

Abstract
Many stressors cause an increase in ventilation in humans. This is predominantly reported as an increase in minute ventilation (). But, the same can be achieved by a wide variety of changes in the depth (tidal volume, V T) and number of breaths (respiratory frequency, ƒR). This review investigates the impact of stressors including: cold, heat, hypoxia, pain and panic on the contributions of ƒR and V T to to see if they differ with different stressors. Where possible we also consider the potential mechanisms that underpin the responses identified, and propose mechanisms by which differences in ƒR and V T are mediated. Our aim being to consider if there is an overall differential control of ƒR and V T that applies in a wide range of conditions. We consider moderating factors, including exercise, sex, intensity and duration of stimuli. For the stressors reviewed, as the stress becomes extreme generally becomes increased more by ƒR than V T. We also present some tentative evidence that the pattern of ƒR and V T could provide some useful diagnostic information for a variety of clinical conditions. In The Physiological Society's year of ‘Making Sense of Stress’, this review has wide‐ranging implications that are not limited to one discipline, but are integrative and relevant for physiology, psychophysiology, neuroscience and pathophysiology.
Keywords: cold, heat, neurophysiology, pain, panic, respiratory frequency, tidal volume, ventilation
Abbreviations
- AMS
acute mountain sickness
- ARDS
acute respiratory distress syndrome
- ƒR
respiratory frequency
- HH
hypobaric hypoxia
- HN
hypobaric normoxia
- HRV
heart rate variability
- HVR
hypoxic ventilatory response
- IH
intermittent hypoxia
- KF
Kölliker‐Fuse
- MAP
mean arterial pressure
- NH
normobaric hypoxia
- NN
normobaric normoxia
- PAG
periaqueductal grey matter
- RPE
rating of perceived exertion
- Tes
oesophageal temperature
- TI
inspiratory time
- TE
expiratory time
alveolar ventilation
wasted ventilation
- VDanat
anatomical’ or ‘airway’ dead space
- VDphys
physiological dead space
minute ventilation
- VT
tidal volume
Introduction
It is widely reported that ‘breathing’ or minute ventilation (, l min−1) increases in response to a variety of external and internal stressors. In this context, ‘stress’ is defined as adverse or demanding physiological, pathophysiological or psychological stimuli that perturb the respiratory system. What is much less frequently reported than , is how is increased, in terms of the relative contribution of tidal volume (V T) and respiratory frequency (ƒR), and the mechanisms driving these contributions. The increase in can constitute hyperpnoea (increase in rate and depth of breathing commensurate with metabolic needs and arterial blood gas tensions), hyperventilation (increase in ventilation that is greater than that required by metabolic needs or arterial blood gas tensions) or tachypnoea (rapid (>20 breaths min−1), shallow breathing). Which of these responses is evoked can, as we shall see, depend on the nature of the stressor.
How is increased is important because of the impact it can have on the more critical measure of alveolar ventilation (), or the air actually available for gas exchange within the lung; this has consequences for the physiological and biochemical status of the body. is not routinely reported, probably because it is more difficult to measure than , requiring the measurement or estimation of physiological dead space (V Dphys). Multiplying this by ƒR ([V Dphys]ƒR) gives ‘wasted ventilation’ () and so, = – .
V Dphys is the sum of two separate components of lung volume, the first is the nose, pharynx and conduction airways which do not contribute in gas exchange. This component is often referred to as ‘anatomical’ or ‘airway’ dead space (V Danat) (Murias et al. 2014) and is typically 2.2 ml (kg body weight)−1, but is not fixed; varying with posture, breath holding and lung volume (Fowler, 1950; Nunn et al. 1959). Simplistically, V Danat is generally assumed to be relatively constant at about 150 ml for a 70 kg adult human. The second component of V Dphys, called ‘alveolar V D’, consists of alveoli that are either well ventilated but poorly perfused (e.g. apex of the lung in upright individuals) or well perfused but poorly ventilated (e.g. as in small airway disease) (Murias et al. 2014). This can vary with body size and other more subtle factors.
It follows that if an increase in is achieved primarily via V T, the effect of V Dphys on is minimised (Sheel & Romer, 2012). In contrast, if an increase in is achieved primarily via ƒR, the effect of V Dphys on is maximised (e.g. panting). An example is given in Table 1 using just V Danat, it demonstrates that the same can result in a ranging between 0 and 6.0 l min−1.
Table 1.
The effects of breathing frequency and tidal volume on alveolar ventilation (adapted from Braun, 1990)
| Minute ventilation (, l min−1) | Breathing frequency (ƒR, breaths min−1) | Tidal volume (V T, l) | Dead space* (, l min−1) | Alveolar ventilation (, l min−1) |
|---|---|---|---|---|
| 7.5 | 50 | 0.15 | 7.5 | 0 |
| 7.5 | 30 | 0.25 | 4.5 | 3.0 |
| 7.5 | 20 | 0.375 | 3.0 | 4.5 |
| 7.5 | 10 | 0.75 | 1.5 | 6.0 |
*An anatomical dead space of 150 ml breath−1 is assumed.
As V Dphys contributes to the work of breathing without contributing to , the work of breathing associated with any given is relatively increased when is achieved by ƒR rather than V T. However, because increasing via ƒR has less of an effect on it also has less of an effect on alveolar and arterial and, therefore, blood gas homeostasis. The differing impact of on the work of breathing and blood gas homeostasis may, in itself, have implications for the respiratory pattern seen in response to differing stressors. Finally, the perfusion of the lung and resulting ventilation:perfusion matching in different stresses (e.g. hypoxia, immersion, gravity) and respiratory disease states can result in hypoxaemia and hypercapnia, which then influence ventilation.
The breathing pattern
The measured represents the combination of a neural output and a mechanical response. The neural output and rhythm arise from excitatory inputs from a variety of sources. They may change during an inspiration (‘Phasic’, e.g. with increasing lung volume (Hering‐Bruer inflation reflex – weak in humans, not seen until V T > 800 ml)) or may be impacted upon by a constant stimulus (‘Tonic’, e.g. body temperature). The profile of neural output at the beginning of neural inspiration plays a role in determining V T. In the present review we focus on short‐term stress‐induced changes in ventilation in response to stressors such as: cold, heat, hypoxia, pain and panic. This excludes consideration of sleep, which has an impact on the respiratory pattern, and has been reviewed elsewhere (Phillipson, 1978).
Breathing patterns have primarily been examined in an attempt to gain an insight into the output of the ‘respiratory centres’ (Widdicombe & Winning, 1974; Richter, 1982). On the basis of rebreathing experiments in cats and humans, Clark & von Euler (1972) suggested ƒR and V T are linked in a definitive way, and that V T is influenced by both respiratory drive (mean rate of inspiratory flow (V T/inspiratory time)) and the duration of inspiration (timing), with the rate of lung inflation being primarily determined by the chemical drive for respiration. When the vagi were cut in anaesthetised animals, and volume feedback thereby prevented, stimulation of breathing caused an increase in V T but not ƒR. Younes & Remmers (1981) state that the breathing pattern is determined by four principal variables: (1) inspiratory‐time profile, (2) inspiratory duration, (3) expiratory volume–time profile, (4) expiratory duration (Fig. 1). Stressors can affect by altering one or more of these variables, which, after addition or subtraction, then alter ƒR and/or V T. Thus, ƒR may be increased by a reduction in both, or either, inspiratory and expiratory duration, or a reduction in one that exceeds the prolongation of the other.
Figure 1. The route by which V T and ƒR may be altered.
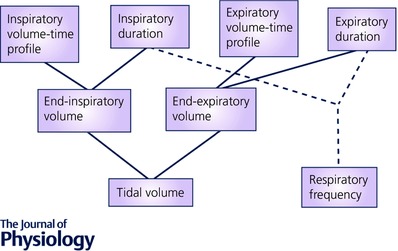
Reproduced from Younes & Remmers, 1981 (with permission).
Several authors (Pearson & Cunningham, 1973; Miller et al. 1974; Gardner, 1977; Bechbache et al. 1979) have pointed out that there are differing response times for changes in V T, inspiratory time (T I) and expiratory time (T E). Cunningham (1974) suggests that the steady state relationships between these variables is not fully re‐established for 20 min after ventilatory drive is altered, but most of the changes occur within 1–2 min.
From the mechanical perspective, it is generally reported that for a given there is a ƒR that minimises muscle work in relation to ventilation. However, Mead (1960) reported that in humans at rest and exercise the resulting ƒR is the one which is least costly in terms of average force. In this context, V T increases until it is at the top of the lung compliance curve and, for a given response, V T becomes smaller as the resistance or elasticity of the respiratory system increases; variables such as temperature can alter compliance and therefore the relationship between V T and ƒR. Additionally, during sustained increases in ventilation in some pulmonary disease states, and in situations where gas density is increased (e.g. hyperbaric environments), the relationship between ƒR and V T can be altered to minimise the work of breathing (Dongelmans et al. 2011; West, 2012). Given that, from a mechanical perspective, an initial increase in via V T is most efficient, when this response is not observed, factors in addition to respiratory mechanical efficiency must be involved. For example, V T is affected by both central neural output (including an inspiratory ‘off‐switch’) and peripheral neural (pulmonary stretch, irritant and J receptor) input to neural inspiration; these factors vary in relation to the absolute lung volume and consequent Functional Residual Capacity at the time (Younes & Remmers, 1981). At high lung volumes mechanical constraints can prevent V T from increasing further, and when the rate of rise of neural output is high, a relatively short amount of time is allowed for lung volume to decrease; as a consequence, mechanical inspiration can begin at a higher absolute volume, limiting V T and driving ƒR. As we shall see, such an inspiratory shift in end‐expiratory lung volume following stimulation of respiratory neurons is a feature of some environmental stressors and helps explain the respiratory response observed.
In order to avoid variations caused by respiratory mechanics, mouth occlusion pressure 100 ms after the onset of inspiration against a closed airway has been measured (P0.1). In this period the respiratory muscles should shorten solely in response to output from the respiratory centres in conscious humans (Whitelaw et al. 1975; Whitelaw & Derenne, 1993). However, this approach does not avoid the reported impact of the use of respiratory instrumentation on breathing pattern. Mouthpieces and nose‐clips have been reported to increase , V T, T I, T E, mean inspiratory flow, and mean expiratory flow, but decrease ƒR (Perez & Tobin, 1985).
Exercise
Exposure to environmental stressors often co‐exists with exercise; this can mask the nature of the response to an environmental stress, which is over‐ridden by the metabolic demands of exercise. Additionally, and with time, the heat production associated with exercise increases body temperature, which can alter lung function (Tipton et al. 2017) and, independently, breathing pattern.
The involvement of central command (that is, the activity of motor and premotor areas of the brain relating to voluntary muscle contractions or ‘effort‐induced modulation of autonomic function’ (Williamson et al. 2006; de Morree et al. 2012) in the rapid ƒR response to exercise is supported by the work of Bell & Duffin (2006), who reported an immediate response of ƒR but not V T on commencement and cessation of exercise, and with a change in the mode of exercise. Although central command, muscle afferent fibres and metabolic stimuli are major determinants of during exercise (Forster et al. 2012), they act with different timings when exercise intensity changes, with metabolic stimuli having a slower time course than the faster inputs of central command and muscle afferents (Duffin, 2014). The ƒR and V T responses to exercise are less well studied than the response, but there are some intriguing studies involving hypnosis and passive exercise that seem to indicate that ƒR and V T are under separate control (Thornton et al. 2001; Bell & Duffin, 2006; Nicolò et al. 2016), with the suggestion that ƒR is less, and V T more determined by metabolic factors. Ohashi et al. (2013) provided evidence from studies examining CO2 homeostasis at rest that suggests that ƒR can be regulated by non‐metabolic factors. Recent evidence suggests that ƒR is a valid marker of rating of perceived exertion (RPE) during cycling exercise (Nicolò et al. 2014, 2016). RPE and ƒR also respond in a similar way to muscle fatigue (Marcora et al. 2008), muscle damage (Davies et al. 2009), hypoxia (Koglin & Kayser, 2013) and increases in body temperature (Hayashi et al. 2006), but appear disassociated from V T (Noble et al. 1973; Nicolò et al. 2016). The close association between RPE and ƒR has led to the suggestion that they share a common regulatory mechanism involving central command (Nicolò et al. 2016).
After the initial response to the onset of exercise, and at low work intensities, the ventilatory response is then primarily achieved by an increase in V T. With increasing intensity (stress), ƒR is also increased (Sheel & Romer, 2012). During more strenuous exercise, once V T reaches 50% to 60% of vital capacity, there is a plateau in V T and further increases in are accomplished solely by increases in ƒR (Fujii et al. 2008b; Sheel & Romer, 2012). As mentioned, airway resistance, lung compliance and the work/force requirements of breathing can influence the response to a stressor. In exercise the initial increase in by V T is efficient but is limited by lung compliance at higher V T where increases in ƒR become preferable from a mechanical perspective.
The additional rise in caused by increases in ƒR is commonly referred to as the ‘tachypnoeic’ breathing pattern of heavy exercise and is achieved via reductions in both inspiratory and expiratory time (Sheel & Romer, 2012). In general, the breathing pattern and interaction of ƒR and V T are highly variable between individual athletes during incremental exercise (Carey et al. 2008). The mode (running, cycling) and rate of exercise can also influence breathing pattern, with breathing becoming entrained to exercise rhythm in some individuals. Evidence is emerging that respiratory and locomotor rhythms during exercise; so‐called ‘locomotor respiratory coupling’ may be responsible (Lunt et al. 2011; Sheel & Romer, 2012).
Therefore, the determinants of the ventilatory response to exercise include: (i) a central medullary rhythm generator/integrator; (ii) neural inputs into this integrator from higher locomotor areas of the central nervous system (central command, central chemoreceptors, direct effect of temperature) and from the periphery (chemoreceptors, mechanoreceptor); and (iii) the regulation of the distribution of efferent motor output to the ventilatory muscles (Sheel & Romer, 2012). The interaction between these mechanisms remains largely unknown; evidence suggests that, in the absence of mechanical ventilatory constraints, there is differential regulation of V T and ƒR during exercise. It would therefore seem important to consider the ƒR and V T responses separately to fully understand the mechanisms underlying exercise hyperpnoea.
The initial response to various stressors includes an increase in the activity of the sympathetic nervous system. This is changed in a global fashion, leading to an increase in cardiovascular function and a release of adrenal catecholamines (Homma & Masaoka, 2008). Since the seminal work of Cannon (1929), the human ventilatory response to external and internal stressors has also been recognised as an important part of the body's preparation during the ‘fight or flight’ response, the prototypic human response to stress (Jansen et al. 1995; Taylor et al. 2000). Hyperventilation is often considered part of this response (Van Diest et al. 2001), but how this hyperventilation is brought about (ƒR and/or V T) has not been thoroughly investigated. The pattern of ƒR and V T may also have wider implications than just ; it is possible that the pattern itself is indicative of other physiological and pathophysiological pathways and states.
It is concluded that, in conscious humans, a variety of factors influence the pattern of breathing. These include: lung volume and the reflexes evoked by inflation, the level of chemoreflex stimulation; the time variability of neural mechanisms; the level of consciousness; and the rhythm of body movements during exercise. It is concluded that how is increased matters. Whilst it is convenient to report only , it can be misleading if different stressors cause an increase in by different combinations of ƒR and V T. Also, important information may be contained in the way is increased.
In the present review we examine the breathing pattern in terms of the ƒR and V T across a range of stressors examining how that response differs depending on the nature of the stress. A review of this scope has not been undertaken previously. Our aim was to consider if there is an overall differential control of ƒR and V T that applies in a wide range of conditions. In addition, if is increased by different combinations of ƒR and V T the question then becomes, how? Therefore, where possible, we also consider the potential mechanisms that underpin the responses identified.
Review methodology
The following stressors were examined: sudden cold exposure and hypothermia, passive and active (involving exercise) hyperthermia, hypoxic exposure, induced pain and panic (triggered by rebreathing CO2 or external sensory stimuli).
