 167), whereas NOS-independent reactions leading to NO formation include nitrite/nitrate reduction (43, 90, 116, 172, 192). NOS activity is also dependent on tetrahydrobiopterin and reducing equivalents in the form of NADPH (72, 185). RSS production relies on the availability of methionine, homocysteine, and cysteine serving as substrates in the methionine recycling and transsulfuration pathway. The enzymes of the transsulfuration pathway are responsible for the formation of cysteine from the essential amino acid methionine and serine. Cysteine is crucial for defining protein structure (disulfide bonds), function (e.g., enzymatic activity), redox signaling (e.g., by acting as redox switches, see The RSI: Sensing and Transducing Elements section), and as a building block for glutathione (GSH) production. The tripeptide GSH (Glu-Cys-Gly) is less toxic for cells than cysteine itself, is present in millimolar concentrations intracellularly, and buffers the cellular antioxidant network, together with ascorbate (vitamin C), ubiquinol (coenzyme Q), and α-tocopherol (vitamin E) (175). The two other amino acids critical to GSH synthesis are glycine (itself, in part, derived from serine) and glutamine (formed from glutamate and interaction with proline) (see Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/ars).
Cofactors such as folate, choline, vitamin B6 (pyridoxal phosphate), and B12 (cobalamin) are critically important for adequate methionine recycling. Interestingly, several metabolic aberrations in the tetrahydrofolate cycle, the methionine recycling capacity, or flux through the transsulfuration pathway are associated with elevated homocysteine concentrations in blood. The latter marks metabolic imbalance and/or inadequate nutrient availability and hence is a marker of risk for cardiovascular disease (95, 138). The transsulfuration pathway enzymes, cysteine-β-synthase (CBS), cysteine-γ-lyase (CSE), and 3-mercaptosulfotransferase (MST), are also responsible for the endogenous production of H2S (144) as well as organic persulfides (CBS) (86) and polysulfides (3-MST) (101, 102). CBS is functionally regulated by NO, its expression enhanced by oxidative stress, and gene transcription hormonally regulated in response to fuel supply (182). These pathways therefore can be considered to be a central hub for intermediary metabolism and a point of intersection for the production of proteins (as building block for tRNA and ribosomal protein synthesis), lipids (via S-adenosylmethionine and choline), and methylation reactions (via S-adenosylmethionine), as well as GSH production and H2S/persulfide signaling. This is in accordance with the recent discovery that the nearly 4 billion-year-old metabolism of the last universal common ancestor (LUCA), the forerunner of all contemporary life forms on Earth, already relied on S-adenosylmethionine-dependent 1-carbon metabolism to make a living by harnessing energy from its primordial geological environment (208).
The interaction of H2S, NO, and O2 is tightly linked to bioenergetics through their convergence in the regulation of mitochondrial function. In cultured cells, hypoxic stress induces CSE translocation from the cytosol to mitochondria to sustain ATP production, presumably via fine-tuning of the electron transport chain and use of sulfide as a mitochondrial substrate (63, 78, 188). Marked changes in metabolic needs and/or mitochondrial function are likely to affect precursor/cofactor availabilities and therefore RSI-mediated sensing and adaptation processes (Box 6).
Box 6. Reactive Species Interactome Precursors and Cofactors.
The precursors and cofactors required to support the functioning of the RSI belong to the oxygen–arginine–methionine metabolome and originate from the same pathways that provide the basic building blocks for proteins, lipids, methyl groups, and DNA/RNA synthesis and are thus important for cell proliferation and repair; this suggests competition between anabolic events and redox signaling. The RSI also regulates the expression and activity of enzymes belonging to intermediary metabolism and stress response, highlighting the interactions between catabolism, bioenergetics, and redox status. The reciprocal nature of these relationships indicates that the RSI serves as a central hub that integrates intermediary metabolism and stress signaling (Fig. 1). In addition, tissue/organ functions need to be coordinated and integrated for the sake of optimal fitness of the entire organism; this is achieved via a central communication system, that is, blood, transporting gases, nutrients, and waste products, as well as erythrocytes, immune cells and platelets. Considering the fundamental role of blood in maintaining systemic homeostasis, the RSI itself may participate in systemic redox regulation (see also the Interorgan Redox Communication Systems section).
How nutrition affects precursor availability
L-arginine uptake and metabolism in the human body
In addition to its role in protein biosynthesis, arginine is a precursor for creatine and NO production. There is the need for endogenous formation of arginine, and for young growing mammals, it has to be provided in the diet (i.e., it is an essential amino acid), but less so in adulthood, where it can be considered to be conditionally essential. (189). Inadequate availability of arginine has been associated with T cell and endothelial dysfunction (129); these effects are not usually observed in healthy adults (24) as endogenous synthesis is sufficient to meet usual demands, except in situations of catabolic stress (e.g., inflammation or infection) (128). The net rate of endogenous de novo arginine synthesis is modulated in relation to provision from the diet and the breakdown of proteins (28).
There is evidence for de novo synthesis of arginine in enterocytes up to the age of 3–5 years (202, 212). Beyond this, more complex interorgan amino acid cooperativity is required, which involves enterocytes and the renal cortex (14, 48, 213) (known as the intestinal–renal axis). In enterocytes, endogenous and dietary glutamine is converted into citrulline via glutamate and ornithine (218). Circulating citrulline is then taken up by cells in the renal cortex and converted into arginine (14, 48, 213). The conversion of argininosuccinate to arginine, the final step in arginine de novo synthesis, requires argininosuccinate lyase, which is almost exclusively found in the renal cortex. Hepatic arginine synthesis is embedded in the metabolic pathway of the urea cycle and therefore results in high flux, but low net production (218).
Approximately 60% of net arginine synthesis occurs in the kidney. However, renal insufficiency does not result in decreased plasma arginine concentration, but in increased citrulline levels (112, 218). The mechanisms underlying the maintenance of plasma arginine concentration are poorly understood, but may involve a compensatory decrease in arginine utilization (28).
Only a small proportion (∼1%) of the overall arginine turnover, but a considerable amount (54%) of circulating arginine, is used for NO production (27). In healthy human adults, the production of NO from L-arginine corresponds to ∼1 mmol/day (173). Citrulline, one of the products of NOS, can be recycled by transamination to arginine via the so-called citrulline/NO cycle or arginine/citrulline cycle (76, 218), although in vitro this cycle is much less efficient than the hepatic urea cycle (218). Of note, the guanidino nitrogen group used to form NO is mostly not derived from dietary arginine, but from carbamoylphosphate and aspartate (see Supplementary Fig. S1).
Arginine moieties in proteins can be methylated to form mono- and dimethylated derivatives, which are released into the circulation upon proteolysis. Circulating concentrations of two of these methylated arginine derivatives (L-NG-methylarginine and asymmetric dimethylarginine) are effective inhibitors of cellular arginine uptake and NOS activity (113). While symmetric dimethylarginine does not act as a direct NOS inhibitor, it can reduce NO production by competing with arginine transport (13).
Methionine recycling, transsulfuration, and one-carbon metabolism
Methionine and cysteine are the two sulfur-containing amino acids (SAAs) incorporated into proteins. Methionine is one of the most hydrophobic amino acids. It has important physiological roles, including the initiation of translation via initiation tRNA (met-tRNAimet) and methylation pathways via S-adenosylmethionine (18, 182), which are important for the formation of cofactors such as biotin and lipoic acid. Despite its importance in physiology and being the seventh most abundant element in higher vertebrates, the extent to which the dietary provision of sulfur-related components adequately supports the needs of sulfur metabolism has received inadequate attention (87, 134).
The main sources of sulfur in the diet are inorganic sulfate (SO42−) and SAAs. Methionine can be converted into cysteine, and with a sufficient supply of the former, adequate amounts of the latter can be formed endogenously from serine. However, as this reaction is irreversible, methionine has to be provided preformed in the diet regardless of cysteine status (152). Dietary methionine is absorbed rapidly and almost completely, and only small amounts are excreted directly following bolus administration. It is eliminated from plasma with a half-life of ∼150 min and a threefold increase in urinary SO42−, another important product of transsulfuration (85).
Healthy adults are in sulfur balance with equilibrium between intake, transsulfuration, and excretion. The conversion of methionine to cysteine via homocysteine is the only catabolic pathway of methionine. Sulfur is excreted via the kidney mainly as free sulfate (SO42−, 77%–92%), esterified sulfate (7%–9%), taurine (2%–6%), cyst(e)ine (0.6%–0.7%), and minor amounts of methionine, homocysteine, cystathionine, N-acetylcysteine, mercaptolactate, mercaptoacetate, thiosulfate, and thiocyanate (182). The net changes represented by external balance do not adequately capture the considerable internal flux associated with the turnover of methionine and cysteine into and from protein and peptide pools, estimated as 32 and 38 mmoles per day, respectively (19). Although the daily production of H2S has not been quantified, it is likely considerably higher than that of NO.
Homocysteine represents a determinant branch point for methionine flow either to cysteine through transsulfuration via pyridoxal phosphate-dependent CBS and CSE to cystathionine and cysteine or remethylation via betaine, folate, or vitamin B12 (cobalamin)-dependent pathways (89, 177) (Fig. 4 and Supplementary Fig. S1). Under physiological conditions, there is a similar flow through each pathway (183), but changes in methionine supply or the relative availability of donors for methylation modify the flow through these pathways. Low methionine availability results in high transsulfuration rates (presumably secondary to systemically increased ROS production), whereas a replacement of methionine by cysteine results in increased remethylation (49). Importantly, modulation of dietary SAA intake can affect plasma redox status (93).
Methionine homoeostasis is achieved by modulation of the balance of protein turnover and the relative rates of transsulfuration and remethylation (64, 151, 152, 183). The reductive adaptations developed during malnutrition limit the capacity for handling large doses of methionine, leading to a high plasma concentration of methionine (178) and increased concentration of homocysteine (88). Similar to poor vitamin B6 status and limited serine availability, perturbations in amino acid status can result in homocysteine accumulation, increased remethylation to methionine, and a concomitant reduction in flux through the transsulfuration pathway. Overall, methionine maintains a very stable plasma concentration at the expense of endogenous sulfate production (87).
The sulfur/nitrogen relationship
In adults consuming a normal diet, there is a strong correlation between urinary sulfate and dietary SAA intake, and urinary S:N ratio reflects dietary S:N ratio (157, 171). This relationship is, however, modulated by unusual dietary patterns, for example, during starvation or on low-protein diets when postprandial urinary sulfate excretion is reduced to a greater extent than urea excretion (74, 97, 118, 119). This suggests that protein restriction results not only in preservation of SAA and replenishment of the nonprotein SAA pool but also complex and tight interconnections between the metabolic pathways of nitrogen and sulfur metabolism to maintain constant plasma concentrations of methionine. Thus, alterations in the S:N ratio of urinary metabolites may hold promise as indicators of a stressed system with unusual metabolic demands.
Impact of microbial–host co-metabolism on components of the RSI
Besides the above mammalian pathways, H2S may also be generated from isothiocyanates, which are particularly prevalent in Brassica vegetables such as broccoli (23, 32), polysulfides contained in garlic (7), and via gut microbial reduction of dietary sulfate (SO42−) (23), cysteine, and protein (23, 119). Microbial H2S generation may contribute to the total body pool of sulfide (170) and may even have blood pressure-lowering effects (191). Likewise, dietary nitrite and nitrate can be reduced by the oral and gut microbial flora to NO, contributing to circulating nitrite levels and mildly lower blood pressure (25, 117); together with the enterosalivary recirculation pathway, this has become known as the mammalian N-oxide cycle (25, 116). It is conceivable that a similar sulfur cycle exists in mammalian organisms and those pools of H2S and NO may give rise to reactive species, including ONOO− and possibly SSNO− (37, 39, 41). Fluctuations in intestinal oxygen gradients may further shape the redox relationships between the gut microbiome and the host metabolome (53).
