

Abstract
Inflammation is a normal physiological process; however, dysregulation of this process may contribute to inflammatory-based chronic disorders and diseases in animals and humans. Therefore, the antioxidant and anti-inflammatory properties of natural products, often recognized in traditional medicine systems, represent therapeutic modalities to reduce or prevent uncontrolled inflammatory processes which in turn potentially ameliorate or prevent sequelae of inflammatory-based symptoms of chronic diseases. We have investigated the antioxidant and anti-inflammatory effects of honokiol (HNK) and modified citrus pectin (MCP) in vitro and examined whether the MCP : HNK combination has synergistic effects on antioxidant and anti-inflammatory properties. Although both HNK and MCP induced a dose-dependent increase in antioxidant activity, the latter has a consistently higher antioxidant effect. The MCP : HNK (9 : 1) combination induced a synergistic effect on antioxidant activity suggesting that the combination is significantly more efficacious than individual compounds. In mouse monocytes, the lipopolysaccharide- (LPS-) induced tumor necrosis-α (TNF-α) synthesis was significantly inhibited by HNK and the MCP : HNK combination in a dose-dependent manner and synergistic effects were clearly demonstrated with the combination on TNF-α inhibition. This combination effect was also evident on inhibition of nuclear factor-kappa B activity, cyclooxygenase-II activity, and lipid peroxidation in mouse monocytes. Further research into the combination is warranted.
1. Introduction
Dysregulated inflammation is often implicated as a pathophysiological phenomenon underlying many chronic diseases in humans and animals. Biomarkers associated with the responses are those that are often involved in the mediation of inflammation: proinflammatory cytokines, nitric oxide, and lipid mediators including cyclooxygenase enzymes and NF-κB factors produced by inflammatory cells [1, 2]. Inflammation is in part characterized by the activation of the subsets of the innate immune system, such as monocytes and macrophages, and the secretion of inflammatory mediators like tumor necrosis factor-α (TNF-α), prostaglandin E2 (derived from cyclooxygenase-II), and nitric oxide [3]. Plant-derived natural products with antioxidant and anti-inflammatory properties are thus potentially beneficial for prevention and treatment of inflammation-associated chronic diseases.
Honokiol (HNK) is a biphenolic neolignan from magnolia bark (Magnolia officinalis) possessing multiple biological activities including antioxidant, anti-inflammatory anxiolytic, antidepressant, and neuroprotective properties [4–7]. It is widely used in Traditional Chinese Medicine and has already been shown in preclinical studies to be an effective multifunctional antioxidant, used for a wide variety of conditions including dermatological disorders [8], cancer prevention and therapeutics [4], neuromodulation [9], and cardiovascular conditions [10]. HNK at high concentrations is also reported to have anticancer and antiangiogenic properties whereas, at low doses (<10 μM), it is a potent scavenger of super oxide and peroxyl radicals [11]. It contains two phenolic groups which can exhibit antioxidant properties like vitamin E. HNK inhibits freed radical-induced lipid peroxidation [12] and prevents oxidative modification of low-density lipoprotein (LDL), reducing the oxLDL-induced cytotoxicity, apoptotic features, and expression of adhesion molecules in endothelial cells [13].
Modified citrus pectin (MCP) is a well-investigated compound characterized from citrus peels that is recommended as a therapeutic agent for immune support [14], cancer [15–21], heavy metal toxicity [22–24], and fibrotic diseases [25–33]. The mechanism of action of MCP in inflammation, fibrosis, and cancer progression is that it works as a competitive inhibitor of extracellular galectin-3 (Gal-3). MCP blocks Gal-3 activity that drives fibrosis by reducing macrophage activity, proinflammatory cytokine expression, and apoptosis, as well as lowering inflammatory markers, thereby reducing tissue fibrosis. At elevated levels in the circulation, Gal-3 is involved in myofibroblast proliferation, inflammation and fibrogenesis, tissue repair, and ventricular and tissue remodeling [34, 35]. An elevated blood level of Gal-3 is associated with higher risk of death in acute decompensated heart failure and chronic heart failure patients [36] and also associated with the progression of autoimmune disease in studies of rheumatoid arthritis [37].
In the inflammatory cascade, the role of TNF-α and NF-κB activity is well understood [1, 38]. The cytokine, TNF-α, mediates early-stage responses of inflammation by regulating the production of other cytokines, including interleukin-1 (IL-1) and IL-6. Because TNF-α is the main mediator of several inflammatory toxic responses to chemicals, it represents a promising target for the prevention of uncontrolled inflammation. TNF-α has also been reported to induce NF-κB production and this protein is inhibited by the presence of antioxidants [39, 40]. Because the expression of many inflammation-associated genes, including iNOS, COX-2, and TNF-α, are known to be modulated by the binding of NF-κB to its specific promoter regions, it represents a good target for suppressing NF-κB activity for the regulation of lipopolysaccharide- (LPS-) induced inflammation [41]. In the present investigation, we have analyzed the anti-inflammatory and antioxidant properties of MCP, HNK, and the MCP : HNK combination to assess potential synergy for anti-inflammatory and antioxidant properties. A ratio of 9 parts of MCP with 1 part of HNK was used based on a profile of clinically relevant dosages, reduced side effects, and bioavailability data on MCP and HNK for a practical and effective oral capsule delivery use.
2. Materials and Methods
2.1. Test Compounds
Stock solution and dilutions of honokiol (HNK, HonoPure®, ecoNugenics, Santa Rosa, CA) and Modified Citrus Pectin (PectaSol-C®, ecoNugenics, Santa Rosa, CA) were prepared in sterile phosphate buffered saline (PBS) for treatment. The MCP : HNK combination was prepared in a ratio of 9 parts of MCP to 1 part of HNK (i.e., nine grams of MCP mixed with 1 gram of HNK) and dilutions of the mix were prepared in PBS. Test compounds were prepared at the highest concentration of 100 mg/ml in sterile PBS at 60°C. The amount of compounds that have been dissolved is calculated based on the weight of undissolved residue separated by centrifugation. The solubility of MCP and HNK in PBS was 76.4% and 100%, respectively. The volume was adjusted to get accurate amount of the compound for treatment based on solubility factor.