A literature search was undertaken using Medline, Pubmed, and Science Direct search engines to identify studies that reported both ƒR and V T in humans following exposure to a stressful environment. Keywords and phrases used included: [‘rate’ OR ‘frequency’ OR ‘ventilation’] AND [‘volume’ OR ‘depth’ OR ‘ventilation’] AND [‘cold water’ OR ‘cool water’ OR ‘hypothermia’ OR ‘heat’ OR ‘hot’ OR ‘hyperthermia’ OR ‘exercise’ OR ‘panic’ OR ‘anxiety’ OR ‘fear’ OR ‘pain’ OR ‘noxious’ OR ‘altitude’ OR ‘hypoxia’]. The reference lists of relevant articles were searched and experts and colleagues working in the area were contacted regarding the existence of any additional studies suitable for inclusion. The searches were undertaken in August 2015 and no restrictions were made on study design, methodology or method of assessing ƒR and V T. There was no blinding to study author, institution or journal at this stage. The methodologies of the reviewed studies were heterogeneous in terms of the: age and sex of participants; clothing and equipment used; and duration and intensity of the stressor.
We have focused on human studies but, in the absence of relevant human data, have included animal models to help determine underlying mechanisms. In terms of mechanistic pathways, as recently reviewed (Dempsey & Smith, 2014), there is no doubt that evidence from animals and animal models of human disease have helped our understanding of the input of peripheral and central chemoreceptors to ventilation, as well as their role in disease states such as heart failure, sleep apnoea and chronic obstructive pulmonary disease. Given this, it is argued that the organisation of peripheral receptors, their central reflex arcs and integration with the central respiratory pattern generator are likely to be similar between animals and humans. Indeed, Hess et al. (2013) have identified the pre‐Bötzinger complex and parafacial regions in the human brainstem and shown inspiratory and expiratory related activity with fMRI. Further evidence of correspondence comes with the role of Phox2b gene in the control of central chemoreception and respiration in humans (Weese‐Mayer et al. 2003) and animals (Gaultier et al. 2004). However, we acknowledge that species differences exist, as with the relative strength of the pulmonary stretch receptors when activated by increases in ventilation brought about by stimulation of the carotid bodies; the dog displays tachycardia and the cat bradycardia; reflecting the predominance of lung inflation versus peripheral chemoreceptor reflexes, respectively. In comparison with humans, in some areas animal responses differ in terms of their magnitude (e.g. the ventilatory patterns to hyperthermia (Robertshaw, 2006); vagal lung inflation reflexes (Guz et al. 1971)), in others they are similar (e.g. respiratory responses to cold water immersion (Datta & Tipton, 2006)).
Where possible, we extracted mean ƒR and V T data before, during and after exposure to the various stressors. In some studies, data were also extracted from a ‘Control’ group who were not exposed to the stress being examined. The percentage changes in both ƒR (x‐axis) and V T (y‐axis) from baseline or control values were then calculated and plotted on a scatter graph. For clarity, the stressors are plotted separately and the associated legend and symbols briefly describe the individual data points.
The magnitude or intensity of a stress is likely to have an influence on the nature of the ventilatory response; this magnitude may change with factors such as absolute temperature or oxygen concentrations. It may also change with time due, for example, to increasing cooling, heating or hypoxia. It may also decrease with time such as with the adaptation of peripheral thermal receptors following a sudden change in skin temperature. As noted, the co‐existence of exercise with environmental stress adds inputs to the breathing pattern compared to those when exposed at rest. We considered these influences (e.g. intensity and duration of stress and co‐existent exercise) in our interpretation of the data.
Results: ventilatory responses
Having reviewed the ventilatory response to exercise in the previous section, in the following sections ƒR and V T data are examined in the following:
Sudden or prolonged cold exposure (12 studies; 176 participants)
Passive and active hyperthermia (13 studies; 153 participants)
Hypoxia (10 studies; 290 participants)
Panic (8 studies; 470 participants)
Pain (4 studies, 56 participants)
Cold exposure
Rapid cooling of the skin by immersion in cold water produces the ‘cold shock’ response (Tipton, 1989); this includes an inspiratory gasp, hyperventilation, tachycardia, peripheral vasoconstriction, hypertension, a marked reduction in cerebral blood flow velocity (Datta & Tipton, 2006; Mantoni et al. 2007; Croft et al. 2013; Barwood et al. 2016) and increased circulating level of catecholamines (Heymans et al. 1930; Tipton & Golden, 1987; Barwood et al. 2014).
The cold shock response is evoked by the dynamic response of the peripheral cold thermoreceptors (Tipton, 1989). However, the precise afferent pathways responsible for the respiratory responses to cold‐water immersion remain to be elucidated. It has been suggested that thermoafferents from the peripheral cold receptors directly stimulate respiratory networks (Keatinge & Evans, 1961; Goode et al. 1975), with the sensitivity of the central chemoreceptors remaining unchanged (Cooper et al. 1976). Part of the respiratory response to cold immersion may result from reflex stimulation of spinal networks including α‐motorneurones innervating the intercostal muscles and diaphragm (Keatinge & Nadel, 1965). Keatinge & Nadel (1965) concluded that the reflex respiratory responses to cold water in the cat are also mediated at midbrain level, with the cerebrum not being essential for the response.
Tipton & Harris (in Datta & Tipton, 2006) report that, following 60 s upright immersion to the diaphragm of rats in 8°C water, c‐fos protein immunohistochemistry identified activated neurones in the nucleus tractus solitarius, area postrema and dorsal motor nucleus; areas known to process cardiovascular and respiratory afferents. No expression was observed with immersion in water at 39°C. The areas generating respiratory rhythm (dorsolateral pons (Kölliker‐Fuse nucleus), ventrolateral medulla between the facial nucleus and the lateral reticular nucleus) were not examined.
In naked individuals the cold shock response peaks in water between 15 and 10°C (Tipton et al. 1991). Below about 14°C there is increasing activation of cold nociceptors (Simone & Kajander, 1997). Studies have shown noxious cold stimuli (rat hindlimb immersion in 4° C water) causes c‐fos activation of neurons in lamina I of the spinal dorsal horn (Todd et al. 2005), suggesting this ascending pathway is involved in transmitting the thermoreceptor volley following cold immersion to the pons (Hinckel & Schroder‐Rosenstock, 1981). The pons has areas controlling eupnoea and apneusis (prolonged inspiration). It (Kölliker‐Fuse (KF) nucleus) allows the frequency of breathing to change very quickly. The Hering‐Breuer afferents project there and control inspiration off‐switching; it is therefore a major regulator of respiratory phase switching (Abdala et al. 2016).
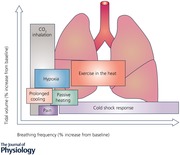
In vagotomised animals, eupnoea is transformed to gasping following removal of the pons or during severe hypoxia (Lumsden, 1924; Paton et al. 2006; Smith et al. 2007), and destruction of neurons in the rostral medulla with neurotoxins leads to the elimination of gasping but not eupnoea (Fung et al. 1994); this region was later shown to be in the pre‐Bötzinger complex (Paton et al. 2006; St‐John et al. 2009). This suggests that the neural mechanisms for gasping may be distinct from those necessary for the generation of eupnoea. That the first respiratory response to immersion in cold water is a gasp, suggests that the cold thermoreceptor volley elicited by cold immersion powerfully excites this area. However, there remains a possibility that the initial ‘gasp’ may be an augmented breath rather than a gasp, defined as the respiratory movement that occurs when the pons is removed or in extreme hypoxia. The hyperventilation seen after the initial gasp of the cold shock response reflects a stress response accompanied by sympathetic overdrive (see Fig. 2 A). The speed of this response, as with the gasp response, suggests activation of an uncomplicated (direct) neural pathway, thought to be mediated through the tegmentum of the midbrain and the hypothalamus (Winters et al. 2000).
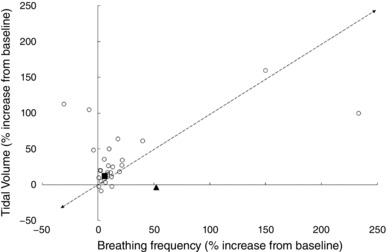
Figure 2. Increase in tidal volume and respiratory frequency in response to the cold shock response cold exposure for ≤120 s (A) and prolonged cooling (>120 s; B).
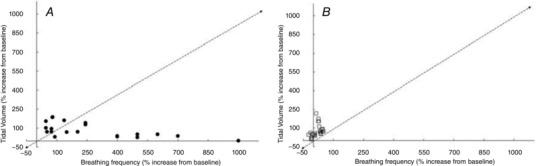
Data extracted from the following references: Barwood et al. (2013, 2014); Frank et al. (1997); Eglin & Tipton (2005); Hayashi et al. (2006); Mantoni et al. (2007, 2008); Button et al. (2015); Eglin et al. (2015); Wagner & Horvath, (1985); Tipton & Golden (1987); Tipton et al. (1991).
The cold shock response peaks during the first minute of immersion and then diminishes over the next minute as the peripheral thermoreceptors adapt. The respiratory response includes a three‐ to fourfold increase in ventilation (Martin & Cooper, 1978). Several studies (Tipton et al. 1991; Eglin & Tipton, 2005; Mantoni et al. 2007) have reported increases in ƒR of > 200% of baseline values, while corresponding V T values did not typically exceed 100% (Fig. 2 A). This is particularly apparent if the respiratory data are recorded at a higher sampling frequency or breath by breath (Mekjavic et al. 1987b; Datta & Tipton, 2006). For example, analysis of ƒR and V T at 10 s periods during the first 20–30 s of immersion in cold water shows that the increase in is primarily determined by an increase in ƒR, with V T not increasing until ƒR begins to fall back towards pre‐immersion levels (Tipton et al. 1991). Thus, analysis of the components of after the first minute of immersion shows an increasing contribution of V T to (Hayward & Eckerson, 1984; Barwood et al. 2014). This relationship between ƒR and V T is similar to that seen between heart rate and stroke volume at high heart rates. On immersion in cold water, the relative increase above resting levels is greater for heart rate than stroke volume (Tipton, 1989). This suggests that in such stressful situations the rate, rather than depth, of the cardiac and respiratory pumps is primary activated. Indeed, it has been concluded that ƒR is a better analogue of respiratory drive on initial immersion in cold water than V T (Tipton et al. 1991).
Prompted by the initial gasp response, part of the cold shock response is an inspiratory shift in end‐expiratory lung volume (Tipton, 1989). As mentioned, different patterns of ƒR and V T are influenced by the starting lung volume (Fig. 1) and rate of rise of neural output. The higher the lung volume and the faster the rate of rise of neural output, the smaller the V T and the higher the ƒR (Younes & Remmers, 1981). Thus, the inspiratory shift in end‐expiratory lung volume resulting from the cold shock gasp response pre‐sets the respiratory system neuronally and mechanically for a frequency driven response with high ƒR and small V T.
After the first 2 min of immersion, and with the attenuation of the cold shock response, the respiratory responses become driven by thermoregulatory demands as represented by shivering, an involuntary form of skeletal muscle exercise (see Fig. 2 B). Thus, the ventilatory stimulus alters from rapid skin cooling (‘thermal – dynamic’) to ‘thermally evoked metabolic’. With this change comes a consequent diminution of the contribution of ƒR to and an increasing contribution of V T. Hayward & Eckerson (1984) report an average 434% increase in ventilation rate on immersion in cold water (0°C), but no differences in the responses of lightly clothed males and females. However, from 10–30 min of exposure to cold water the in males was, on average, 20% greater than that of females, with at this time being primarily driven by the metabolic demands of shivering, and the increase in being predominantly caused by increased V T (Hayward & Eckerson, 1984). Similarly, during 120 min exposure to 10°C cold air, the increased in men resulted from a greater V T (Wagner & Horvath, 1985). The increases in during cold air exposures at 10 and 15°C in women were solely due to increases in V T; however, in men the increases in resulted from increases in V T at 15°C and both V T and ƒR at 10°C (Wagner & Horvath, 1985).
Frank et al. (1997) induced mild deep body hypothermia in humans by intravenous infusion of 30 or 40 ml kg−1 of cold saline (4°C) on two separate days. V T increased by between 60% and 170% on completion of the cold infusion, ƒR was unchanged. Similarly, when data for a 2 h cold air exposure at either 10, 15 or 20°C were averaged, Wagner & Horvath (1985) found that although increased, probably as a result of shivering, only V T increased; there was no change, or a slight reduction, in ƒR.
The surface area exposed and insulation of any clothing worn during cold exposure may also be relevant and help to describe some of the findings presented in Fig. 2. Simonova & Kozyreva (1994) have previously demonstrated that during forearm cooling the increase in was caused by an increase in ƒR; but when the hand and face were cooled the increase in was caused by an increase in V T. Whilst there are differences between the responses evoked by face compared to forearm cooing (diving response cf. cold pressor), it is more difficult to explain why hand and forearm cooling should produce such a difference in the ventilatory response. It is possible that the density of peripheral thermoreceptors located around the face is partially responsible for these observations. However, research in this area is sparse.
These findings, and the data presented in Fig. 1, demonstrate a clear variation in the way is increased on cold exposure depending on the nature (duration, location and rate (intensity) of cooling) of that exposure. Sudden cooling of the skin on immersion, sufficient to evoke a large dynamic response of the cutaneous cold receptors, results in an increase in primarily as a consequence of an increase in ƒR (Fig. 2 A). Longer exposures to cold and associated deep body cooling results in a similar response to that seen with exercise; an increase in due to a combination of increases in V T and ƒR (Fig. 2 B). This is not unreasonable given that shivering is a form of skeletal muscle exercise with similar metabolic and chemical consequences and, therefore, requirements for .
Hyperthermia and exercise in the heat
Over 100 years ago Haldane (1905) established that hyperthermia is a potent stimulus for increasing . It is now well established that hyperthermia causes hyperventilation, an increase in leading to hypocapnia and cerebral hypo‐perfusion in resting and exercising humans, which can reduce cerebral blood flow by as much as 20%, potentially decreasing oxygenation and brain convective cooling (Haldane, 1905; Gaudio & Abramson, 1968; Nielsen & Nybo, 2003; Fujii et al. 2015; Hayashi, 2015; Tsuji et al. 2016). Thus, a clear difference from the thermal panting of animals is that hyperthermia‐induced hyperventilation in humans can cause a reduction in the arterial carbon dioxide tension () that is proportional to the increase in . However, the precise physiological mechanisms responsible for this hyperthermia‐induced hyperventilation in humans are poorly understood. Potential causative factors include increases in (i) rectal temperature (White, 2006), (ii) temperature of the hypothalamus (Boden et al. 2000), (iii) peripheral chemoreceptor sensitivity, possibly specific to O2 (Fujii et al. 2008a). Although hyperthermia increases peripheral chemoreceptor ventilatory O2 drive in resting humans, the relative contribution of this drive to hyperthermia‐induced hyperventilation is small (∼20%), and appears to be unaffected by increasing deep body temperature (Fujii et al. 2008a).
The variability seen in the data in Fig. 3 is due to the usual sources (sex, age, body composition, aerobic fitness) observed across most stressors. Two of the more important sources include the magnitude of the increases in deep body temperature and the co‐existence of exercise. Several studies have reported that hyperthermia‐induced increases in at rest (passive) result from increased ƒR with little or no change in V T (Vejby‐Christensen & Petersen, 1973a,b; Petersen & Vejby‐Christensen, 1977; Chu et al. 2007; Curtis et al. 2007; Fan et al. 2008). However, others have reported that hyperthermia can result in increases in ƒR and V T (Fung et al. 1994; Cabanac & White, 1995; White, 2006). The latter findings are more consistent with the data presented in Fig. 3, and with the reported relationship in humans between hyperthermia‐induced hyperventilation and reduction in noted above. These discrepancies could be due to the different methodologies and degree of hyperthermia. For example, in the studies of Cabanac & White (1995), passive heating of humans to an oesophageal temperature (T es) of 38.5°C was required for hyperthermic‐induced hyperventilation. Petersen & Vejby‐Christensen (1977) reported a deep body temperature threshold near 38°C for the onset of a relative hyperventilation. Tsuji et al. (2012b) have reported a threshold at an oesophageal temperature of about 38.2°C. Passive heating that raised T es by ∼1.5°C was not reported to influence V T (Curtis et al. 2007). Above these temperatures, gradually increased with increasing T es (Fujii et al. 2008a).