Interorgan Redox Communication Systems
One of the primary functions of the circulatory system in mammals is to efficiently transport oxygen, nutrients, and waste products around the system. With one exception (136), little consideration has been given to its further potential role in acting to communicate and maintain whole-body redox status in relation to external environmental conditions and internal metabolic needs.
Individual cells within a given tissue/organ need to sense their microenvironment in relation to that of the entire organism to achieve a metabolic status that adequately enables the needs of their preferred activity (Box 5). Similarly, the cells need to relate their individual redox status within a composite redox state that matches with other cells and organs. This necessitates reciprocal sensing of intra- and extracellular redox poise. This has to embrace cell membrane behavior and an interorgan communication system that connects the various contributing elements and provides a readout of the global redox poise. Extracellular fluids such as interstitial fluid, lymph, and blood are especially well suited as the connecting medium (36). We propose that the entirety of protein thiols serves as an important redox buffer and that extracellular thiol status helps to mark the global redox state in health and disease.
Quantifying oxidative stress: early attempts
In vitro, oxidative stress is typically assessed by determining the concentration of reduced/oxidized glutathione, antioxidant enzyme (SOD, catalase, glutathione peroxidase) activity, and/or the levels of (anti)oxidants and the potential for free radical scavenging using fluorescent molecules, with all their limitations (205) including their lack of specificity for ROS versus RSS (47). More often than not, little distinction is made between assessing capacity to cope with an oxidant burden, determining the magnitude of exposure to an oxidant, and the balance between exposure and antioxidant capacity. Earlier attempts at quantifying oxidative stress in vivo by measuring levels of select products of lipid, protein, or DNA oxidation (e.g., 8-isoprostanes, malondialdehyde, protein carbonyls, 8-hydroxyguanosine) often met with disappointment, in that all these biomarkers showed distinct profile scaling with oxidative stress burden, but considerable temporal heterogeneity. The reason for this divergence is not immediately apparent and is likely due to the fact that some markers are only bystanders of oxidative damage, while others are actual regulatory nodes of the antioxidant network. Insights into the systems architecture of the redox network and its modus operandi are required to interpret this information. Valuable insight might be derived from information about metabolic fluxes (the direction of travel within metabolic pathways), which can be achieved by either applying stable isotope labeling methodology or monitoring natural isotopic fractionation using high-sensitivity, high-resolution mass spectrometry (fluxomics).
Assessment of low-molecular-weight and protein thiols in plasma/serum
Based upon the assumption that thiols play a determinant role in redox regulation, measuring the ratio of reduced over oxidized forms of small aminothiols such as cysteine or glutathione is potentially more powerful than measuring levels of individual oxidation products inasmuch as it informs us about the status of a dynamic system that serves to shuttle nutrients between cells/organs and electrons between the intracellular and extracellular milieu. Often misunderstood, it is not the electrochemical redox potentials (of e.g., GSH/GSSG) that drive the biology; those concentration ratios are simply the outcome of fast enzymatic processes related to thiol transport, degradation, and regeneration (57). The ratios of reduced/oxidized thiols show diurnal variation (12) and decrease with age, but the redox couples of cysteine and glutathione, for example, are not in equilibrium (92). This indicates that ratios are maintained at defined levels, which might be presumed to confer benefits that are as yet unclear. However, even those measurements reflect only a small part of the overall thiol redox network as it fails to capture the large protein-bound thiol pool (196) and kinetically controlled exchange reactions with free thiols in the intra- and extracellular compartments (Fig. 5). While the overall complexity of this system has been appreciated already some time ago (196), little is known about central regulatory nodes governing these equilibria.
A recent report suggests that cysteine and glutathione redox status are associated with mortality from coronary artery disease (143). Another highly significant association was found between total free thiol status in serum and clinical outcome in unrelated clinical conditions (61, 105). Given the overall complexity of the extracellular antioxidant network and its link to intracellular redox status, the latter was utterly unexpected. A simplistic view of total free thiol levels in a given compartment could be interpreted as a direct reflection of the balance between oxidants and antioxidant capacity (or overall redox poise). Indeed, a decrease of reduced thiols or an increase of oxidized thiols has been found in patients with blood disorders, cancer, cardiovascular disease, diabetes, inflammatory disease, kidney disease, metabolic disease, neurological disease, skin disease, and thyroid disease (3, 108, 110, 123, 133, 180, 220). Thiol oxidation has also been associated with risk factors, including aging, smoking, obesity, and alcohol abuse (69). Both within cells (75) and in blood (195), proteins constitute by far the largest pool of redox-active thiols. Approximately 60% of the total thiol groups in serum/plasma are accounted for by the single free cysteine (Cys34) of albumin (195). Thus, when instead of the ratio of free and oxidized thiols only free thiols are measured, adjustment for total serum protein can be seen as an indirect way of accounting for total thiol content.
In renal transplant recipients, protein-adjusted serum-free thiols were found to predict graft failure and patient survival (61, 105). In a small cohort of stable chronic heart failure patients, there was a positive association between protein-adjusted serum-free thiols and a favorable disease outcome (61, 105). Interestingly, a study evaluating the concentration of serum protein thiols in a wide range of species concluded that free thiol levels are positively associated with life span, suggesting that control of redox status has retained its importance from evolution to modern-day (patho)physiology (146).
Studies relating serum-free thiol levels to other components of the redox network are lacking. In this context, disentangling the relationship between overall thiol redox status (free reduced and oxidized, as well as protein-bound thiols), which is likely affected by cellular uptake and reduction processes, and production/metabolism of both NO and H2S/sulfide would seem to be important. We here propose that protein thiols in the extracellular fluid play a fundamental role in communication between different body compartments, acting as sentinels of distant danger, transporters of specific substrates, and as dynamic entities that reflect a readout of global thiol redox status. Rather than relying on the integrity of a single protein to fulfill this important function, there is greater security with greater buffering capability, and in nature, the entire protein thiol pool may play a role for this purpose. Nevertheless, albumin is likely to play a more important role quantitatively simply based on its abundance in the extracellular compartment (195) and the extent to which it transports small aminothiols. While mixed disulfide formation can occur nonenzymatically via attack of reactive protein thiolates (e.g., Cys34 of albumin) at the disulfide bond of oxidized thiols, the reverse process that would regenerate the free thiol is very slow. This implies that a significant portion of these reactions must be controlled through the activity of specific thiol oxidoreductases such as glutaredoxin, protein disulfide isomerase, and thioredoxin/thioredoxin reductase (70). Besides mixed disulfide formation (S-thiolation), other sulfhydryl modifications, including S-nitrosylation, S-sulfuration (sulfhydration), S-oxidation, and S-acylation (e.g., S-palmitoylation), may also reversibly decrease free thiol availability, but information on the practical relevance of these processes is limited. This may be particularly relevant for cysteinylation and persulfidation of protein thiols as micromolar concentrations of per/polysulfides were detected in biological tissues (86) and therefore have the potential to affect the measurement of total free thiols.
Intracellular redox regulation, bioenergetics, and intermediary metabolism
The same fundamental regulatory principles that operate in the extracellular compartment likely apply to intracellular redox state (Fig. 5). Much emphasis has been devoted to the process of S-cysteinylation and S-glutathionylation (46, 66), although other post-translational modifications may be of similar importance. Crosstalk between cell compartments of widely different redox status (nucleus, endoplasmic reticulum, peroxisomes, and mitochondria) is of significance in this context (30, 51, 161) since the same reactive species that modulate mitochondrial activity and dynamics (130, 211) may also link overall redox poise to intracellular signaling, metabolic control, and bioenergetic status (188). Mitochondrial dysfunction has recently been demonstrated to remodel one-carbon metabolism (4) and the latter is fundamental for mammalian health and disease (50).
These regulatory principles would provide the opportunity to prioritize options for metabolic adjustments in individual cells in relation to the overall redox status of the organism, which might be achieved as a consequence of exchange of redox information across cell membranes (Fig. 5). Although the cystine/glutamate antiporter has been implicated (34), the elements connecting intra- and extracellular spaces to exchange this information are currently unknown. An effective communication across cell membranes requires moieties that reliably operate under widely different redox conditions, that is, under oxidative as well as reductive stress. There are limited choices chemically that fulfill the need of electron exchange under those extremes, but one interesting possibility may involve the persulfide/perthiyl radical couple (10).
Redox state and cell survival
Redox status is also linked to cell survival (193, 222). Effective repair of damage is cardinal to survival and resilience. DNA repair mechanisms have been studied most extensively, not least because the survival of organisms depends on faithful transmission of genetic information from one cell to the next (and across generations), from the level of DNA replication over chromosomal distribution to the repair of damage incurred. This involves surveillance systems for structural monitoring and orchestration of the sophisticated repair processes during normal functioning (31). Spontaneous DNA lesions are common events (82), and the response to DNA damage is principally orchestrated through activation of sensors, transducers, and effectors. This allows resolving problems induced by physical or chemical stresses while limiting unnecessary maintenance. Several DNA repair enzymes, including poly-ADP-ribose polymerase, are under redox control (3), which may provide a mechanistic explanation for the observed association between plasma-free thiols and active disease. Similar processes are at work under conditions of strong emotional stresses, which are known to be associated with oxidative stress, poor immune function and health, lower telomerase activity, and telomere shortening (52). These observations are akin to what happens during normal aging, just at a higher pace. Thus, life stresses of any sort, perceived or real, seem to lead to accelerated (premature) aging. Since parallel monitoring of multiple stresses is energetically costly, integrated stress sensing will be a preferred option; bacteria realize this through distributed sensing of metabolic fluxes (106). A sizable portion of an organism's energy is spent processing sensory information (162), which enables stress tolerance; as the ability of cells to generate adequate levels of energy declines, housekeeping and acute repair processes are compromised and physiological function starts failing.
Perspective: How Redox Biology and Insights into the Regulation of the RSI May Transform Personalized Medicine
The regulatory concepts outlined above, as viewed from the perspective of systems biology, may provide useful guidance in any search for prevention and treatment approaches in the emerging area of redox medicine. Based upon the notion that (i) a set of factors plays a dominant role in sensing environmental changes (O2, amino acid substrates around the arginine-methionine metabolome, and its cofactors), (ii) another set of factors acts as transducing elements (cysteine-based redox relays and the dynamic interplay of free reduced and oxidized, and protein-bound thiols), and that (iii) there are stable end products of the RSI (S,N,O-based stable metabolites), it is possible to create a conceptual framework, which places the complexities of these interactions into context and allows to interrogate the quality and quantity of the balance of forces among different components. The specificity of this approach will be guided not by predefined levels of normality of particular readouts (as commonly practiced in current medicine), but rather by understanding what determines the metabolic fluxes through these pathways (Fig. 6). Due to the interacting nature of the pattern and direction of flow through many pathways within a complex system, the nature and outcome of these interactions will differ depending upon context and circumstance from individual to individual. This underlies the basis of personalized medicine and indicates the diagnostic basis needed to develop rational therapeutic interventions that adequately meet the individual situation (Box 7). In the following, we will present our understanding of redox regulation in the context of health and disease and suggest methods to interrogate the activity status of its components and how to utilize these insights for the benefit of a personalized medicine approach.
FIG. 6.