2.2. Antioxidant Activity
The principle of the antioxidant assay is based on the formation of a ferryl myoglobin radical from metmyoglobin and hydrogen peroxide, which oxidizes the 2,2′-azino-bis(3-ethylbenzthiazoline-6-sulfonic acid) (ABTS) to produce a radical cation, ABTS+, a soluble chromogen that is green in color and can be measured in a spectrophotometer at 405 nm. Antioxidants suppress the production of the radical cation in a concentration dependent manner and the color intensity decreases proportionally. Trolox, a water-soluble vitamin E analog, was used as a control antioxidant for analyzing the antioxidant activity of HNK, MCP, and MCP : HNK (9 : 1). In a 96-well plate, the assay was set up with 10 µl of increasing concentrations of compounds (0–500 µg/ml) and 20 µl of myoglobin working solution per the protocol described in Antioxidant Assay Kit (Sigma-Aldrich, St. Louis, MO). Afterward, 150 μl of ABTS working solution containing 0.0075% H2O2 was added and incubated at room temperature for 5 min. The reaction was stopped by adding 100 µl stop solution and absorbance was measured at 405 nm in a Bio-Rad Benchtop microplate reader. The decrease in absorbance indicated the antioxidant activity of HNK, MCP, and MCP : HNK (9 : 1) equivalent to the Trolox standard, which was plotted against concentrations of compounds [42].
2.3. Lipopolysaccharide-Induced TNF-α Synthesis
Mouse monocytes (0.5 × 106/ml) were plated in 24-well plates and starved overnight by growing in minimal essential medium containing 0.5% fetal bovine serum and antibiotics. On the following day, the plates were replaced with fresh starving medium and treated with increasing concentrations of HNK, MCP, and MCP : HNK (9 : 1) in the presence and absence of LPS. The compound was added initially and after incubation for 2 h at 37°C; 20 ng/ml LPS was added to induce an inflammatory response. The plate was incubated for an additional 4 h and culture medium was collected, centrifuged, and stored at −80°C. TNF-α produced and secreted into the medium by the cells was analyzed by ELISA protocol using the mouse TNF-α Quantikine ELISA kit (R&D systems, Minneapolis, MN) as per manufacturer's instructions [43].
2.4. Lipid Peroxidation
RAW 264.7 mouse monocytes (3 × 106 cells/5 ml) were treated with increasing concentrations of MCP, HNK, and MCP : HNK (9 : 1) (0–2000 µg/ml) at 37°C for 72 h before they were challenged with 20 µM H2O2 overnight. The cell lysate was prepared per the Lipid Peroxidation Assay protocol (Sigma-Aldrich, St. Louis, MO) and protein concentrations were determined. The cell lysate (200 µl) was analyzed for inhibition of lipid peroxidation per manufacturer's protocol. Lipid peroxidation is determined by the reaction of malondialdehyde (MDA) with thiobarbituric acid (TBA) to form a calorimetric (532 nm) product proportional to the MDA present. To form the MDA-TBA adduct 600 µl of TBA solution was added to the 200 µl of lysate and incubated at 95°C for 1 h. The reaction mix was cooled in an ice bath for 10 min and absorbance recorded at 532 nm in a Beckman spectrophotometer. MDA standards (0–2 nmoles) reacted with TBA were used as a standard for the calculation of lipid peroxidation activity. The reaction mix attains pink color and decrease in absorbance indicated the inhibition of adduct formation. The percentage of inhibition was calculated based on untreated cells and plotted against concentrations of compounds [42].
2.5. Nitric Oxide Synthesis
RAW 264.7 monocytes (1 × 106 cells/ml) were seeded in starving phenol-free minimum essential medium (MEM) containing 0.5% FBS and antibiotics overnight in 24-well plates. On the following day, the starving medium was replaced with fresh medium and cells were treated with increasing concentrations of compounds for 2 h followed by NO stimulation with LPS (20 ng/ml) for a total of 24 h. Supernatants were collected after centrifugation and used for analysis of nitrite and nitrate levels using the nitric oxide quantitation kit (Active Motif, Carlsbad, CA) [43].
2.6. NF-κB Activation
RAW 264.7 cells (3 × 106/5 ml) were incubated with increasing concentrations of HNK, MCP, and MCP : HNK (9 : 1) for 48 h and nuclear proteins were extracted with TransAM NF-κB p65 activity ELISA kit (Active Motif, Carlsbad, CA) per manufacturer's instructions. Protein concentration of the nuclear lysate was determined and lysate equivalent to 20 µg protein was analyzed for NF-κB activity using the TransAM NF-κB p65 activity kit. In the TransAM kit, the NF-κB consensus site (5′-GGGACTTTCC-3′) is immobilized on the ELISA plate and the active form of NF-κB contained in the nuclear extract will specifically bind to the nucleotide. The complex can be detected with NF-κB primary and secondary antibody reactions followed by substrate color reactions. The ELISA plates were read at 450 nm with a reference wavelength of 655 nm in a microplate reader. The decrease in the absorbance compared to untreated sample indicated the inhibition of NF-κB activity (%) which was plotted against concentrations of compounds [44].
2.7. Cyclooxygenase-II Activity
Cyclooxygenase (COX) activity assay kit from Abcam (Cambridge, MA) and COX-II enzyme from Sigma-Aldrich (St. Louis, MO) were used for analyzing the effect of MCH, HNK, and MCH:HNK (9 : 1) on COX-II activity. The assay kit uses a chemiluminescent substrate to detect the peroxidative activity of the COX-II enzyme. Ibuprofen (nonsteroidal anti-inflammatory drug, NSAID) is used as a positive control in the assay. After inhibition with increasing concentrations of MCP, HNK, or MCP : HNK (9 : 1), the residual activity of COX-II is measured by addition of a luminescent substrate and arachidonic acid. Light emission will start immediately and is directly proportional to the COX-II activity in the sample which is measured quickly by using a Veritas Luminometer (Turner Biosystems, Sunnyvale, CA) equipped with injectors for both substrate and arachidonic acid. The relative light units (RLU) recorded by the luminometer were used to calculate the percent inhibition of COX-II activity by MCH, HNK, or MCH:HNK (9 : 1) according to the following formula: percent inhibition (1 − average net inhibitor RLU/average net RLU for uninhibited) × 100. The inhibition percentage was plotted against PMF concentrations of compounds.
2.8. CompuSyn Analysis
To determine the synergistic/additive/antagonistic effects between MCP and HNK, data on various biochemical parameters were analyzed further using CompuSyn software (CompuSyn Inc., Paramus, NJ). This program is based on Chou and Talalay's multiple drug effect equations [45] and it defines synergism as a more-than-expected additive effect and antagonism as a less-than-expected additive effect. The combination index (CI) was calculated by the Chou-Talalay equations for multiple drug effects, which consider both potency (inhibitory concentration values) and shape (slope, m) of the dose-effect curve [45, 46].