Figure 3. Increase in tidal volume and respiratory frequency in response to passive heating (A) and the combination of exercise and heat exposure (B).
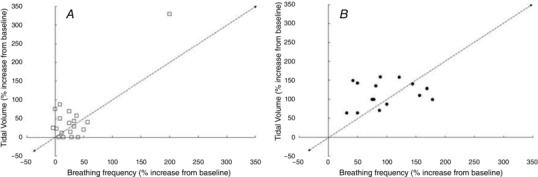
Data extracted from the following references: Cabanac & White (1995); Curtis et al. (2007, 2008b); Fan et al. (2008); Fujii et al. (2008a,b, 2012); Hayashi et al. (2006, 2011); Nelson et al. (2011, 2012); Tsuji et al. (2012a,b).
Hey et al. (1966) reported that during quiet breathing and in hyperpnoea there is a linear relation between and V t, up to a V t equal to about half of the vital capacity. Further increases in occur without further change in V t. The authors report that this relationship is essentially unaltered by wide variations in and , by metabolic acidaemia, some drugs which affect respiration, and by moderate muscular exercise. When, however, the body temperature is raised, ƒR is relatively increased.
During exercise in the heat, the identification of the thermal component of the increase in is complicated by the added demand of metabolism (Fig. 3 B). Thus, temporal changes in ventilation during exercise in the heat are likely to be directly associated with: (i) temperature per se, (ii) the type and intensity of exercise, which may produce metabolites influencing ventilation (Chu et al. 2007), or (iii) an interaction of both. Many of the factors controlling and influencing ventilation, including output from central command, input from central or peripheral chemoreceptors, input from muscle mechanoreceptors and metaboreceptors, the mechanical properties of muscle and connective tissue and the release of mediators influencing smooth muscle and bronchial tone (e.g. HSP20, E‐type prostaglandins), are influenced by temperature (Hayashi 2015; Tipton et al. 2017). As a consequence, there is a strong association between body temperature and tachypnoea during exercise. Hayashi et al. (2006) suggest that this may operate, in part, by increasing central command. Hayashi (2015) postulates that during exercise, output from central command and input from muscle mechanoreceptors and metaboreceptors via group III and IV muscle afferents modulate and contribute to hyperthermia‐induced hyperventilation. However, changes in deep body temperature cannot explain the fast response of ƒR to exercise and to variations in power output; these occur too fast to be explained by the invariably slow rise in body temperature.
Inspiratory duration has been reported to have an inverse relationship to body temperature, without directly affecting the off‐switch to inspiration threshold (Grunstein et al. 1973; Bradley et al. 1974). Animal studies have implicated descending hypothalamic projections as well as cutaneous afferents in this response (Widdicombe, 1964). Tsuji et al. (2016) report that the oesophageal temperature‐dependent increases in during light and moderate exercise occur due to increases in ƒR and in spite of falling V T. It is likely that alterations in cerebral blood flow during exercise impact upon the ventilatory drive, as reduced cerebral blood flow will activate thermoregulatory responses including hyperventilation; this will be further driven by CO2 accumulation in the brain stimulating central chemoreceptors (Tsuji et al. 2016).
Six consecutive days of heat acclimation (37°C, 50% relative humidity) has been shown to have no effect on V T or ƒR at rest or during an exercise‐heat test (Fujii et al. 2012), and dehydration with maintained mean arterial pressure (MAP) has no effect on absolute V T or ƒR, or the T es‐dependent slope changes of V T or ƒR (Fujii et al. 2008b). Fujii et al. (2008a) have also demonstrated that is reduced by hyperoxia, induced by breathing 100% O2 (an effect attributed to a dis‐facilitation of carotid body chemoreceptors), and that the absolute changes in gradually increase with increasing T es. These findings suggest that the ventilatory drive from peripheral chemoreceptors increases with increased deep body temperature in resting normoxic conditions. In both animal and human studies, hypoxic and hyperthermic stimuli have been shown to positively interact (Natalino et al. 1977; Eyzaguirre & Zapata, 1984; Chu et al. 2007; Curtis et al. 2007) in terms of their influence on resting or exercise ventilation. The direct influence of temperature on chemosensitive tissues may help explain this positive interaction between hypoxia and hyperthermia (Natalino et al. 1977; Chu et al. 2007; Petersen & Vejby‐Christensen, 1977).
It is concluded that, in humans, increases in ƒR and V T appear to be responsible for the higher during passive hyperthermia (denoted by the square symbols; Fig. 3 A). However, when deep body temperature is increased by approximately 1.5–2°C, or reaches 38.2–38.5°C, i.e. when the stress becomes more intense, ƒR appears to play an increasing role, perhaps as part of a vestigial panting response?
Hypoxia
Blood gases are controlled via central and peripheral respiratory chemoreceptor responses with different gains and delays. The slower acting central chemoreceptors are located throughout the brain stem (Nattie & Li, 2009) and respond to increases in brain tissue CO2/[H+]. The faster responding peripheral chemoreceptors (carotid bodies) are located at the bifurcation of the common carotid arteries and respond to low arterial O2 and high arterial CO2. The O2–CO2 stimulus interaction is multiplicative and results in an augmented carotid body response in hypoxia and a blunted CO2 response in hyperoxia (Lahiri et al. 1987); however, over time the ventilatory response to CO2 is restored in humans and other species (Dahan et al. 2007; Forster et al. 1971; Dempsey et al. 2014).
On ascent to high altitude, there is an immediate increase in , known as the hypoxic ventilatory response (HVR; Schoene, 2001). This increase is mediated primarily by the carotid bodies (∼90%) and partially by the aortic bodies (∼10%) (Heymans et al. 1930; Timmers et al. 2003). Carotid and aortic bodies contain glomus (type I) cells, which release neurotransmitters in response to hypoxia, causing depolarization of nearby afferent nerve endings (Prabhakar, 2000; Timmers et al. 2003). In humans, glomic tissues other than the peripheral chemoreceptors, such as the glomus trigeminale, the glomus jugulare, the glomus pulmonare and the intrapulmonary neuroepithelial bodies, serve no discernible role in ventilatory responsiveness to hypoxia (Whipp, 1984). The carotid bodies therefore play a vital role in constraining the fall of arterial in response to the challenge of hypoxaemia, and carotid chemoreceptor denervation leads to permanent abolition of normocapnic ventilatory responses to hypoxia, and reduced ventilatory responses to hypercapnia (Timmers et al. 2003).
Mekjavic et al. (1987a) studied breathing during rest and incremental exercise in normoxic and hypoxic ( = 0.12) environments. They reported that and mean inspiratory flow (V T/T I) increased with exercise intensity and were augmented in the hypoxic environment. The ratio between T I and total breath durations increased with exercise intensity, but was unaffected by hypoxia. V T and T I had linear, coinciding ranges in normoxic and hypoxic conditions up to V T/T I values of about 2.5 l s−1. At higher values T I continued to decrease, whilst V T tended to level off, an effect which was more evident in the hypoxic condition. The authors suggest that the hypoxic augmentation of exercise hyperponea is primarily brought about by an enhancement of central inspiratory drive, with the timing component being largely unaffected by the hypoxic environment. At low to moderate levels of exercise hyperpnoea inspiratory off‐switch mechanisms are essentially unaffected by moderate hypoxia. This may explain why the relatively low metabolic demand of shivering thermogenesis is unaffected by normobaric hypoxia (Blatteis & Lutherer, 1976).
Given that hypoxia is a chemical stimulus requiring, in part, increased for its correction, an increase in via V T on exposure to altitude can be regarded as an appropriate response. In support of this, Mateika et al. (2004) reported an increase in V T with no change in ƒR during intermittent exposure to hypoxia.
The increased ventilatory drive during exposure to hypoxia, when in an awake state, appears to be primarily linked to an increase in V T in humans (Fig. 4). The hyperventilation induced by the HVR results in hypocapnia which ‘brakes’ the ventilatory response.
Figure 4. Increase in tidal volume and respiratory frequency in response to hypoxic exposure.
Filled triangle represents individuals who required supplementary oxygen and the filled square represents those who did not require supplementary oxygen at altitude (5200 m) (Bernardi et al. 2006). Data extracted from the following references: Berssenbrugge et al. (1983); Easton & Anthonisen (1988); Georgopoulos et al. (1989); Morelli et al. (2004); Bernardi et al. (2006); Curtis et al. (2007); Savourey et al. (2007); Faiss et al. (2013); Richard et al. (2014); Puthon et al. (2016).
It seems that, as with other stressors, the severity of the hypoxic stress appears to influence the pattern of the breathing response. During severe hypoxia a pronounced increase in ƒR and decrease in V T has been reported, resulting in either a decrease (Gautier & Bonora, 1980) or increase (Miller & Tenney, 1975), in (for a review see Teppema & Dahan, 2010). Persistent 5% hypoxia will arrest breathing (eupnoea) and trigger gasping – the latter being an intrinsic auto‐resuscitative mechanism. If hypoxia persists then gasping stops in adults but not neonates (St‐John, 1998). With regard to the effects of CNS hypoxia on ventilatory control, central respiratory depression has been reported (Schmidt et al. 1995). However, different respiratory‐related neurons in the medulla or hypothalamus have been observed to both depolarize and hyperpolarize in response to physiological levels of hypoxia. The results obtained appear to be critically dependent on such methodological factors as state of consciousness, whether the carotid chemoreceptors are intact and the duration of hypoxia (Bisgard & Neubauer, 1994; Curran et al. 2000). However, when the response observed to hypoxia of any intensity or duration is an increase in ventilation, it must be concluded that the net effect is excitatory.
Bernardi et al. (2006) reported that successful high altitude climbers, who could reach the highest summits without oxygen, were characterised by a lower ventilatory sensitivity to hypoxia and a slower ƒR, compared to those who did not succeed or needed supplemental oxygen. These findings were supported by Puthon et al. (2016) who reported that ƒR was larger in control participants compared with elite high‐altitude climbers, while V T was larger in climbers compared with controls during maximal incremental cycling tests in both normoxic and hypoxic conditions.
Schoene et al. (1984) have previously demonstrated that the drop in arterial oxygen saturation that occurs with exercise at altitude is inversely related to HVR, and sojourners with high HVR may perform better at extreme altitude. From Fig. 4 it is notable that those requiring supplemental oxygen during 15 days at 5200 m (filled triangle) increased by an increase in ƒR. Those not requiring supplemental oxygen (filled square) increase by an equivalent increase in ƒR and V T. Faulhaber et al. (2014) reported that individuals susceptible to acute mountain sickness (AMS) have a higher ƒR and smaller V T at rest in acute hypoxia compared with those not susceptible to AMS. Petersen & Vejby‐Christensen (1977) concluded that those with a respiratory control system that is particularly sensitive to CO2 and/or hypoxia tend to increase by a relatively larger increase in ƒR compared to V T. Why these differing responses are observed is a matter of conjecture; perhaps they represent the difference between an adaptive (chemically driven) response to altitude and a maladaptive (stress driven) response, underpinned by individual differences in the sensitivity of the chemoreceptors?
Whilst these somewhat disparate findings are difficult to interpret, they share one common theme: those who perform better at altitude or suffer less AMS, tend to increase their by V T rather than ƒR and thus their should be better for a given amount of respiratory effort. As mentioned, the work of breathing and consequent heat production and oxygen demand by the respiratory muscles are higher per unit of if is increased by ƒR rather than V T (more breaths required for the same if ƒR is major route for the increase in ).
It is well established that hypoxic stimuli have an interactive effect on multiple superimposed stimuli including, but not limited to: exercise; CO2; medication; narcotics; alcohol; and various disease states such as asthma and chronic obstructive pulmonary disease (Caruana‐Montaldo et al. 2000). Similarly, different responses are seen during exposure to hypoxia when sleeping (Berssenbrugge et al. 1983; Smith et al. 2007). Severe normobaric (NH) and hypobaric hypoxia (HH) also have different effects on ventilatory drive (Richard & Koehle, 2012). Savourey et al. (2003, 2007) observed a greater V T in NH compared to HH, while ƒR tended to be lower for a given in NH as opposed to HH. ƒR was also significantly higher in HH than NH after 5 min, but reached similar values in both conditions at the end of the 40 min of exposure. Richard & Koehle (2012) also demonstrated that V T was higher in NH than HH at 5 min and 30 min, and greater than in normobaric normoxia (NN) over an average of 5 h; however, no differences between conditions were reported in ƒR.
Research examining sex differences in the ventilatory response to hypoxia in healthy, awake humans has provided somewhat conflicting results (Aitken et al. 1986; Van Klaveren & Demedts, 1998; Tarbichi et al. 2003; Morelli et al. 2004). It is likely that variations in age, body morphology, menstrual cycle stage and inter‐individual variability may have contributed to these findings; however, studies in awake animals have also produced similar contradictory data (Morelli et al. 2004).
Panic and pain
When conscious, in addition to the brainstem, many mid‐ and forebrain regions are also involved in breathing, including corticostriatal circuitry (Critchley et al. 2015). Humans can voluntarily override automatic respiratory pattern generation when playing wind instruments, speaking or swallowing, for example. Emotion and behaviour can also affect respiratory motor output through mechanisms that are not influenced by metabolic demands (Homma & Masaoka, 2008). Indeed, psychophysiological stress‐induced respiratory reactions produce changes in ventilation which often precede changes in metabolic processes (Suess et al. 1980).
Unpleasant emotions such as panic or fear result in deviations from eupnoea that are distinct from those produced in response to metabolic requirements. Areas of the forebrain, limbic and cortical structures and hypothalamus have been implicated in systems that process information from the external environment and directly stimulate spinal respiratory motor neurones (Masaoka et al. 2005; Homma & Masaoka, 2008). Additionally, the sensation of pain (defined as an unpleasant sensory experience associated with actual or potential tissue damage) has been shown to affect the respiratory cycle through a mixture of behavioural response and a direct effect on medullary respiratory centres (Sarton et al. 1997; Ward, 2002). It should be noted that any increase in ventilation due anxiety, panic or pain will decrease and thereby the drive to breathe.
Panic
Panic is defined as sudden uncontrollable or overwhelming fear or anxiety. Although centrally mediated, panic can be evoked by a wide range of real or apparent ‘threats’. The most consistent physiological change observed during laboratory‐induced panic attacks is hyperventilation (Niccolai et al. 2009).
Authors have identified areas of the limbic system that have a central role in fear and panic responses (Homma & Masaoka, 2008). In particular, the temporal pole is associated with evaluation of an uncertain or dangerous environment, and the amygdala processes negative emotions such as anxiety and fear (Halgren et al. 1978; Reiman et al. 1989; Davis, 1992; Masaoka & Homma, 2004). The involvement of the temporal lobe is consistent with patients with focal temporal lobe epileptic seizures who experience pre‐ictal fear or anxiety (Masaoka & Homma, 2004). In human studies, electrical activity in the amygdala was increased during anticipatory anxiety and Pavlovian fear‐conditioning; the electrical activity was synchronous with the onset of inspiration and often occurred before the patient reported experiencing symptoms of panic or anxiety (Le Doux, 1998; Masaoka & Homma, 2004; Homma & Masaoka, 2008). Animal studies have identified an interruption of normal respiratory cycle and an increase in ƒR following electrical stimulation of the amygdala (Harper et al. 1984).
Carnevali et al. (2013), using rats selectively bred for high or low anxiety behaviour, have reported elevated ƒR responses, reduced sniffing and increased incidence of sighs in novel environments, anxiety‐inducing restraint and acoustically stressful stimuli in high anxiety rats, with no habituation to repetitive stimuli. The authors conclude that these responses resemble those observed in anxious and panic patients, and that respiratory pattern may represent a promising method for assessing anxiety states in pre‐clinical investigation. Human individual responses vary depending on learned emotional reactions; an increase in ƒR has been associated with individual trait anxiety scores (Masaoka & Homma, 2000); individuals with high anxiety scores have large increases in ƒR compared to those that are not anxious. Variation in the responses of individuals result from the interplay of factors such as memory, cognition and emotion (Masaoka & Homma, 2000; Tracey & Mantyh, 2007).