Simplified scheme visualizing the consequences of a reduced redox buffering capacity (redox reserve) due to bioenergetic limitations. Under normal physiological conditions, extracellular redox poise is subject to diurnal variations, fluctuating between more reducing (−) and more oxidizing (+) conditions (12). The capacity of a cell/organism to deal with changes in reductive and oxidative load is intimately linked to mitochondrial function. Enhanced mitochondrial activity is associated with higher oxidative stress, which affects the redox status of the local tissue microenvironment, while the activity of the mitochondrial respiratory chain itself and mitochondrial intermediary metabolism are modulated by the local and global redox status (upper cartoon). An adequate bioenergetic reserve capacity allows redox stresses (oxidative stress induced by, for example, strenuous physical activity or reductive stress due to chronic overfeeding) into either direction to be comfortably accommodated without incurring damage to cellular constituents (upper panels); physiological redox fluctuations experienced in daily life situations are well within the normal buffering capacity. Mitochondrial dysfunction leads to impaired cellular bioenergetics, resulting in narrowing of a cell's or an organism's ability to buffer redox stresses, inflicting damage and/or compromising adaptive capacities (middle panels), which can result in severe impairment of redox regulatory events upon further bioenergetic challenge (lower panels). Under these conditions, significant damage may be inflicted (shaded areas), demanding the allocation of additional energy to cellular repair processes. According to these relationships, chronic stress triggers a vicious cycle that can lead to a condition associated with severely limited redox reserve capacity, compromising cellular surveillance and repair mechanisms and inviting cascading network failures. This notion is consistent with the links of AMP-activated protein kinase and aberrant mitochondrial gene expression with cellular stress, aging/degenerative processes and immune processes (186), molecular system energetics (159, 160) in general, and the emerging concept of bioenergetic health, where redox biology controls the interface between bioenergetics, autophagy, and circadian control of metabolism (29, 209). See also Box 7.
Box 7. Chemical Biology of the Reactive Species Interactome: The Long and Winding Road from Evolution to Personalized Medicine.
The challenge of developing a dynamic map of redox interactions is related to placing the fundamental chemistry within a given physiological process or disease mechanism into spatial and temporal context. Although the RSI concept begins disentangling the fundamental interplay between the different reactive species and their biological targets, before their use in personalized medicine, specific mechanism-based biomarker panels need to be developed. To this end, the evolutionary origin of redox regulation, based fundamentally on the handling of various sulfur species, may provide a useful pointer since “Nature does not waste a good chemical reaction”.
A case in point is nitrogen fixation and nitrate/nitrite reduction to make NO. Bacteria can produce NO either via ammonia oxidation (e.g., Nitrosomonas) or denitrification (e.g., Pseudomonas), giving rise to NO emanations from soil; plants use NO to stimulate cyclic GMP. Nitrite reductase is a P450-like enzyme that evolved into a NO synthase (NOS) using another source that connects important proliferative and immune responses to arginine metabolism. This evolution helps today's mammalian organisms regulate host defense mechanisms via modulation of mitochondrial respiration and intermediary metabolism (affecting bioenergetics and immunometabolism).
The ultimate thermodynamic equation that connects evolution with precision medicine is that describing Gibbs free energy: ΔG = ΔH − TΔS, where the fundamental reactions of small molecules depend mostly on ΔH or the energy of covalent bond making. However, in the biological context, ΔS means there are ever-increasing differentiated states/niches defined by the process of evolution. Precision medicine is by definition where ΔH meets ΔS. To get there involves a series of approximations: first, the ΔH of the fundamental chemical reactions (The RSI: Sensing and Transducing Elements section); second, interaction of the reactive species metabolome with macromolecules; third, cellular compartmentalization; fourth, tissue interactions; fifth, systemic physiological and inflammatory reactions; sixth, the genomic and epigenetic interaction with these processes; and seventh, the interaction between macroorganisms and microbiome, virome, and exposome (210).
Thermodynamics can tell us into which direction redox reactions may proceed, but it cannot tell us anything about their speed or indeed whether those reactions can ever be fast enough to be of biological significance. Overcoming energetic barriers by using substrate-specific enzymatic catalysis is an important first step, but just the beginning. Redox tunes systems through reaction rates (affecting concentrations), timing, and location—these factors provide precise targets that in nature are exploited to improve biological fitness. The challenge is to unveil how this chemical redox biology operates in health and disease using multibiomarker analysis (via omics) and imaging tools to determine location and timing. Such redox mapping will hold the key to novel therapeutics.
What defines health?
Contemporary medicine still uses the same organ-based classification system physicians developed centuries ago to define disease symptoms. We have become better and better at treating diseases, but not necessarily at identifying the root cause of illness or just “feeling sick” (166). More often than not, disease itself is equated with dysfunction. The converse is our limited ability to define and measure what constitutes health. In medical diagnostic terms, it often means staying within perceived limits of the distribution of variations in simple physiological readouts (such as blood pressure, heart rate, peripheral oxygen saturation, and white blood count) and the absence of overt signs of organ/tissue damage. Systems biology has taught us much about the emergence of unpredicted states arising from the complex interaction of multiple, nonlinear interconnected systems and these lessons are now being incorporated into what constitutes systems medicine. To this end, good health is not merely a state defined by the absence of overt symptoms of disease, but includes overall mental and physical well-being and resilience to stress. Biomarkers of good health are in short supply and limited to a few markers of nutritional adequacy (e.g., amino acid, protein, and micronutrient status) and perceived levels of normality of markers associated with inflammation, oxidative stress cell aging (telomere length), and DNA methylation. However, the latter two tend to differ markedly between individuals and ethnicities and provide limited information about the origin of the problem or the reserve capacity of the regulatory systems affected. From a systems perspective, as long as an organism is alive, one can consider its subsystems to be functioning; however, perturbations may force them to deviate from their normal equilibria and setpoints. If severe enough, these perturbations may become obvious and observable as disease phenotype (e.g., elevated blood pressure, hyperglycemia), which may be interpreted as the price paid by the organism to stay fully operational. To this end, accelerated aging (and aging per se) may be viewed as a process that leads to a compromised capacity for repair and/or adaptive change, eventually culminating in system failure and death (217).
Personalized medicine
Personalized medicine aims at providing individually tailored prevention and treatment strategies for patients. One important driver for this development is the realization that generic treatment approaches are not optimally addressing patients' needs. With an aging population and ever-rising healthcare costs, the idea is to provide tailored solutions matched to specific deviations from the desired equilibrium with the aim to improve population health. In the words of the Horizon2020 Advisory Group, personalized medicine is a medical model using characterization of individuals' phenotypes and genotypes (e.g., molecular profiling, medical imaging, and lifestyle data) for tailoring the right therapeutic strategy for the right person at the right time and/or to determine the predisposition to disease and/or to deliver timely and targeted prevention (see https://ec.europa.eu/research/health/index.cfm?pg=policy&policyname=personalised). The fascination with this concept transcends medicine as it goes well beyond the treatment with medical products to include a better understanding of how particular biological mechanisms and interactions of our body with environmental, nutritional, and infectious stresses [known as the exposome (210)] affect health and disease along the life course. How can these high expectations be fulfilled?
Targeted redox metabolomics for understanding disease processes
Great methodological strides have been made in recent years to capture the entirety of genetic and transcriptomic events in an affordable manner. Beyond the use of multiple omics techniques for deep phenotyping, however, a sound understanding of the key drivers and mechanisms underlying disease processes and resilience is required. Importantly, the mere integration of genomics, transcriptomics, and proteomics does not provide any information about enzyme function/activity and metabolic pathways. Metabolomics, that is, the analysis of stable products of metabolic pathways, may offer a deeper level of information about the state of an organ or the entire organism and may therefore be a key enabling technique for personal/precision medicine (6). Much of the recent success of metabolomic approaches is linked to the rapid advancement in mass spectrometry-based techniques over the last two decades, but many technical obstacles are still to be overcome to extend the level of coverage typical of genomic approaches to metabolic events. Measuring the entire metabolome of every patient may remain beyond reach for some time, but a targeted approach covering key aspects appears realistic even today.
Despite the advancements in untargeted metabolomics to identify and analyze complex patterns of metabolites, some of the low-abundance, polar, or volatile metabolites still pose analytical challenges; in particular, many of the highly reactive species of fleeting existence, for example, H2S and its per- and polysulfides belonging to the RSI, are currently only reliably quantifiable after chemical trapping, providing more stable derivatives (86, 99). A disadvantage of such approaches is that they tend to perturb natural equilibria, not only complicating the interpretation of changes recorded but also introducing bias as their disappearance can elicit adaptive reactions not normally seen under physiological conditions.
What to measure where and how?
To adequately capture the substrates utilized by the RSI for sensing and transduction processes, we propose to concentrate on those parts of intermediary metabolism that are central to the regulation of redox status and production of essential building blocks of life such as DNA/RNA, vitamins, amino acids/proteins, and fatty acids/lipids. Emphasis should be placed on integrative biomarkers faithfully reporting on multiple mechanisms operating in synchrony. Circulating homocysteine concentrations are a pertinent example as they represent a combined readout of methionine and vitamin B6/12 availability, one-carbon metabolism (methionine recycling and tetrahydrofolate pathways), and flux through the transsulfuration pathway, an ancient pathway established almost 4 bya (208); there may well be many more among the markers we routinely assess today. Thus, besides capturing as many species as possible around these pathways to monitor bodily sulfur/SAA handling and H2S production, the arginine metabolome will be another sensible target to capture as it provides insight into nitrogen handling and NO-related processes. To describe the dynamics of the thiol regulatory system, we suggest quantifying the entirety of free and protein-bound thiols; differences in concentration of stable end products of the RSI in different compartments may serve as readouts of the sensing and adaptation system in operation. For sulfur hydrosulfide, per- and polysulfides, thiosulfate, and sulfate would seem to be useful readouts, complemented by ammonium, urea, nitrite, nitrate, nitroso, and nitrosyl species for the nitrogen products. Relevant clinically accessible compartments to analyze include blood (plasma and cellular components), saliva, sweat, urine, and exhaled breath.
To gain insight into the regulation of the entire system, whole-body responses to various stressors ought to be explored first in healthy individuals subjected to, for example, exercise versus rest, hypoxia versus normoxia, heat versus cold, fasting versus controlled feeding, and exposure to frequently prescribed or over-the-counter drugs such as anti-inflammatories. This will form the basis for subsequent interrogations in diseased individuals and patients on specific drug regimens and allow disentangling the underlying redox network architecture using specific patient cohorts. Once we understand how the system is structured, organized, and able to sense in a way that enables control and regulation, we will be in a position to intervene causally to achieve defined objectives.
Summary and Outlook: Disentangling the RSI Using Redox Metabolomics to Enable Precision Medicine
The interactions between individual constituents of the RSI and key biological targets of the antioxidant network are complex. Hoping to disentangle the in vivo relationships for the benefit of defining treatment opportunities for redox medicine will require quantifying stable decomposition products and downstream metabolites of RSS, RNS, and ROS interaction, along with their substrates, in multiple biofluids, in health, and disease. Since we are not dealing with on/off reactions, it would be naive to expect that measuring a few biomarkers in health and comparing them with levels one observes in overt disease would suffice. The perils of oversimplification are nowhere more apparent than in the interpretation of circulating homocysteine concentrations (197). To capture the dynamics of regulation, we ought to measure the same set of readouts in healthy individuals of different sex/age under varying levels of physiological perturbation, (sedentary/exercise, normoxia/hypoxia, starved/fed, etc.) and compare their steady-state levels under these conditions with concentrations in sex/age-matched patients suffering from specific diseases. By carefully matching the physiologic/metabolic phenotypes to the profile of biochemical markers, we should be able to learn enough to decode the underlying network architecture. In turn, this will enable us to identify new avenues for targeted therapeutic modulation of redox signaling. Besides guiding pharmacological treatments, those insights may also aid in risk-stratifying patients before undergoing major surgery, improve recovery from critical illness or major trauma, and inform nutritional priorities for healthy aging.