2.9. Statistical Analysis
Mean and standard deviation estimates were calculated using Excel software. The data were analyzed statistically by paired t test (GraphPad Prism software, La Jolla, CA) and p values were used to determine the significant difference between treatment groups.
3. Results
3.1. Antioxidant Activity
The results of antioxidant assay presented in Figure 1 showed that both HNK and MCP induced a dose-dependent increase in the antioxidant activity, the latter showing a consistently higher response than the former. MCP : HNK (9 : 1) mix showed significantly higher antioxidant activity than the single agents alone, especially at 200 and 500 μg/ml concentrations.
Figure 1.
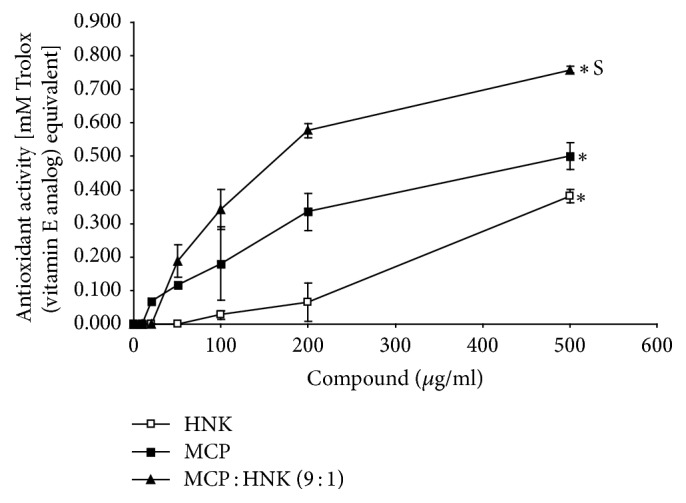
Antioxidant activity of HNK, MCP, and MCP : HNK (9 : 1). The activity equivalent to mM Trolox (vitamin E analog) was analyzed using Antioxidant Assay Kit (Sigma-Aldrich, MO) and plotted against compound concentrations. Antioxidant activity curves were statistically analyzed by paired t test; ∗p < 0.05 for HNK versus MCP, HNK versus MCP : HNK (9 : 1), and MCP versus MCP : HNK (9 : 1); S, synergism between MCP and HNK.
3.2. Inhibition of LPS-Induced TNF-α Production
When RAW 264.7 mouse monocyte cells were treated with LPS (20 ng/ml), proinflammatory cytokine TNF-α is induced (875.35 pg/ml) and secreted into the medium. Treatment of mouse monocytes with increasing concentrations of HNK inhibited the LPS-induced TNF-α synthesis significantly in a dose-dependent manner (Figure 2). As compared to HNK, MCP showed essentially no inhibitory effect on LPS-induced TNF-α synthesis by monocytes. However, the MCP : HNK (9 : 1) combination showed significantly better inhibitory effect than HNK alone. The highest dose of MCP : HNK (9 : 1) (5000 ug/ml) has almost completely inhibited the LPS-induced TNF-α synthesis in monocytes.
Figure 2.
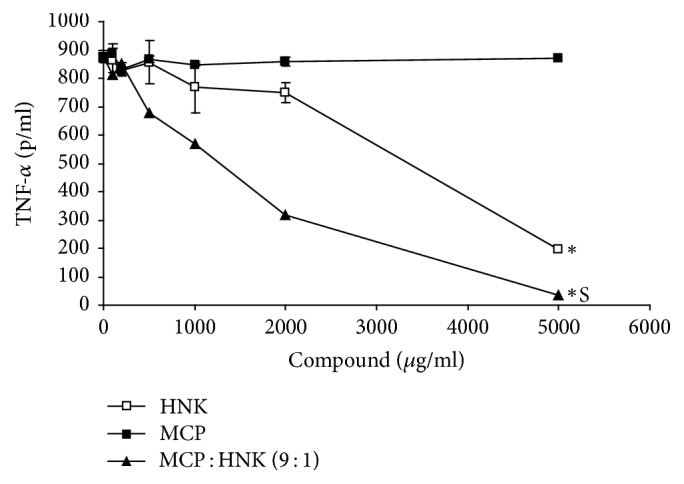
Inhibition of LPS-induced TNF-α (pg/ml) production by HNK, MCP, and MCP : HNK (9 : 1) in RAW 264.7 mouse monocyte cell line. The cells were treated with compounds and/or LPS in starvation medium and TNF-α analyzed by ELISA. Inhibition curves were analyzed by paired t test; ∗p < 0.05 for HNK versus MCP, and HNK versus MCP : HNK (9 : 1); S, synergism between MCP and HNK.
3.3. Inhibition of Lipid Peroxidation
The effect of HNK, MCP, and MCP : HNK (9 : 1) on H2O2-induced lipid peroxidation is shown in Figure 3. HNK has significant inhibitory effect on lipid peroxidation with 58% inhibition at 500 μg/ml concentration. Higher concentrations of HNK beyond 500 μg/ml dose do not inhibit the lipid peroxidation further. MCP and MCP : HNK (9 : 1) mix inhibit lipid peroxidation slightly with 20% reduction at 500 μg/ml dose. MCP demonstrated a steady inhibition of lipid peroxidation up to 200 μg/ml dose which remained stable with increase in dosage.
Figure 3.
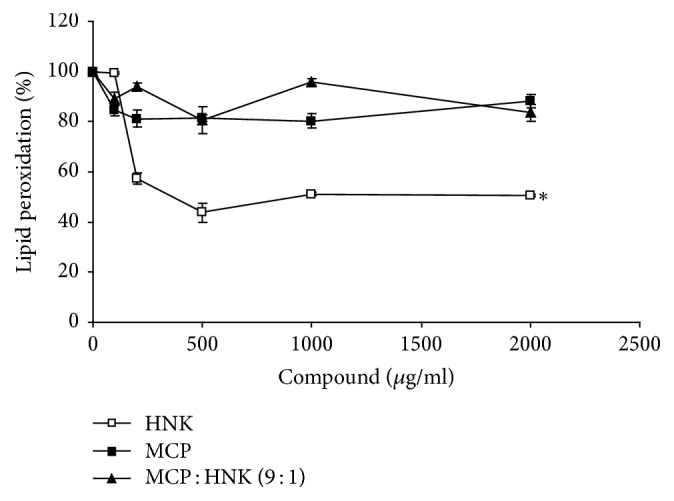
Inhibition of lipid peroxidation by HNK, MCP, and MCP : HNK (9 : 1) in RAW 264.7 mouse monocyte cell line. Lipid peroxidation was significantly inhibited by HNK treatment of monocytes. The treatment groups were compared using paired t test; ∗p < 0.05 for HNK versus MCP and HNK versus MCP : HNK (9 : 1). MCP and MCP : HNK (9 : 1) treatments were not statistically significant.