There may be a significant evolutionary reason for the relationship between emotion and respiration. The olfactory system is the only sensory modality that is not processed by the thalamus. Instead, information ascends directly to the limbic system (Masaoka et al. 2005). Direct stimulation of the olfactory limbic system can alter respiratory patterns in humans, with unpleasant odours increasing ƒR and reducing V T (similar to panting). Perception of odour is dependent on aspiration (sniffing) and it is therefore possible that the relationship between the limbic system and respiration has a role in the detection of danger and evolutionary defence mechanisms (Homma & Masaoka, 2008).
Cunningham & Gardner (1972, 1977) and Gardner (1977) remarked upon the possible shortening of resting expiratory time (T E) due to anxiety in their subjects. Rigg et al. (1977) reported that and ƒR increased significantly on performance of mental arithmetic during both air breathing and rebreathing, whilst V T was unchanged.
From Fig. 5 it appears that the response to ‘panic’ is primarily achieved by an increase in V T. However, it is important to note that in the studies presented in Fig. 5 panic was often (18 out of 22 studies) induced by carbon dioxide inhalation; this will evoke the respiratory response appropriate for hypercapnia (increase alveolar ventilation via an increase in V T) rather than panic per se. With a progressive increase in hypercapnia V T increases first, ƒR does not increase until hypercapnia reaches a significant level, around half the vital capacity (Gelfand & Lambertsen, 1973; Duffin et al., 2000). Hypercapnia in the intact animal augments V T and expiratory flow rate. Hypercapnia and hypoxia cause a prominent decrease in spontaneous T E, so that a larger V T is expired in a shorter period (Rebuck et al. 1976), in part due to the recruitment of expiratory muscles (Gaitier et al. 1973). Inverse proportionalities have been observed between V T and T I and T E during CO2 inhalation for both the mean values at differing steady states (Cunningham et al. 1972), and breath‐by‐breath values during rebreathing (Clark & von Euler, 1972). During rapid changes in stimulation, these proportionalities may be altered due to differing response times for the changes in V T, T I and T E. Hypercapnia independently inhibits the respiratory off‐switch mechanism (increases its threshold) while increasing the rate of rise of inspiratory activity (von Euler & Trippenbach, 1976). Thus, whilst CO2 exposure causes a robust fear response and panic symptom ratings in healthy volunteers and panic disorder patients (Leibold et al. 2016), it is not valid for looking at the ventilatory response to panic. Such studies are more appropriate for investigating the ventilatory response to hypercapnia.
Figure 5. Increase in tidal volume and respiratory frequency in response to ‘panic’ induced by CO2 inhalation (hypercarbia).
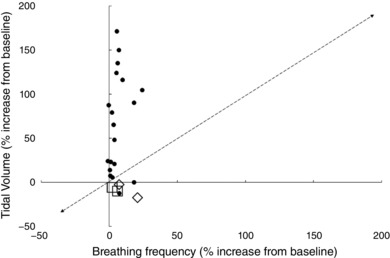
Open diamonds represent anxiety associated with anticipatory electric shock (Masaoka & Homma, 2001) and open squares represent social anxiety associated with being touched by an individual (Wilhelm et al. 2001). Data extracted from the following references: Papp et al. (1997); Pine et al. (1998, 2000); Gorman et al. (2000); Masaoka & Homma (2001); Wilhelm et al. (2001); Bailey & Holt (2002); Hoit et al. (2007).
The studies that induced panic by social anxiety (male touching the wrist of a female) or fear conditioning using unpleasant sensory stimuli, show a small increase in ƒR and a reduction in V T with remaining unchanged (Wilhelm et al. 2001). An increase in ƒR is seen with anxiety produced by breathlessness and mechanical discomfort (Gorman et al. 2000). Such an increase is apparent in patients that panic following respiratory challenge, as well as those exposed to other sensory stimuli and anticipatory anxiety. Cohorts that panic following audio‐visual, olfactory, touch or electric shock conditioning do not exhibit an increase in V T (Masaoka & Homma, 1997; Masaoka & Homma, 2001; Wilhelm et al. 2001).
It is concluded that the ventilatory pattern evoked by panic is confused by the method often used to evoke it (CO2 inhalation – illustrated by the filled circles in Fig. 5) which preferentially increases V T rather than ƒR. Those human studies which have not been confounded by hypercapnia (Rigg et al. 1977; Gorman et al. 2000; Masaoka & Homma, 2000; Wilhelm et al. 2001) and animal studies (Harper et al. 1984; Carnevali et al. 2013), have often reported an increase in ƒR in response to panic, particularly in individuals and animals with high baseline levels of anxiety.
Pain
‘Pain’ is a generic term that covers a range of types (e.g. somatic, visceral, neuropathic) and can be chronic or acute. Individual sensitivities to pain vary (e.g. there is a sex difference), as does the type of pain evoked by different situations and disease states (Tracey & Mantyh, 2007). Pain can be evoked by a variety of stimuli including temperature, mechanical and chemical stimuli, and the nerve fibres associated with the transmission of pain vary, including both small myelinated (Aα–Aδ) and unmyelinated (C) fibres. It is perhaps unsurprising therefore that ‘pain’ can produce a variety of responses depending on how, and in whom, it is evoked. In addition, there is a relative paucity of papers that provide details of the components of the ventilatory response to pain, those that do are presented in Fig. 6.
Figure 6. Increase in tidal volume and respiratory frequency in response to painful stimuli.
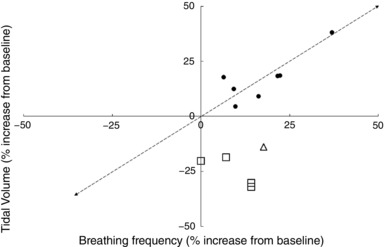
The filled circles represent data from research in which pain was induced by a combination of ischaemia and cutaneous electrical stimulation (Sarton et al. 1997). The open triangles represent pain experienced before an intercostobrachial nerve block in hand injuries (Bourke, 1997) and the open squares represent post‐cholecystectomy operative pain (Joris et al. 1992). Data extracted from the following references: Duranti et al. (1991); Joris et al. (1992); Bourke (1997); Sarton et al. (1997).
The experience of pain calls for appraisal of the affected body area in a cognitive and emotional process. Transmission of nociceptive information occurs in several ways. The spinothalamic tract enables localisation and discrimination of pain by by‐passing respiratory centres and communicating directly with the thalamus and sensory cortex (Tracey & Dickenson, 2012). Contrastingly, the spinoreticulothalamic tract transmits through the pons and medulla to the thalamus and higher centres that make up the ‘pain matrix’ (Tracey & Mantyh, 2007). This matrix includes the amgydala and hippocampus, along with areas such as the anterior cingulate and prefrontal cortex that are responsible for giving emotional significance to a sensory stimulus (von Leupoltd et al. 2009). Willer (1975) demonstrated that the anticipation of pain was enough to increase ƒR, suggesting behavioural breathing control by higher centres may be responsible for changes in associated with acute pain.
Unfortunately, the direct role of centres that make up the pain matrix is unclear and poorly defined due to the interplay of factors such as memory, cognition and emotion (Tracey & Mantyh, 2007). This interplay results in the ‘individualised’ response to pain demonstrated in studies; higher perceived pain scores correlate with larger changes in respiratory parameters. Individualised responses are also evident in panic studies and may reflect learned emotional reactions to perceived threats in the external environment.
The spinomesencephalic tract communicates nociceptive information from the dorsal horn to the periaqueductal grey matter (PAG) and mediates the ‘fight or flight’ response (Tracey & Mantyh, 2007; Subramanian & Holstege, 2014). Acute pain can induce a wide range of sympatho‐adrenal responses that can indirectly increase (Christensen, 1991). Weissman et al. (1986) reported that the increase in following the infusion of cortisol, epinephrine and glucagon to simulate plasma levels seen in patients following trauma, was due to increased V T rather than ƒR.
A direct relationship between nociceptive afferents and respiratory networks explains the changes in with painful stimuli (Sarton et al. 1997). An increase in ƒR is often seen in surgical patients despite adequate anaesthesia (Rosenberg et al. 1980). Higher cortical centres, such as those involved in the pain matrix, are supressed by anaesthetic agents (Eger et al. 1972). Patients under deep anaesthesia still exhibit a respiratory change following surgical stimulation in the absence of cardiovascular or somatic response (Eger et al. 1972; Borgbjerg et al. 1995). This finding suggests that nociceptive information can be transmitted directly to bulbar respiratory nuclei without involving higher centres. Waldrop et al. (1984) further support this suggestion by demonstrating increased phrenic nerve activity in decerebrate cats following noxious heating of the skin. The dorsolateral PAG has a high density of opiate receptors and electrical stimulation of this area results in profound analgesia. Interestingly, stimulation of different areas of the PAG (ventrolateral) can also result in changes in the respiratory pattern, such as slow, deep breathing, tachypnoea and breath holding (Subramanian & Holstege, 2014). Due to the close proximity of networks involved in pain transmission and those responsible for respiratory control in the medulla, pons and PAG, stimulation of nociceptive afferents may cause changes in respiratory function through action on any of these structures (Sarton et al. 1997; Subramanian & Holstege, 2014). For example, Bourke (1997) studying the administration of regional anaesthesia to four patients with trauma to the upper extremities and with an average pain score of 7.1/10; reports that relief of pain was associated with decreased in all four patients.
From Fig. 6 it appears that the response to ‘pain’ is generally achieved by a combination of an increase in V T and ƒR. However, as noted above when discussing panic, it is important to note how the pain is induced. Pain that is produced by methods that alter the biochemical status of the body (ischaemia, hypoxia, hypercapnia) are likely to evoke the respiratory response appropriate for these stressors, rather than pain per se. It is interesting to note that perioperative pain seems to be associated with an increase in ƒR and fall in V T. This may involve a conscious component in situations where there is a relationship between the depth of each breath and the level of post‐operative pain.
Duranti et al. (1991) identified a decrease in T I and T E during cutaneous electrical stimulation in humans, resulting in an increase in ƒR. V T did not change, but increased with deep muscle electrical stimulation and ischaemic pain from a tourniquet; however, a larger increase in ƒR was also seen, along with an increase in (it is worth noting that use of a tourniquet with resulting ischaemia represents an additional chemically derived drive to ventilation). The authors inferred that the increase in seen during painful stimuli resulted from an increase in the drive to breathe, rather than a change in timing of components during the respiratory cycle. Sarton et al. (1997) confirmed this by describing an initial increase in ƒR and V T after induction of pain (scored 4.5 to 5.5 on a 10 point scale) by transcutaneous electrical stimulation. During prolonged painful stimuli, the V T returned to resting values whilst the ƒR remained elevated. Further testing whilst participants inhaled different CO2 and O2 gas mixes enabled authors to investigate whether pain induced a change in via the peripheral or central chemoreflexes. It was concluded that pain induced a ‘chemoreflex‐independent tonic ventilatory drive’ (Sarton et al. 1997). It was unclear whether a stronger pain stimulus would have had more of an effect.
Pain seems to increase through variations in both ƒR and V T (Duranti et al. 1991; Sarton et al. 1997; Ward, 2002). Higher cortical centres interact to produce a behavioural control of breathing that can be seen during anticipation of painful stimuli (Willer, 1975). In decerebrate unanaesthetised rats, mechanical nociceptive stimulation of a forelimb evoked abrupt increases in the rate of phrenic discharge with little effect on amplitude, whereas apnoea was evoked with the same stimulus applied to the cornea (Boscan & Paton, 2001); this illustrates the diversity of the respiratory responses evoked by pain stimuli. However, as noted, anaesthetic agents that suppress higher centres do not prevent the increase in respiratory rate resulting from surgical stimulation (Rosenberg et al. 1980). It would therefore appear that the ventilatory response to pain is complex and results from interplay of factors including direct tissue damage, neural circuits in the midbrain and brainstem, along with emotional processing and memory. Further research is required to fully understand the mechanisms by which painful stimuli affect ventilation.
In conclusion, and as with panic, some of the methods used to evoke pain (e.g. hypercapnia, ischaemia, hyperoxia) also influence by other mechanisms and are therefore not valid to investigate the influence of pain on ventilation. When they are, V T and ƒR appear to operate independently.
Discussion
At rest, the relative contributions of ƒR and V T to may be determined by energy optimisation of the neural mechanisms and respiratory muscles (to minimise the work of breathing), as well as summation of feed‐forward and feedback drives. With the onset of a stressor, differences in the relative contributions of ƒR and V T to may reflect the separate neural circuits sub‐serving the different stressors. These neural circuits clearly differ depending on whether the stimulus is psychological, chemical or acute/chronic, moderate/intense. With the stressors reviewed, as the stress becomes extreme, generally becomes increased more by ƒR than V T. However, this switch to increasing from increases in V T to increases in ƒR occurs, as described, in some situations for mechanical reasons. The contribution of mechanical and neural mechanisms to the increase in seen with exposure to each stressor examined remains to be determined.
The afferent neural pathways for pain and temperature are anatomically very close and extremes of heat and cold activate hot and cold nociceptors. Part of the ventilatory response to these stressors may arise due to direct stimulation at the spinal level of α‐motorneurones innervating the intercostal muscles and diaphragm. Neuroanatomic studies corroborate the hypothesis that ascending neurons involved in transmitting the thermoreceptor volley following cold immersion are in the pons (Hinckel & Schroder‐Rosenstock, 1981) very close to the neurons involved in respiratory phase switching. Evidence (Lumsden, 1924; St‐John et al. 1984; Fung et al. 1994; St‐John & Paton, 2004; Paton et al. 2006) suggests that the gasping and high ƒR seen with cold shock and other thermal and psychological stressors may involve pathways that are distinct from those necessary for the generation of eupnoea. Pacemakers have only been found in the pre‐Bötzinger complex of rat and mouse and they are only active during gasping; during eupnoea they are hyperpolarised (i.e. below the pacemaking membrane voltage threshold) and embedded in a synaptic network. The latter depends on reciprocal inhibitory synapses between subpopulations of inspiratory and expiratory neurones of which there are at least six types (Paton et al. 2006; Smith et al. 2009; St‐John et al. 2009).
Breathing results from a combination of complex and interacting mechanisms involving the central nervous system, peripheral afferent feedback and chemical drives (Fig. 7). The relative contributions of these mechanisms are highly dynamic and state dependent. The essential neural circuitry resides within the pons and medulla and has the capability to generate the eupnoeic pattern as well as co‐ordinate cranial and motor activities (St‐John, 1998; St‐John & Paton, 2003, 2004); separation of these neural entities produces pathological changes in inspiratory and expiratory patterns (Abdala et al. 2009b; Smith et al. 2013). For example, sequential rostro‐to‐caudal pontine transection results in apneusis (mid‐pontine transection) and gasping (ponto‐medullary transection) (St‐John, 1998; Paton et al. 2006; Smith et al. 2007).
Figure 7. The brainstem respiratory network.