Supplementary Material
Abbreviations Used
- ADMA
asymmetric dimethylarginine
- bya
billion years ago
- CBS
cysteine-β-synthase
- CSE
cysteine-γ-lyase
- GSH
glutathione
- LUCA
last universal common ancestor
- MST
3-mercaptosulfotransferase
- NO
nitric oxide
- NOS
nitric oxide synthases
- RNS
reactive nitrogen species
- ROS
reactive oxygen species
- RSI
reactive species interactome
- RSS
reactive sulfur species
- S:N ratio
sulfur-to-nitrogen ratio
- SAA
sulfur-containing amino acids
- SAM
S-adenosyl-methionine
- TS
transsulfuration
Acknowledgments
The authors are indebted to many of their colleagues for stimulating discussions over the years; special thanks go to Takaaki Akaike, Anthony R. Butler, Giuseppe Cirino, Michael P. Frenneaux, Michael P. Grocott, Dean Jones, Malte Kelm, Hideo Kimura, John F. Martin, Salvador Moncada, Jerome Santolini, Helmut Sies, Mervyn Singer, David Whitlock, and Steven Wootton. M.M.C.K. acknowledges financial support from the German Research Council (DFG CO 1305/2-1, SFB1116, IRTG1902) and the Forschungskommission of the Universitätsklinikum Düsseldorf; C.L.B. acknowledges support by the US National Science Foundation (CHE 1566065); and P.N. acknowledges support from the Hungarian National Science Foundation (OTKA; grant No.: K 109843). H.v.G. was supported by a grant from the Dutch Kidney Foundation (IP13-114). M.F. is supported by the UK Medical Research Council (G1001536) and the National Institute for Health Research through the NIHR Southampton Biomedical Research Centre. Artwork for the figures was adapted from Servier Medical Art (www.servier.com/Powerpoint-image-bank).
Author Disclosure Statement
M.F. is a member of the Scientific Advisory Board of AOBiome, LLC, a company commercializing the use of ammonia-oxidizing bacteria for the treatment of inflammatory skin diseases and hypertension. None of the other authors have any conflict of interests to declare.
In this context, the word “code” is used to describe a set of principles, rather than a carrier of information like in the genetic code.
We here use the most traditional notation of perthionitrite (SSNO−) as the molecule was originally named by Fritz Seel and coworkers [see Cortese-Krott et al. (37) for a comprehensive review]; however, the notation as nitrosopersulfide (ONSS−) is also correct.
References
- 1.Aragones J, Fraisl P, Baes M, and Carmeliet P. Oxygen sensors at the crossroad of metabolism. Cell Metab 9: 11–22, 2009 [DOI] [PubMed] [Google Scholar]
- 2.Bailey TS, Henthorn HA, and Pluth MD. The intersection of NO and H2S: persulfides generate NO from nitrite through polysulfide formation. Inorg Chem 55: 12618–12625, 2016 [DOI] [PubMed] [Google Scholar]
- 3.Banne AF, Amiri A, and Pero RW. Reduced level of serum thiols in patients with a diagnosis of active disease. J Anti Aging Med 6: 327–334, 2003 [DOI] [PubMed] [Google Scholar]
- 4.Bao XR, Ong S-E, Goldberger O, Peng J, Sharma R, Thompson DA, Vafai SB, Cox AG, Marutani E, and Ichinose F. Mitochondrial dysfunction remodels one-carbon metabolism in human cells. Elife 5: e10575, 2016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Basudhar D, Ridnour LA, Cheng R, Kesarwala AH, Heinecke J, and Wink DA. Biological signaling by small inorganic molecules. Coord Chem Rev 306: 708–723, 2016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Beger RD, Dunn W, Schmidt MA, Gross SS, Kirwan JA, Cascante M, Brennan L, Wishart DS, Oresic M, and Hankemeier T. Metabolomics enables precision medicine: “A White Paper, Community Perspective.” Metabolomics 12: 149, 2016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Benavides GA, Squadrito GL, Mills RW, Patel HD, Isbell TS, Patel RP, Darley-Usmar VM, Doeller JE, and Kraus DW. Hydrogen sulfide mediates the vasoactivity of garlic. Proc Natl Acad Sci U S A 104: 17977–17982, 2007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Benzie IF. Evolution of antioxidant defence mechanisms. Eur J Nutr 39: 53–61, 2000 [DOI] [PubMed] [Google Scholar]
- 9.Bernstein M. Prebiotic materials from on and off the early Earth. Philos Trans R Soc Lond B Biol Sci 361: 1689–1700, 2006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Bianco CL, Chavez TA, Sosa V, Saund SS, Nguyen QN, Tantillo DJ, Ichimura AS, Toscano JP, and Fukuto JM. The chemical biology of the persulfide (RSSH)/perthiyl (RSS.) redox couple and possible role in biological redox signaling. Free Radic Biol Med 101: 20–31, 2016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Blackstone E, Morrison M, and Roth MB. H2S induces a suspended animation-like state in mice. Science 308: 518, 2005 [DOI] [PubMed] [Google Scholar]
- 12.Blanco RA, Ziegler TR, Carlson BA, Cheng PY, Park Y, Cotsonis GA, Accardi CJ, and Jones DP. Diurnal variation in glutathione and cysteine redox states in human plasma. Am J Clin Nutr 86: 1016–1023, 2007 [DOI] [PubMed] [Google Scholar]
- 13.Bode-Böger SM, Scalera F, Kielstein JT, Martens-Lobenhoffer J, Breithardt G, Fobker M, and Reinecke H. Symmetrical dimethylarginine: a new combined parameter for renal function and extent of coronary artery disease. J Am Soc Nephrol 17: 1128–1134, 2006 [DOI] [PubMed] [Google Scholar]
- 14.Borsook H. and Dubnoff JW. The conversion of citrulline to arginine in the kidney. J Biol Chem 141: 717–738, 1941 [Google Scholar]
- 15.Brandes N, Schmitt S, and Jakob U. Thiol-based redox switches in eukaryotic proteins. Antioxid Redox Signal 11: 997–1014, 2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Brandes RP, Weissmann N, and Schroder K. Nox family NADPH oxidases: molecular mechanisms of activation. Free Radic Biol Med 76: 208–226, 2014 [DOI] [PubMed] [Google Scholar]
- 17.Broniowska KA. and Hogg N. The chemical biology of S-nitrosothiols. Antioxid Redox Signal 17: 969–980, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Brosnan JT. and Brosnan ME. The sulfur-containing amino acids: an overview. J Nutr 136: 1636S–1640S, 2006 [DOI] [PubMed] [Google Scholar]
- 19.Bunker VW, Lawson MS, Stansfield MF, and Clayton BE. Nitrogen balance studies in apparently healthy elderly people and those who are housebound. Br J Nutr 57: 211–221, 1987 [DOI] [PubMed] [Google Scholar]
- 20.Caetano-Anolles G, Yafremava LS, Gee H, Caetano-Anolles D, Kim HS, and Mittenthal JE. The origin and evolution of modern metabolism. Int J Biochem Cell Biol 41: 285–297, 2009 [DOI] [PubMed] [Google Scholar]
- 21.Cannio R, Fiorentino G, Morana A, Rossi M, and Bartolucci S. Oxygen: friend or foe? Archaeal superoxide dismutases in the protection of intra- and extracellular oxidative stress. Front Biosci 5: D768–D779, 2000 [DOI] [PubMed] [Google Scholar]
- 22.Capaccioni F, Coradini A, Filacchione G, Erard S, Arnold G, Drossart P, De Sanctis MC, Bockelee-Morvan D, Capria MT, Tosi F, Leyrat C, Schmitt B, Quirico E, Cerroni P, Mennella V, Raponi A, Ciarniello M, McCord T, Moroz L, Palomba E, Ammannito E, Barucci MA, Bellucci G, Benkhoff J, Bibring JP, Blanco A, Blecka M, Carlson R, Carsenty U, Colangeli L, Combes M, Combi M, Crovisier J, Encrenaz T, Federico C, Fink U, Fonti S, Ip WH, Irwin P, Jaumann R, Kuehrt E, Langevin Y, Magni G, Mottola S, Orofino V, Palumbo P, Piccioni G, Schade U, Taylor F, Tiphene D, Tozzi GP, Beck P, Biver N, Bonal L, Combe JP, Despan D, Flamini E, Fornasier S, Frigeri A, Grassi D, Gudipati M, Longobardo A, Markus K, Merlin F, Orosei R, Rinaldi G, Stephan K, Cartacci M, Cicchetti A, Giuppi S, Hello Y, Henry F, Jacquinod S, Noschese R, Peter G, Politi R, Reess JM, and Semery A. Cometary science. The organic-rich surface of comet 67P/Churyumov-Gerasimenko as seen by VIRTIS/Rosetta. Science 347: aaa0628, 2015 [DOI] [PubMed] [Google Scholar]
- 23.Carbonero F, Benefiel AC, Alizadeh-Ghamsari AH, and Gaskins HR. Microbial pathways in colonic sulfur metabolism and links with health and disease. Front Physiol 3: 448, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Carey GP, Kime Z, Rogers QR, Morris JG, Hargrove D, Buffington CA, and Brusilow SW. An arginine-deficient diet in humans does not evoke hyperammonemia or orotic aciduria. J Nutr 117: 1734–1739, 1987 [DOI] [PubMed] [Google Scholar]
- 25.Carlstrom M, Liu M, Yang T, Zollbrecht C, Huang LY, Peleli M, Borniquel S, Kishikawa H, Hezel M, Persson AEG, Weitzberg E, and Lundberg JO. Cross-talk between nitrate-nitrite-NO and NO synthase pathways in control of vascular NO homeostasis. Antioxid Redox Signal 23: 295–306, 2015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Casas AI, Dao VT-V, Daiber A, Maghzal GJ, Di Lisa F, Kaludercic N, Leach S, Cuadrado A, Jaquet V, and Seredenina T. Reactive oxygen-related diseases: therapeutic targets and emerging clinical indications. Antioxid Redox Signal 23: 1171–1185, 2015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Castillo L, Beaumier L, Ajami AM, and Young VR. Whole body nitric oxide synthesis in healthy men determined from [15 N] arginine-to-[15 N]citrulline labeling. Proc Natl Acad Sci U S A 93: 11460–11465, 1996 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Castillo L, Chapman TE, Sanchez M, Yu YM, Burke JF, Ajami AM, Vogt J, and Young VR. Plasma arginine and citrulline kinetics in adults given adequate and arginine-free diets. Proc Natl Acad Sci U S A 90: 7749–7753, 1993 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Chacko BK, Kramer PA, Ravi S, Benavides GA, Mitchell T, Dranka BP, Ferrick D, Singal AK, Ballinger SW, Bailey SM, Hardy RW, Zhang J, Zhi D, and Darley-Usmar VM. The bioenergetic health index: a new concept in mitochondrial translational research. Clin Sci (Lond) 127: 367–373, 2014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Chandel NS. Evolution of mitochondria as signaling organelles. Cell Metab 22: 204–206, 2015 [DOI] [PubMed] [Google Scholar]
- 31.Ciccia A. and Elledge SJ. The DNA damage response: making it safe to play with knives. Mol Cell 40: 179–204, 2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Citi V, Martelli A, Testai L, Marino A, Breschi MC, and Calderone V. Hydrogen sulfide releasing capacity of natural isothiocyanates: is it a reliable explanation for the multiple biological effects of Brassicaceae? Planta Med 80: 610–613, 2014 [DOI] [PubMed] [Google Scholar]
- 33.Cody GD. Transition metal sulfides and the origins of metabolism. Annu Rev Earth Planet Sci 32: 569–599, 2004 [Google Scholar]
