

Abstract
In higher plant thylakoids, the heterogeneous distribution of photosynthetic protein complexes is a determinant for the formation of grana, stacks of membrane discs that are densely populated with Photosystem II (PSII) and its light harvesting complex (LHCII). PSII associates with LHCII to form the PSII-LHCII supercomplex, a crucial component for solar energy conversion. Here, we report a biochemical, structural and functional characterization of pairs of PSII-LHCII supercomplexes, which were isolated under physiologically-relevant cation concentrations. Using single-particle cryo-electron microscopy, we determined the three-dimensional structure of paired C2S2M PSII-LHCII supercomplexes at 14 Å resolution. The two supercomplexes interact on their stromal sides through a specific overlap between apposing LHCII trimers and via physical connections that span the stromal gap, one of which is likely formed by interactions between the N-terminal loops of two Lhcb4 monomeric LHCII subunits. Fast chlorophyll fluorescence induction analysis showed that paired PSII-LHCII supercomplexes are energetically coupled. Molecular dynamics simulations revealed that additional flexible physical connections may form between the apposing LHCII trimers of paired PSII-LHCII supercomplexes in appressed thylakoid membranes. Our findings provide new insights into how interactions between pairs of PSII-LHCII supercomplexes can link adjacent thylakoids to mediate the stacking of grana membranes.
Introduction
Photosystem II (PSII) is a multisubunit pigment-protein complex that is embedded in the thylakoid membranes of oxygenic photosynthetic organisms. It uses solar energy to catalyse the splitting of water into dioxygen, protons and electrons, thus providing the molecular oxygen and chemical energy that sustain most of the life on Earth1.
PSII forms supercomplexes that are composed of two moieties: a central PSII core containing the catalytic reaction center and a peripheral antenna system responsible for harvesting light and transferring energy to the reaction center. The PSII core, occurring mainly in dimeric form2–4, is highly conserved among all oxygenic photosynthetic organisms, with the exception of the extrinsic proteins of the oxygen evolving complex (PsbO, PsbV and PsbU in cyanobacteria and diatoms; PsbO, PsbP and PsbQ in plants and green algae)5. Conversely, the peripheral antenna system is evolutionarily divergent; it is composed of extrinsic phycobilisomes in cyanobacteria, while it consists of intrinsic LHCII light harvesting complexes in green algae and plants5, 6. The evolution of different antenna systems was accompanied by the differentiation of thylakoid membranes into functionally distinct appressed (stacked) and non-appressed (unstacked) regions, which in higher plants are called grana and stroma lamellae, respectively7.
Two related processes are believed to govern grana stacking: (i) interactions between adjacent membrane surfaces8 and (ii) lateral segregation of photosystems within the thylakoid membrane plane9, 10. At neutral pH, thylakoid membranes carry a net negative charge, and the maintenance of the grana stacks requires shielding cations (e.g., Mg2+, K+ and Na+)11, 12. Indeed, it has long been known that exposure of thylakoid membranes to low-ionic strength buffers induces grana unstacking, and the readdition of low concentrations of divalent cations (≥ 5 mM Mg2+) or higher concentrations of monovalent cations (≥ 150 mM K+ or Na+) results in spontaneous restacking11. Photosynthetic membrane proteins are segregated into different thylakoid domains; grana regions contain mainly PSII and LHCII, complexes with flat stromal surfaces that do not project into the narrow stromal gap between thylakoids. Conversely, stroma lamellae and grana end membranes accommodate Photosystem I (PSI), with its light harvesting antenna complex (LHCI), and ATP synthase (ATP-ase), which have bulky stromal protrusions. Unlike the photosystems, cytochrome b 6 /f (Cyt b 6 /f) is evenly distributed between stacked and unstacked thylakoid regions13, 14. This lateral heterogeneity between PSI and PSII is thought to be intimately linked to membrane appression, allowing higher plants to optimize photosynthesis in ever-changing light conditions15.
Grana formation seems to be driven by interactions between the stromal domains of LHCIIs that face each other from adjacent stacked membranes16, 17. However, grana with apparently normal stacked architecture form in Arabidopsis mutants that are virtually devoid of LHCII18, suggesting that other mechanisms are involved in grana formation19. In addition, it has been proposed that the ordered assembly of semi-crystalline 2D arrays of PSII-LHCII supercomplexes in grana thylakoids might promote grana stacking by mediating specific contacts between adjacent membrane discs20. While the lateral heterogeneity of thylakoid membrane proteins appears to drive grana staking, there seems to be a reciprocal relationship where thylakoid architecture enforces this lateral heterogeneity; the unstacking of grana with low-ionic strength buffer causes the random redistribution of photosystems within the lateral thylakoid membrane plane as well as the detachment of some LHCII from supercomplexes21, 22.
In plant grana thylakoids, the PSII core associates with a variable number of LHCII antennas to form different types of PSII-LHCII supercomplexes14, 23, 24. The C2S2 supercomplex consists of a dimeric PSII core (C2), which strongly binds two LHCII trimers (S-trimers) via two copies of the monomeric Lhcb4 and Lhcb5 subunits. Larger C2S2M1-2 supercomplexes contain two copies of monomeric Lhcb6, with one or two additional LHCII trimers (M-trimers) bound with moderate strength to the dimeric PSII core via Lhcb4 and Lhcb614.
Two 3D maps of the C2S2 PSII-LHCII supercomplex isolated from spinach have been obtained by single-particle cryo-electron microscopy (cryo-EM); the first structure was generated 17 years ago at 17 Å resolution25, while the second structure was recently solved at 3.2 Å resolution26. The new structure’s significant improvement in resolution allowed the precise positioning of the pigments bound to the PSII-LHCII supercomplex. Thus, there is now a clear structural description for the energy transfer from the peripheral LHCII antennas to the PSII reaction center. The next step for the mechanistic understanding of plant photosynthesis is to expand this structural description to the organization and coordination of multiple PSII-LHCII supercomplexes within the grana stack. Here, we pursue this goal with a structural and functional description of paired C2S2M PSII-LHCII supercomplexes that can link adjacent grana membranes.
Results and Discussion
Isolation of paired PSII-LHCII supercomplexes
The differentiation of thylakoid membranes into grana and stroma lamellae is a ubiquitous feature of higher plant chloroplasts7. Even though the number and diameter of discs within grana stacks may vary depending on fluctuating environmental conditions27–29, different plant species grown under the same illumination regime can differ intrinsically in their grana organization30. We used pea plants as starting material for PSII-LHCII supercomplex isolation because their chloroplasts contain large, highly stacked grana30.
In order to investigate how PSII-LHCII supercomplexes interact with each other within grana stacks, thylakoids were isolated from pea leaves in the presence of divalent cation concentrations (i.e., 5 mM Mg2+) that resemble the native chloroplast ionic environment31, preserving the stacked morphology of the grana membranes. PSII-LHCII supercomplexes were subsequently isolated by a quick (1 min) direct solubilization of stacked thylakoids with the mild detergent n-dodecyl-α-D-maltoside (α-DDM) followed by sucrose density gradient ultracentrifugation in the presence of the same concentration of divalent cations, according to an optimized protocol previously described32. Physiological cation concentrations were maintained throughout the entire purification, and either Ca2+ or Mg2+ cations in the sucrose gradient buffer produced the same isolation profile (data not shown). It is important to point out that our isolation procedure greatly differs from that used by Wei et al.26 to produce the recent cryo-EM structure of the unpaired C2S2 plant supercomplex. In their study, grana membranes were first isolated by solubilizing thylakoids with Triton X-100 and then unstacked by washing with a metal chelating agent (i.e., EDTA), followed by further solubilization with α-DDM and ultracentrifugation. The buffers were devoid of salt throughout the purification, and thus the resulting structure lacks the native interactions between supercomplexes found within stacked grana.
Cryo-EM structure of paired C2S2M supercomplexes at 14 Å resolution
To investigate the structure of the isolated PSII-LHCII supercomplexes, the detergent-solubilized PSII-LHCII particles were frozen in vitreous ice and then imaged by transmission electron microscopy using a direct electron detector. From 6,834 micrographs collected (Supplementary Fig. 1a), 33,729 particles were manually picked for further data processing. After several image sorting steps and iterative rounds of 2D classification (Supplementary Fig. 1b), 6,776 particles corresponding to contaminants and blurred or broken complexes were discarded. The initial model (Supplementary Fig. 1c), built from 44 representative 2D classes, was used for the 3D classification of 26,953 particles, resulting in three classes: paired C2S2M supercomplexes (~53%), unpaired C2S2M supercomplexes (~21%) and paired C2S2 supercomplexes (~26%) (Supplementary Fig. 1d). From this classification, the majority of the isolated particles were found to be in a paired conformation (~79%), with the C2S2M form (~72%) more abundant than the C2S2 (~28%). Most particles contained LHCII M-trimers (~74%), irrespective of their paired or unpaired behaviour. This finding agrees with previous results showing that C2S2M is the predominant supercomplex in the thylakoid membranes of pea plants grown in moderate light intensity33, the same light condition used in this study.
Paired C2S2M particles, representing the most abundant 3D class and the largest isolated supercomplexes, were further analysed (Supplementary Fig. 1e,f). Several refinement cycles led to a final 3D cryo-EM map at an overall resolution of 14 Å (Fig. 1a–d), as judged by the “gold standard” Fourier shell correlation (FSC = 0.143) criterion34 (Supplementary Fig. 2). The 3D map depicts two C2S2M supercomplexes with their stromal surfaces facing each other. The supercomplexes do not completely overlap, but rather have a rotational offset of ~35° around the membrane plane’s normal vector (Fig. 1d). The 3D map of each C2S2M supercomplex, including the detergent shell, is 280 Å along its long-axis in the membrane plane, 200 Å along the M-trimer side and 160 Å along the S-trimer side (Fig. 1e). The thickness of the supercomplex is ~50 Å at its periphery, while the central protrusions extend for another ~50 Å on the luminal side (Fig. 1a). The central regions of the paired supercomplexes are connected on their stromal sides by two distinct densities that we have termed the “hinge” and “knot” as a description of their shapes (Fig. 1e,f). The ~20 Å stromal distance between the paired supercomplexes is consistent with reported stromal gap values for grana thylakoids, which range from 2 nm14 to 3.2–3.6 nm20, 35. In total, the supercomplex pair has a thickness of 220 Å, which is compatible with the in situ stacking periodicity measured for intact pea chloroplasts20.