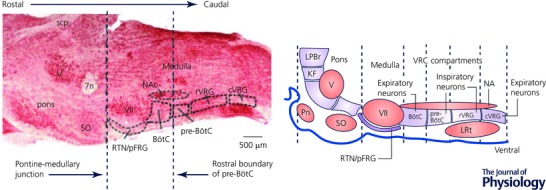
The brainstem respiratory network is organized into compartments that extend from the pons to caudal areas of the medulla. These include in the pons the Kolliker‐Fuse nucleus (KF) and the lateral parabrachial region (LPBr). In the medulla exists the ventral respiratory column (VRC) consisting of (rostral to caudal) the Bötzinger complex (BötC), pre‐Bötzinger complex (Pre‐BötC), the rostral and caudal ventral respiratory groups (r/c VRG). The VRC is located ventral to the nucleus ambiguus (NA), caudal to the facial nucleus (VII) and extends to the lateral reticular nucleus (LRt). Much interconnectivity between the Bötzinger and pre‐Bötzinger complexes is reciprocally inhibitory whereas the projections from central chemoreceptive areas coexisting within the retrotrapezoid nucleus (RTN) and the parafacial respiratory group (pFRG) are excitatory to the VRC. The KF plays a dominant role in cranial–spinal motor co‐ordination and respiratory phase switching. Visceral afferents such as lung inflation and peripheral chemoreceptor inputs relay via the nucleus tractus solitarii in the dorsomedial medulla (not shown) and connect directly or indirectly (via the pons) to the VRC. The image on the left is a parasagittal section from a rat brainstem. Abbreviations: Pn, ventral pontine nucleus; LRt, lateral reticular nucleus; SO, superior olive; Scp, superior cerebellar peduncle; V, motor nucleus of the trigeminal nerve.
Impinging on the brainstem respiratory circuitry are afferent feedback signals, most notably from vagal pulmonary inflation (stretch) receptors and peripheral arterial chemoreceptors. The former provide, via the pons, graded timing signals to off‐switch inspiration (von Euler, 1983), whereas the latter are activated by systemic hypoxia, hypercapnia and protons, and stimulate ventilation to ensure blood gas homeostasis. Lung inflation is an important mechanism for increasing respiratory frequency at the expense of reducing tidal volume; denervation of the stretch receptors results in apneusis (St‐John, 1998). Peripheral chemoreceptors also excite central chemoreceptors (Takakura et al. 2006), which reside on the ventral surface of the medulla oblongata in the retrofacial nucleus. In the latter nucleus, neurones respond to hypercapnia and their activity is a trigger for forced expiration manifested by abdominal muscle contraction (Abdala et al. 2009a; Feldman et al. 2013). Although absent at rest, forced expiration also occurs during: (i) exercise, where it plays a major role in shortening expiratory time to allow frequency to increase, and (ii) during straining and the defensive reflexes of cough, sneeze and emesis, where expiratory effort provides propulsive force.
The ultimate determinant of the pattern of ventilatory response of a particular stimulant will reside within the site(s) of termination of the activated afferent fibres, and their onward connectivity patterns (Fig. 7). There is a need for classical high resolution tract tracing to link specific afferent inputs to the respiratory network, as well as demonstration of the functional effects of activating those afferents on breathing, perhaps by adoption of optogenetics. Nevertheless, based on the known functional effects of various brain regions, it would be possible to make informed proposals as to how ƒR and V T are mediated by sensory afferent drives. Higher brain areas such as the insular cortex, amygdala and hypothalamus are known to produce changes in respiration associated with the conditioned, emotional responses that accompany affective behavioural responses (Timms, 1981; Pascoe & Kapp, 1985). These types of circuits undoubtedly co‐ordinate ‘fight or flight’ or ‘play dead’ responses capable of increasing and decreasing ƒR and V T (Applegate et al. 1983), and are mediated either via direct spinal connections to respiratory motor neurones that may bypass the brainstem respiratory network, or engage brainstem respiratory neuronal machinery.
Recent studies have demonstrated the importance of the pons, in particular the Kölliker‐Fuse (KF) nucleus, for both the normal eupoeic breathing pattern and regulation of the timing of the phase transition of respiration. Indeed, based on the patterns of respiration that can be evoked by modulating the pons and KF excitability (Chamberlin & Saper, 1994), we propose that the KF is one site for mediating changes in ƒR and V T. For example, activation of the KF slows respiration (Dutschmann & Herbert, 2006), its inhibition increases frequency (Abdala et al. 2016) whereas its opiate modulation depresses neural V T (Levitt et al. 2015). These effects are most likely mediated by orchestration of the ventral respiratory column machinery, located in the ventrolateral medulla. This core oscillating network comprises multiple types of expiratory (in the Bötzinger complex primarily) and inspiratory neurones (in the pre‐Bötzinger complex) that are inter‐connected with inhibitory synapses and output to cranial and spinal motoneurones. In isolation from the pons, including the KF, the ventral respiratory column generates apneusis and gasping (St‐John & Paton, 2004; Paton et al. 2006; Smith et al. 2007; St‐John et al. 2009). Given the range of respiratory responses that can be elicited from the ventral respiratory column when connected to the pons it is a most likely site for directly mediating ƒR and/or V T changes.
Is there clinical relevance to understanding the pattern of breathing?
ƒR and V T are altered by a variety of diseases. For example, tachypnoea, particularly during exercise, is a prominent feature in various pulmonary, cardiovascular, chest wall and neuromuscular diseases. Congestive heart failure patients can respond to exercise with tachypnoeic hyperventilation, the severity of which is prognostic of morbidity and mortality (Johnson, 2000; Olson et al. 2008; Dempsey & Smith, 2014)
We also consider changes in reflex respiratory responses following stimulation of peripheral receptors to be revealing. An example of this is the substantial exaggeration of the hypoxic ventilatory response in human patients with systolic heart failure (Ponikowski et al. 2001). Indeed, these patients have sensitisation of their peripheral chemoreceptor reflex, but when this actually occurs in relation to the onset of heart failure is not known. Thus, future research to determine the relative onset of hypoxia ventilatory response sensitivity in patients with heart failure (i.e. coincident with, or after the onset) will be important to establishing the prognostic value of this respiratory change and its consequent value in initiating early and appropriate preventative therapy.
Respiratory pattern shifts have been reported in animal experiments of lung disease, with vagal mechanisms being implicated in simulated pulmonary embolism (Guz & Trenchard, 1971), acute chemical pneumonia (Trenchard & Guz, 1972) and experimental bronchial asthma (Gold et al. 1972). The involvement of J receptors in producing rapid, shallow breathing has also been suggested (Anand & Paintal, 1980). However, the relevance of these data for humans remains, as noted in the Introduction, debatable due to species variations in vagal function (Guz et al. 1970).
In terms of human data, almost a century ago Professor J. B. S. Haldane described ‘the effects of shallow breathing’ in patients suffering from ‘soldier's heart’ (Haldane et al. 1919): now known as postural tachycardia syndrome (Schondorf & Low, 1993; Mathias et al. 2011). The condition was coined ‘soldier's heart’ owing to its frequent occurrence among military personnel in the First World War. Although equivocal, it appears that the onset of the syndrome can be linked to infection, trauma, surgery or stress (Mathias et al. 2011). The condition was characterised by breathlessness on exertion, rapid pulse, fainting attacks, giddiness, exhaustion, lassitude, headache and irritability; with the symptoms exacerbated by common stimuli in daily life, including modest exertion, food ingestion and heat (Haldane et al. 1919; Mathias et al. 2011). The breathing of patients with the condition was described as ‘remarkable’ by Haldane in the initial studies after the Great War (Haldane et al. 1919). Their respiration was rapid “from 20 to 60 or more per minute’ and shallow, ranging from ‘250 to 350 c.c. or thereabouts” and their alveolar CO2 was abnormally low (Haldane et al. 1919). Consequently, it was concluded that patients with soldier's heart were in the same state as a healthy individual at high altitudes. From this review we know this applies, in particular, to those who tend to do badly at altitude (see ‘Hypoxia’ above and Fig. 4). It is certainly revealing that almost a century later, we still have a limited understanding of the regulation of ventilation during various psychological stresses.
From a clinical perspective, we have reported in this review that the relationship between ƒR and V T may provide insights into those susceptible to AMS and perioperative pain. Indeed, a change in breathing pattern at rest whether ƒR, V T and/or forced expiratory breathing can provide prognostic information and early clinical insight into disease onset, permitting early therapeutic intervention. Either the changes in breathing pattern at rest (e.g. inspiration to expiration duration ratio), or changes to the established ƒR and V T responses to stressors, could help with diagnosis and assist in stratification of medicine. An example is the emergence of forced expiration (stage II) that accompanies development of hypertension when rats are exposed to repeated bouts of intermittent hypoxia (IH) (Zoccal et al. 2008). Also, IH is caused by central and obstructive sleep apnoea, and chronic, severe IH causes hypertension or heart failure. Thus, future research should test the hypothesis that a change in breathing pattern is indicative of the development of sleep apnoea and cardiovascular disease. Moreover, we speculate that there will be a correlation between breathing patterns and mortality and morbidity in cardiovascular disease.
Whilst we have mainly considered the relationship between ƒR and V T, the effect of dead space ventilation also has clinical significance. Measuring the dead space fraction (V D/V T) has been shown to have prognostic value in patients with acute respiratory distress syndrome (ARDS). In this fraction, V D refers to alveolar dead space resulting from alveoli that are ventilated but not perfused. The affected airways are unable to take part in gas exchange and cause arterial hypoxaemia due to right‐to‐left intrapulmonary shunting. Nuckton et al. (2002) found that for every increase of 0.05 in dead space fraction, risk of mortality from ARDS increased by 45%. Subsequent studies showed V D/V T is persistently elevated through acute, subacute and intermediate phases, and only decreases in those that survive ARDS (Kallet et al. 2004; Raurich et al. 2010). Positive end expiratory pressure (PEEP) decreases V D/V T by recruiting more of the lung, until the point that overexpansion injury occurs. Although bedside monitoring of dead space fraction is difficult, with various methods such as volumetric capnography, it could provide invaluable insight into the clinical responsiveness to treatment of ARDS, as well as identify the most severely affected individuals requiring therapeutic intervention.
Conclusions
We have examined the ventilatory response to a wide range of stressors with the aim of considering if there is an overall differential control of ƒR and V T that applies in a wide range of conditions and if so, why. It appears that for the stressors reviewed (exercise, cooling, heating, hypoxia, pain and panic), as the stress becomes more intense generally becomes increased more by ƒR than V T, regardless of whether this was the case at lower intensities of stress. We have suggested that, given the range of respiratory responses that can be elicited from the ventral respiratory column when connected to the pons, it is a most likely site for directly mediating ƒR and/or V T changes. We were tempted to speculate that the ventilatory response evoked by a stress depends on the nature of that stress: with metabolic/chemical stimuli normally resulting in an increase in both ƒR and V T; and non‐metabolic/non‐chemical intense stimuli (thermal, nociceptive, psychological) primarily resulting in an increase in ƒR. However, current understanding of the pathways involved, the complex integration, the multiple mechanical and physiological determinants of the ventilatory response and the difficulty of experimentally evoking and isolating a standardised stressor, make this suggestion impossible to test definitively at this time.
We conclude that a careful analysis of the breathing pattern of a patient might be informative diagnostically; there are likely to be associations between breathing pattern and potential causes of a disease; these could inform effective treatment strategy. In the areas of AMS, disease‐related hypoxic ventilatory responses and clinical psychology we have presented some tentative evidence to support the idea that the pattern of the ƒR and V T response of an individual may provide information regarding their physiological and pathophysiological state. Much more investigation is required in this area to determine the utility of this approach.
Although comprehensive, the current review is not systematic in nature; the data it sought were often not reported or not the main purpose of the study reviewed. Indeed, with the reporting of ƒR and V T typically neglected in the relevant scientific literature, it was difficult to draw together sufficient evidence to reach a conclusion. The data we have examined in some areas are confused by the complex nature of some of the methodologies adopted (competing stimuli, confounding factors), the duration and intensity of stimuli and individual variation. Finally, we have not considered in detail ventilation:perfusion matching differences caused by different stressors and their impact on the breathing pattern, or the respiratory muscles recruited as a result of different stressors and their impact on the breathing pattern observed; much work remains to be done in these areas.
The strength of this review is that its findings have implications for teaching, research methodology and future physiological and clinical research. The findings are not limited to one discipline, but are integrative and relevant for physiology, psychophysiology, neuroscience and pathophysiology. They highlight an important consideration that has been overlooked in some research areas and applied situations. More research is required to gain further insight into the patterns of the ventilatory responses produced by stressors and what they reveal. Such research should not limit itself to ƒR and V T, but consider all of the variables presented in Fig. 1.
Additional information
Competing interests
None declared.
Author contributions
M.J.T. conceived the review, co‐wrote the cold sections and took overall editorial control; A.H. wrote the first draft of the clinical sections and pain and panic section; J.F.R.P. wrote the neurophysiological sections; J.T.C. did the analysis of existing papers and produced the first draft of the manuscript, minus the neurophysiological and clinical sections. All authors reviewed, revised and approved the final manuscript.
Acknowledgements
The authors thank Matt Wilkes for review and Carole Tipton for her proofreading.
Biographies
Mike Tipton is Professor of Human and Applied Physiology, he is interested in the physiological and pathological integrative response of humans to extreme environments.

Abbi Harper is an Anaesthetics Trainee from Severn Deanery with an interest in Intensive Care Medicine and Extreme Environment Physiology. She is a MBBS Graduate from Newcastle University and holder of UIAA Diploma in Mountain Medicine.

Julian Paton is a Professor in Translational Cardiovascular and Respiratory physiology and uses animal models to inform novel interventional, device and pharmacological approaches for human diseases such as hypertension, sleep apnoea and heart failure.

Joseph Costello is a Senior Lecturer in exercise physiology. His current research interests are directed toward understanding the effects of various stressors (e.g. exercise, temperature and clothing) on human performance.