3.4. Inhibition of NF-κB Activity
The results of NF-κB activity assay presented in Figure 4 showed that HNK treatment of RAW 264.7 mouse monocytes has no significant inhibitory effect on NF-κB activity. However, MCP inhibited the NF-κB activity significantly with about 35% and 40% inhibition at 500 ug/ml and 2000 μg/ml doses, respectively. On the other hand, MCP : HNK (9 : 1) combination demonstrated the highest inhibitory effect at every concentration, with about 60% inhibition at 500 ug/ml and 2000 μg/ml doses.
Figure 4.
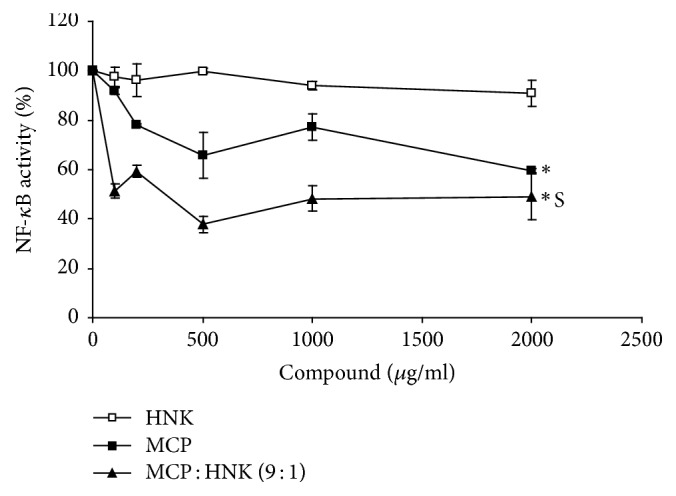
Inhibition of NF-κB (p65) activity by HNK, MCP, and MCP : HNK (9 : 1) in RAW 264.7 mouse monocyte cell line. NF-kB activity is inhibited significantly by MCP and MCP : HNK (9 : 1) treatment of monocytes and not by HNK alone. The treatment groups were compared using paired t test; ∗p < 0.05 for HNK versus MCP and HNK versus MCP : HNK (9 : 1). No significant difference was observed between MCP and MCP : HNK (9 : 1) treatment. S, synergism between MCP and HNK.
3.5. Inhibition of COX-II Activity
HNK, MCP, and MCP : HNK (9 : 1) inhibit COX-II activity (Figure 5). The inhibition level plateaued between 10 and 100 μg/ml and then further increased at a concentration >200 μg/ml in all treatment groups. MCP alone demonstrates greater inhibition profile than HNK and MCP : HNK (9 : 1) combination, with about 85% inhibition of COX-II activity at 200 μg/ml concentration.
Figure 5.
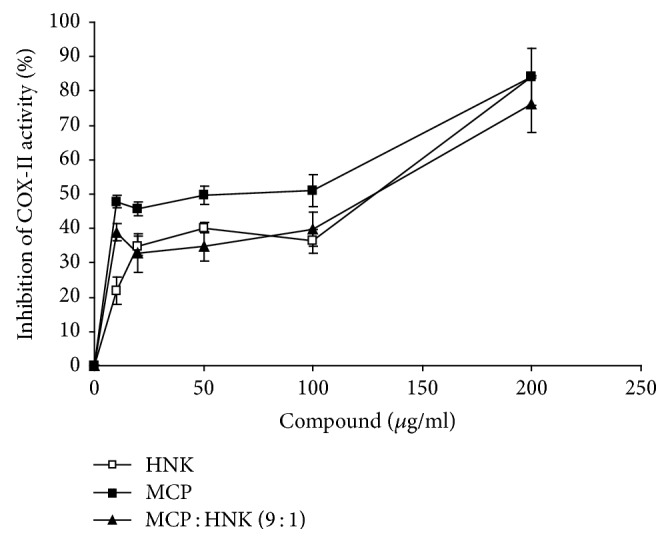
Inhibition of cyclooxygenase-II (COX-II) activity by HNK, MCP, and MCP : HNK (9 : 1) treatment. The inhibition curves were compared by paired t test; p < 0.05 for HNK versus MCP and MCP versus MCP : HNK (9 : 1). No significant difference was observed between MCP and MCP : HNK (9 : 1) treatment.
3.6. Effect of HNK, MCP, and MCP : HNK (9 : 1) on Nitric Oxide Synthesis
The concentration of nitrite and nitrate levels is an indication of nitric oxide synthesis. The effect of HNK, MCP, and MCP : HNK (9 : 1) on nitrite and nitrate concentration in RAW264.7 mouse monocytes is presented in Figures 6(a)–6(c). All three compounds failed to show any significant effect on nitrite synthesis. However, MCP and the MCP : HNK (9 : 1) mixture elevated the nitrate concentration in a dose-dependent manner, without any significant difference between the groups. In the case of HNK treated cells, nitrate level remained the same without any change and the data matched nitrite concentration. It is quite possible that nitrogen content in MCP may be contributing to the spike in nitrate levels.
Figure 6.

Effect of HNK, MCP, and MCP : HNK (9 : 1) on nitric oxide synthesis in RAW 264.7 mouse monocyte cell line. (a) Effect on nitrite concentration in cells treated with increasing concentrations of compounds. No significant difference was noticed among HNK, MCP, and MCP : HNK (9 : 1) treated monocytes. (b) Effect on nitrate concentration in cells treated with increasing concentrations of HNK, MCP, and MCP : HNK (9 : 1). Treatment groups were compared using paired t test; ∗p < 0.05 for HNK versus MCP and HNK versus MCP : HNK (9 : 1). No significant difference between MCP and MCP : HNK (9 : 1) treatment groups. (c) Effect on nitrite + nitrate concentration in cells treated with increasing concentrations of HNK, MCP, and MCP : HNK (9 : 1). Treatment groups were compared using paired t test; ∗p < 0.05 for HNK versus MCP and HNK versus MCP : HNK (9 : 1). No significant difference is noticed between MCP and MCP : HNK (9 : 1).