Figure 1.

Three-dimensional cryo-EM density map of paired C2S2M PSII-LHCII supercomplexes. (a–c) Side view (a) and end views (b,c) within the membrane plane. (d) Top view towards the luminal surface of the paired supercomplexes, rotated 90° from panel (a). Solid and dashed lines extending from the top (cyan) and bottom (yellow) supercomplexes, respectively, illustrate the ~35° rotational offset between the supercomplexes. (e) The same view as panel (d) with the top supercomplex removed, revealing the top view of the lower supercomplex’s stromal surface from the perspective of the stromal gap. The stromal connections are also displayed. (f) Structural features of the “hinge” (magenta) and “knot” (red) stromal connections shown in different orientations starting on the left with the same side view as panel (a). Contact sites to the upper and lower supercomplexes are indicated with cyan and yellow mesh densities, respectively.
Model fitting reveals different degrees of overlap between facing LHCII S- and M-trimers
To interpret our 3D cryo-EM map, we performed rigid-body fitting using the atomic coordinates from two structures: the 3.2 Å cryo-EM structure of the unpaired C2S2 PSII-LHCII supercomplex from spinach26 (PDB ID: 3JCU) for modeling the central dimeric PSII core together with the monomeric Lhcb4 and Lhcb5 subunits, and the 2.5 Å crystal structure of the LHCII trimer from pea36 (PDB ID: 2BHW) for modeling the LHCII S- and M-trimers. The atomic coordinates of the PSII dimeric core, the LHCII trimers, and the monomeric Lhcb4 and Lhcb5 fit well into the cryo-EM density map (Fig. 2). While modeling the PSII dimeric core, we did not fit the extrinsic subunits PsbP, PsbQ and PsbTn, since previous biochemical and proteomic characterization showed that PsbQ and PsbTn are absent from the supercomplexes and PsbP is present in sub-stoichiometric amounts relative to the core subunits32, 37. Although a higher-resolution crystal structure of spinach Lhcb4 (2.8 Å) is available38, it lacks 73 amino acid residues at the N-terminus, which were nearly all detected in our sample by mass spectrometry (Supplementary Table 1). Thus, we decided to fit our cryo-EM map with the more complete structure of Lhcb4 obtained by Wei et al.26. An atomic structure is not yet available for the monomeric Lhcb6 antenna, which links the LHCII M-trimer to the CP47 subunit of the PSII core. To model Lhcb6 within our cryo-EM map, we used the 3D structure predicted by the PHYRE2 program39 for the P. sativum Lhcb6 protein sequence, which showed 50% coverage in the mass spectrometry analysis of our sample (see Supplementary Table 1).
Figure 2.

Fitting the cryo-EM density map of paired C2S2M supercomplexes with high-resolution structures. Side view within the membrane plane (a) and top view towards the luminal surface (b) of the paired C2S2M supercomplexes. The following structures were placed into the cryo-EM map by rigid-body fitting: the PSII dimeric core and monomeric Lhcb4 and Lhcb5 from spinach26 (PDB: 3JCU depleted of subunits PsbP, PsbQ, PsbTn; upper PSII dimer in cyan, lower PSII dimer in blue, upper Lhcb4 in pale red, lower Lhcb4 in dark red, upper Lhcb5 in pale brown, lower Lhcb5 in dark brown), the LHCII trimer from pea36 (PDB: 2BHW; upper S-trimers in pink, lower S-trimers in violet, upper M-trimer in pale green, lower M-trimer in dark green), the predicted structure of monomeric Lhcb6 from pea generated by the PHYRE2 algorithm39 (upper Lhcb6 in yellow, lower Lhcb6 in orange). (c–e) Schematic representations showing the positions of the LHCII trimers (c,d) and of all fitted supercomplex components (e), superimposed on the cryo-EM density map, showing end views within the membrane plane (c,d) and a top view towards the luminal surface (e). Colors match the structures in panels (a,b); solid lines for upper supercomplex components, dashed lines for lower supercomplex components.
Despite the limited resolution of our 3D map, fitting high-resolution components of the PSII-LHCII supercomplex clearly showed that LHCII trimers face each other across the stromal gap with various degrees of overlap. On the side of the paired supercomplexes containing only S-trimers, the apposing LHCII trimers completely overlapped and were in close proximity to the core complex, with the same position observed in the structure by Wei et al.26 (Fig. 2b,d). Conversely, on the opposite side of the paired supercomplexes containing additional M-trimers, the S-trimers were more distantly positioned with respect to the core complex if compared to the structure by Wei et al.26, and the apposing S- and M-trimers only partially overlapped (Fig. 2b,c). The overlaps observed between pairs of facing LHCII trimers, including S- with S- and S- with M- trimers, were specific and closely resembled two arrangements observed in the crystal structure of multilayer packed LHCII from spinach reported by Wan et al.40. The crystal packing of LHCII in this study, obtained with a physiologically-relevant cation concentration and in the presence of native membrane lipids, may replicate the stacking of LHCII in native grana membranes. In these crystals, cations are sandwiched between the neighbouring LHCII layers to screen the surface charges and mediate interactions by forming strong salt bridges between highly conserved negatively charged residues on facing LHCII trimers. Although we did not resolve physical contacts between apposing LHCII trimers in our cryo-EM map (Fig. 1a), the conservation of these negatively charged residues in the Lhcb1 and Lhcb2 subunits was confirmed in our preparation (Supplementary Fig. 3a, Supplementary Table 1). This evidence provides a plausible explanation for the complete overlap observed between two S-trimers (Fig. 2b,d), which are composed of Lhcb1 and Lhcb23, and thus have affinity for each other across the stromal gap due to salt bridge formation. It also helps explain the partial overlap between S- and M-trimers on the other side of the paired supercomplexes (Fig. 2b,c); the M-trimer contains Lhcb341, which may lack affinity for other LHCII subunits, thus reducing trimer overlap on that side.
Our cryo-EM map provides clear evidence at intermediate resolution for interactions between apposing LHCII trimers within paired PSII-LHCII supercomplexes. In so doing, it bridges the resolution gap between previous reports of LHCII interactions from X-ray crystallography of isolated LHCII trimers40 and cryo-electron tomography of C2S2 supercomplex arrays embedded within intact grana membranes20.
“Hinge” and “knot” densities make physical connections across the stromal gap
Well-defined electron densities were visible in the stromal gap between the paired supercomplexes, strongly indicating the presence of two different centrally positioned physical connections with distinct shapes and specific connections to the two C2S2M supercomplexes.
Towards the side of the paired supercomplexes containing only S-trimers, we observed a dimer of bulky densities with a “hinge” shape, which made defined contacts to PSII core subunits in each of the facing supercomplexes (Figs 1f and 3a,b). Comparison to the Wei et al.26 structure showed that this “hinge” density connects one PSII monomer in the upper supercomplex to one PSII monomer in the lower supercomplex, binding areas containing loops of the D1, CP43 and D2 proteins (Fig. 3b). Extensive biochemical and proteomic characterization of the sample used in this cryo-EM study32, 37 revealed the presence of an additional subunit of the PSII core that was not found in the Wei et al.26 unpaired C2S2 structure, a 10 kDa protein named PsbR. To date, the structure of PsbR is unknown and its positioning is still controversial. Although an extrinsic luminal location was hypothesized for PsbR based on its capacity to crosslink with the luminal PsbP subunit, PsbR’s hydrophobic C-terminus was suggested to be responsible for anchoring PsbR to the thylakoid membrane, leading to the protein’s insolubility42. Studies with Arabidopsis psbR knockout mutants revealed a reduction of the amount of PsbP bound to the PSII core and indicated a role for PsbR in stabilizing the supramolecular organization of PSII-LHCII supercomplexes43. However, it is not yet clear whether PsbR provides a direct docking site for PsbP or rather if it indirectly enhances the binding of PsbP on the luminal side by inducing a structural rearrangement in the PSII core. Taking all of this evidence into account, a possible interpretation of the “hinge” density in our cryo-EM map is that it is an extrinsic protein, potentially corresponding to PsbR. This protein, interacting with the stromal loops of the PSII core proteins, may induce a structural reorganization of PSII on the luminal side that favors the binding of PsbP, which stabilizes the association of LHCIIs with PSII within the membrane plane. In addition, by making connections that span the stromal gap between facing supercomplexes, the “hinge” can facilitate the stacking of adjacent grana thylakoids.
Figure 3.