References
- Abdala AP, Rybak IA, Smith JC & Paton JF (2009a). Abdominal expiratory activity in the rat brainstem‐spinal cord in situ: patterns, origins and implications for respiratory rhythm generation. J Physiol 587, 3539–3559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Abdala AP, RybakH IA, Smith JC, Zoccal DB, Machado BH, St‐John WM & Paton JF (2009b). Multiple pontomedullary mechanisms of respiratory rhythmogenesis. Respir Physiol Neurobiol 168, 19–25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Abdala AP, Toward MA, Dutschmann M, Bissonnette JM & Paton JF (2016). Deficiency of GABAergic synaptic inhibition in the Kölliker‐Fuse area underlies respiratory dysrhythmia in a mouse model of Rett syndrome. J Physiol 594, 223–237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aitken ML, Franklin JL, Pierson DJ & Schoene RB (1986). Influence of body size and gender on control of ventilation. J App Physiol 60, 1894–1899. [DOI] [PubMed] [Google Scholar]
- Anand A & Paintal AS (1980) Reflex effects following selective stimulation of J receptors in the cat. J Physiol 299, 553–572. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Applegate CD, Kapp BS, Underwood MD & McNall CL (1983). Autonomic and somatomotor effects of amygdala central N. stimulation in awake rabbits. Physiol Behav 31, 353–360. [DOI] [PubMed] [Google Scholar]
- Bailey EF & Hoit JD (2002). Speaking and breathing in high respiratory drive. J Speech Lang Hear Res 45, 89–99. [DOI] [PubMed] [Google Scholar]
- Barwood MJ, Burrows H, Cessford J & Goodall S (2016). "Float first and kick for your life": Psychophysiological basis for safety behaviour on accidental short‐term cold water immersion. Physiol Behav 154, 83–89. [DOI] [PubMed] [Google Scholar]
- Barwood MJ, Corbett J, Green R, Smith T, Tomlin P, Weir‐Blankenstein L & Tipton MJ (2013). Acute anxiety increases the magnitude of the cold shock response before and after habituation. Eur J Appl Physiol 113, 681–689. [DOI] [PubMed] [Google Scholar]
- Barwood MJ, Corbett J & Wagstaff CR (2014). Habituation of the cold shock response may include a significant perceptual component. Aviat Space Environ Med 85, 167–171. [DOI] [PubMed] [Google Scholar]
- Bechbach RR, Chow HHK, Duffin J, Orsini EC (1979). The effects of hypercapnia, hypoxia, exercise and anxiety on the pattern of breathing in man. J Physiol 293, 285–300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bell HJ & Duffin J (2006). Rapid increases in ventilation accompany the transition from passive to active movement. Resp Physiol Neurobiol 152, 128–142. [DOI] [PubMed] [Google Scholar]
- Bernardi L, Schneider A, Pomidori L, Paolucci E & Cogo A (2006). Hypoxic ventilatory response in successful extreme altitude climbers. Eur Respir J 27, 165‐171. [DOI] [PubMed] [Google Scholar]
- Berssenbrugge A, Dempsey J, Iber C, Skatrud J & Wilson P (1983). Mechanisms of hypoxia‐induced periodic breathing during sleep in humans. J Physiol 343, 507–524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bisgard GE & Neubauer JA (1994). Peripheral and central effects of hypoxia In Regulation of Breathing (2nd edn), eds Dempsey JA. & Pack AI, pp. 617–668. Dekker, New York. [Google Scholar]
- Blatteis CM & Lutherer LO (1976). Effect of altitude exposure on thermoregulatory response of man to cold. J Appl Physiol 41, 848–858. [DOI] [PubMed] [Google Scholar]
- Boden AG, Harris MC & Parkes MJ (2000). The preoptic area in the hypothalamus is the source of the additional respiratory drive at raised body temperature in anaesthetised rats. Exp Physiol 85, 527–537. [PubMed] [Google Scholar]
- Borgbjerg FM, Nielsen K & Franks J (1995). Experimental pain stimulates respiration and attenuates morphine‐induced respiratory depression: a controlled study in human volunteers. Pain 64, 123–128. [DOI] [PubMed] [Google Scholar]
- Boscan P & Paton JF (2001). Role of the solitary tract nucleus in mediating nociceptive evoked cardiorespiratory responses. Auton Neurosci 86, 170–182. [DOI] [PubMed] [Google Scholar]
- Bourke DL (1997). Respiratory effects of regional anesthesia during acute pain. Reg Anesth 18, 361–365. [PubMed] [Google Scholar]
- Bradley GW, von Euler C, Marttila I & Ross B (1974). Steady state effects of CO2 and temperature on the relationship between lung volume and inspiratory duration (Hering‐Breuer threshold curve). Acta Physiol Scand 92, 351–363. [DOI] [PubMed] [Google Scholar]
- Braun SR (1990). Respiratory rate and pattern In Clinical Methods: The History, Physical, and Laboratory Examinations, eds Walker HK, Hall WD. & Hurst JW, pp. 226–230, Butterworth Publishers, Stoneham, MA, USA. [PubMed] [Google Scholar]
- Button C, Croft JL, Cotter JD, Graham MJ & Lucas SJ (2015). Integrative physiological and behavioural responses to sudden cold‐water immersion are similar in skilled and less‐skilled swimmers. Physiol Behav 138, 254–259. [DOI] [PubMed] [Google Scholar]
- Cabanac M & White MD (1995). Core temperature thresholds for hyperpnea during passive hyperthermia in humans. Eur J Appl Physiol Occup Physiol 71, 71–76. [DOI] [PubMed] [Google Scholar]
- Cannon WB (1929). Bodily Changes in Pain, Fear and Rage, 2nd edn Appleton and Company, New York. [Google Scholar]
- Carey D, Pliego G & Raymond R (2008). How endurance athletes breathe during incremental exercise to fatigue: interaction of tidal volume and frequency. J Ex Physiol 11, 44–51. [Google Scholar]
- Carnevali L, Sgoifo A, Trombini M, Landgraf R, Neumann ID & Nalivaiko E (2013) Different patterns of respiration in rat lines selectively bred for high or low anxiety. PLoS One 8, e64519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caruana‐Montaldo B, Gleeson K & Zwillich CW (2000). The control of breathing in clinical practice. Chest 117, 205–225. [DOI] [PubMed] [Google Scholar]
- Chamberlin NL & Saper CB (1994). Topographic organization of respiratory responses to glutamate microstimulation of the parabrachial nucleus in the rat. J Neurosci 14, 6500–6510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Christensen NJ (1991). The biochemical assessment of sympathoadrenal activity in man. Clin Auton Res 1, 167–172. [DOI] [PubMed] [Google Scholar]
- Chu AL, Jay O & White MD (2007). The effects of hyperthermia and hypoxia on ventilation during low‐intensity steady‐state exercise. Am J Physiol Regul Integr Comp Physiol 292, R195–R203. [DOI] [PubMed] [Google Scholar]
- Clark FJ & von Euler C (1972). On the regulation of depth and rate of breathing. J Physiol 222, 267–295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cooper KE, Martin S & Riben P (1976). Respiratory and other responses in subjects immersed in cold water. J Appl Physiol 40, 903–910. [DOI] [PubMed] [Google Scholar]
- Critchley HD, Nicotra A, Chiesa PA, Nagai Y, Gray MA, Minati L & Bernardi L (2015). Slow breathing and hypoxic challenge: cardiorespiratory consequences and their central neural substrate. PLoS One 10, e0127082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Croft JL, Button C, Hodge K, Lucas SJ, Barwood MJ & Cotter JD (2013). Responses to sudden cold‐water immersion in inexperienced swimmers following training. Aviat Space Environ Med 84, 850–855. [DOI] [PubMed] [Google Scholar]
- Cunningham DJC (1974). Integrative aspects of the regulation of breathing: a personal view In MTP Int Rev Sci Physiol, series 1, vol. 2, Respiration, ed. Widdicombe JG, p. 310 Butterworths, London. [Google Scholar]
- Cunningham DJC & Gardner WN (1972). The relationship between tidal volume and inspiratory and expiratory times during steady‐state CO2 inhalation in man. J Physiol 227, 50P–51P. [PubMed] [Google Scholar]
- Cunningham DJC & Gardner WN (1977). A quantitative description of the pattern of breathing during steady‐state CO2 inhalation in man, with special emphasis on expiration. J Physiol 272, 613–632. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cunningham DJC, Pearson SB & Gardner WN (1972). Regulation of respiratory frequency and tidal volume at various ventilations. Arch Fisiol 69, 443–446. [Google Scholar]
- Curran AK, Rodman JR, Eastwood PR, Henderson KS, Dempsey JA & Smith CA (2000) Ventilation responses to specific CNS hypoxia in sleeping dogs. J Appl Physiol 88, 1840–1852. [DOI] [PubMed] [Google Scholar]
- Curtis AN, Walsh ML & White MD (2007). Influence of passive hyperthermia on human ventilation during rest and isocapnic hypoxia. Appl Physiol Nutr Metab 32, 721–732. [DOI] [PubMed] [Google Scholar]
- Dahan A, Nieuwenhuijs D & Teppema L (2007). Plasticity of central chemoreceptors: effect of bilateral carotid body resection on central CO2 sensitivity. PLoS Med 4, e239 https://doi.org/10.1371/journal.pmed.0040239 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Datta A & Tipton M (2006). Respiratory responses to cold water immersion: neural pathways, interactions, and clinical consequences awake and asleep. J Appl Physiol 100, 2057–2064. [DOI] [PubMed] [Google Scholar]
- Davis M (1992). The role of the amygdala in fear and anxiety. Annu Rev Neurosci 15, 353–375. [DOI] [PubMed] [Google Scholar]
- Davies RC, Rowlands AV & Easton RG (2009). Effect of exercise‐induced muscle damage on ventilatory and perceived exertion responses to moderate and severe intensity cycle exercise. Eur J Appl Physiol 107, 11–19. [DOI] [PubMed] [Google Scholar]
- De Morree HM, Klein C & Marcora SM (2012). Perception of effort reflects central motor command during movement execution. Psychophysiology 49, 1242–1253. [DOI] [PubMed] [Google Scholar]
- Dempsey JA, Powell FL, Bisgard GE, Blain GM, Poulin MJ & Smith CA (2014). Role of chemoreception in cardiorespiratory acclimatization to, and deacclimatization from, hypoxia. J Appl Physiol 116, 858–866. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dempsey JA & Smith CA (2014). Pathophysiology of human ventilatory control. Eur Respir J 44, 495–512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dongelmans DA, Paulus F, Veelo DP, Binnekade JM, Vroom MB & Schultz MJ (2011). Adaptive support ventilation may deliver unwanted respiratory rate‐tidal volume combinations in patients with acute lung injury ventilated according to an open lung concept. Anesthesiology 114, 1138–1143. [DOI] [PubMed] [Google Scholar]
- Duffin J (2014). The fast exercise drive to breathe. J Physiol 592, 445–451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duffin J, Mohan RM, Vasiliou P, Stephenson R & Mahamed S (2000). A model of the chemoreflex control of breathing in humans: Model parameters measurement. Respir Physiol 120, 13–26. [DOI] [PubMed] [Google Scholar]
- Duranti R, Pantaleo T, Bellini F, Bongianni F & Scano G (1991). Respiratory responses induced by the activation of somatic nociceptive afferents in humans, J Appl Physiol 71, 2440–2448. [DOI] [PubMed] [Google Scholar]
- Dutschmann M & Herbert H (2006). The Kölliker‐Fuse nucleus gates the postinspiratory phase of the respiratory cycle to control inspiratory off‐switch and upper airway resistance in rat. Eur J Neurosci 24, 1071–1084. [DOI] [PubMed] [Google Scholar]
- Easton PA & Anthonisen NR (1988). Carobin diaoxide effects on the ventilatory response to sustained hypoxia. J Appl Physiol 64, 1451–1456. [DOI] [PubMed] [Google Scholar]
- Eger EI, Dolan WM, Stevens WC, Miller RD & Way WL (1972). Surgical stimulation antagonises the respiratory depression produced by forane. Anesthesiology 36, 544–549. [DOI] [PubMed] [Google Scholar]
- Eglin CM, Butt G, Howden S, Nash T & Costello JT (2015). Rapid habituation of the cold shock response. Extrem Physiol Med 4 (Suppl. 1), A38. [Google Scholar]
- Eglin CM & Tipton MJ (2005). Repeated cold showers as a method of habituating humans to the initial responses to cold water immersion. Eur J Appl Physiol 93, 624–629. [DOI] [PubMed] [Google Scholar]
- Eyzaguirre C & Zapata P (1984). Perspectives in carotid body research. J Appl Physiol 57, 931–957. [DOI] [PubMed] [Google Scholar]
- Faiss R, Pialoux V, Sartori C, Faes C, Deriaz O & Millet GP (2013). Ventilation, oxidative stress, and nitric oxide in hypobaric versus normobaric hypoxia. Med Sci Sports Exerc 45, 253–260. [DOI] [PubMed] [Google Scholar]
- Fan J‐L, Cotter JD, Lucas RA, Thomas K, Wilson L & Ainslie PN (2008). Human cardiorespiratory and cerebrovascular function during severe passive hyperthermia: effects of mild hypohydration. J Appl Physiol 105, 433–445. [DOI] [PubMed] [Google Scholar]
- Faulhaber M, Wille M, Gatterer H, Heinrich D & Burtscher M (2014). Resting arterial oxygen saturation and breathing frequency as predictors for acute mountain sickness development: a prospective cohort study. Sleep Breath 18, 669–674. [DOI] [PubMed] [Google Scholar]
- Feldman JL, Del Negro CA & Gray PA (2013). Understanding the rhythm of breathing: so near, yet so far. Annu Rev Physiol 75, 423–452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Forster HV, Dempsey JA, Birnbaum ML, Reddan WG, Thoden J, Grover RF & Rankin J (1971). Effect of chronic exposure to hypoxia on ventilatory response to CO2 and hypoxia. J Appl Physiol 31, 586–592. [DOI] [PubMed] [Google Scholar]
- Forster HV, Haouzi P & Dempsey JA (2012). Control of breathing during exercise. Compr Physiol 2, 743–777. [DOI] [PubMed] [Google Scholar]
- Fowler WS (1950). Lung function studies IV. Postural changes in respiratory dead space and functional residual capacity. J Clin Invest 29, 1437–1438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frank, SM , Higgins MS., Fleisher LA, Sitzmann JV, Hershel R, & Breslow MJ (1997). Adrenergic, respiratory, and cardiovascular effects of core cooling in humans. Am J Physiol Regulatory Integrative Comp Physiol 41, R557–R562. [DOI] [PubMed] [Google Scholar]
- Fujii N, Honda Y, Hayashi K, Kondo N, Koga S & Nishiyasu T (2008a). Effects of chemoreflexes on hyperthermic hyperventilation and cerebral blood velocity in resting heated humans. Exp Physiol 93, 994–1001. [DOI] [PubMed] [Google Scholar]
- Fujii N, Honda Y, Hayashi K, Kondo N & Nishiyasu T (2008b). Effect of hypohydration on hyperthermic hyperpnea and cutaneous vasodilation during exercise in men. J Appl Physiol 105, 1509–1518. [DOI] [PubMed] [Google Scholar]
- Fujii N, Honda Y, Ogawa T, Tsuji B, Kondo N, Koga S & Nishiyasu T (2012). Short‐term exercise‐heat acclimation enhances skin vasodilation but not hyperthermic hyperpnea in humans exercising in a hot environment. Eur J Appl Physiol 112, 295–307. [DOI] [PubMed] [Google Scholar]
- Fujii N, Tsuji B, Honda Y, Kondo N & Nishiyasu T (2015). Effect of short‐term exercise‐heat acclimation on ventilatory and cerebral blood flow responses to passive heating at rest in humans. J Appl Physiol 119, 435–444. [DOI] [PubMed] [Google Scholar]
- Fung ML, Wang W & St‐John WM (1994). Medullary loci critical for expression of gasping in adult rats. J Physiol 480, 597–611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gaitier H, Remmers JE & Bartlett D Jr (1973). Control of the duration of expiration. Resp Physiol 18, 205–221. [DOI] [PubMed] [Google Scholar]
- Gardner WN (1977). The relation between tidal volume and inspiratory and expiratory times during steady‐state carbon dioxide inhalation in man. J Physiol 272, 591–611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gaudio R Jr & Abramson N (1968). Heat‐induced hyperventilation. J Appl Physiol 25, 742–746. [DOI] [PubMed] [Google Scholar]