3.7. Synergism between MCP and HNK
Table 1 shows the dose-effect relationship between MCP and HNK for various biochemical parameters analyzed in the present investigation. The combination index (CI) estimates demonstrated synergism between MCP and HNK for inhibition of LPS-induced TNF-α because the CI values were below 1 at ED50, ED75, and ED90 levels. The drug reduction index (DRI) indicated the potential reduction of MCP and HNK amounts to achieve an ED50 effect. A synergistic effect between MCP and HNK was also observed for antioxidant activity with CI values between 0.3 and 0.7 (synergism) at ED50, ED75, and ED90 levels. The DRI values for antioxidant effect indicated the dose reduction possible by combining MCP and HNK. For lipid peroxidation, MCP and HNK combination has additive effect at an ED50 level and low level of antagonism at ED75 and ED90 levels. CompuSyn analysis also showed strong level of synergism between MCP and HNK for inhibition of NF-κB activity.
Table 1.
Dose-effect relationship between modified citrus pectin (MCP) and honokiol (HNK).
| Biochemical traits | CI at ED50 | CI at ED75 | CI at ED90 | DRI-MCP ED50 | DRI-HNK ED50 |
|---|---|---|---|---|---|
| Inhibition of LPS-induced TNF-α | 0.549 | 0.515 | 0.510 | 1.825 | 706.268 |
| Antioxidant activity | 0.670 | 0.546 | 0.446 | 33.682 | 1.562 |
| Inhibition of lipid peroxidation | 1.070 | 1.765 | 1.189 | — | — |
| Nitric oxide synthesis (NO2 + NO3) | 2.932 | 4.675 | 5.684 | — | — |
| Inhibition of NFκB activity | 0.068 | 0.131 | 0.254 | 14.749 | 917.283 |
| Inhibition of COX-II activity | 2.248 | 3.080 | 1.870 | — | — |
CI, combination index; DRI, drug reduction index; ED, effective dose; CI values by CompuSyn analysis: <0.1: very strong synergism; 0.1–0.3: strong synergism; 0.3–0.7: synergism; 0.7–0.85: moderate synergism; 0.85–0.90: slight synergism; 0.90–1.10: nearly additive; 1.10–1.20: slight antagonism; 1.20–1.45: moderate antagonism; 1.45–3.3: antagonism; 3.3–10: strong antagonism; >10: very strong antagonism.
4. Discussion
Overproduction of inflammatory mediators such as NF-κB, TNF-α, COX-II, and NO produced by macrophages, neutrophils, and other immune cells is very much involved in the pathogenesis of chronic diseases, for example, atherosclerosis, arthritis, type 2 diabetes, and cancer [47–49]. Therefore, identification of natural product-derived extracts or compounds that controls the production of inflammatory mediators is an extremely attractive therapeutic or preventative modality for these diseases [50]. In the present investigation, both HNK and MCP treatment have enhanced the antioxidant activity in monocytes. Furthermore, by using CompuSyn analysis we showed that the combination of MCP and HNK has synergistic effect on the antioxidant activity of monocytes. The CI values of the MCP : HNK (9 : 1) combination for antioxidant activity are <1 at ED50, ED75, or ED90 levels, thereby indicating the strong synergism between MCP and HNK for antioxidant activity. Such synergistic effects for antioxidant activity have been observed for combinations of extracts from spices (anise, cardamom, clove, and cinnamon) [51] and also between black tea and curcumin [52], which may form the basis for a therapeutic that has superior effects compared to single agents.
The dysregulation of the redox system is implicated in numerous human disorders including neoplasia, inflammation, degenerative diseases, and environmental exposures [53, 54]. HNK has been shown to reduce intracellular super oxide 2-fold and therefore free radical scavenging activity is implicated as an important attribute of HNK [11]. Further, HNK inhibits copper-induced oxidative modification of LDL [12] and iron-stimulated lipid peroxidation [13]. It is also known that oxygen overproduction stimulates lipid peroxidation and the present investigation showed that HNK inhibits lipid peroxidation which may involve both initiation and propagation of peroxidation process [12].
NF-κB regulates the transcription of several genes, including iNOS, COX-II, TNF-α, and IL-6 and is thus important for the development of inflammation-associated diseases [1, 50, 55]. NF-κB is primarily composed of proteins with molecular mass of 50 (p50) and 65 kDa (p65) and is retained in the cytoplasm by inhibitor of κB (I-κB). From its unstimulated form, NF-κB is activated by a wide variety of inflammatory stimuli like LPS. Most of these inflammatory mediators induce the phosphorylation-dependent degradation of I-κB proteins, allowing active NF-κB to translocate into the nucleus, where it regulates several genes including iNOS, COX-II, TNF-α, and IL-6 [55]. In the present investigation, we have shown that both MCP and MCP : HNK (9 : 1) combination have significant inhibitory effect on NF-κB activity whereas HNK alone showed little inhibition of NF-κB activity. Furthermore, very strong synergism between MCP and HNK for inhibition of NF-κB activity and also the DRI values (Table 1) may facilitate the reduction of MCP and HNK amounts in the combination to achieve the defined therapeutic effect. The inhibition profile of TNF-α by these compounds also follows a similar pattern as NF-κB activity. HNK demonstrated the strongest inhibitory effect whereas MCP failed to show any inhibition of TNF-α unlike NF-κB. However, CompuSyn analysis showed the synergism between MCP and HNK for TNF-α -inhibition with CI values of <1 at ED50, ED75, and ED90 levels which were similar to NF-κB inhibition. Furthermore, all three compounds including the MCP : HNK (9 : 1) combination inhibited COX-II activity. The MCP : HNK combination does show an additive effect for inhibition of lipid peroxidation at ED50 level. In summary, MCP : HNK combination appears to be superior to individual agents due to the synergism for enhanced antioxidant activity, inhibition of NF-κB activity, and LPS-induced TNF-α synthesis.
5. Conclusion
The current investigation demonstrated the antioxidant and anti-inflammatory effects of MCP and HNK and the MCP : HNK in vitro. The MCP : HNK combination in general demonstrated significantly better effects than the single compounds due to the synergistic effects on antioxidant activity, inhibition of LPS-induced TNF-alpha production, and NF-κB activity. This demonstration of synergistic antioxidant and anti-inflammatory properties of MCP and HNK suggests a potential application for the amelioration of effects due to oxidative stress and free radical damage.
Acknowledgments
This study was funded by a research grant from ecoNugenics, Inc.