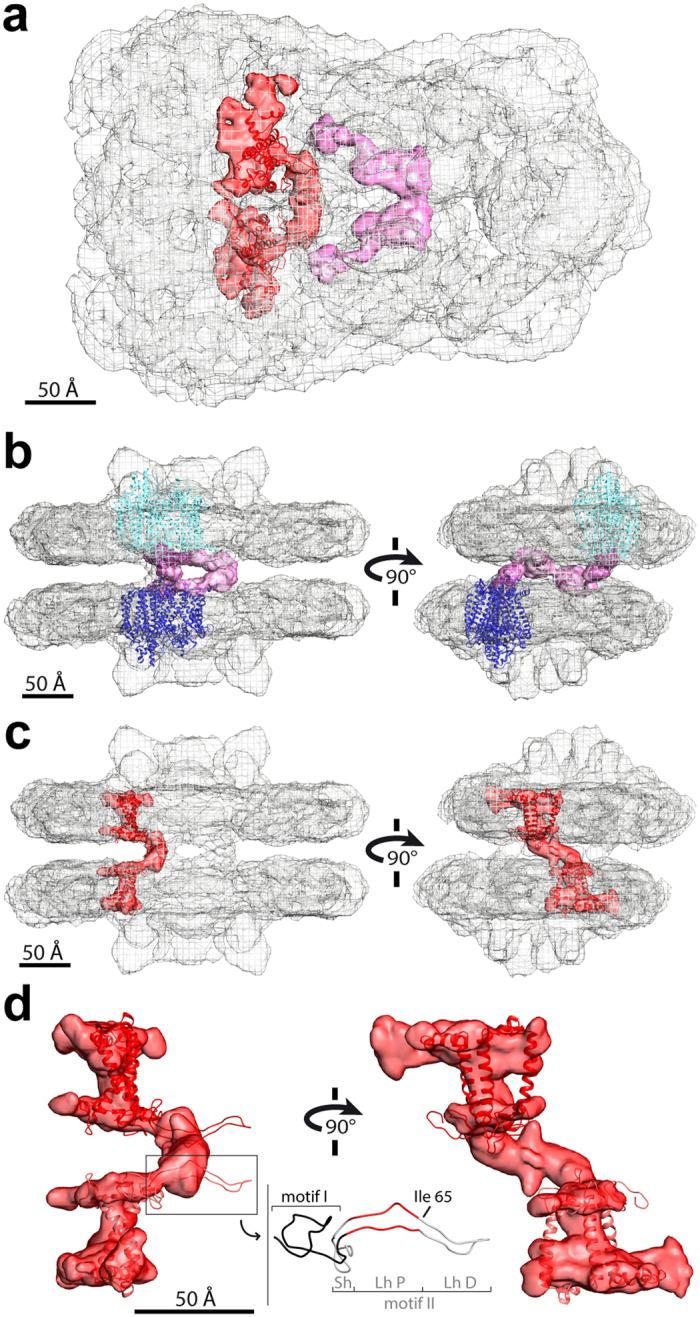
“Hinge” and “knot” connections cross the stromal gap of paired C2S2M supercomplexes. (a) Top view towards the luminal surface, showing the cryo-EM density map of paired C2S2M supercomplexes (grey mesh), with the stromal “hinge” density in magenta and the stromal “knot” density plus adjoining supercomplex region in red. Fitting of the high-resolution structure of Lhcb4 from spinach26 (PDB 3JCU: chain r; upper Lhcb4 in lighter red, lower Lhcb4 in darker red) as in Fig. 2 is shown. (b) Side and end views within the membrane plane showing the connections between the paired supercomplexes and the “hinge” (connecting supercomplex density in violet mesh). The hinge connects one PSII monomer of the upper supercomplex to one PSII monomer of the lower supercomplex (PDB 3JCU: chains A, C and D, corresponding to the subunits D1, CP43 and D2; upper PSII core in cyan, lower PSII core in blue)26. (c) Side and end views within the membrane plane showing the connections between the paired supercomplexes and the “knot”, which links an Lhcb4 of the upper supercomplex with an Lhcb4 of the lower supercomplex. Fitting of the high-resolution structure of Lhcb4 from spinach26 as in panel (a). (d) Enlarged views of the “knot” density plus adjoining supercomplex region fit with two Lhcb4 subunits, as in panel (c). The inset shows the N-terminal loop of the Lhcb4 structure by Wei et al.26, composed of motif I (Pro12–Lys41) in black and motif II (Pro42–Phe87) in grey and red. Motif II is subdivided into the short hairpin (Sh, Ala73–Phe87, grey), the proximal part of the long hairpin that fits within the “knot” cryo-EM density (Lh P, Pro42–Gln47 and Ile66–Ser72, red) and the distal part of the long hairpin that protrudes from the “knot” density (Lh D, Thr48–Ile65, grey). The flexible Ile65 residue, which separates the proximal and distal segments of the long hairpin, is indicated.
Towards the side of the paired supercomplexes containing the M-trimers, we observed a defined stromal density with a “knot” shape, forming an arm that extends diagonally across the stromal gap and contacts the supercomplexes on the opposite sides from the “hinge” binding sites (Figs 1f and 3a,c,d). This density is characterized by a symmetric and well-structured appearance, even when no D2 point group symmetry is imposed during image processing (data not shown). From its position in our fitted cryo-EM map, it is evident that the “knot” connection arises from the stromal surfaces of the Lhcb4 subunits, and thus can be assigned with high confidence to the N-terminal loops of two Lhcb4 subunits that face each other from two apposing supercomplexes (Fig. 3c,d). Lhcb4 contains a flexible ~90 amino acid N-terminal loop, easily lost during the crystallization process38, with dimensions that support its possible involvement in forming the “knot”. According to the structure by Wei et al.26, the N-terminal loop of Lhcb4 forms two motifs with irregular coil structures: motif I (Pro12–Lys41) and motif II (Pro42–Phe87) (Fig. 3d). Motif I forms a short loop near the stromal surface. Motif II contains a short hairpin loop (Ala73–Phe87) and a second hairpin loop that is approximately 40 Å long (Pro42–Ser72), extending into the stromal gap and then running nearly parallel to the stromal surface. In our structure, the connecting “knot” density that we attributed to the N-terminal loop of Lhcb4 confidently accommodates the proximal part of the motif II long hairpin formed approximately by Pro42–Gln47 and Ile66–Ser72. However, the distal part of the hairpin, corresponding approximately to Thr48–Ile65, protrudes from our 3D map (Fig. 3c,d). Spectroscopic analyses44 revealed that Ile65 is highly mobile, likely serving as a point of flexibility in the loop, while the Thr48–Ile65 region contains a helical stretch and a random coil. It is therefore plausible that the distal part of the long hairpin is responsible for “tying the knot” by homodimerizing to form the structured area at the center of the connecting density between the two Lhcb4 subunits.
The stromal gap-spanning connection between apposing Lhcb4 subunits may be secured by cation-mediated ionic interactions through the formation of salt bridges. Although the N-terminus of Lhcb4 contains some positively-charged amino acids, it is characterized by an overall net negative charge (Supplementary Fig. 3b). In particular, the distal part of the motif II long hairpin (Thr48–Ile65) is enriched in negatively-charged amino acids (Supplementary Fig. 3b) that are highly conserved in higher plants (Supplementary Fig. 4). Cations may screen these surface charges, forming strong salt bridges between negatively charged residues that mediate the interaction between the ends of two facing hairpin loops. Conversely, the proximal Ile66–Ser72 portion of the long hairpin as well as the short hairpin (Ala73–Phe87) each contain a Thr residue that becomes phosphorylated under high-light conditions45, indicating that these regions may regulate the homo-interactions of Lhcb4. Taken together, these results suggest that the Lhcb4 N-terminal loop may assume multiple conformations. Upon environmental light changes, phosphorylation of Lhcb4 might induce a conformational change in its N-terminal loop that “unties the knot”, allowing the redistribution of Lhcb4 from PSII-LHCII supercomplexes to PSII dimers and monomers45, while potentially enabling dynamic modulation of PSII-LHCII supercomplex pairing and ultimately the degree of grana staking.
Cations mediate the physical connection between paired PSII-LHCII supercomplexes
Despite the well-established requirement of shielding cations for the maintenance of grana stacking11, 12, the role of cations in PSII organization within grana remains unclear. Therefore, we assessed the potential function of cations in mediating the pairing of isolated PSII-LHCII supercomplexes. First, we isolated PSII-LHCII supercomplexes with sucrose density gradients that either contained (+glut) or lacked (-glut) glutaraldehyde, a cross-linking reagent often used to obtain information on protein interactions and the oligomerization state of complexes46, 47. Similar density gradient profiles of solubilized thylakoids were obtained for both conditions (Fig. 4a). PSII-LHCII supercomplexes, present in bands α3 and α3G, migrated to the same position on the sucrose gradient (Fig. 4a) and showed similar spectroscopic properties (Supplementary Fig. 5a). EM analysis, in cryo conditions for the α3 particles (Supplementary Fig. 1a) and by negative stain for the α3G particles (Supplementary Fig. 5b), showed that both samples contained paired supercomplexes. Next, buffer exchange was used to remove salt from both the + glut and −glut samples. Negative stain EM revealed that the majority of the supercomplexes detached from their paired partner upon salt removal (Fig. 4b), while the glutaraldehyde fixation was effective in maintaining the stromal connections between paired supercomplexes (Fig. 4c). Native polyacrylamide gel electrophoresis (PAGE) of the isolated supercomplexes (Supplementary Fig. 5c) confirmed that + glut particles (α3G) were in a paired oligomerization state, whereas -glut particles (α3) were unpaired. These results strongly support the hypothesis that the stromal connections between apposing supercomplexes are mediated by ionic bonds, which can be easily lost by changing the ionic strength of the buffers used during the isolation procedure. Notably, previous studies that reported the purification of paired PSII-LHCII supercomplexes also used cations in their isolation buffers2, 25. Recently, we observed a higher-order PSII-LHCII supercomplex oligomerization state consisting of two PSII-LHCII supercomplexes sitting side-by-side in the membrane plane (i.e., megacomplex), interacting with another two side-by-side supercomplexes across the stromal gap48. These paired megacomplexes correspond to band α5 of the sucrose gradient shown in Fig. 4a. Native PAGE showed that the removal of cations disrupted the stromal connections between facing megacomplexes (α5) unless these megacomplexes had been crosslinked with glutaraldehyde (α5G) (Supplementary Fig. 5c). In contrast, the lateral interactions between two supercomplexes were maintained without crosslinking, suggesting a specific role of salts in mediating the stromal connections between supercomplexes in adjacent thylakoid membranes.
Figure 4.