- Gaultier C, Amiel J, Dauger S, Trang H, Lyonnet S, Gallego J & Simonneau M (2004). Genetics and early disturbances of breathing control. Pediatr Res 55, 729–733. [DOI] [PubMed] [Google Scholar]
- Gautier H & Bonora M (1980). Possible alterations in brain monoamine metabolism during hypoxia‐induced tachypnea in cats. J Appl Physiol 49, 769–777. [DOI] [PubMed] [Google Scholar]
- Gefand R & Lambertsen C (1973). Dynamic respiratory response to abrupt change of inspired CO2 at normal and high PO2 . J Appl Physiol 35, 903‐913. [DOI] [PubMed] [Google Scholar]
- Georgopoulos D, Berezanski D & Anthonisen NR (1989). Effects of CO2 breathing on ventilatory response to sustained hypoxia in normal adults. J Appl Physiol 66, 1071–1078. [DOI] [PubMed] [Google Scholar]
- Gold WM, Kessler GF & Yu DYC (1972). Role of vagus nerves in experimental asthma in allergic dogs. J Appl Physiol 33, 719–725. [DOI] [PubMed] [Google Scholar]
- Goode RC, Duffin J, Miller R, Romet TT, Chant W & Ackles A (1975). Sudden cold water immersion. Respir Physiol 23, 301–310. [DOI] [PubMed] [Google Scholar]
- Gorman JM, Kent J, Martinez J, Browne S, Coplan J & Papp L (2000). Physiological changes during carbon dioxide inhalation in patients with panic disorder, major depression, and premenstrual dysphoric disorder: evidence for a central fear mechanism. Arch Gen Psychiatry 58, 125–131. [DOI] [PubMed] [Google Scholar]
- Grunstein M, Younes M & Milic‐Emili J (1973). Control of tidal volume and respiratory frequency in anaesthetized cats. J Appl Physiol 35, 463–476. [DOI] [PubMed] [Google Scholar]
- Guz A, Noble MIM, Eisele JH & Trenchard D (1970). The role of vagal inflation reflexes in man and other animals In Ciba Foundation Symposium on Breathing: Hering‐Breuer Centenary Symposium, ed. Porter R, pp. 17–40. Churchill, London. [Google Scholar]
- Guz A & Trenchard D (1971). The role of nonmyelinated vagal afferent fibers from the lungs in the genesis of tachypnea in the rabbit. J Physiol 213, 345–371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haldane JS (1905). The influence of high air temperatures. J Hyg 5, 494–513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haldane JS, Meakins JC & Priestley JG (1919). The effects of shallow breathing. J Physiol 52, 433–453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Halgren E, Walter R, Cherlow D & Crandall P (1978). Mental phenomema evoked by electrical stimulation of the human hippocampal formation and amygdala. Brain 101, 83–117. [DOI] [PubMed] [Google Scholar]
- Harper RM, Frysinger RC, Trelease RB & Marks JD (1984). State‐dependant alternation of respiratory cycle timing by stimulation of the central nucleus of the amygdala. Brain Res 306, 1–8. [DOI] [PubMed] [Google Scholar]
- Hayashi, K (2015). Ventilatory response to increasing body temperature: characteristics and effect on central fatigue. J Phys Fitness Sports Med 4, 143–149. [Google Scholar]
- Hayashi K, Honda Y, Miyakawa N, Fujii N, Ichinose M, Koga S, Kondo N & Nishiyasu T (2011). Effect of CO2 on the ventilatory sensitivity to rising body temperature during exercise. J Appl Physiol 110, 1334–1341. [DOI] [PubMed] [Google Scholar]
- Hayashi, K , Honda Yasushi, Ogawa Takeshi, Kondo Narihiko & Nishiyasu Takeshi (2006). Relationship between ventilator response and body temperature during prolonged submaximal exercise. J Appl Physiol 100, 414–420. [DOI] [PubMed] [Google Scholar]
- Hayward JS & Eckerson JD (1984). Physiological responses and survival time prediction for humans in ice water. Aviat Space Environ Med 55, 206–212. [PubMed] [Google Scholar]
- Hess A, Yu L, Klein I, De Mazancourt M, Jebrak G, Mal H, Brugière O, Fournier M, Courbage M, Dauriat G, Schouman‐Clayes E, Clerici C & Mangin L (2013). Neural mechanisms underlying breathing complexity. PLoS One 8, e75740. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hey EN, Lloyd BB, Cunningham DJC, Jukes MGM & Bolton DPG (1966). Effects of various respiratory stimuli on the depth and frequency of breathing in man. Resp Physiol 1, 193–205. [DOI] [PubMed] [Google Scholar]
- Heymans C, Bouckhaert JJ & Dautrebande L (1930). Sinus carotidien et reflexes respiratoires II. Influences respiratoires reflexes de l'acidose, de l'alcalose, de l'anhydride carbonique, de l'ion hydrogene et de l'anoxeme. Sinus carotidiens et echanges respiratores dans les poumons et au dela des poumons. Arch Intern Pharmacodyn 39, 400–448. [Google Scholar]
- Hinckel P & Schroder‐Rosenstock K (1981). Responses of pontine units to skin‐temperature changes in the guinea‐pig. J Physiol 314, 89–94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoit JD, Lansing RW & Perona KE (2007). Speaking‐related dyspnea in healthy adults. J Speech Lang Hear Res 50, 361–374. [DOI] [PubMed] [Google Scholar]
- Homma I & Masaoka Y (2008). Breathing rhythms and emotions. Exp Physiol 93, 1011–1021. [DOI] [PubMed] [Google Scholar]
- Jansen ASP, Nguyen XV, Karpitskiy V, Mettenleiter TC & Loewy AD (1995). Central command neurons of the sympathetic nervous system: basis of the fight‐or flight response. Science 270, 644–646. [DOI] [PubMed] [Google Scholar]
- Johnson RL Jr (2000) Gas exchange efficiency in congestive heart failure. Circulation 101, 2774–2776. [DOI] [PubMed] [Google Scholar]
- Joris J, Cigarini M, Legrand M, Jacquet N, De Groote D, Franchimont P & Lamy M (1992). Metabolic and respiratory changes after cholecystectomy performed via laparotomy or laparoscopy. Br J Anaesth 69, 341–345. [DOI] [PubMed] [Google Scholar]
- Kallet RH, Alonso JA, Pittet JF & Matthay MA (2004). Prognostic value of the pulmonary dead‐space fraction during the first 6 days of acute respiratory distress syndrome. Respiratory Care 49, 1008–1014. [PubMed] [Google Scholar]
- Keatinge WR & Evans M (1961). The respiratory and cardiovascular response to immersion in cold and warm water. Quart J Exp Physiol 46, 83–94. [DOI] [PubMed] [Google Scholar]
- Keatinge WR & Nadel JA (1965). Immediate respiratory response to sudden cooling of the skin. J Appl Physiol 20, 65–69. [DOI] [PubMed] [Google Scholar]
- Koglin L & Kayser B (2013). Control and sensation of breathing during cycling exercise in hypoxia under naloxone: a randomised controlled crossover trial. Extreme Physiol Med 2, 1; http://extremephysiolmed.biomedcentral.com/articles/10.1186/2046-7648-2-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lahiri S, Mulligan E, Andronikou S, Shirahata M & Mokashi A (1987). Carotid body chemosensory function in prolonged normobaric hyperoxia in the cat. J Appl Physiol 62, 1924–1931. [DOI] [PubMed] [Google Scholar]
- Le Doux JE (1998). The Emotional Brain: The Mysterious Underpinnings of Emotional Life. Simon and Schuster, New York. [Google Scholar]
- Leibold NK, van den Hove DL & Viechtbauer W (2016). CO2 exposure as translational cross‐species experimental model for panic. Transl Psychiatry 6, e885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levitt ES, Abdala AP, Paton JF, Bissonnette JM & Williams JT (2015). μ Opioid receptor activation hyperpolarizes respiratory‐controlling Kölliker‐Fuse neurons and suppresses post‐inspiratory drive. J Physiol 593, 4453–4469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lumsden T (1924). Effects of bulbar anaemia on respiratory movements. J Physiol 59, 1vii–1x.16993675 [Google Scholar]
- Lunt HC, Corbett J, Barwood M & Tipton MJ (2011). Cycling cadence affects heart rate variability. Physiol Measurement 32, 1133–1145. [DOI] [PubMed] [Google Scholar]
- Mantoni T, Belhage B, Pedersen LM & Pott FC (2007). Reduced cerebral perfusion on sudden immersion in ice water: a possible cause of drowning. Aviat Space Environ Med 78, 374–376. [PubMed] [Google Scholar]
- Mantoni T, Rasmussen JH, Belhage B & Pott FC (2008). Voluntary respiratory control and cerebral blood flow velocity upon ice‐water immersion. Aviat Space Environ Med 79, 765–768. [DOI] [PubMed] [Google Scholar]
- Marcora SM, Bosio A & Morree HM (2008). Locomotor muscle fatigue increases cardiorespiratory responses and reduces performance during intense cycling exercise independently from metabolic stress. Am J Physiol Regul Integr Comp Physiol 294, R874–R883. [DOI] [PubMed] [Google Scholar]
- Martin S & Cooper KE (1978). The relationship of deep and surface skin temperature to the ventilatory responses elicited during cold water immersion. Can J Physiol Pharmacol 56, 999–1004. [DOI] [PubMed] [Google Scholar]
- Masaoka Y & Homma I (1997). Anxiety and respiratory pattern: their relationship during mental stress and physical load. Int J Psychophysiol 27, 153–159. [DOI] [PubMed] [Google Scholar]
- Masaoka Y & Homma I (2000). The source generator of respiratory‐related anxiety potential in the human brain. Neurosci Lett 283, 21–24. [DOI] [PubMed] [Google Scholar]
- Masaoka Y & Homma I (2001). The effect of anticipatory anxiety on breathing and metabolism in humans. Respir Physiol 128, 171–177. [DOI] [PubMed] [Google Scholar]
- Masaoka Y & Homma I (2004). Amygdala and emotional breathing In Post‐genomic Perspectives in Modelling and Control of Breathing, ed. Champagnat J, pp. 9–14. Kluwer Academic/Plenum Publishers, New York. [Google Scholar]
- Masaoka Y, Koiwa N & Homma I (2005). Inspiratory phase‐locked alpha oscillation in human olfaction: source generators estimated by a dipole tracing method. J Physiol 566, 979–997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mateika JH, Mendello C, Obeid D & Badr MS (2004). Peripheral chemoreflex responsiveness is increased at elevated levels of carbon dioxide after episodic hypoxia in awake humans. J Appl Physiol 96, 1197–1205. [DOI] [PubMed] [Google Scholar]
- Mathias CJ, Low DA, Iodice V, Owens AP, Kirbis M & Grahame R (2011). Postural tachycardia syndrome – current experience and concepts. Nat Rev Neurol 8, 22–34. [DOI] [PubMed] [Google Scholar]
- Mead J (1960). Control of respiratory frequency. J Appl Physiol 15, 325–336. [Google Scholar]
- Mekjavic IB, Eiken O, La Prairie A & Banister EW (1987a). The pattern of breathing during hypoxic exercise. Eur J Appl Physiol Occup Physiol 56, 619–622. [DOI] [PubMed] [Google Scholar]
- Mekjavic IB, La Prairie A, Burke W & Lindborg B (1987b). Respiratory drive during sudden cold water immersion. Respiratory Physiology 70, 121–130. [DOI] [PubMed] [Google Scholar]
- Miller JP, Cunningham DJC, Lloyd BB & Young JM (1974). The transient respiratory effects in man of sudden changes in alveolar CO2 in hypoxia and in high oxygen. Resp Physiol 20, 17–31. [DOI] [PubMed] [Google Scholar]
- Miller MJ & Tenney SM (1975). Hypoxia‐induced tachypnea in carotid‐deafferented cats. Respir Physiol 23, 31–39. [DOI] [PubMed] [Google Scholar]
- Morelli C, Badr MS & Mateika JH (2004). Ventilatory responses to carbon dioxide at low and high levels of oxygen are elevated after episodic hypoxia in men compared with women. J Appl Physiol 97, 1673–1680. [DOI] [PubMed] [Google Scholar]
- Murias G, Blanch L & Lucangelo U (2014). The physiology of ventilation. Respiratory Care 59, 1795–1807. [DOI] [PubMed] [Google Scholar]
- Natalino MR, Zwillich CW & Weil JV (1977). Effects of hyperthermia on hypoxic ventilatory response in normal man. J Lab Clin Med 89, 564–572. [PubMed] [Google Scholar]
- Nattie E & Li A (2009). Central chemoreception is a complex system function that involves multiple brain stem sites. J Appl Physiol 106, 1464–1466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nielsen B & Nybo L (2003). Cerebral changes during exercise in the heat. Sports Med 33, 1–11. [DOI] [PubMed] [Google Scholar]
- Nelson MD, Haykowsky MJ, Stickland MK, Altamirano‐Diaz LA, Willie CK, Smith KJ, Petersen SR & Ainslie PN (2011). Reductions in cerebral blood flow during passive heat stress in humans: partitioning the mechanisms. J Physiol 589, 4053–4064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Niccolai V, van Duinen M & Griez E (2009). Respiratory patterns in panic disorder reviewed: a focus on biological challenge tests [Review]. Acta Psychiatr Scand 120, 167–177. [DOI] [PubMed] [Google Scholar]
- Nicolo A, Bazzucchi I, Haxhi J, Felici F & Sacchetti M (2014). Comparing continuous and intermittent exercise: an “isoeffort” and “isotime” approach. PLoS One 9, e94990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nicolo A, Marcora SM & Sacchetti M (2016). Respiratory frequency is strongly associated with perceived exertion during time trials of different duration. J Sports Sci 34, 1199–1206. [DOI] [PubMed] [Google Scholar]
- Noble BJ, Metz KF, Pandolf KB & Cafarelli E (1973). Perceptual response to exercise: a multiple regression study. Med Sci Sports 5, 104–109. [PubMed] [Google Scholar]
- Nuckton TJ, Alonso JA, Kallet RH, Daniel BM, Pittet JF, Eisner MD & Matthay MA. (2002). Pulmonary dead‐space fraction as a risk factor for death in the acute respiratory distress syndrome. New Eng J Med 346, 1281–1286. [DOI] [PubMed] [Google Scholar]
- Nunn JF, Campbell EJ & Peckett BW (1959). Anatomical subdivisions of the volume of respiratory dead space and effect of position of the jaw. J Appl Physiol 14, 174–176. [DOI] [PubMed] [Google Scholar]
- Ohashi S, Izumizaki M, Atsumi T & Holmes I (2013). CO2 homeostasis is maintained in conscious humans by regulation of tidal volume, but not of respiratory rhythm. Respir Physiol Neurobiol 186, 155–163. [DOI] [PubMed] [Google Scholar]
- Olson LJ, Arruda‐Olson AM, Somers VK, Scott CG & Johnson BD (2008). Exercise oscillatory ventilation: instability of breathing control associated with advanced heart failure. Chest 133, 474–481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Papp LA, Martinez JM, Klein DF, Coplan JD, Norman RG, Cole R & de Jesus MJ (1997). Respiratory psychophysiology of panic disorder: three respiratory challenges in 98 subjects. Am J Psychiatry 154, 1557–1565. [DOI] [PubMed] [Google Scholar]
- Pascoe JP & Kapp BS (1985). Electrophysiological characteristics of amygdaloid central nucleus neurons during Pavlovian fear conditioning in the rabbit. Behav Brain Res 16, 117–133. [DOI] [PubMed] [Google Scholar]
- Paton JF, Abdala AP, Koizumi H, Smith JC & St‐John WM (2006). Respiratory rhythm generation during gasping depends on persistent sodium current. Nat Neurosci 9, 311–313. [DOI] [PubMed] [Google Scholar]
- Pearson SB & Cunningham DJC (1973). Some observations on the relation between ventilation, tidal volume and frequency in man in various steady and transient states. Acta Neurobiol 33, 177–188. [PubMed] [Google Scholar]
- Perez W & Tobin MJ (1985). Separation of factors responsible for changing breathing pattern induced by instrumentation. J Appl Physiol 59, 1515–1520. [DOI] [PubMed] [Google Scholar]
- Petersen ES & Vejby‐Christensen H (1977). Effects of body temperature on ventilatory response to hypoxia and breathing pattern in man. J Appl Physiol 42, 492–500. [DOI] [PubMed] [Google Scholar]
- Phillipson ES (1978). Control of breathing during sleep. Am Rev Respir Dis 118, 909–939. [DOI] [PubMed] [Google Scholar]
- Pine DS, Coplan JD, Papp LA, Klein RG, Martinez JM, Kovalenko P, Tancer N, Moreau D, Dummit ES 3rd, Shaffer D, Klein DF & Gorman JM (1998). Ventilatory physiology of children and adolescents with anxiety disorders. Arch Gen Psychiatry 55, 123–129. [DOI] [PubMed] [Google Scholar]
- Pine DS, Klein, RG , Coplan JD, Papp LA, et al (2000). Differential carbon dioxide sensitivity in childhood anxiety disorders and non‐ill comparison group. Arch Gen Psychiatry 57, 960–967. [DOI] [PubMed] [Google Scholar]