Abbreviations
- ABTS:
2,2′-azino-bis(3-ethylbenzthiazoline-6-sulfonic acid)
- CI:
Combination index
- COX-II:
Cyclooxygenase-II
- DRI:
Drug reduction index
- ELISA:
Enzyme linked immunosorbent assay
- ED:
Effective dose
- Gal-3:
Galectin-3
- H2O2:
Hydrogen peroxide
- HNK:
Honokiol
- I-κB:
Inhibitor of κB
- IL-1:
Interleukin-1
- IL-6:
Interleukin-6
- iNOS:
Inducible nitric oxide synthase
- LDL:
Low-density lipoprotein
- LPS:
Lipopolysaccharide
- MCP:
Modified citrus pectin
- MDA:
Malondialdehyde
- MEM:
Minimum essential medium
- NF-κB:
Nuclear factor-kappa B
- NSAID:
Nonsteroidal anti-inflammatory drug
- PBS:
Phosphate buffered saline
- TBA:
Thiobarbituric acid.
Ethical Approval
This investigation did not involve human subjects or animal species and therefore approvals from the institutional committees were not required. The investigation was performed adhering to the rules and regulations.
Conflicts of Interest
The author Barry Wilk discloses employment at a dietary supplement company. Isaac Eliaz discloses ownership of a dietary supplement company. Cheppail Ramachandran and Steven J. Melnick do not have any conflicts of interest in the research presented.
Authors' Contributions
Cheppail Ramachandran, Barry Wilk, Steven J. Melnick, and Isaac Eliaz were responsible for study design, experimentation, and manuscript preparation.
References
- 1.Lawrence T. The nuclear factor NF-κB pathway in inflammation. Cold Spring Harbor perspectives in biology. 2009;1(6) doi: 10.1101/cshperspect.a001651.a001651 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Kanterman J., Sade-Feldman M., Baniyash M. New insights into chronic inflammation-induced immunosuppression. Seminars in Cancer Biology. 2012;22(4):307–318. doi: 10.1016/j.semcancer.2012.02.008. [DOI] [PubMed] [Google Scholar]
- 3.Hseu Y.-C., Wu F.-Y., Wu J.-J., et al. Anti-inflammatory potential of Antrodia Camphorata through inhibition of iNOS, COX-2 and cytokines via the NF-κB pathway. International Immunopharmacology. 2005;5(13):1914–1925. doi: 10.1016/j.intimp.2005.06.013. [DOI] [PubMed] [Google Scholar]
- 4.Arora S., Singh S., Piazza G. A., Contreras C. M., Panyam J., Singh A. P. Honokiol: a novel natural agent for cancer prevention and therapy. Current Molecular Medicine. 2012;12(10):1244–1252. doi: 10.2174/156652412803833508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Zhang P., Liu X., Zhu Y., Chen S., Zhou D., Wang Y. Honokiol inhibits the inflammatory reaction during cerebral ischemia reperfusion by suppressing NF-κB activation and cytokine production of glial cells. Neuroscience Letters. 2013;534(1):123–127. doi: 10.1016/j.neulet.2012.11.052. [DOI] [PubMed] [Google Scholar]
- 6.Sulakhiya K., Kumar P., Gurjar S. S., Barua C. C., Hazarika N. K. Beneficial effect of honokiol on lipopolysaccharide induced anxiety-like behavior and liver damage in mice. Pharmacology Biochemistry and Behavior. 2015;132:79–87. doi: 10.1016/j.pbb.2015.02.015. [DOI] [PubMed] [Google Scholar]
- 7.Sulakhiya K., Kumar P., Jangra A., et al. Honokiol abrogates lipopolysaccharide-induced depressive like behavior by impeding neuroinflammation and oxido-nitrosative stress in mice. European Journal of Pharmacology. 2015;744:124–131. doi: 10.1016/j.ejphar.2014.09.049. [DOI] [PubMed] [Google Scholar]
- 8.Shen J., Man K., Huang P., et al. Honokiol and magnolol as multifunctional antioxidative molecules for dermatologic disorders. Molecules. 2010;15(9):6452–6465. doi: 10.3390/molecules15096452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Woodbury A., Yu S. P., Wei L., García P. Neuro-modulating effects of honokiol: A review. Frontiers in Neurology. 2013;4 doi: 10.3389/fneur.2013.00130.Article 130 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Zhang G.-S., Wang R.-J., Zhang H.-N., Zhang G.-P., Luo M.-S., Luo J.-D. Effects of chronic treatment with honokiol in spontaneously hypertensive rats. Biological and Pharmaceutical Bulletin. 2010;33(3):427–431. doi: 10.1248/bpb.33.427. [DOI] [PubMed] [Google Scholar]
- 11.Dikalov S., Losik T., Arbiser J. L. Honokiol is a potent scavenger of superoxide and peroxyl radicals. Biochemical Pharmacology. 2008;76(5):589–596. doi: 10.1016/j.bcp.2008.06.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Chiu J.-H., Ho C.-T., Wei Y.-H., Lui W.-Y., Hong C.-Y. In vitro and in vivo protective effect of honokiol on rat liver from peroxidative injury. Life Sciences. 1997;61(19):1961–1971. doi: 10.1016/S0024-3205(97)00836-9. [DOI] [PubMed] [Google Scholar]
- 13.Ou H.-C., Chou F.-P., Lin T.-M., Yang C.-H., Sheu W. H.-H. Protective effects of honokiol against oxidized LDL-induced cytotoxicity and adhesion molecule expression in endothelial cells. Chemico-Biological Interactions. 2006;161(1):1–13. doi: 10.1016/j.cbi.2006.02.006. [DOI] [PubMed] [Google Scholar]
- 14.Ramachandran C., Wilk B. J., Hotchkiss A., Chau H., Eliaz I., Melnick S. J. Activation of Human T-Helper/Inducer Cell, T-Cytotoxic Cell, B-Cell, and Natural Killer (NK)-Cells and induction of Natural Killer Cell Activity against K562 Chronic Myeloid Leukemia Cells with Modified Citrus Pectin. BMC Complementary and Alternative Medicine. 2011;11, article no. 59 doi: 10.1186/1472-6882-11-59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Hossein G., Keshavarz M., Ahmadi S., Naderi N. Synergistic effects of PectaSol-C modified citrus pectin an inhibitor of galectin-3 and paclitaxel on apoptosis of human SKOV-3 ovarian cancer cells. Asian Pacific Journal of Cancer Prevention. 2013;14(12):7561–7568. doi: 10.7314/APJCP.2013.14.12.7561. [DOI] [PubMed] [Google Scholar]