Salt removal causes unpairing of purified pea PSII-LHCII supercomplexes. (a) Isolation of PSII-LHCII supercomplexes from stacked pea thylakoid membranes by sucrose density gradients either containing ( + glut) or lacking glutaraldehyde (−glut). (b,c) Electron micrographs of uranyl acetate-stained PSII-LHCII supercomplexes contained in sucrose gradient bands α3 (b) and α3G (c) upon salt removal. Scale bar, 50 nm.
Paired PSII-LHCII supercomplexes are energetically coupled
Having assessed the requirement of cations for maintaining the structural connections between paired supercomplexes, we next sought to understand the function of supercomplex pairing by analyzing the fast chlorophyll a fluorescence induction curve49, which can be used to provide an estimation of the excitonic connectivity between PSII units50.
The fast phase of the chlorophyll a fluorescence induction curve is known as the OJIP transient, where O corresponds to the minimal fluorescence F0, J and I are inflections, and P is the peak corresponding to the maximal fluorescence Fm. Three rise phases of the F0 to Fm kinetics are distinguishable: O–J (0–2 ms), J–I (2–30 ms), and I-P (30–1000 ms)49. In the analysis of the OJIP transient, the major inflection points of the fast fluorescence induction curve are used for the calculation of several parameters characterizing the structure and photochemical activity of a photosynthetic sample50.
OJIP curves were recorded for isolated PSII-LHCII supercomplexes that had been diluted 1,000-fold either in the standard sucrose gradient buffer containing divalent cations or in a similar buffer that lacked salts (Fig. 5a). Compared to supercomplex pairs, unpaired supercomplexes in salt-free buffer showed increased values of F0 and reduced values of Fm, with a concomitant strong reduction of the OJIP curve. The differences in OJIP curve shape and F0/Fm ratio may reflect different redox states of the plastoquinone QA and differences in macromolecular organization. In particular, the rise of the F0 under low ionic conditions might be due to the disconnection of some LHCII from the supercomplexes, as suggested by negative stain EM that showed the detachment of LHCII trimers from PSII-LHCII particles upon salt removal (Fig. 4b).
Figure 5.

Estimation of the PSII excitonic connectivity in paired PSII-LHCII supercomplexes. (a) Chlorophyll a fluorescence induction curves (i.e., the OJIP transient) plotted on a logarithmic time scale (up to 1 s) for the paired PSII-LHCII supercomplexes prepared at 1.25 mg mL−1 in the sucrose gradient buffer and diluted 1,000-fold either in the sucrose gradient buffer (control, black line) or in a similar salt-free buffer (no salts, red line). The excitation light intensity was 2,400 μmol photons m−2 s−1, with an emission peak at 630 nm. (b) The initial 300 µs from the OJIP transient in panel (a) replotted on a linear time scale. The normalized O–J phase of the OJIP curve is labeled W (solid line), and the theoretical exponential curve corresponding to the unconnected system is labeled WE (dashed line). For calculation of W and WE see Supplementary Table 2. (c) Plot of WE – W using the values from panel (b). Graphs display the mean values ± standard deviations of six replicates.
Since the shape of the induction curve is influenced by excitation energy transfer between PSII units, commonly referred to as PSII connectivity51 or grouping52, the sigmoidicity of the initial phase of the fast fluorescence transient can be used to estimate the degree of PSII connectivity51. To estimate the energetic PSII connectivity in paired and unpaired PSII-LHCII supercomplexes, we evaluated the O–J phase of the OJIP transient using the method developed by Strasser and Stirbet53. In this approach, the degree of the PSII connectivity can be measured and compared between different samples by estimating the sigmoidicity of the chlorophyll a fluorescence induction curve during the first few microseconds. For the first 300 µs of the OJIP curves recorded in Fig. 5a, we calculated the normalized O–J phase (labelled W) and the theoretical exponential curve corresponding to an unconnected system (labelled WE, for definitions and calculation of these parameters see Supplementary Table 2). The increase in sigmoidicity from the theoretical unconnected curve WE to the experimental curve W was greater for the cation-containing sample than for the sample in salt-free buffer (Fig. 5b). The greater increase in sigmoidicity for the paired PSII-LHCII supercomplexes was further emphasized by plotting the difference between WE and W (Fig. 5c).
The O–J phase of the OJIP curve was also used to estimate the parameters of connectivity between PSII units49–53 (for definitions see Supplementary Table 2). The O–J curvature parameter C and the connectivity among PSII units parameter p were over two times higher in paired PSII-LHCII supercomplexes (C ≈ 0.35, p ≈ 0.30) than in unpaired supercomplexes treated with salt-free buffer (C ≈ 0.14, p ≈ 0.13) (Table 1). In addition, the overall grouping probability p2G and the probability of connectivity between PSII units ω (for definitions see Supplementary Table 2), indicated a significantly higher connectivity in paired PSII-LHCII supercomplexes (p2G ≈ 0.21, ω ≈ 0.20) than in unpaired supercomplexes (p2G ≈ 0.15, ω ≈ 0.06) (Table 1).
Table 1.
Selected parameters derived from fast fluorescence kinetic measurements to estimate the energetic connectivity between PSII units. Parameters for the paired PSII-LHCII supercomplexes prepared at 1.25 mg mL−1 in the sucrose gradient buffer and diluted 1,000-fold either in the sucrose gradient buffer (control) or in a similar salt-free buffer (no salts) were calculated according to methods previously reported49–53 as defined in Supplementary Table 2.
| C | p 2G | p | ω | |
|---|---|---|---|---|
| control | 0.35 ± 0.03 | 0.21 ± 0.02 | 0.30 ± 0.02 | 0.20 ± 0.01 |
| no salts | 0.14 ± 0.03 | 0.15 ± 0.03 | 0.13 ± 0.02 | 0.06 ± 0.01 |
Despite the limitations of calculations based on OJIP curve measurements, these results support the conclusion that excitation energy migrates between supercomplex pairs that interact across the stromal gap. This suggests that energetic coupling is possible between two reaction center cores that are not members of the same PSII dimer. However, at the limited resolution of our cryo-EM map, we cannot deduce the precise route for this excitation energy diffusion. A plausible hypothesis is that energy crosses the ~20 Å stromal gap via interactions between overlapping LHCII trimers, which may be precisely positioned to face each other by the more robust “knot” and “hinge” physical connections.
Molecular dynamics simulations predict additional transient interactions between overlapping LHCII trimers
To further investigate the interactions of paired C2S2M PSII-LHCII supercomplexes across the stromal gap, we ran molecular dynamics simulations using an atomic model generated from the fit of our cryo-EM density map (Fig. 2). After running the simulation for 7 ns, the backbone atoms had an average root mean square fluctuation (RMSF) of 0.23 nm compared to the reference time-averaged structure, with the central PSII core showing higher rigidity than the peripheral LHCII antennae system (Fig. 6 and Supplementary Fig. 6). Within several ps of starting the simulation, visible connections formed between the two apposing supercomplexes embedded within the appressed thylakoid membranes. Although the amino acid residues that form these connections varied during the simulation, they belong to the LHCII trimers and the monomeric Lhcb subunits (see Supplementary Video). These results suggest that flexible physical connections might occur between the antenna systems of PSII-LHCII supercomplexes that face each other across the stromal gap. The stromal N-terminal loops of Lhcb4 subunits positioned at the “knot” density seen in our cryo-EM map immediately formed connections that lasted throughout the entire timeframe of the simulation; conversely, the connections observed between the overlapping LHCII trimers were transient (see Supplementary Video). No strong connections formed at the position of the “hinge” density seen in the cryo-EM map, in agreement with the hypothesis that the proteins comprising this structure are not present in our fitted atomic model. The results of these molecular dynamics simulations are consistent with our cryo-EM study and may explain why only the most durable connections are well resolved in our cryo-EM map of paired C2S2M supercomplexes (Fig. 3).
Figure 6.

Molecular dynamics simulations. (a,b) Root mean square fluctuations (RMSF) for the backbone atoms of the paired C2S2M PSII-LHCII supercomplexes at the end of the 7 ns molecular dynamics simulation. Supercomplexes are colored according to RMSF values, and thylakoid membranes are grey. Side view within the membrane plane (a) and top view towards the luminal surface (b) of the paired C2S2M supercomplexes, with interactions that match the position of the “knot” connection in the stromal gap indicated with a black triangle. (c,d) Corresponding side view (c) and top view (d) of the paired C2S2M supercomplexes colored by subunit identity as in the Supplementary Video and Fig. 2.
Conclusions
The structure reported here for paired C2S2M PSII-LHCII supercomplexes is representative of the predominant form of PSII in pea plants grown at moderate light intensity. The interaction observed between apposing supercomplexes is mediated by physical connections that span the stromal gap, one of which is likely provided by the mutual interaction of two Lhcb4 subunits, possibly through salt bridges between negatively-charged amino acids in their N-termini. In addition, there is a specific overlap between facing LHCII trimers that may be due to the formation of salt bridges between highly conserved negatively-charged residues of the Lhcb subunits in facing LHCII trimers. The preservation of the PSII-LHCII supercomplex paired architecture requires the maintenance of physiological cation concentrations in all the buffers used during the purification procedure. This suggests that cation salt bridges mediate both the physical connections across the stromal gap as well as the transient interactions between facing LHCII trimers.
Migration of excitation energy occurs between supercomplexes that interact across the stromal gap. Molecular dynamics simulations of the paired PSII-LHCII supercomplexes suggest different roles for the physical connections and the overlapping LHCII trimers; the former likely serve a structural function to hold the supercomplexes together, whereas the latter may enable energy transfer between PSII cores embedded in adjacent thylakoid membranes. Thus, the stromal gap-spanning interactions between PSII-LHCII supercomplexes likely enable both grana stacking and efficient energy transfer between adjacent thylakoids.