- Ponikowski P, Chua TP, Anker SD, Francis DP, Doehner W, Banasiak W, Poole‐Wilson PA, Piepoli MF & Coats AJ (2001). Peripheral chemoreceptor hypersensitivity: an ominous sign in patients with chronic heart failure. Circulation 104, 544–549. [DOI] [PubMed] [Google Scholar]
- Prabhakar NR (2000). Oxygen sensing by the carotid body chemoreceptors. J Appl Physiol 88, 2287–2295. [DOI] [PubMed] [Google Scholar]
- Puthon L, Bouzat P, Rupp T, Robach P, Favre‐Juvin A & Verges S (2016). Physiological characteristics of elite high‐altitude climbers. Scand J Med Sci Sports. 26, 1052–1059. [DOI] [PubMed] [Google Scholar]
- Raurich JM, Vilar M, Colomar A, Ibanez J, Ayestaran I, Perez‐Barcena J & Llompart‐Pou JA (2010). Prognostic value of the pulmonary dead‐space fraction during the early and intermediate phases of acute respiratory distress syndrome. Respiratory Care 55, 282–287. [PubMed] [Google Scholar]
- Rebuck AS, Rigg JRA & Saunders NS (1976). Respiratory frequency response to progressive isocapnic hypoxia. J Physiol 258, 19–31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reiman EM, Fusselman MJ, Fox PT & Raichle ME (1989). Neuroanatomical correlates of anticipatory anxiety. Science 243, 1071–1074. [DOI] [PubMed] [Google Scholar]
- Richard NA & Koehle MS (2012). Differences in cardio‐ventilatory responses to hypobaric and normobaric hypoxia: a review. Aviat Space Environ Med 83, 677–84. [DOI] [PubMed] [Google Scholar]
- Richard NA, Sahota IS, Widmer N, Ferguson S, Sheel AW & Koehle MS (2014). Acute mountain sickness, chemosensitivity, and cardiorespiratory responses in humans exposed to hypobaric and normobaric hypoxia. J Appl Physiol 116, 945–952. [DOI] [PubMed] [Google Scholar]
- Richter DW (1982). Generation and maintenance of the respiratory rhythm. J Exp Biol 100, 93–107. [DOI] [PubMed] [Google Scholar]
- Rigg JR, Inman EM, Saunders NA, Leeder SR & Jones NL (1977). Interaction of mental factors with hypercapnic ventilatory drive in man. Clin Sci Mol Med 52, 269–275. [DOI] [PubMed] [Google Scholar]
- Robertshaw D (2006). Mechanisms for the control of respiratory evaporative heat loss in panting animals. J Appl Physiol 101, 664–668. [DOI] [PubMed] [Google Scholar]
- Rosenberg M, Tobias R, Bourke D & Kamat V (1980). Respiratory responses to surgical stimulation during enflurane anesthesia. Anesthesiology 52, 166–167. [PubMed] [Google Scholar]
- Sarton E, Dahan, A , Teppema L, Berkenbosch A, van den Elsen M & van Kleef J (1997). Influence of acute pain induced by activation of cutaneous nociceptors on ventilatory control. Anaesthesiology 87, 289–296. [DOI] [PubMed] [Google Scholar]
- Savourey G, Launay JC, Besnard Y, Guinet‐Lebreton A, Alonso A, et al. (2007). Normo or hypobaric hypoxic tests: propositions for the determination of the individual susceptibility to altitude illnesses. Eur J Appl Physiol 100, 193–205. [DOI] [PubMed] [Google Scholar]
- Savourey G, Launay JC, Besnard Y, Guinet A & Travers S (2003). Normo and hypobaric hypoxia: are there any physiological differences? Eur J Appl Physiol 89, 122–6. [DOI] [PubMed] [Google Scholar]
- Schoene RB (2001). Limits of human lung function at high altitude. J Exp Biol 204, 3121–3127. [DOI] [PubMed] [Google Scholar]
- Schoene RB, Lahiri S, Hackett PH, et al (1984). Relationship of hypoxic ventilatory response to exercise performance on Mount Everest. J Appl Physiol 56, 1478–1483. [DOI] [PubMed] [Google Scholar]
- Schondorf R & Low PA (1993). Idiopathic postural orthostatic tachycardia syndrome: An attenuated form of acute pandysautonomia? Neurology 43, 132–137. [DOI] [PubMed] [Google Scholar]
- Sheel AW & Romer LM (2012). Ventilation and respiratory mechanics. Compr Physiol 2, 1093–1142. [DOI] [PubMed] [Google Scholar]
- Simone DA & Kajander KC (1997). Responses of cutaneous A‐fiber nociceptors to noxious cold. J Neurophysiol 77, 2049–2060. [DOI] [PubMed] [Google Scholar]
- Simonova TG & Kozyreva TV (1994). Respiratory response to local skin cooling at artificial modulation of skin thermoreceptors. Proc Sixth Int Conf on Environmental Ergonomics 38–39. [Google Scholar]
- Schmidt C, Bellingham MC & Richter DW (1995). Adenosinergic modulation of respiratory neurones and hypoxic responses in the anaesthetized cat. J Physiol 483, 769–781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith JC, Abdala AP, Borgmann A, Rybak IA & Paton JF (2013). Brainstem respiratory networks: building blocks and microcircuits. Trends Neurosci 36, 152–162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith JC, Abdala AP, Koizumi H, Rybak IA & Paton JF (2007). Spatial and functional architecture of the mammalian brain stem respiratory network: a hierarchy of three oscillatory mechanisms. J Neurophysiol 98, 3370–3387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith JC, Abdala AP, Rybak IA & Paton JF (2009). Structural and functional architecture of respiratory networks in the mammalian brainstem. Philos Trans R Soc Lond B Biol Sci 364, 2577–2587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- St‐John WM (1998). Neurogenesis of patterns of automatic ventilatory activity. Prog Neurobiol 56, 97–117. [DOI] [PubMed] [Google Scholar]
- St‐John WM, Bledsoe TA & Sokol HW (1984). Identification of medullary loci critical for neurogenesis of gasping. J Appl Physiol 59, 1008–1019. [DOI] [PubMed] [Google Scholar]
- St‐John WM & Paton JF (2003). Defining eupnoea. Respir Physiol Neurobiol 139, 97–103. [DOI] [PubMed] [Google Scholar]
- St‐John WM & Paton JF (2004). Role of pontile mechanisms in the neurogenesis of eupnoea. Respir Physiol Neurobiol 143, 321–332. [DOI] [PubMed] [Google Scholar]
- St‐John WM, Stornetta RL, Guyenet PG & Paton JF (2009). Location and properties of respiratory neurones with putative intrinsic bursting properties in the rat in situ. J Physiol 587, 3175–3188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Subramanian HH & Holstege G (2014). The midbrain periaqueductal gray changes the eupneic respiratory rhythm into a breathing pattern necessary for survivial of the individual and of the species. Prog Brain Res 212, 351–384. [DOI] [PubMed] [Google Scholar]
- Suess WM, Alexander AB, Smith DD, Sweeney HW & Marion RJ (1980). The effects of psychological stress on respiration: a preliminary study of anxiety and hyperventilation. Psychophysiology 17, 535–540. [DOI] [PubMed] [Google Scholar]
- Takakura AC, Moreira TS, Colombari E, West GH, Stornetta RL, Guyenet PG (2006). Peripheral chemoreceptor inputs to retrotrapezoid nucleus (RTN) CO2‐sensitive neurons in rats. J Physiol 572, 503–523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tarbichi AG, Rowley JA, Shkoukani MA, Mahadevan K & Badr MS (2003). Lack of gender difference in ventilatory chemoresponsiveness and post‐hypoxic ventilatory decline. Respir Physiol Neurobiol 137, 41–50. [DOI] [PubMed] [Google Scholar]
- Taylor SE, Klein LC, Lewis BP, Gruenewald TL, Gurung RA & Updegraff JA (2000). Biobehavioral responses to stress in females: tend‐and‐befriend, not fight‐or‐flight. Psychol Rev 107, 411–429. [DOI] [PubMed] [Google Scholar]
- Teppema LJ & Dahan A (2010). The ventilatory response to hypoxia in mammals: mechanisms, measurement, and analysis. Physiol Rev 90, 675–754. [DOI] [PubMed] [Google Scholar]
- Thornton JM, Guz A, Murphy K, Griffin AR, Pedersen DL, Kardos A, Leff A, Adams L, Casadei B & Paterson DJ (2001). Identification of higher rain centres that may encode the cardiorespiratory response to exercise in humans. J Physiol 533, 823–836. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Timmers HJ, Wieling W, Karemaker MJ & Lenders JW (2003). Denervation of carotid baro‐ and chemoreceptors in humans. J Physiol 553, 3–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Timms RJ (1981). A study of the amygdaloid defence reaction showing the value of Althesin anaesthesia in studies of the functions of the fore‐brain in cats. Pflugers Arch 391, 49–56. [DOI] [PubMed] [Google Scholar]
- Tipton MJ (1989). The initial responses to cold‐water immersion in man. Clin Sci 77, 581–588. [DOI] [PubMed] [Google Scholar]
- Tipton MJ & Golden FS (1987). The influence of regional insulation on the initial responses to cold immersion. Aviat Space Environ Med 58, 1192–1196. [PubMed] [Google Scholar]
- Tipton MJ, Kadinopoulos P, Roiz de Sa D & Barwood MJ (2017). Changes in lung function during exercise are independently mediated by increases in deep body temperature. BMJ Open Sport & Exercise Medicine, https://doi.org/10.1136/bmjsem-2016-000210 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tipton MJ, Stubbs DA & Elliott DH (1991). Human initial responses to immersion in cold water at 3 temperatures and following hyperventilation. J Appl Physiol 70, 317–322. [DOI] [PubMed] [Google Scholar]
- Todd AJ, Spike RC, Young S & Puskar Z (2005). Fos induction in lamina I projection neurons in response to noxious thermal stimuli. Neuroscience 131, 209–217. [DOI] [PubMed] [Google Scholar]
- Tracey I & Dickenson A (2012). SnapShot: pain perception. Cell 148, 1308–1308. [DOI] [PubMed] [Google Scholar]
- Tracey I & Mantyh P (2007). The cerebral signature for pain perception and its modulation [Review]. Neuron 55, 377–391. [DOI] [PubMed] [Google Scholar]
- Trenchard D & Guz A (1972). Role of pulmonary vagal afferent nerve fibers in the development of rapid shallow breathing in lung inflammation. Clin Sci 42, 251–263. [DOI] [PubMed] [Google Scholar]
- Tsuji B, Honda Y, Fujii N, Kondo N & Nishiyasu T (2012a). Effect of initial core temperature on hyperthermic hyperventilation during prolonged submaximal exercise in the heat. Am J Physiol Regul Integr Comp Physiol 302, R94–R102. [DOI] [PubMed] [Google Scholar]
- Tsuji B, Honda Y, Fujii N, Kondo N & Nishiyasu T (2012b). Comparison of hyperthermic hyperventilation during passive heating and prolonged light and moderate exercise in the heat. J Appl Physiol 113, 1388–1397. [DOI] [PubMed] [Google Scholar]
- Tsuji B, Hayashi K, Kondo N & Nishiyashi T (2016). Characteristics of hyperthermia‐induced hyperventilation in humans. Temperature 3, 146–160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Diest I, Winters W, Devriese S, Vercamst E, Han JN, Van de Woestijne KP & Van den Bergh O (2001). Hyperventilation beyond fight/flight: respiratory responses during emotional imagery. Psychophysiology 38, 961–968. [DOI] [PubMed] [Google Scholar]
- Van Klaveren RJ & Demedts M (1998). Determinants of the hypercapnic and hypoxic response in normal man. Respir Physiol 113, 157–165. [DOI] [PubMed] [Google Scholar]
- Vejby‐Christensen H & Petersen ES (1973a). Effect of body temperature and hypoxia on the ventilatory CO2 response in man. Respir Physiol 19, 322–332. [DOI] [PubMed] [Google Scholar]
- Vejby‐Christensen H & Petersen ES (1973b). The effect of raised body temperature on the ventilatory CO2 response in man. Bull Physiopathol Respir 9, 672–675. [PubMed] [Google Scholar]
- von Euler C (1983). On the central pattern generator for the basic breathing rhythmicity. J Appl Physiol Respir Environ Exerc Physiol 55, 1647–1659 [DOI] [PubMed] [Google Scholar]
- von Euler C & Trippenbach T (1976). Excitability changes of the inspiratory “off‐switch” mechanism tested by electrical stimulation in nucleus parabrachialis in the cat. Acta Physiol Scand 97, 175–188. [DOI] [PubMed] [Google Scholar]
- von Leupoltd A, Sommer T, Kegat S, Baumann HJ, Klose H, Dahme B & Buchel C (2009). Dyspnea and pain share emotion‐related brain network. Neuroimage 48, 200–206. [DOI] [PubMed] [Google Scholar]
- Wagner JA & Horvath SM (1985). Influences of age and gender on human thermoregulatory responses to cold exposures. J Appl Physiol 58, 180–186. [DOI] [PubMed] [Google Scholar]
- Waldrop TG, Millhorn DE, Eldridge FL & Klingler LE (1984). Respiratory responses to noxious and nonnoxious heating of skin in cats. J Appl Physiol 57, 1738–1741. [DOI] [PubMed] [Google Scholar]
- Ward LM (2002). Synchronous neural oscillations and cognitive processes. Trends in cognitive sciences 7, 553–559. [DOI] [PubMed] [Google Scholar]
- Weese‐Mayer DE, Berry‐Kravis EM, Zhou L, Maher BS, Silvestri JM, Curran ME & Marazita ML (2003). Idiopathic congenital central hypoventilation syndrome: analysis of genes pertinent to early autonomic nervous system embryologic development and identification of mutations in PHOX2b. Am J Med Genet A 123A, 267–278. [DOI] [PubMed] [Google Scholar]
- Weissman C, Askanazi J, Forse RA, Hyman AI, Milic‐Emili J & Kinney JM (1986). The metabolic and ventilatory response to the infusion of stress hormones. Ann Surg 203, 408–412. [DOI] [PMC free article] [PubMed] [Google Scholar]
- West JB (2012). The physiological legacy of the Fenn, Rahn, and Otis school. Am J Physiol Lung Cell Mol Physiol 303, L845–L851, [DOI] [PubMed] [Google Scholar]
- Whipp BJ (1984). Carotid bodies and breathing in humans. Thorax 49, 1081–1084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- White MD (2006). Components and mechanisms of thermal hyperpnea. J Appl Physiol 101, 655–663. [DOI] [PubMed] [Google Scholar]
- Whitelaw WA & Derenne JP (1993). Airway occlusion pressure. J Appl Physiol 74, 1475–1483. [DOI] [PubMed] [Google Scholar]
- Whitelaw WA, Derenne J‐P & Milic‐Emili J (1975). Occlusion pressure as a measure of respiratory centre output in conscious man. Resp Physiol 23, 181–199. [DOI] [PubMed] [Google Scholar]
- Widdicombe JG (1964). Respiratory reflexes In Handbook of Physiology, section 2, vol. 1. eds Fenn WO. & Rahn H, pp. 585–630. American Physiological Society, Washington DC. [Google Scholar]
- Widdicombe JG & Winning A (1974). Effects of hypoxia, hypercapnia and changes in body temperature on the pattern of breathing in cats. Resp Physiol 21, 203–221. [DOI] [PubMed] [Google Scholar]
- Wilhelm FH, Gevirtz R & Roth WT (2001). Respiratory dysregulation in anxiety, functional cardiac, and pain disorders. Assessment, phenomenology, and treatment. Behaviour Modification 25, 513–545. [DOI] [PubMed] [Google Scholar]
- Williamson JW, Fadel PJ & Mitchell JH (2006). New insights into central cardiovascular control during exercise in humans: a central command update. Exp Physiol 91, 51–58. [DOI] [PubMed] [Google Scholar]
- Willer JC (1975). The influence of anticipation of pain on heart and respiratory rates and on the pain reflex in man [in French: Influence de l'anticipation de la douleur sur les frequencies cardiaque et respiratoire et sur le reflexe nociceptive chez l'homme]. Physiology and Behaviour 15, 411–415. [DOI] [PubMed] [Google Scholar]
- Winters PM, McCabe PM, Green EJ & Schneiderman N (2000). Stress responses, coping, and cardiovascular neurobiology: central nervous system circuitry underlying learned and unlearned affective responses to stressful stimuli In Stress, Coping and Cardiovascular Disease. eds Mccabe P, Schneiderman N, Field TM. & Wellens AR.pp. 1–49. Routledge, UK. [Google Scholar]
- Younes MK & Remmers JE (1981). Control of tidal volume and respiratory frequency In Regulation of Breathing, part 1, vol.17, ed. Hornbein TF, pp. 621–671. Marcel Dekker, New York. [Google Scholar]
- Zoccal DB, Simms AE, Bonagamba LG, Braga VA, Pickering AE, Paton JF & Machado BH (2008). Increased sympathetic outflow in juvenile rats submitted to chronic intermittent hypoxia correlates with enhanced expiratory activity. J Physiol 586, 3253–3265. [DOI] [PMC free article] [PubMed] [Google Scholar]