- 16.Tehranian N., Sepehri H., Mehdipour P., et al. Combination effect of pectasol and Doxorubicin on viability, cell cycle arrest and apoptosis in DU-145 and lncap prostate cancer cell lines. Cell Biology International. 2012;36(7):601–610. doi: 10.1042/CBI20110309. [DOI] [PubMed] [Google Scholar]
- 17.Jiang J., Eliaz I., Sliva D. Synergistic and additive effects of modified citrus pectin with two polybotanical compounds, in the suppression of invasive behavior of human breast and prostate cancer cells. Integrative Cancer Therapies. 2013;12(2):145–152. doi: 10.1177/1534735412442369. [DOI] [PubMed] [Google Scholar]
- 18.Yan J., Katz A. PectaSol-C modified citrus pectin induces apoptosis and inhibition of proliferation in human and mouse androgen-dependent and-independent prostate cancer cells. Integrative Cancer Therapies. 2010;9(2):197–203. doi: 10.1177/1534735410369672. [DOI] [PubMed] [Google Scholar]
- 19.Liu H.-Y., Huang Z.-L., Yang G.-H., Lu W.-Q., Yu N.-R. Inhibitory effect of modified citrus pectin on liver metastases in a mouse colon cancer model. World Journal of Gastroenterology. 2008;14(48):7386–7391. doi: 10.3748/wjg.14.7386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Azémar M., Hildenbrand B., Haering B., Heim M. E., Unger C. Clinical benefit in patients with advanced solid tumors treated with modified citrus pectin: a prospective pilot study. Clinical Medicine Insights: Oncology. 2007;1:73–80. [Google Scholar]
- 21.Guess B. W., Scholz M. C., Strum S. B., Lam R. Y., Johnson H. J., Jennrich R. I. Modified citrus pectin (MCP) increases the prostate-specific antigen doubling time in men with prostate cancer: A phase II pilot study. Prostate Cancer and Prostatic Diseases. 2003;6(4):301–304. doi: 10.1038/sj.pcan.4500679. [DOI] [PubMed] [Google Scholar]
- 22.Zhao Z. Y., Liang L., Fan X., Yu Z., et al. The role of modified citrus pectin as an effective chelator of lead in children hospitalized with toxic lead levels. Alternative Therapies in Health and Medicine. 2008;14(4):34–38. [PubMed] [Google Scholar]
- 23.Eliaz I., Weil E., Wilk B. Integrative medicine and the role of modified citrus pectin/alginates in heavy metal chelation and detoxification - Five case reports. Forschende Komplementarmedizin. 2007;14(6):358–364. doi: 10.1159/000109829. [DOI] [PubMed] [Google Scholar]
- 24.Eliaz I., Hotchkiss A. T., Fishman M. L., Rode D. The effect of modified citrus pectin on urinary excretion of toxic elements. Phytotherapy Research. 2006;20(10):859–864. doi: 10.1002/ptr.1953. [DOI] [PubMed] [Google Scholar]
- 25.Martinez-Martinez E., Ibarrola J., Calvier L., et al. Galectin-3 blockade reduces renal fibrosis in two normotensive experimental models of renal damage. PLoS One. 2016;11(11):p. e0166272. doi: 10.1371/journal.pone.0166272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Abu-Elsaad N. M., Elkashef W. F. Modified citrus pectin stops progression of liver fibrosis by inhibiting galectin-3 and inducing apoptosis of stellate cells. Canadian Journal of Physiology and Pharmacology. 2016;94(5):554–562. doi: 10.1139/cjpp-2015-0284. [DOI] [PubMed] [Google Scholar]
- 27.Martínez-Martínez E., Calvier L., Rossignol P., et al. Galectin-3 inhibition prevents adipose tissue remodelling in obesity. International Journal of Obesity. 2016;40(6):1034–1038. doi: 10.1038/ijo.2016.19. [DOI] [PubMed] [Google Scholar]
- 28.Vergaro G., Prud'Homme M., Fazal L., et al. Inhibition of Galectin-3 Pathway Prevents Isoproterenol-Induced Left Ventricular Dysfunction and Fibrosis in Mice. Hypertension. 2016;67(3):606–612. doi: 10.1161/HYPERTENSIONAHA.115.06161. [DOI] [PubMed] [Google Scholar]
- 29.Martínez-Martínez E., López-Ándres N., Jurado-López R., et al. Galectin-3 participates in cardiovascular remodeling associated with obesity. Hypertension. 2015;66(5):961–969. doi: 10.1161/HYPERTENSIONAHA.115.06032. [DOI] [PubMed] [Google Scholar]
- 30.Martínez-Martínez E., Calvier L., Fernández-Celis A., et al. Galectin-3 blockade inhibits cardiac inflammation and fibrosis in experimental hyperaldosteronism and hypertension. Hypertension. 2015;66(4):767–775. doi: 10.1161/HYPERTENSIONAHA.115.05876. [DOI] [PubMed] [Google Scholar]
- 31.Calvier L., Martinez-Martinez E., Miana M., et al. The impact of galectin-3 inhibition on aldosterone-induced cardiac and renal injuries. JACC: Heart Failure. 2015;3(1):59–67. doi: 10.1016/j.jchf.2014.08.002. [DOI] [PubMed] [Google Scholar]
- 32.Calvier L., Miana M., Reboul P., et al. Galectin-3 mediates aldosterone-induced vascular fibrosis. Arteriosclerosis, Thrombosis, and Vascular Biology. 2013;33(1):67–75. doi: 10.1161/ATVBAHA.112.300569. [DOI] [PubMed] [Google Scholar]
- 33.Kolatsi-Joannou M., Price K. L., Winyard P. J., Long D. A. Modified citrus pectin reduces galectin-3 expression and disease severity in experimental acute kidney injury. PLoS ONE. 2011;6(4) doi: 10.1371/journal.pone.0018683.e18683 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.de Boer R. A., van Veldhuisen D. J., Gansevoort R. T., et al. The fibrosis marker galectin-3 and outcome in the general population. Journal of Internal Medicine. 2012;272(1):55–64. doi: 10.1111/j.1365-2796.2011.02476.x. [DOI] [PubMed] [Google Scholar]