- 34.Conrad M. and Sato H. The oxidative stress-inducible cystine/glutamate antiporter, system x (c) (-): cystine supplier and beyond. Amino Acids 42: 231–246, 2012 [DOI] [PubMed] [Google Scholar]
- 35.Cooper CE, Patel RP, Brookes PS, and Darley-Usmar VM. Nanotransducers in cellular redox signaling: modification of thiols by reactive oxygen and nitrogen species. Trends Biochem Sci 27: 489–492, 2002 [DOI] [PubMed] [Google Scholar]
- 36.Corkey BE. and Shirihai O. Metabolic master regulators: sharing information among multiple systems. Trends Endocrinol Metab 23: 594–601, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Cortese-Krott MM, Butler AR, Woollins JD, and Feelisch M. Inorganic sulfur–nitrogen compounds: from gunpowder chemistry to the forefront of biological signaling. Dalton Transact 45: 5908–5919, 2016 [DOI] [PubMed] [Google Scholar]
- 38.Cortese-Krott MM, Fernandez BO, Kelm M, Butler AR, and Feelisch M. On the chemical biology of the nitrite/sulfide interaction. Nitric Oxide 46: 14–24, 2015 [DOI] [PubMed] [Google Scholar]
- 39.Cortese-Krott MM, Fernandez BO, Santos JL, Mergia E, Grman M, Nagy P, Kelm M, Butler A, and Feelisch M. Nitrosopersulfide (SSNO(-)) accounts for sustained NO bioactivity of S-nitrosothiols following reaction with sulfide. Redox Biol 2: 234–244, 2014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Cortese-Krott MM. and Kelm M. Endothelial nitric oxide synthase in red blood cells: key to a new erythrocrine function? Redox Biol 2: 251–258, 2014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Cortese-Krott MM, Kuhnle GG, Dyson A, Fernandez BO, Grman M, DuMond JF, Barrow MP, McLeod G, Nakagawa H, Ondrias K, Nagy P, King SB, Saavedra JE, Keefer LK, Singer M, Kelm M, Butler AR, and Feelisch M. Key bioactive reaction products of the NO/H2S interaction are S/N-hybrid species, polysulfides, and nitroxyl. Proc Natl Acad Sci U S A 112: E4651–E4660, 2015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Cortese-Krott MM, Pullmann D, and Feelisch M. Nitrosopersulfide (SSNO-) targets the Keap-1/Nrf2 redox system. Pharmacol Res 113: 490–499, 2016 [DOI] [PubMed] [Google Scholar]
- 43.Cosby K, Partovi KS, Crawford JH, Patel RP, Reiter CD, Martyr S, Yang BK, Waclawiw MA, Zalos G, Xu X, Huang KT, Shields H, Kim-Shapiro DB, Schechter AN, Cannon Iii RO, and Gladwin MT. Nitrite reduction to nitric oxide by deoxyhemoglobin vasodilates the human circulation. Nat Med 9: 1498–1505, 2003 [DOI] [PubMed] [Google Scholar]
- 44.Crane BR, Sudhamsu J, and Patel BA. Bacterial nitric oxide synthases. Annu Rev Biochem 79: 445–470, 2010 [DOI] [PubMed] [Google Scholar]
- 45.Crowe SA, Dossing LN, Beukes NJ, Bau M, Kruger SJ, Frei R, and Canfield DE. Atmospheric oxygenation three billion years ago. Nature 501: 535–538, 2013 [DOI] [PubMed] [Google Scholar]
- 46.Dalle-Donne I, Rossi R, Giustarini D, Colombo R, and Milzani A. S-glutathionylation in protein redox regulation. Free Radic Biol Med 43: 883–898, 2007 [DOI] [PubMed] [Google Scholar]
- 47.DeLeon ER, Gao Y, Huang E, Arif M, Arora N, Divietro A, Patel S, and Olson KR. A case of mistaken identity: are reactive oxygen species actually reactive sulfide species? Am J Physiol Regul Integr Comp Physiol 310: R549–R560, 2016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Dhanakoti SN, Brosnan JT, Herzberg GR, and Brosnan ME. Renal arginine synthesis: studies in vitro and in vivo. Am J Physiol 259: E437–E442, 1990 [DOI] [PubMed] [Google Scholar]
- 49.Di Buono M, Wykes LJ, Cole DEC, Ball RO, and Pencharz PB. Regulation of sulfur amino acid metabolism in men in response to changes in sulfur amino acid intakes. J Nutr 133: 733–739, 2003 [DOI] [PubMed] [Google Scholar]
- 50.Ducker GS. and Rabinowitz JD. One-carbon metabolism in health and disease. Cell Metab 25: 27–42, 2017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Elsasser TH, Kahl S, Capuco AV, and Schmidt W. Effects of stress on endocrine and metabolic processes and redirection: cross talk between subcellular compartments. Domest Anim Endocrinol 43: 132–145, 2012 [DOI] [PubMed] [Google Scholar]
- 52.Epel ES, Blackburn EH, Lin J, Dhabhar FS, Adler NE, Morrow JD, and Cawthon RM. Accelerated telomere shortening in response to life stress. Proc Natl Acad Sci U S A 101: 17312–17315, 2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Espey MG. Role of oxygen gradients in shaping redox relationships between the human intestine and its microbiota. Free Radic Biol Med 55: 130–140, 2013 [DOI] [PubMed] [Google Scholar]
- 54.Espinosa-Diez C, Miguel V, Mennerich D, Kietzmann T, Sanchez-Perez P, Cadenas S, and Lamas S. Antioxidant responses and cellular adjustments to oxidative stress. Redox Biol 6: 183–197, 2015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Feelisch M, Fernandez BO, Bryan NS, Garcia-Saura MF, Bauer S, Whitlock DR, Ford PC, Janero DR, Rodriguez J, and Ashrafian H. Tissue processing of nitrite in hypoxia an intricate interplay of nitric oxide-generating and-scavenging systems. J Biol Chem 283: 33927–33934, 2008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Feelisch M. and Martin JF. The early role of nitric oxide in evolution. Trends Ecol Evol 10: 496–499, 1995 [DOI] [PubMed] [Google Scholar]
- 57.Flohe L. The fairytale of the GSSG/GSH redox potential. Biochim Biophys Acta 1830: 3139–3142, 2013 [DOI] [PubMed] [Google Scholar]
- 58.Förstermann U. and Sessa WC. Nitric oxide synthases: regulation and function. Eur Heart J 33: 829–837, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Foster MW, McMahon TJ, and Stamler JS. S-nitrosylation in health and disease. Trends Mol Med 9: 160–168, 2003 [DOI] [PubMed] [Google Scholar]
- 60.Francoleon NE, Carrington SJ, and Fukuto JM. The reaction of H(2)S with oxidized thiols: generation of persulfides and implications to H(2)S biology. Arch Biochem Biophys 516: 146–153, 2011 [DOI] [PubMed] [Google Scholar]
- 61.Frenay AS, de Borst MH, Bachtler M, Tschopp N, Keyzer CA, van den Berg E, Bakker SJ, Feelisch M, Pasch A, and van Goor H. Serum free sulfhydryl status is associated with patient and graft survival in renal transplant recipients. Free Radic Biol Med 99: 345–351, 2016 [DOI] [PubMed] [Google Scholar]
- 62.Frigaard NU. and Dahl C. Sulfur metabolism in phototrophic sulfur bacteria. Adv Microb Physiol 54: 103–200, 2009 [DOI] [PubMed] [Google Scholar]
- 63.Fu M, Zhang W, Wu L, Yang G, Li H, and Wang R. Hydrogen sulfide (H2S) metabolism in mitochondria and its regulatory role in energy production. Proc Natl Acad Sci U S A 109: 2943–2948, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Fukagawa NK, Ajami AM, and Young VR. Plasma methionine and cysteine kinetics in response to an intravenous glutathione infusion in adult humans. Am J Physiol Endocrinol Metab 270: E209–E214, 1996 [DOI] [PubMed] [Google Scholar]
- 65.Fukuto JM, Carrington SJ, Tantillo DJ, Harrison JG, Ignarro LJ, Freeman BA, Chen A, and Wink DA. Small molecule signaling agents: the integrated chemistry and biochemistry of nitrogen oxides, oxides of carbon, dioxygen, hydrogen sulfide, and their derived species. Chem Res Toxicol 25: 769–793, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Ghezzi P. and Chan P. Redox proteomics applied to the thiol secretome. Antioxid Redox Signal 26: 399–312, 2017 [DOI] [PubMed] [Google Scholar]
- 67.Giles GI. and Jacob C. Reactive sulfur species: an emerging concept in oxidative stress. Biol Chem 383: 375–388, 2002 [DOI] [PubMed] [Google Scholar]
- 68.Go Y-M, Chandler JD, and Jones DP. The cysteine proteome. Free Radic Biol Med 84: 227–245, 2015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Go Y-M. and Jones DP. Cysteine/cystine redox signaling in cardiovascular disease. Free Radic Biol Med 50: 495–509, 2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Go Y-M. and Jones DP. The redox proteome. J Biol Chem 288: 26512–26520, 2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Godfrey KM, Lillycrop KA, Burdge GC, Gluckman PD, and Hanson MA. Epigenetic mechanisms and the mismatch concept of the developmental origins of health and disease. Pediatr Res 61: 5r–10r, 2007 [DOI] [PubMed] [Google Scholar]
- 72.Griffith OW. and Stuehr DJ. Nitric oxide synthases: properties and catalytic mechanism. Annu Rev Physiol 57: 707–734, 1995 [DOI] [PubMed] [Google Scholar]
- 73.Halliwell B. and Gutteridge JMC. Free Radicals in Biology and Medicine, 3rd Ed. New York, NY: Oxford University Press Inc., 1999. [Google Scholar]
- 74.Hamadeh MJ, Schiffrin A, and Hoffer LJ. Sulfate production depicts fed-state adaptation to protein restriction in humans. Am J Physiol Endocrinol Metab 281: E341–E348, 2001 [DOI] [PubMed] [Google Scholar]
- 75.Hansen RE, Roth D, and Winther JR. Quantifying the global cellular thiol–disulfide status. Proc Natl Acad Sci U S A 106: 422–427, 2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Hecker M, Sessa WC, Harris HJ, Anggard EE, and Vane JR. The metabolism of L-arginine and its significance for the biosynthesis of endothelium-derived relaxing factor: cultured endothelial cells recycle L-citrulline to L-arginine. Proc Natl Acad Sci U S A 87: 8612–8616, 1990 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Heindel JJ, Balbus J, Birnbaum L, Brune-Drisse MN, Grandjean P, Gray K, Landrigan PJ, Sly PD, Suk W, Cory Slechta D, Thompson C, and Hanson M. Developmental origins of health and disease: integrating environmental influences. Endocrinology 156: 3416–3421, 2015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Hildebrandt TM. and Grieshaber MK. Three enzymatic activities catalyze the oxidation of sulfide to thiosulfate in mammalian and invertebrate mitochondria. FEBS J 275: 3352–3361, 2008 [DOI] [PubMed] [Google Scholar]
- 79.Hill BG, Dranka BP, Bailey SM, Lancaster JR, Jr, and Darley-Usmar VM. What part of NO don't you understand? Some answers to the cardinal questions in nitric oxide biology. J Biol Chem 285: 19699–19704, 2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Hochachka P. and Somero G. Biochemical Adaptation, Mechanism and Process in Physiological Evolution. New York, NY: Oxford University Press, 2002. [Google Scholar]
- 81.Hodson R. Precision medicine. Nature 537: S49, 2016 [DOI] [PubMed] [Google Scholar]
- 82.Hoeijmakers JHJ. DNA damage, aging, and cancer. N Engl J Med 361: 1475–1485, 2009 [DOI] [PubMed] [Google Scholar]
- 83.Holmes FL. Claude Bernard, the “Milieu Intérieur,” and regulatory physiology. Hist Philos Life Sci 8: 3–25, 1986 [PubMed] [Google Scholar]