It remains to be tested how the physical connections and energetic coupling we observed in isolated paired C2S2M supercomplexes compare to PSII-LHCII supercomplexes within intact thylakoid membranes. Exploration of this interplay between supercomplex organization and thylakoid architecture will benefit from an extensive visualization of supercomplexes within the native thylakoid membranes using in situ cryo-electron tomography54. Until now, evidences of contacts between PSII-LHCII supercomplexes occurring in adjacent thylakoid membranes were provided by cryo-electron tomography20, revealing interactions mediated by the stromal surfaces of both the LHCIIs and the PSII reaction center dimers. In this study, the geometry observed for the interaction of supercomplexes in two adjacent membranes was different from that seen in our study, but this may be due to the specific arrangement in 2D arrays of the supercomplexes observed in the tomograms. Moreover, Arabidopsis mutants respond to a lack of conventional LHCII trimers by generating Lhcb5-containing trimers18, 55, and to a lack of PSII cores by accumulating large amounts of LHCII organized into three dimensional structures56 to ultimately maintain their grana architecture. Therefore, the functional interactions we characterized for purified pairs of C2S2M PSII-LHCII supercomplexes are likely one of the several possible mechanisms occurring in vivo within adjacent thylakoid membranes that plants adopted to secure a functional grana stacking among different flexible strategies evolved to thrive in ever-changing light conditions.
Methods
Plant growth and isolation of thylakoid membranes
Pea plants (Pisum sativum L., var. Palladio nano) were grown in a growth chamber (SANYO MLR-351H) at 20 °C and 60% humidity, with a 8 h light/16 h dark photoperiod under 150 µmol photons m−2 s−1.
Thylakoid membranes were isolated from three-week-old leaves using the following procedure that maintains the stacked granal organization. These samples are hereafter referred to as stacked thylakoid membranes. Pea leaves were disrupted by grinding with a blender in 50 mM HEPES pH 7.5, 300 mM sucrose and 5 mM MgCl2. The suspension was filtered through four cotton cloth layers, and the filtrate was centrifuged at 1,500 g for 10 min. The pellet was washed once by centrifugation in the same buffer and then homogenized in 5 mM MgCl2 and diluted 1:1 with 50 mM MES pH 6.0, 400 mM sucrose, 15 mM NaCl and 5 mM MgCl2, followed by 10 min centrifugation at 3,000 g. The resulting pellet of stacked thylakoid membranes was washed once by centrifugation in 25 mM MES pH 6.0, 10 mM NaCl and 5 mM MgCl2, and then suspended and stored in 25 mM MES pH 6.0, 10 mM NaCl, 5 mM MgCl2 and 2 M glycine betaine. When necessary, stacked thylakoids were flash frozen in liquid nitrogen and stored at −80 °C.
Purification of pea PSII-LHCII supercomplexes
PSII-LHCII supercomplexes were isolated according to a previously optimized protocol32. Briefly, stacked thylakoid membranes at a Chl concentration of 1 mg mL−1 were treated with 50 mM n-dodecyl-α-D-maltoside (α-DDM) for 1 min at 4 °C in the dark. Phenylmethylsulphonylfluoride (500 mM) was present during the solubilization to inhibit protease activity. After centrifuging at 21,000 g for 10 min at 4 °C, 700 µl of the supernatant were added to the top of a linear sucrose gradient, prepared by freezing and thawing ultracentrifuge tubes filled with a buffer consisting of 0.65 M sucrose, 25 mM MES pH 5.7, 10 mM NaCl, 5 mM CaCl2 and 0.03% (w/v) of α-DDM (sucrose gradient buffer). For the crosslinking experiment with glutaraldehyde, 0.1% (v/v) glutaraldehyde was added to the sucrose gradient buffer. Centrifugation was carried out at 100,000 g for 12 h at 4 °C (Surespin 630 rotor, Thermo Scientific). The sucrose band containing PSII-LHCII supercomplexes was harvested and, if necessary, concentrated to a Chl concentration above 1.25 mg mL−1 by membrane filtration with an Amicon Ultra 100 kDa cutoff device (Millipore) and then stored at −80 °C. Remaining glutaraldehyde was inactivated after fractionation by adding Tris-HCl pH 8.0 to a final concentration of 80 mM.
Spectroscopic analyses
Absorption spectra in native conditions were recorded using a Lambda25 spectrophotometer (Perkin Elmer) at 12 °C.
Gel electrophoresis
Separation of thylakoid membrane protein complexes and PSII-LHCII complexes was performed using large pore blue native polyacrylamide gel electrophoresis57 (lpBN-PAGE), adopting the protocol previously described48.
Mass spectrometry analyses
PSII-LHCII supercomplexes were centrifuged at 20,000 g for 10 min at 4 °C, and the resulting pellet was rinsed in 10 mM HEPES pH 7.5. To remove the adhered pigments, proteins were precipitated in ice-cold acetone overnight at −20 °C. The extracting solution was centrifuged at 20,000 g for 20 min at 4 °C, and the denatured proteins were subsequently re-solubilized in a buffer consisting of 50 mM Tris-HCl pH 8.0, 7 M urea and 2 M thiourea. Insoluble material was removed by centrifuging at 15,000 g for 10 min. Protein concentration was determined using the Bradford assay58. Proteins at a concentration of 0.5 mg mL−1 were reduced with 10 mM DTT for 30 min at 37 °C and alkylated with 20 mM iodoacetamide for 30 min at room temperature in the dark. The protein in-solution digestion was conducted by adding Trypsin/Lys-C Mix (Promega, WI, USA) at a final protein:protease ratio of 25:1 (w/w), followed by overnight incubation at 37 °C. Peptide desalting was conducted by solid phase extraction59 using 30 mg Oasis HLB cartridges (Waters, MA, USA). Peptides were dried and then dissolved in 30 µl of LC-MS/MS mobile phase A (water containing 0.1% (v/v) formic acid).
LC-MS/MS analyses were performed using a micro-LC Eksigent Technologies (Dublin, USA) system, with a Halo Fused C18 column (0.5 × 100 mm, 2.7 μm; Eksigent Technologies Dublin, USA) as the stationary phase. The mobile phase was a mixture of 0.1% (v/v) formic acid in water (A) and 0.1% (v/v) formic acid in acetonitrile (B), eluting at a flow-rate of 15.0 μL min−1 and at an increasing concentration of solvent B from 2% to 40% over 30 min. The injection volume was 4.0 μL. The LC system was interfaced with a 5600 + TripleTOFTM system (AB Sciex, Concord, Canada) equipped with DuoSprayTM Ion Source and CDS (Calibrant Delivery System). The mass spectrometer was operated in information dependent acquisition (IDA) mode. Peptide profiling was performed using a mass range of 100–1600 Da (TOF scan with an accumulation time of 100.0 ms), followed by a MS/MS product ion scan from 200 to 1250 Da (accumulation time of 5.0 ms) with the abundance threshold set at 30 cps. The ion source parameters in electrospray positive mode were set as follows: curtain gas (N2) at 25 psig, nebulizer gas GAS1 at 25 psig, and GAS2 at 20 psig, ion spray floating voltage (ISFV) at 5000 V, source temperature at 450 °C and declustering potential at 25 V.
MS data were acquired with Analyst TF 1.7 (AB SCIEX, Concord, Canada). Raw files were processed with the search engine ProteinPilotTM v.5.0.1.0, 4895 (AB Sciex, Concord, Canada) using the Paragon algorithm v.5.0.1.0, 4874. The following sample parameters were used: Trypsin/Lys-C digestion, cysteine alkylation set to carbamidomethylation and no special factors. Processing parameters were set to “Biological modification”. All data files were searched, thorough ID search effort, using a UniProtKB/TrEMBL database containing Viridiplantae proteins (version 2016.09.02, with a total of 3,825,803 sequences), or a de novo database obtained by generating open reading frames from all six frames of each transcript in the P. sativum transcriptome (p.sativum_csfl_reftransV1 downloaded from https://www.coolseasonfoodlegume.org/organism/Pisum/sativum/reftrans/v1) concatenated with a reversed “decoy” version of the “forward” database. After searching, we accepted protein IDs that had a ProteinPilot Unused Score of at least 1.3 (equivalent to a 95% confidence interval) as a cutoff threshold and an estimated local false discovery rate (FDR) not higher than 1% according to Rardin et al.60.
Sequence analysis
Clustal Omega61 and Jalview62 software were used for multiple sequence alignment and conservation analysis, respectively. A phylogenetic tree was generated with ClustalW2.0 software63 using the UPGMA method and PAM250 substitution matrix.
Cryo-EM data collection
PSII-LHCII supercomplexes were prepared at 1 mg mL−1 in the sucrose gradient buffer. 4 μL of sample were applied to a glow-discharged lacey carbon grid (200 Cu mesh, Quantifoil) within the chamber of a Vitrobot (mark 3, FEI). After 60 s incubation at 100% humidity and 21 °C, a quick wash with 4 μL of 10 mM Hepes pH 7.5 was performed to remove sugar, excess solution was blotted from both sides for 4 s, and the grid was plunge-frozen in a liquid ethane/propane mixture. Samples were exposed to only dim green light during the grid preparation procedure. Data collection was performed on a Titan Krios microscope (FEI) operated at 300 kV using EPU automated acquisition software (FEI). Spanning a defocus range −1 to −3 μm, 6,834 micrographs were recorded on a Falcon II direct electron detector (FEI) at 59,000 magnification (image pixel size of 1.4 Å), with a total dose of 47.5 e−/Å2 fractionated over 7 frames (1.5 s exposure, dose rate of 30 e−/Å2/s).