- 35.Lala R. I., Puschita M., Darabantiu D., Pilat L. Galectin-3 in heart failure pathology – “another brick in the wall”? Acta Cardiologica. 2015;70(3):323–331. doi: 10.2143/AC.70.3.3080637. doi: 10.2143/AC.70.3.3080637. [DOI] [PubMed] [Google Scholar]
- 36.de Boer R. A., Voors A. A., Muntendam P., van Gilst W. H., van Veldhuisen D. J. Galectin-3: a novel mediator of heart failure development and progression. European Journal of Heart Failure. 2009;11(9):811–817. doi: 10.1093/eurjhf/hfp097. [DOI] [PubMed] [Google Scholar]
- 37.Ohshima S., Kuchen S., Seemayer C. A., et al. Galectin 3 and its binding protein in rheumatoid arthritis. Arthritis & Rheumatology. 2003;48(10):2788–2795. doi: 10.1002/art.11287. [DOI] [PubMed] [Google Scholar]
- 38.Baeck C., Tacke F. Balance of inflammatory pathways and interplay of immune cells in the liver during homeostasis and injury. EXCLI Journal Experimental and Clinical Sciences. 2014;13:67–81. [PMC free article] [PubMed] [Google Scholar]
- 39.Osborn L., Kunkel S., Nabel G. J. Tumor necrosis factor-alpha and interleukin 1stimulate the human immunodeficiency virus enhancer by activation of the nuclear factor κB. Proceedings of the National Academy of Sciences USA. 1989;86(7):2336–2340. doi: 10.1073/pnas.86.7.2336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Beg A. A., Finco T. S., Nantermet P. V., Baldwin A. S., Jr. Tumor necrosis factor and interleukin-1 lead to phosphorylation and loss of IκBα: a mechanism for NF-κB activation. Molecular and Cellular Biology. 1993;13(6):3301–3310. doi: 10.1128/MCB.13.6.3301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Baima E. T., Guzova J. A., Mathialagan S., et al. Novel insights into the cellular mechanisms of the anti-inflammatory effects of NF-κB essential modulator binding domain peptides. The Journal of Biological Chemistry. 2010;285(18):13498–13506. doi: 10.1074/jbc.M109.099895. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Ramachandran C., Quirin K.-W., Escalon E., Melnick S. J. Improved neuroprotective effects by combining Bacopa monnieri and Rosmarinus officinalis supercritical CO2 extracts. Journal of Evidence-Based Complementary and Alternative Medicine. 2014;19(2):119–127. doi: 10.1177/2156587214524577. [DOI] [PubMed] [Google Scholar]
- 43.Aviram A., Tsoukias N. M., Melnick S. J., Resek A. P., Ramachandran C. Inhibition of nitric oxide synthesis in mouse macrophage cells by feverfew supercritical extract. Phytotherapy Research. 2012;26(4):541–545. doi: 10.1002/ptr.3594. [DOI] [PubMed] [Google Scholar]
- 44.Nair P. K. R., Melnick S. J., Ramachandran R., Escalon E., Ramachandran C. Mechanism of macrophage activation by (1,4)-α-d-glucan isolated from Tinospora cordifolia. International Immunopharmacology. 2006;6(12):1815–1824. doi: 10.1016/j.intimp.2006.07.028. [DOI] [PubMed] [Google Scholar]
- 45.Chou T. C., Talalay P. Analysis of combined drug effects: a new look at a very old problem. Trends in Pharmacological Sciences. 1983;4:450–454. doi: 10.1016/0165-6147(83)90490-X. [DOI] [Google Scholar]
- 46.Kapadia G. J., Rao G. S., Ramachandran C., Iida A., Suzuki N., Tokuda H. Synergistic cytotoxicity of red beetroot (Beta vulgaris L.) extract with doxorubicin in human pancreatic, breast and prostate cancer cell lines. Journal of complementary & integrative medicine. 2013;10 doi: 10.1515/jcim-2013-0007. [DOI] [PubMed] [Google Scholar]
- 47.Kumar S., Pandey A. K. Free radicals: health implications and their mitigation by herbals. British Journal of Medicine and Medical Research. 2015;7(6):438–457. doi: 10.9734/bjmmr/2015/16284. [DOI] [Google Scholar]
- 48.Poyton R. O., Ball K. A., Castello P. R. Mitochondrial generation of free radicals and hypoxic signaling. Trends in Endocrinology and Metabolism. 2009;20(7):332–340. doi: 10.1016/j.tem.2009.04.001. [DOI] [PubMed] [Google Scholar]
- 49.Libby P. Inflammatory mechanisms: the molecular basis of inflammation and disease. Nutrition Reviews. 2007;65(3):S140–S146. doi: 10.1111/j.1753-4887.2007.tb00352.x. [DOI] [PubMed] [Google Scholar]
- 50.Prasad S., Phromnoi K., Yadav V. R., Chaturvedi M. M., Aggarwal B. B. Targeting inflammatory pathways by flavonoids for prevention and treatment of cancer. Planta Medica. 2010;76(11):1044–1063. doi: 10.1055/s-0030-1250111. [DOI] [PubMed] [Google Scholar]
- 51.Patra K., Jana S., Mandal D. P., Bhattacharjee S. Evaluation of the antioxidant activity of extracts and active principles of commonly consumed Indian spices. Journal of Environmental Pathology, Toxicology and Oncology. 2016;35(4):299–315. doi: 10.1615/JEnvironPatholToxicolOncol.2016016387. [DOI] [PubMed] [Google Scholar]
- 52.Alm-Eldeen A. A., Mona M. H., Shati A. A., El-Mekkawy H. I. Synergistic effect of black tea and curcumin in improving the hepatotoxicity induced by aflatoxin B1 in rats. Toxicology and Industrial Health. 2015;31(12):1269–1280. doi: 10.1177/0748233713491807. [DOI] [PubMed] [Google Scholar]
- 53.Barnes P. J., Karin M. Nuclear factor-κB—a pivotal transcription factor in chronic inflammatory diseases. The New England Journal of Medicine. 1997;336(15):1066–1071. doi: 10.1056/NEJM199704103361506. [DOI] [PubMed] [Google Scholar]
- 54.Bickers D. R., Athar M. Oxidative stress in the pathogenesis of skin disease. Journal of Investigative Dermatology. 2006;126(12):2565–2575. doi: 10.1038/sj.jid.5700340. [DOI] [PubMed] [Google Scholar]
- 55.Baldwin A. S., Jr. The NF-κB and IκB proteins: new discoveries and insights. Annual Review of Immunology. 1996;14:649–683. doi: 10.1146/annurev.immunol.14.1.649. [DOI] [PubMed] [Google Scholar]