- 84.Hood L. and Friend SH. Predictive, personalized, preventive, participatory (P4) cancer medicine. Nat Rev Clin Oncol 8: 184–187, 2011 [DOI] [PubMed] [Google Scholar]
- 85.Horowitz JH, Rypins EB, Henderson JM, Heymsfield SB, Moffitt SD, Bain RP, Chawla RK, Bleier JC, and Rudman D. Evidence for impairment of transsulfuration pathway in cirrhosis. Gastroenterology 81: 668–675, 1981 [PubMed] [Google Scholar]
- 86.Ida T, Sawa T, Ihara H, Tsuchiya Y, Watanabe Y, Kumagai Y, Suematsu M, Motohashi H, Fujii S, Matsunaga T, Yamamoto M, Ono K, Devarie-Baez NO, Xian M, Fukuto JM, and Akaike T. Reactive cysteine persulfides and S-polythiolation regulate oxidative stress and redox signaling. Proc Natl Acad Sci U S A 111: 7606–7611, 2014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Ingenbleek Y. The nutritional relationship linking sulfur to nitrogen in living organisms. J Nutr 136: 1641S–1651S, 2006 [DOI] [PubMed] [Google Scholar]
- 88.Ingenbleek Y, Hardillier E, and Jung L. Subclinical protein malnutrition is a determinant of hyperhomocysteinemia. Nutrition 18: 40–46, 2002 [DOI] [PubMed] [Google Scholar]
- 89.Ingenbleek Y. and Young VR. The essentiality of sulfur is closely related to nitrogen metabolism: a clue to hyperhomocysteinaemia. Nutr Res Rev 17: 135–151, 2004 [DOI] [PubMed] [Google Scholar]
- 90.Jansson EA, Huang L, Malkey R, Govoni M, Nihlen C, Olsson A, Stensdotter M, Petersson J, Holm L, Weitzberg E, and Lundberg JO. A mammalian functional nitrate reductase that regulates nitrite and nitric oxide homeostasis. Nat Chem Biol 4: 411–417, 2008 [DOI] [PubMed] [Google Scholar]
- 91.Jones DP. Redefining oxidative stress. Antioxid Redox Signal 8: 1865–1879, 2006 [DOI] [PubMed] [Google Scholar]
- 92.Jones DP, Mody VC, Jr, Carlson JL, Lynn MJ, and Sternberg P., Jr Redox analysis of human plasma allows separation of pro-oxidant events of aging from decline in antioxidant defenses. Free Radic Biol Med 33: 1290–1300, 2002 [DOI] [PubMed] [Google Scholar]
- 93.Jones DP, Park Y, Gletsu-Miller N, Liang Y, Yu T, Accardi CJ, and Ziegler TR. Dietary sulfur amino acid effects on fasting plasma cysteine/cystine redox potential in humans. Nutrition 27: 199–205, 2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Jones DP. and Sies H. The redox code. Antiox Redox Signal 23: 734–746, 2015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Joseph J, Handy DE, and Loscalzo J. Quo vadis: whither homocysteine research? Cardiovasc Toxicol 9: 53–63, 2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Jourd'heuil D, Jourd'heuil FL, Kutchukian PS, Musah RA, Wink DA, and Grisham MB. Reaction of superoxide and nitric oxide with peroxynitrite implications for peroxynitrite-mediated oxidation reactions in vivo. J Biol Chem 276: 28799–28805, 2001 [DOI] [PubMed] [Google Scholar]
- 97.Jourdan M, Glock C, Margen S, and Bradfield RB. Sulfate, acid-base, and mineral balances of obese women during weight-loss. Am J Clin Nutr 33: 236–243, 1980 [DOI] [PubMed] [Google Scholar]
- 98.Kalapos MP. A theoretical approach to the link between oxidoreductions and pyrite formation in the early stage of evolution. Biochim Biophys Acta 1553: 218–222, 2002 [DOI] [PubMed] [Google Scholar]
- 99.Kamyshny A, Jr., Gun J, Rizkov D, Voitsekovski T, and Lev O. Equilibrium distribution of polysulfide ions in aqueous solutions at different temperatures by rapid single phase derivatization. Environ Sci Technol 41: 2395–2400, 2007 [DOI] [PubMed] [Google Scholar]
- 100.Kemp M, Go YM, and Jones DP. Nonequilibrium thermodynamics of thiol/disulfide redox systems: a perspective on redox systems biology. Free Radic Biol Med 44: 921–937, 2008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Kimura H. Signaling of hydrogen sulfide and polysulfides. Antioxid Redox Signal 22: 347–349, 2014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Kimura Y, Toyofuku Y, Koike S, Shibuya N, Nagahara N, Lefer D, Ogasawara Y, and Kimura H. Identification of H2S3 and H2S produced by 3-mercaptopyruvate sulfurtransferase in the brain. Sci Rep 5: 14774, 2015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Knoops B, Loumaye E, and Van Der Eecken V. Evolution of the peroxiredoxins. Subcell Biochem 44: 27–40, 2007 [DOI] [PubMed] [Google Scholar]
- 104.Kolluru GK, Shen X, Bir SC, and Kevil CG. Hydrogen sulfide chemical biology: pathophysiological roles and detection. Nitric Oxide 35: 5–20, 2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Koning AM, Meijers WC, Pasch A, Leuvenink HG, Frenay AR, Dekker MM, Feelisch M, de Boer RA, and van Goor H. Serum free thiols in chronic heart failure. Pharmacol Res 111: 452–458, 2016 [DOI] [PubMed] [Google Scholar]
- 106.Kotte O, Zaugg JB, and Heinemann M. Bacterial adaptation through distributed sensing of metabolic fluxes. Mol Syst Biol 6: 355, 2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Kultz D. Molecular and evolutionary basis of the cellular stress response. Annu Rev Physiol 67: 225–257, 2005 [DOI] [PubMed] [Google Scholar]
- 108.Kundi H, Ates I, Kiziltunc E, Cetin M, Cicekcioglu H, Neselioglu S, Erel O, and Ornek E. A novel oxidative stress marker in acute myocardial infarction; thiol/disulphide homeostasis. Am J Emerg Med 33: 1567–1571, 2015 [DOI] [PubMed] [Google Scholar]
- 109.Kurland CG. and Andersson SG. Origin and evolution of the mitochondrial proteome. Microbiol Mol Biol Rev 64: 786–820, 2000 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Lalwani P, de Souza GKBB, de Lima DSN, Passos LFS, Boechat AL, and Lima ES. Serum thiols as a biomarker of disease activity in lupus nephritis. PLoS One 10: e0119947, 2015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Lane N. Power, Sex, Suicide: Mitochondria and the Meaning of Life. Oxford, UK: Oxford University Press, 2005. [Google Scholar]
- 112.Lau T, Owen W, Yu YM, Noviski N, Lyons J, Zurakowski D, Tsay R, Ajami A, Young VR, and Castillo L. Arginine, citrulline, and nitric oxide metabolism in end-stage renal disease patients. J Clin Invest 105: 1217–1225, 2000 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Leiper J. and Nandi M. The therapeutic potential of targeting endogenous inhibitors of nitric oxide synthesis. Nat Rev Drug Discov 10: 277–291, 2011 [DOI] [PubMed] [Google Scholar]
- 114.Lim SS, Vos T, Flaxman AD, Danaei G, Shibuya K, Adair-Rohani H, Amann M, Anderson HR, Andrews KG, Aryee M, Atkinson C, Bacchus LJ, Bahalim AN, Balakrishnan K, Balmes J, Barker-Collo S, Baxter A, Bell ML, Blore JD, Blyth F, Bonner C, Borges G, Bourne R, Boussinesq M, Brauer M, Brooks P, Bruce NG, Brunekreef B, Bryan-Hancock C, Bucello C, Buchbinder R, Bull F, Burnett RT, Byers TE, Calabria B, Carapetis J, Carnahan E, Chafe Z, Charlson F, Chen H, Chen JS, Cheng AT, Child JC, Cohen A, Colson KE, Cowie BC, Darby S, Darling S, Davis A, Degenhardt L, Dentener F, Des Jarlais DC, Devries K, Dherani M, Ding EL, Dorsey ER, Driscoll T, Edmond K, Ali SE, Engell RE, Erwin PJ, Fahimi S, Falder G, Farzadfar F, Ferrari A, Finucane MM, Flaxman S, Fowkes FG, Freedman G, Freeman MK, Gakidou E, Ghosh S, Giovannucci E, Gmel G, Graham K, Grainger R, Grant B, Gunnell D, Gutierrez HR, Hall W, Hoek HW, Hogan A, Hosgood HD, 3rd, Hoy D, Hu H, Hubbell BJ, Hutchings SJ, Ibeanusi SE, Jacklyn GL, Jasrasaria R, Jonas JB, Kan H, Kanis JA, Kassebaum N, Kawakami N, Khang YH, Khatibzadeh S, Khoo JP, Kok C, Laden F, Lalloo R, Lan Q, Lathlean T, Leasher JL, Leigh J, Li Y, Lin JK, Lipshultz SE, London S, Lozano R, Lu Y, Mak J, Malekzadeh R, Mallinger L, Marcenes W, March L, Marks R, Martin R, McGale P, McGrath J, Mehta S, Mensah GA, Merriman TR, Micha R, Michaud C, Mishra V, Mohd Hanafiah K, Mokdad AA, Morawska L, Mozaffarian D, Murphy T, Naghavi M, Neal B, Nelson PK, Nolla JM, Norman R, Olives C, Omer SB, Orchard J, Osborne R, Ostro B, Page A, Pandey KD, Parry CD, Passmore E, Patra J, Pearce N, Pelizzari PM, Petzold M, Phillips MR, Pope D, Pope CA, 3rd, Powles J, Rao M, Razavi H, Rehfuess EA, Rehm JT, Ritz B, Rivara FP, Roberts T, Robinson C, Rodriguez-Portales JA, Romieu I, Room R, Rosenfeld LC, Roy A, Rushton L, Salomon JA, Sampson U, Sanchez-Riera L, Sanman E, Sapkota A, Seedat S, Shi P, Shield K, Shivakoti R, Singh GM, Sleet DA, Smith E, Smith KR, Stapelberg NJ, Steenland K, Stockl H, Stovner LJ, Straif K, Straney L, Thurston GD, Tran JH, Van Dingenen R, van Donkelaar A, Veerman JL, Vijayakumar L, Weintraub R, Weissman MM, White RA, Whiteford H, Wiersma ST, Wilkinson JD, Williams HC, Williams W, Wilson N, Woolf AD, Yip P, Zielinski JM, Lopez AD, Murray CJ, Ezzati M, AlMazroa MA, and Memish ZA. A comparative risk assessment of burden of disease and injury attributable to 67 risk factors and risk factor clusters in 21 regions, 1990–2010: a systematic analysis for the Global Burden of Disease Study 2010. Lancet 380: 2224–2260, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Liu X, Miller MJ, Joshi MS, Thomas DD, and Lancaster JR. Accelerated reaction of nitric oxide with O2 within the hydrophobic interior of biological membranes. Proc Nat Acad Sci U S A 95: 2175–2179, 1998 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Lundberg JO, Gladwin MT, Ahluwalia A, Benjamin N, Bryan NS, Butler A, Cabrales P, Fago A, Feelisch M, Ford PC, Freeman BA, Frenneaux M, Friedman J, Kelm M, Kevil CG, Kim-Shapiro DB, Kozlov AV, Lancaster JR, Jr., Lefer DJ, McColl K, McCurry K, Patel RP, Petersson J, Rassaf T, Reutov VP, Richter-Addo GB, Schechter A, Shiva S, Tsuchiya K, van Faassen EE, Webb AJ, Zuckerbraun BS, Zweier JL, and Weitzberg E. Nitrate and nitrite in biology, nutrition and therapeutics. Nat Chem Biol 5: 865–869, 2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Lundberg JO, Gladwin MT, and Weitzberg E. Strategies to increase nitric oxide signalling in cardiovascular disease. Nat Rev Drug Discov 14: 623–641, 2015 [DOI] [PubMed] [Google Scholar]
- 118.Magee EA, Curno R, Edmond LM, and Cummings JH. Contribution of dietary protein and inorganic sulfur to urinary sulfate: toward a biomarker of inorganic sulfur intake. Am J Clin Nutr 80: 137–142, 2004 [DOI] [PubMed] [Google Scholar]
- 119.Magee EA, Richardson CJ, Hughes R, and Cummings JH. Contribution of dietary protein to sulfide production in the large intestine: an in vitro and a controlled feeding study in humans. Am J Clin Nutr 72: 1488–1494, 2000 [DOI] [PubMed] [Google Scholar]