Data processing and 3D reconstruction
Beam-induced motion was corrected by aligning the image frames with in-house developed software (https://github.com/dtegunov/k2align). Aligned images were used for single particle analysis. All image processing steps were performed using the Scipion platform (http://scipion.cnb.csic.es), which is an image processing framework that integrates several software packages into a unified interface64. For estimating the objective lens defocus parameters (i.e., contrast transfer function/CTF) in the transmission electron micrographs, we used the CTFFIND4 program65. For this project, a total of 33,729 cryo-EM particles were manually extracted from 6,834 micrographs using the particle picking tool of Xmipp66. These particles were classified in 2D using Xmipp67 and then in 3D using Relion68. Using e2initialmodel.py of EMAN2.169 and Ransac of Xmipp70, averages assigned to the PSII-LHCII supercomplex were used to generate several initial models and subsequently select among them the one with the highest score to be used as an unbiased low-resolution 3D template for refinement and classification. This template was used as an initial model for Relion 3D classification68. After 3D classification, 14,291 particles were identified as paired C2S2M supercomplexes, 7,021 particles as paired C2S2 supercomplexes and 5,641 particles as unpaired C2S2M supercomplexes. The paired C2S2M particles were processed further with 3D refinement, using the Relion auto-refine algorithm68 and D2 point group symmetry. After refinement, we used the multireference alignability method, recently implemented in Scipion, to select the “best particles” in the dataset according to the reference map and provide the alignment accuracy for each particle used in the reconstruction71. The final cryo-EM model was validated using the soft-alignment validation approach recently described by Vargas et al.71.
Fitting of atomic models into cryo-EM maps
UCSF Chimera72 software was used to model atomic structures into the 3D cryo-EM reconstruction. Surface-rendered views of the EM density were calculated at a threshold of 2.5 σ. Atomic co-ordinates from spinach26 (PDB ID: 3JCU) were used for modeling both the central dimeric PSII core (removing chains P, Q and U, corresponding to the extrinsic subunits PsbP, PsbQ and PsbTn, respectively) and the monomeric Lhcb4 and Lhcb5 subunits. Atomic coordinates from pea36 (PDB ID: 2BHW) were used for fitting the LHCII S- and M-trimers. For the monomeric Lhcb6, we fitted the 3D structure predicted by the PHYRE2 algorithm39 for the P. sativum Lhcb6 protein sequence derived from the corresponding transcript (p.sativum_csfl_reftransV1_0079196_5/148-357). Local fitting and adjustment of the Lhcb6 subunit and the LHCII trimers in the cryo-EM maps were performed by visual inspection, aided in the case of the S-trimer by its known localization in the available C2S2 atomic structure26.
Negative stain EM
PSII-LHCII supercomplexes were prepared at 1 mg mL−1 in the sucrose gradient buffer. To remove salts from the sample in a centrifugation step, we used Amicon Ultra 100 kDa cutoff buffer exchange spin columns (Millipore) and a salt-free buffer containing 25 mM MES pH 5.7 and 0.03% (w/v) α-DDM at a final ratio 1:60 (v/v). Concentrated samples were diluted to a final Chl concentration of ~30 µg mL−1 either in the sucrose gradient buffer or in the buffer with no salts. Samples were applied to glow discharged carbon-coated copper grids, washed quickly with distilled water and negatively stained with 2% (w/v) uranyl acetate. A FEI Tecnai F20-ST transmission electron microscope, equipped with a field emission gun (FEG) operated at 200 kV, was used for acquisition of micrographs, recorded at 38,000 magnification on a Gatan Orius 4.0 K × 2.7 K CCD camera.
Chlorophyll a fluorescence induction measurements
PSII-LHCII supercomplexes prepared at 1.25 mg mL−1 in the sucrose gradient buffer were used for the experiments. Samples were diluted to a final Chl concentration of 1.25 μg mL−1 in the sucrose gradient buffer or in a similar buffer devoid of any salt. The fluorescence induction OJIP transient was measured at room temperature, with a FL3500 double modulation fluorometer (Photon Systems Instruments). Dark-adapted samples were illuminated for 1 s with continuous actinic light (2,400 μmol photons m−2 s−1, emission peak at 630 nm). The first reliable point of the transient is measured at t0 = 0.02 ms after the onset of illumination. After recording fast fluorescence transients, the OJIP curves were analyzed according to methods previously reported49–53 to estimate the energetic connectivity between PSII units. Results shown in graphs and tables are presented as the mean value ± standard deviation of six replicates.
Molecular dynamics simulations
The atomic model generated by fitting the 3D cryo-EM map of the paired C2S2M supercomplexes (Fig. 2) was embedded within a pair of thylakoid membranes constituted by phosphatidylglycerol (PG), digalactosyldiacylglycerol (DGDG), monogalactosyldiacylglycerol (MGDG) and sulfoquinovosyl-diacylglycerol (SQDG) molecules, according to the detailed thylakoid membrane composition of higher plants proposed by van Eerden et al.73. The starting structure for the molecular dynamics simulations was the atomic coordinates of the protein components modeled into the cryo-EM 3D map, with the exclusion of any bound cofactors, and the predicted structure of Lhcb6. Prior to the insertion of the paired PSII-LHCII supercomplexes into the pair of membranes, the supercomplexes and the membranes were separately energy minimized and equilibrated. Subsequently, the supercomplex pair was embedded into the thylakoid membranes and solvated with water molecules and Na+ and Cl− ions according to van Eerden et al.73 in order to reach a globally uncharged system.
A 7 ns fine-grained simulation was performed with the GROMACS 4.6.6 package74, with the system interactions modeled by the GROMOS G53a6 force field75. The system was simulated using the Nosé-Hoover thermostat to control the temperature76, 77. By setting the thermal bath temperature at 300 K, the paired PSII-LHCII supercomplex, the pair of thylakoid membranes and the solvent were independently coupled every 0.5 ps. The pressure was semi-isotropically coupled every 2 ps to a reference pressure of 1 bar using the Parrinello-Rahman barostat78, and a compressibility of 4.6 × 10−5 bar−1 was applied to the system. Electrostatic and Van der Waals interactions were calculated using the Particle Mesh Ewald potential with a 1.4 nm cutoff. The mobility of the PSII-LHCII supercomplex pair was determined by calculating the root mean square fluctuations (RMSF) of backbone atoms (N, Cα and C atoms) during the molecular dynamics simulations with respect to the reference time-averaged structure:
where is the i−th backbone atoms and denotes the time-average.
Data availability
The data that support the findings of this study are available from the corresponding author upon request. The cryo-EM map was deposited at the Electron Microscopy Data Bank (EMDB) with accession code EMD-3825.
Addendum
After the completion of this manuscript, a paper was published by van Bezouwen et al.79, which reported a 5.3 Å structure of the higher plant unpaired C2S2M2 supercomplex. This PSII-LHCII supercomplex was isolated from Arabidopsis and its structure determined by single particle cryo-EM. In this work, the supramolecular organization of the subunits in the C2S2M2 is described in particular with respect to the location and orientation of trimeric and monomeric LHCII, and the location of all the chlorophylls in the subunits and their role in the energy flow from the peripheral light harvesting complexes to the PSII core is discussed. It is worth noting that, despite the lower resolution of our cryo-EM map, the fitting of the C2S2M components, and in particular of the LHCII trimers and the Lhcb6 subunit, is perfectly in accordance with that proposed by van Bezouwen et al.79. It is also worth noting that most of the chlorophylls responsible for energy connections observed in this recent structure are on the stromal side of the LHCII complexes. This evidence may suggest their involvement in the energy exchange between PSII cores located in adjacent thylakoid membranes in vivo, in accordance to the increase of PSII connectivity measured in our isolated paired PSII-LHCII supercomplexes.
Electronic supplementary material
Acknowledgements
This work was supported by the Italian Ministry of Education, University and Research, “Futuro in Ricerca 2013” program RBFR1334SB to C.P. and P.B. The authors acknowledge the support of employees and the use of experimental resources of Instruct, through Instruct Access Projects PID:1112 and PID:1363; Prof. Wolfgang Baumeister and Prof. Jürgen Plitzko for access to cryo-EM instrumentation and Gunter Pfeifer for assistance with cryo-EM data collection. Computational resources were provided by HPC@POLITO (The HPC Polito Project. Available: http://www.hpc.polito.it). We gratefully acknowledge the support of NVIDIA Corporation with the donation of the Quadro GP100 GPU used for this research. The authors thank Dr. Andrea Fantuzzi for helpful discussion.
Author Contributions
C.P. conceived the study. C.P., J.M.C., G.Z., E.M., and G.S. supervised the study. P.A. isolated PSII-LHCII supercomplexes. B.D.E. performed the preparation of cryo-EM grids and data acquisition. P.A., P.B. and A.G. manually picked particles from cryo-EM dataset. R.M., J.V. and C.O.S.S. performed the image processing. P.A., C.P., P.B., A.G., and G.Z. performed the model fitting. P.A. prepared samples for mass spectrometry and analysed mass spectrometry data. M.M. performed mass spectrometry data acquisition. C.P. and P.A. prepared the EM grids in negative stain and A.C. performed image acquisition. P.A. performed gel electrophoresis, sequence alignments and phylogenetic analysis. P.A. and C.P. performed fluorescence analyses. A.G. performed molecular dynamics simulations. C.P. wrote the article. B.D.E. critically read and revised the manuscript. All authors reviewed the results and approved the final version of the manuscript.
Competing Interests
The authors declare that they have no competing interests.