- 120.Marcolongo JP, Morzan UN, Zeida A, Scherlis DA, and Olabe JA. Nitrosodisulfide [S 2 NO]−(perthionitrite) is a true intermediate during the “cross-talk” of nitrosyl and sulfide. Phys Chem Chem Phys 18: 30047–30052, 2016 [DOI] [PubMed] [Google Scholar]
- 121.Mattson MP. Hormesis and disease resistance: activation of cellular stress response pathways. Hum Exp Toxicol 27: 155–162, 2008 [DOI] [PubMed] [Google Scholar]
- 122.McGlynn SE, Kanik I, and Russell MJ. Peptide and RNA contributions to iron-sulphur chemical gardens as life's first inorganic compartments, catalysts, capacitors and condensers. Philos Trans A Math Phys Eng Sci 370: 3007–3022, 2012 [DOI] [PubMed] [Google Scholar]
- 123.Milevoj Kopčinović L, Domijan A-M, Posavac K, Čepelak I, Žanić Grubišić T, and Rumora L. Systemic redox imbalance in stable chronic obstructive pulmonary disease. Biomarkers 21: 692–698, 2016 [DOI] [PubMed] [Google Scholar]
- 124.Miller AF. Superoxide dismutases: ancient enzymes and new insights. FEBS Lett 586: 585–595, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Miller SL. A production of amino acids under possible primitive earth conditions. Science 117: 528–529, 1953 [DOI] [PubMed] [Google Scholar]
- 126.Mishanina TV, Libiad M, and Banerjee R. Biogenesis of reactive sulfur species for signaling by hydrogen sulfide oxidation pathways. Nat Chem Biol 11: 457–464, 2015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Moncada S, Palmer RM, and Higgs EA. Nitric oxide: physiology, pathophysiology, and pharmacology. Pharmacol Rev 43: 109–142, 1991 [PubMed] [Google Scholar]
- 128.Morris SM. Arginine metabolism: boundaries of our knowledge. J Nutr 137: 1602s–1609s, 2007 [DOI] [PubMed] [Google Scholar]
- 129.Morris SM. Arginases and arginine deficiency syndromes. Curr Opin Clin Nutr Metab Care 15: 64–70, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Murphy MP. Mitochondrial thiols in antioxidant protection and redox signaling: distinct roles for glutathionylation and other thiol modifications. Antioxid Redox Signal 16: 476–495, 2012 [DOI] [PubMed] [Google Scholar]
- 131.Nagy P. Mechanistic chemical perspective of hydrogen sulfide signaling. Methods Enzymol 554: 3–29, 2015 [DOI] [PubMed] [Google Scholar]
- 132.Nagy P. and Winterbourn CC. Redox chemistry of biological thiols. Adv Mol Toxicol 4: 183–222, 2010 [Google Scholar]
- 133.Neuman RB, Bloom HL, Shukrullah I, Darrow LA, Kleinbaum D, Jones DP, and Dudley SC. Oxidative stress markers are associated with persistent atrial fibrillation. Clin Chem 53: 1652–1657, 2007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Nimni ME, Han B, and Cordoba F. Are we getting enough sulfur in our diet? Nutr Metab (Lond) 4: 24, 2007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Nisbet E. and Sleep N. The habitat and nature of early life. Nature 409: 1083–1091, 2001 [DOI] [PubMed] [Google Scholar]
- 136.Nocito L, Kleckner AS, Yoo EJ, Jones IV AR, Liesa M, and Corkey BE. The extracellular redox state modulates mitochondrial function, gluconeogenesis, and glycogen synthesis in murine hepatocytes. PLoS One 10: e0122818, 2015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Novoselov SV. and Gladyshev VN. Non-animal origin of animal thioredoxin reductases: implications for selenocysteine evolution and evolution of protein function through carboxy-terminal extensions. Protein Sci 12: 372–378, 2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Nygård O, Vollset SE, Refsum H, Stensvold I, Tverdal A, Nordrehaug JE, Ueland PM, and Kvåle G. Total plasma homocysteine and cardiovascular risk profile: the hordaland homocysteine study. JAMA 274: 1526–1533, 1995 [DOI] [PubMed] [Google Scholar]
- 139.Olson KR. and Straub KD. The role of hydrogen sulfide in evolution and the evolution of hydrogen sulfide in metabolism and signaling. Physiology 31: 60–72, 2016 [DOI] [PubMed] [Google Scholar]
- 140.Ono K, Akaike T, Sawa T, Kumagai Y, Wink DA, Tantillo DJ, Hobbs AJ, Nagy P, Xian M, Lin J, and Fukuto JM. Redox chemistry and chemical biology of H2S, hydropersulfides, and derived species: implications of their possible biological activity and utility. Free Radic Biol Med 77: 82–94, 2014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Pakos‐Zebrucka K, Koryga I, Mnich K, Ljujic M, Samali A, and Gorman AM. The integrated stress response. EMBO Rep 17: 1374–1395, 2016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Patel BH, Percivalle C, Ritson DJ, Duffy CD, and Sutherland JD. Common origins of RNA, protein and lipid precursors in a cyanosulfidic protometabolism. Nat Chem 7: 301–307, 2015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Patel RS, Ghasemzadeh N, Eapen DJ, Sher S, Arshad S, Ko YA, Veledar E, Samady H, Zafari AM, Sperling L, Vaccarino V, Jones DP, and Quyyumi AA. Novel biomarker of oxidative stress is associated with risk of death in patients with coronary artery disease. Circulation 133: 361–369, 2016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Paul BD. and Snyder SH. Modes of physiologic H2S signaling in the brain and peripheral tissues. Antioxid Redox Signal 22: 411–423, 2015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Paul BD. and Snyder SH. H2S: a novel gasotransmitter that signals by sulfhydration. Trends Biochem Sci 40: 687–700, 2015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Pero RW, Hoppe C, and Sheng Y. Serum thiols as a surrogate estimate of DNA repair correlates to mammalian life span. J Anti Aging Med 3: 241–249, 2000 [Google Scholar]
- 147.Pizzarello S. Molecular asymmetry in prebiotic chemistry: an account from meteorites. Life (Basel) 6: 18, 2016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Pratt AJ. Prebiological evolution and the metabolic origins of life. Artif Life 17: 203–217, 2011 [DOI] [PubMed] [Google Scholar]
- 149.Pross A. What is Life?: How Chemistry Becomes Biology. Oxford, UK: Oxford University Press, 2016. [Google Scholar]
- 150.Quirós PM, Mottis A, and Auwerx J. Mitonuclear communication in homeostasis and stress. Nat Rev Mol Cell Biol 17: 213–226, 2016 [DOI] [PubMed] [Google Scholar]
- 151.Raguso CA, Ajami AM, Gleason R, and Young VR. Effect of cystine intake on methionine kinetics and oxidation determined with oral tracers of methionine and cysteine in healthy adults. Am J Clin Nutr 66: 283–292, 1997 [DOI] [PubMed] [Google Scholar]
- 152.Raguso CA, Regan MM, and Young VR. Cysteine kinetics and oxidation at different intakes of methionine and cystine in young adults. Am J Clin Nutr 71: 491–499, 2000 [DOI] [PubMed] [Google Scholar]
- 153.Raiswell R. and Canfield DE. The iron biogeochemical cycle past and present. Geochem Perspect 1: 1–220, 2012 [Google Scholar]
- 154.Raymond J. and Blankenship RE. The origin of the oxygen-evolving complex. Coord Chem Rev 252: 377–383, 2008 [Google Scholar]
- 155.Richardson AR, Somerville GA, and Sonenshein AL. Regulating the intersection of metabolism and pathogenesis in gram-positive bacteria. Microbiol Spectr 3, 2015; DOI: 10.1128/microbiolspec [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Rudolph V. and Freeman BA. Cardiovascular consequences when nitric oxide and lipid signaling converge. Circ Res 105: 511–522, 2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Sabry ZI, Shadarevian SB, Cowan JW, and Campbell JA. Relationship of dietary intake of sulphur amino-acids to urinary excretion of inorganic sulphate in man. Nature 206: 931–933, 1965 [DOI] [PubMed] [Google Scholar]
- 158.Sagan C. and Khare BN. Long-wavelength ultraviolet photoproduction of amino acids on the primitive Earth. Science 173: 417–420, 1971 [DOI] [PubMed] [Google Scholar]
- 159.Saks VA. Molecular System Bioenergetics: Energy for Life. Hoboken, NJ: John Wiley & Sons; 2007 [Google Scholar]
- 160.Salminen A. and Kaarniranta K. AMP-activated protein kinase (AMPK) controls the aging process via an integrated signaling network. Ageing Res Rev 11: 230–241, 2012 [DOI] [PubMed] [Google Scholar]
- 161.Sandalio LM. and Romero-Puertas MC. Peroxisomes sense and respond to environmental cues by regulating ROS and RNS signalling networks. Ann Bot 116: 475–485, 2015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Sartori P, Granger L, Lee CF, and Horowitz JM. Thermodynamic costs of information processing in sensory adaptation. PLoS Comput Biol 10: e1003974, 2014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Schoepp-Cothenet B, van Lis R, Philippot P, Magalon A, Russell MJ, and Nitschke W. The ineluctable requirement for the trans-iron elements molybdenum and/or tungsten in the origin of life. Sci Rep 2: 263, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Schopf JW. Geological evidence of oxygenic photosynthesis and the biotic response to the 2400-2200 ma “great oxidation event.” Biochemistry (Mosc) 79: 165–177, 2014 [DOI] [PubMed] [Google Scholar]
- 165.Searcy DG. and Lee SH. Sulfur reduction by human erythrocytes. J Exp Zool 282: 310–322, 1998 [DOI] [PubMed] [Google Scholar]
- 166.Selye H. Stress and disease. Science 122: 625–631, 1955 [DOI] [PubMed] [Google Scholar]
- 167.Sessa WC. eNOS at a glance. J Cell Sci 117: 2427–2429, 2004 [DOI] [PubMed] [Google Scholar]
- 168.Shanks III, WCP. Theory of deposit formation in volcanogenic massive sulfide occurrence model: U.S. Geological Survey Scientific Investigations Report 2010-5070-C, Chap. 18, 2012, pp. 12
- 169.Shanks III. WCP and Koski RA. Introduction in volcanogenic massive sulfide occurrence model. In: US Geological Survey Scientific Investigations Report 2010-5070-C, 2012, pp. 5–7
- 170.Shen X, Carlstrom M, Borniquel S, Jadert C, Kevil CG, and Lundberg JO. Microbial regulation of host hydrogen sulfide bioavailability and metabolism. Free Radic Biol Med 60: 195–200, 2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Sherman H. and Hawk P. On the elimination of nitrogen, sulphates, and phosphates after the ingestion of proteid food. Am J Physiol 4: 25–49, 1900 [Google Scholar]
- 172.Shiva S, Wang X, Ringwood LA, Xu X, Yuditskaya S, Annavajjhala V, Miyajima H, Hogg N, Harris ZL, and Gladwin MT. Ceruloplasmin is a NO oxidase and nitrite synthase that determines endocrine NO homeostasis. Nat Chem Biol 2: 486–493, 2006 [DOI] [PubMed] [Google Scholar]
- 173.Siervo M, Stephan B, Feelisch M, and Bluck L. Measurement of in vivo nitric oxide synthesis in humans using stable isotopic methods: a systematic review. Free Radic Biol Med 51: 795–804, 2011 [DOI] [PubMed] [Google Scholar]