Footnotes
Electronic supplementary material
Supplementary information accompanies this paper at doi:10.1038/s41598-017-10700-8
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Barber J. Photosystem II: an enzyme of global significance. Biochem. Soc. Trans. 2006;34:619–631. doi: 10.1042/BST0340619. [DOI] [PubMed] [Google Scholar]
- 2.Boekema EJ, et al. Supramolecular structure of the photosystem II complex from green plants and cyanobacteria. Proc. Natl. Acad. Sci. 1995;92:175–179. doi: 10.1073/pnas.92.1.175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Hankamer B, et al. Isolation and characterization of monomeric and dimeric photosystem II complexes from spinach and their relevance to the organisation of photosystem II in vivo. Eur. J. Biochem. 1997;243:422–429. doi: 10.1111/j.1432-1033.1997.0422a.x. [DOI] [PubMed] [Google Scholar]
- 4.Danielsson R, et al. Dimeric and monomeric organization of photosystem II. Distribution of five distinct complexes in the different domains of the thylakoid membrane. J. Biol. Chem. 2006;281:14241–14249. doi: 10.1074/jbc.M600634200. [DOI] [PubMed] [Google Scholar]
- 5.Pagliano C, Saracco G, Barber J. Structural, functional and auxiliary proteins of photosystem II. Photosynth. Res. 2013;116:167–188. doi: 10.1007/s11120-013-9803-8. [DOI] [PubMed] [Google Scholar]
- 6.Ballottari M, Girardon J, Dall’Osto L, Bassi R. Evolution and functional properties of Photosystem II light harvesting complexes in eukaryotes. Biochim. Biophys. Acta - Bioenerg. 2012;1817:143–157. doi: 10.1016/j.bbabio.2011.06.005. [DOI] [PubMed] [Google Scholar]
- 7.Nevo R, et al. Architecture of thylakoid membrane networks. Lipids Photosynth. Essent. Regul. Funct. 2009;30:295–328. doi: 10.1007/978-90-481-2863-1_14. [DOI] [Google Scholar]
- 8.Kim E-H, Chow WS, Horton P, Anderson JM. Entropy-assisted stacking of thylakoid membranes. Biochim. Biophys. Acta. 2005;1708:187–195. doi: 10.1016/j.bbabio.2005.03.011. [DOI] [PubMed] [Google Scholar]
- 9.Borodich A, Rojdestvenski I, Cottam M. Lateral heterogeneity of photosystems in thylakoid membranes studied by Brownian dynamics simulations. Biophys. J. 2003;85:774–789. doi: 10.1016/S0006-3495(03)74519-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Kirchhoff H. Significance of protein crowding, order and mobility for photosynthetic membrane functions. Biochem. Soc. Trans. 2008;36:967–970. doi: 10.1042/BST0360967. [DOI] [PubMed] [Google Scholar]
- 11.Izawa S, Good NE. Effect of salts and electron transport on the conformation of isolated chloroplasts. II. Electron Microscopy. Plant Physiol. 1966;41:544–552. doi: 10.1104/pp.41.3.544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Barber J. Influence of Surface Charges on Thylakoid Structure and Function. Annu. Rev. Plant Physiol. 1982;33:261–295. doi: 10.1146/annurev.pp.33.060182.001401. [DOI] [Google Scholar]
- 13.Albertsson P. A quantitative model of the domain structure of the photosynthetic membrane. Trends Plant Sci. 2001;6:349–358. doi: 10.1016/S1360-1385(01)02021-0. [DOI] [PubMed] [Google Scholar]
- 14.Dekker JP, Boekema EJ. Supramolecular organization of thylakoid membrane proteins in green plants. Biochim. Biophys. Acta - Bioenerg. 2005;1706:12–39. doi: 10.1016/j.bbabio.2004.09.009. [DOI] [PubMed] [Google Scholar]
- 15.Anderson JM, Chow WS, De Las Rivas J. Dynamic flexibility in the structure and function of photosystem II in higher plant thylakoid membranes: the grana enigma. Photosynth. Res. 2008;98:575–587. doi: 10.1007/s11120-008-9381-3. [DOI] [PubMed] [Google Scholar]
- 16.Mullet JE. The amino acid sequence of the polypeptide segment which regulates membrane adhesion (grana stacking) in chloroplasts. J. Biol. Chem. 1983;258:9941–9948. [PubMed] [Google Scholar]
- 17.Day DA, Ryrie IJ, Fuad N. Investigations of the role of the main light-harvesting chlorophyll-protein complex in thylakoid membranes. Reconstitution of depleted membranes from intermittent-light-grown plants with the isolated complex. J. Cell Biol. 1984;98:163–172. doi: 10.1083/jcb.98.1.163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Andersson J, et al. Absence of the Lhcb1 and Lhcb2 proteins of the light-harvesting complex of photosystem II - effects on photosynthesis, grana stacking and fitness. Plant J. 2003;35:350–361. doi: 10.1046/j.1365-313X.2003.01811.x. [DOI] [PubMed] [Google Scholar]
- 19.Pribil, M., Labs, M. & Leister, D. Structure and dynamics of thylakoids in land plants. 65, 1955–1972 (2014). [DOI] [PubMed]
- 20.Daum B, Nicastro D, Austin J, McIntosh JR, Kühlbrandt W. Arrangement of Photosystem II and ATP Synthase in chloroplast membranes of spinach and pea. Plant Cell. 2010;22:1299–1312. doi: 10.1105/tpc.109.071431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Garab G, Mustardy L. Role of LHCII-containing macrodomains in the structure, function and dynamics of grana. Aust. J. Plant Physiol. 1999;27:648–658. [Google Scholar]
- 22.Kirchhoff H, et al. Structural and functional self-organization of Photosystem II in grana thylakoids. Biochim. Biophys. Acta - Bioenerg. 2007;1767:1180–1188. doi: 10.1016/j.bbabio.2007.05.009. [DOI] [PubMed] [Google Scholar]
- 23.Caffarri S, et al. Functional architecture of higher plant photosystem II supercomplexes. EMBO J. 2009;28:3052–3063. doi: 10.1038/emboj.2009.232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Kouřil R, Dekker JP, Boekema EJ. Supramolecular organization of photosystem II in green plants. Biochim. Biophys. Acta - Bioenerg. 2012;1817:2–12. doi: 10.1016/j.bbabio.2011.05.024. [DOI] [PubMed] [Google Scholar]
- 25.Nield J, et al. 3D map of the plant photosystem II supercomplex obtained by cryoelectron microscopy and single particle analysis. Nat. Struct. Biol. 2000;7:44–47. doi: 10.1038/71242. [DOI] [PubMed] [Google Scholar]
- 26.Wei X, et al. Structure of spinach photosystem II–LHCII supercomplex at 3.2 Å resolution. Nature. 2016;1:1–18. doi: 10.1038/nature19832. [DOI] [PubMed] [Google Scholar]
- 27.Boardman NK. Comparative photosynthesis of sun and shade plants. Annu. Rev. Plant Physiol. 1977;28:355–377. doi: 10.1146/annurev.pp.28.060177.002035. [DOI] [Google Scholar]
- 28.Anderson JM. Photoregulation of the composition, function, and structure of thylakoid membranes. Annu. Rev. Plant Physiol. 1986;37:93–136. doi: 10.1146/annurev.pp.37.060186.000521. [DOI] [Google Scholar]
- 29.Jia H, Liggins JR, Chow WS. Acclimation of leaves to low light produces large grana: the origin of the predominant attractive force at work. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2012;367:3494–3502. doi: 10.1098/rstb.2012.0071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Rumak I, et al. 3-D modelling of chloroplast structure under (Mg2+) magnesium ion treatment. Relationship between thylakoid membrane arrangement and stacking. Biochim. Biophys. Acta - Bioenerg. 2010;1797:1736–1748. doi: 10.1016/j.bbabio.2010.07.001. [DOI] [PubMed] [Google Scholar]
- 31.Schröppel-Meier G, Kaiser WM. Ion homeostasis in chloroplasts under salinity and mineral deficiency: II. Solute distribution between chloroplasts and extrachloroplastic space under excess or deficiency of sulfate, phosphate, or magnesium. Plant Physiol. 1988;87:828–832. doi: 10.1104/pp.87.4.828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Barera S, Pagliano C, Pape T, Saracco G, Barber J. Characterization of PSII-LHCII supercomplexes isolated from pea thylakoid membrane by one-step treatment with α- and β-dodecyl-D-maltoside. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2012;367:3389–3399. doi: 10.1098/rstb.2012.0056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Albanese P, et al. Dynamic reorganization of photosystem II supercomple xes in response to variations in light intensities. Biochim. Biophys. Acta - Bioenerg. 2016;1857:1651–1660. doi: 10.1016/j.bbabio.2016.06.011. [DOI] [PubMed] [Google Scholar]
- 34.Scheres SHW, Chen S. Prevention of overfitting in cryo-EM structure determination. Nat. Methods. 2012;9:853–854. doi: 10.1038/nmeth.2115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Kirchhoff H, et al. Dynamic control of protein diffusion within the granal thylakoid lumen. Proc. Natl. Acad. Sci. 2011;108:20248–20253. doi: 10.1073/pnas.1104141109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Standfuss JT, van Scheltinga AC, Lamborghini M, Kühlbrandt W. Mechanisms of photoprotection and nonphotochemical quenching in pea light-harvesting complex at 2.5 Å resolution. EMBO J. 2005;24:919–928. doi: 10.1038/sj.emboj.7600585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Pagliano C, et al. Proteomic characterization and three-dimensional electron microscopy study of PSII-LHCII supercomplexes from higher plants. Biochim. Biophys. Acta. 2014;1837:1454–1462. doi: 10.1016/j.bbabio.2013.11.004. [DOI] [PubMed] [Google Scholar]
- 38.Pan X, et al. Structural insights into energy regulation of light-harvesting complex CP29 from spinach. Nat. Struct. Mol. Biol. 2011;18:309–315. doi: 10.1038/nsmb.2008. [DOI] [PubMed] [Google Scholar]
- 39.Kelley LA, Mezulis S, Yates CM, Wass MN, Sternberg MJE. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015;10:845–858. doi: 10.1038/nprot.2015.053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Wan T, et al. Crystal structure of a multilayer packed major Light-Harvesting Complex: implications for grana stacking in higher plants. Mol. Plant. 2014;7:916–919. doi: 10.1093/mp/ssu005. [DOI] [PubMed] [Google Scholar]
- 41.Boekema EJ, van Roon H, van Breemen JFL, Dekker JP. Supramolecular organization of photosystem II and its light-harvesting antenna in partially solubilized photosystem II membranes. Eur. J. Biochem. 1999;266:444–452. doi: 10.1046/j.1432-1327.1999.00876.x. [DOI] [PubMed] [Google Scholar]
- 42.Roose JL, Frankel LK, Mummadisetti MP, Bricker TM. The extrinsic proteins of photosystem II: update. Planta. 2016;243:889–908. doi: 10.1007/s00425-015-2462-6. [DOI] [PubMed] [Google Scholar]
- 43.Allahverdiyeva Y, et al. Arabidopsis plants lacking PsbQ and PsbR subunits of the oxygen-evolving complex show altered PSII super-complex organization and short-term adaptive mechanisms. Plant J. 2013;75:671–684. doi: 10.1111/tpj.12230. [DOI] [PubMed] [Google Scholar]
- 44.Shabestari MH, Wolfs CJAM, Spruijt RB, van Amerongen H, Huber M. Exploring the structure of the 100 amino-acid residue long N-terminus of the plant antenna protein CP29. Biophys. J. 2014;106:1349–1358. doi: 10.1016/j.bpj.2013.11.4506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Fristedt R, Vener AV. High light induced disassembly of Photosystem II supercomplexes in Arabidopsis requires STN7-dependent phosphorylation of CP29. PLoS One. 2011;6 doi: 10.1371/journal.pone.0024565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Fadouloglou VE, Kokkinidis M, Glykos NM. Determination of protein oligomerization state: Two approaches based on glutaraldehyde crosslinking. Anal. Biochem. 2008;373:404–406. doi: 10.1016/j.ab.2007.10.027. [DOI] [PubMed] [Google Scholar]
- 47.Kastner B, et al. GraFix: sample preparation for single-particle electron cryomicroscopy. Nat. Methods. 2008;5:53–55. doi: 10.1038/nmeth1139. [DOI] [PubMed] [Google Scholar]
- 48.Albanese P, et al. Isolation of novel PSII-LHCII megacomplexes from pea plants characterized by a combination of proteomics and electron microscopy. Photosynth. Res. 2016;130:19–31. doi: 10.1007/s11120-016-0216-3. [DOI] [PubMed] [Google Scholar]
- 49.Stirbet A. & Govindjee. On the relation between the Kautsky effect (chlorophyll a fluorescence induction) and Photosystem II: Basics and applications of the OJIP fluorescence transient. J. Photochem. Photobiol. B Biol. 2011;104:236–257. doi: 10.1016/j.jphotobiol.2010.12.010. [DOI] [PubMed] [Google Scholar]
- 50.Stirbet A. Excitonic connectivity between photosystem II units: what is it, and how to measure it? Photosynth. Res. 2013;116:189–214. doi: 10.1007/s11120-013-9863-9. [DOI] [PubMed] [Google Scholar]
- 51.Joliot A, Joliot P. Etude cinetique de la reaction photochimique liberant l’oxygene au cours de la photosynthese. Comptes Rendus l’Académie des Sci. 1964;258:4622–4625. [PubMed] [Google Scholar]
- 52.Strasser, R. J. In Chloroplast Development (eds Argyroudi-Akoyunoglou, J. & Akoyunoglou, G.) Elsevier Biomedical, Amsterdam, 513–538 (1978).