- 174.Sies H. Oxidative Stress. London, United Kingdom: Academic Press, 1985, p. 522 [Google Scholar]
- 175.Sies H. Glutathione and its role in cellular functions. Free Radic Biol Med 27: 916–921, 1999 [DOI] [PubMed] [Google Scholar]
- 176.Sies H. Oxidative stress: a concept in redox biology and medicine. Redox Biol 4: 180–183, 2015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Singh S, Padovani D, Leslie RA, Chiku T, and Banerjee R. Relative contributions of cystathionine beta-synthase and gamma-cystathionase to H2S biogenesis via alternative trans-sulfuration reactions. J Biol Chem 284: 22457–22466, 2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Snyderman SE, Holt LE, Jr., Nortn PM, Roitman E, and Phansalkar SV. The plasma aminogram. I. Influence of the level of protein intake and a comparison of whole protein and amino acid diets. Pediatr Res 2: 131–144, 1968 [DOI] [PubMed] [Google Scholar]
- 179.Stamler JS, Lamas S, and Fang FC. Nitrosylation. The prototypic redox-based signaling mechanism. Cell 106: 675–683, 2001 [DOI] [PubMed] [Google Scholar]
- 180.Stephenson ST, Brown LAS, Helms MN, Qu H, Brown SD, Brown MR, and Fitzpatrick AM. Cysteine oxidation impairs systemic glucocorticoid responsiveness in children with difficult-to-treat asthma. J Allergy Clin Immunol 136: 454–461.e9, 2015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Steudel R. Mechanism for the formation of elemental sulfur from aqueous sulfide in chemical and microbiological desulfurization processes. Ind Eng Chem Res 35: 1417–1423, 1996 [Google Scholar]
- 182.Stipanuk MH. Sulfur amino acid metabolism: pathways for production and removal of homocysteine and cysteine. Annu Rev Nutr 24: 539–577, 2004 [DOI] [PubMed] [Google Scholar]
- 183.Storch KJ, Wagner DA, Burke JF, and Young VR. [1-13C; methyl-2H3]methionine kinetics in humans: methionine conservation and cystine sparing. Am J Physiol 258: E790–E798, 1990 [DOI] [PubMed] [Google Scholar]
- 184.Stubbe J. and van der Donk WA. Protein radicals in enzyme catalysis. Chem Rev 98: 705–762, 1998 [DOI] [PubMed] [Google Scholar]
- 185.Stuehr DJ. Enzymes of the L-arginine to nitric oxide pathway. J Nutr 134: 2748S–2751S; discussion 2765S–2767S, 2004 [DOI] [PubMed] [Google Scholar]
- 186.Suhm T. and Ott M. Mitochondrial translation and cellular stress response. Cell Tissue Res 367: 21–31, 2017 [DOI] [PubMed] [Google Scholar]
- 187.Szabo C. Hydrogen sulphide and its therapeutic potential. Nat Rev Drug Discov 6: 917–935, 2007 [DOI] [PubMed] [Google Scholar]
- 188.Szabo C, Ransy C, Módis K, Andriamihaja M, Murghes B, Coletta C, Olah G, Yanagi K, and Bouillaud F. Regulation of mitochondrial bioenergetic function by hydrogen sulfide. Part I. Biochemical and physiological mechanisms. Br J Pharmacol 171: 2099–2122, 2014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Tapiero H, Mathe G, Couvreur P, and Tew KD. Dossier: free amino acids in human health and pathologies - I. Arginine. Biomed Pharmacother 56: 439–445, 2002 [DOI] [PubMed] [Google Scholar]
- 190.Thomas DD, Ridnour LA, Isenberg JS, Flores-Santana W, Switzer CH, Donzelli S, Hussain P, Vecoli C, Paolocci N, Ambs S, Colton CA, Harris CC, Roberts DD, and Wink DA. The chemical biology of nitric oxide: implications in cellular signaling. Free Radic Biol Med 45: 18–31, 2008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.Tomasova L, Dobrowolski L, Jurkowska H, Wrobel M, Huc T, Ondrias K, Ostaszewski R, and Ufnal M. Intracolonic hydrogen sulfide lowers blood pressure in rats. Nitric Oxide 60: 50–58, 2016 [DOI] [PubMed] [Google Scholar]
- 192.Totzeck M, Hendgen-Cotta UB, Luedike P, Berenbrink M, Klare JP, Steinhoff HJ, Semmler D, Shiva S, Williams D, Kipar A, Gladwin MT, Schrader J, Kelm M, Cossins AR, and Rassaf T. Nitrite regulates hypoxic vasodilation via myoglobin-dependent nitric oxide generation. Circulation 126: 325–334, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 193.Trachootham D, Lu W, Ogasawara MA, Nilsa RD, and Huang P. Redox regulation of cell survival. Antioxid Redox Signal 10: 1343–1374, 2008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Trusheim MR, Burgess B, Hu SX, Long T, Averbuch SD, Flynn AA, Lieftucht A, Mazumder A, Milloy J, Shaw PM, Swank D, Wang J, Berndt ER, Goodsaid F, and Palmer MC. Quantifying factors for the success of stratified medicine. Nat Rev Drug Discov 10: 817–833, 2011 [DOI] [PubMed] [Google Scholar]
- 195.Turell L, Radi R, and Alvarez B. The thiol pool in human plasma: the central contribution of albumin to redox processes. Free Radic Biol Med 65: 244–253, 2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196.Ueland PM. Homocysteine species as components of plasma redox thiol status. Clin Chem 41: 340–342, 1995 [PubMed] [Google Scholar]
- 197.Ueland PM. and Loscalzo J. Homocysteine and cardiovascular risk: the perils of reductionism in a complex system. Clin Chem 58: 1623–1625, 2012 [DOI] [PubMed] [Google Scholar]
- 198.Urey HC. On the early chemical history of the Earth and the origin of Life. Proc Natl Acad Sci U S A 38: 351–363, 1952 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Veit F, Pak O, Brandes RP, and Weissmann N. Hypoxia-dependent reactive oxygen species signaling in the pulmonary circulation: focus on ion channels. Antioxid Redox Signal 22: 537–552, 2015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.Wächtershäuser G. Before enzymes and templates: theory of surface metabolism. Microbiol Rev 52: 452–484, 1988 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201.Wächtershäuser G. On the chemistry and evolution of the pioneer organism. Chem Biodivers 4: 584–602, 2007 [DOI] [PubMed] [Google Scholar]
- 202.Wakabayashi Y, Henslee JG, and Jones ME. Pyrroline-5-carboxylate synthesis from glutamate by rat intestinal mucosa. Subcellular localization and temperature stability. J Biol Chem 258: 3873–3882, 1983 [PubMed] [Google Scholar]
- 203.Wang H, Naghavi M, Allen C, Barber RM, Bhutta ZA, Carter A, Casey DC, Charlson FJ, Chen AZ, and Coates MM. Global, regional, and national life expectancy, all-cause mortality, and cause-specific mortality for 249 causes of death, 1980–2015: a systematic analysis for the global burden of disease study 2015. Lancet 388: 1459–1544, 2016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 204.Wang R. Physiological implications of hydrogen sulfide: a whiff exploration that blossomed. Physiol Rev 92: 791–896, 2012 [DOI] [PubMed] [Google Scholar]
- 205.Wardman P. Fluorescent and luminescent probes for measurement of oxidative and nitrosative species in cells and tissues: progress, pitfalls, and prospects. Free Radic Biol Med 43: 995–1022, 2007 [DOI] [PubMed] [Google Scholar]
- 206.Watson JD. Type 2 diabetes as a redox disease. Lancet (London, England) 383: 841–843, 2014 [DOI] [PubMed] [Google Scholar]
- 207.Waypa GB, Smith KA, and Schumacker PT. O2 sensing, mitochondria and ROS signaling: the fog is lifting. Mol Aspects Med 47–48: 76–89, 2016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 208.Weiss MC, Sousa FL, Mrnjavac N, Neukirchen S, Roettger M, Nelson-Sathi S, and Martin WF. The physiology and habitat of the last universal common ancestor. Nat Microbiol 1: 16116, 2016 [DOI] [PubMed] [Google Scholar]
- 209.Wende AR, Young ME, Chatham J, Zhang J, Rajasekaran NS, and Darley-Usmar VM. Redox biology and the interface between bioenergetics, autophagy and circadian control of metabolism. Free Radic Biol Med 100: 94–107, 2016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 210.Wild CP. The exposome: from concept to utility. Int J Epidemiol 41: 24–32, 2012 [DOI] [PubMed] [Google Scholar]
- 211.Willems Peter HGM, Rossignol R, Dieteren Cindy EJ, Murphy Michael P, and Koopman Werner JH. Redox homeostasis and mitochondrial dynamics. Cell Metab 22: 207–218, 2015 [DOI] [PubMed] [Google Scholar]
- 212.Windmueller HG. and Spaeth AE. Uptake and metabolism of plasma glutamine by the small intestine. J Biol Chem 249: 5070–5079, 1974 [PubMed] [Google Scholar]
- 213.Windmueller HG. and Spaeth AE. Source and fate of circulating citrulline. Am J Physiol 241: E473–E480, 1981 [DOI] [PubMed] [Google Scholar]
- 214.Wink DA, Darbyshire JF, Nims RW, Saavedra JE, and Ford PC. Reactions of the bioregulatory agent nitric oxide in oxygenated aqueous media: determination of the kinetics for oxidation and nitrosation by intermediates generated in the nitric oxide/oxygen reaction. Chem Res Toxicol 6: 23–27, 1993 [DOI] [PubMed] [Google Scholar]
- 215.Winterbourn CC. Reconciling the chemistry and biology of reactive oxygen species. Nat Chem Biol 4: 278–286, 2008 [DOI] [PubMed] [Google Scholar]
- 216.Winterbourn CC. and Hampton MB. Thiol chemistry and specificity in redox signaling. Free Radic Biol Med 45: 549–561, 2008 [DOI] [PubMed] [Google Scholar]
- 217.Wootton S. and Jackson A. Influence of under-nutrition in early life on growth, body composition and metabolic competence. In: Long-Term Consequences of Early Environment; Growth, Development and the Lifespan Developmental Perspective (Society for the Study of Human Biology Symposium Series; 37), edited by Henry CJK. and Ulijaszek SJ. Cambridge, England: Cambridge University Press, 1996, pp. 109–123 [Google Scholar]
- 218.Wu G. and Morris SM., Jr Arginine metabolism: nitric oxide and beyond. Biochem J 336 (Pt 1): 1–17, 1998 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 219.Yamasaki H. and Cohen MF. Biological consilience of hydrogen sulfide and nitric oxide in plants: gases of primordial earth linking plant, microbial and animal physiologies. Nitric Oxide 55–56: 91–100, 2016 [DOI] [PubMed] [Google Scholar]
- 220.Yazici C, Köse K, Utaş S, Tanrikulu E, and Taşlidere N. A novel approach in psoriasis: first usage of known protein oxidation markers to prove oxidative stress. Arch Dermatol Res 308: 207–212, 2016 [DOI] [PubMed] [Google Scholar]
- 221.Zamocky M, Gasselhuber B, Furtmuller PG, and Obinger C. Molecular evolution of hydrogen peroxide degrading enzymes. Arch Biochem Biophys 525: 131–144, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 222.Zhang Y, Du Y, Le W, Wang K, Kieffer N, and Zhang J. Redox control of the survival of healthy and diseased cells. Antioxid Redox Signal 15: 2867–2908, 2011 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.