- 53.Strasser RJ, Stirbet AD. Estimation of the energetic connectivity of PS II centres in plants using the fluorescence rise O-J-I-P - Fitting of experimental data to three different PS II models. Math. Comput. Simul. 2001;56:451–461. doi: 10.1016/S0378-4754(01)00314-7. [DOI] [Google Scholar]
- 54.Asano S, Engel BD, Baumeister W. In Situ Cryo-Electron Tomography: A Post-Reductionist Approach to Structural Biology. J. Mol. Biol. 2016;428:332–343. doi: 10.1016/j.jmb.2015.09.030. [DOI] [PubMed] [Google Scholar]
- 55.Ruban AV, et al. Plants lacking the main light-harvesting complex retain photosystem II macro-organization. Nature. 2003;421:648–652. doi: 10.1038/nature01344. [DOI] [PubMed] [Google Scholar]
- 56.Belgio E, Ungerer P, Ruban AV. Light-harvesting superstructures of green plant chloroplasts lacking photosystems. Plant Cell Environ. 2015;38:2035–2047. doi: 10.1111/pce.12528. [DOI] [PubMed] [Google Scholar]
- 57.Järvi S, Suorsa M, Paakkarinen V, Aro E-M. Optimized native gel systems for separation of thylakoid protein complexes: novel super- and mega-complexes. Biochem. J. 2011;439:207–214. doi: 10.1042/BJ20102155. [DOI] [PubMed] [Google Scholar]
- 58.Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976;72:248–254. doi: 10.1016/0003-2697(76)90527-3. [DOI] [PubMed] [Google Scholar]
- 59.Guo X, Kristal BS. The use of under-loaded C18 solid-phase extraction plates increases reproducibility of analysis of tryptic peptides from unfractionated human plasma. Anal. Biochem. 2012;426:86–90. doi: 10.1016/j.ab.2012.04.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Rardin MJ, et al. MS1 peptide ion intensity chromatograms in MS2 (SWATH) Data Independent Acquisitions. Improving post acquisition analysis of proteomic experiments. Mol. Cell. Proteomics. 2015;14:2405–2419. doi: 10.1074/mcp.O115.048181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Sievers F, et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2014;7:539–539. doi: 10.1038/msb.2011.75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Waterhouse AM, Procter JB, Martin DMA, Clamp M, Barton GJ. Jalview Version 2-a multiple sequence alignment editor and analysis workbench. Bioinformatics. 2009;25:1189–1191. doi: 10.1093/bioinformatics/btp033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Larkin MA, et al. Clustal W and Clustal X version 2.0. Bioinformatics. 2007;23:2947–2948. doi: 10.1093/bioinformatics/btm404. [DOI] [PubMed] [Google Scholar]
- 64.de la Rosa-Trevín JM, et al. Scipion: A software framework toward integration, reproducibility and validation in 3D electron microscopy. J. Struct. Biol. 2016;195:93–99. doi: 10.1016/j.jsb.2016.04.010. [DOI] [PubMed] [Google Scholar]
- 65.Rohou A, Grigorieff N. CTFFIND4: Fast and accurate defocus estimation from electron micrographs. J. Struct. Biol. 2015;192:216–221. doi: 10.1016/j.jsb.2015.08.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Abrishami V, et al. A pattern matching approach to the automatic selection of particles from low-contrast electron micrographs. Bioinformatics. 2013;29:2460–2468. doi: 10.1093/bioinformatics/btt429. [DOI] [PubMed] [Google Scholar]
- 67.Sorzano COS, et al. A clustering approach to multireference alignment of single-particle projections in electron microscopy. J. Struct. Biol. 2010;171:197–206. doi: 10.1016/j.jsb.2010.03.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Scheres SHW. RELION: Implementation of a Bayesian approach to cryo-EM structure determination. J. Struct. Biol. 2012;180:519–530. doi: 10.1016/j.jsb.2012.09.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Bell JM, Chen M, Baldwin PR, Ludtke SJ. High resolution single particle refinement in EMAN2.1. Methods. 2016;100:25–34. doi: 10.1016/j.ymeth.2016.02.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Vargas J, Alvarez-Cabrera A-L, Marabini R, Carazo JM, Sorzano COS. Efficient initial volume determination from electron microscopy images of single particles. Bioinformatics. 2014;30:2891–2898. doi: 10.1093/bioinformatics/btu404. [DOI] [PubMed] [Google Scholar]
- 71.Vargas J, Melero R, Gomez-Blanco J, Carazo J-M, Sorzano COS. Quantitative analysis of 3D alignment quality: its impact on soft-validation, particle pruning and homogeneity analysis. Sci. Rep. 2017;7 doi: 10.1038/s41598-017-06526-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Pettersen EF, et al. UCSF Chimera-a visualization system for exploratory research and analysis. J. Comput. Chem. 2004;25:1605–1612. doi: 10.1002/jcc.20084. [DOI] [PubMed] [Google Scholar]
- 73.Van Eerden FJ, De Jong DH, De Vries AH, Wassenaar TA, Marrink SJ. Characterization of thylakoid lipid membranes from cyanobacteria and higher plants by molecular dynamics simulations. Biochim. Biophys. Acta - Biomembr. 2015;1848:1319–1330. doi: 10.1016/j.bbamem.2015.02.025. [DOI] [PubMed] [Google Scholar]
- 74.Hess B, Kutzner C, Van Der Spoel D, Lindahl E. GROMACS 4: Algorithms for highly efficient, load-balanced, and scalable molecular simulation. J. Chem. Theory Comput. 2008;4:435–447. doi: 10.1021/ct700301q. [DOI] [PubMed] [Google Scholar]
- 75.Oostenbrink C, Villa A, Mark AE, Van Gunsteren WF. A biomolecular force field based on the free enthalpy of hydration and solvation: The GROMOS force-field parameter sets 53A5 and 53A6. J. Comput. Chem. 2004;25:1656–1676. doi: 10.1002/jcc.20090. [DOI] [PubMed] [Google Scholar]
- 76.Hoover WG, Ladd AJC, Moran B. High-strain-rate plastic flow studied via nonequilibrium molecular dynamics. Phys. Rev. Lett. 1982;48:1818–1820. doi: 10.1103/PhysRevLett.48.1818. [DOI] [Google Scholar]
- 77.Nosé S. A molecular dynamics method for simulations in the canonical ensemble. Mol. Phys. 1984;52:255–268. doi: 10.1080/00268978400101201. [DOI] [Google Scholar]
- 78.Parrinello M, Rahman A. Polymorphic transitions in single crystals: A new molecular dynamics method. J. Appl. Phys. 1981;52:7182–7190. doi: 10.1063/1.328693. [DOI] [Google Scholar]
- 79.van Bezouwen LS, et al. Subunit and chlorophyll organization of the plant photosystem II supercomplex. Nat. Plants. 2017;3 doi: 10.1038/nplants.2017.80. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon request. The cryo-EM map was deposited at the Electron Microscopy Data Bank (EMDB) with accession code EMD-3825.