


Abstract
Properties of poliovirus RNA-dependent RNA polymerase (3Dpol) including optimal conditions for primer extension, processivity and the rate of dissociation from primer-template (koff) were examined in the presence and absence of viral protein 3AB. Primer-dependent polymerization was examined on templates of 407 or 1499 nt primed such that fully extended products would be 296 or 1388 nt, respectively. Maximal primer extension was achieved with low rNTP concentrations (50–100 µM) using pH 7 and low (<1 mM) MgCl2 and KCl (<20 mM) concentrations. However, high activity (about half maximal) was also observed with 500 µM rNTPs providing that higher MgCl2 levels (3–5 mM) were used. The enhancement observed with the former conditions appeared to result from a large increase in the initial level or active enzyme that associated with the primer. 3AB increased the number of extended primers at all conditions with no apparent change in processivity. The koff values for the polymerase bound to primer-template were 0.011 ± 0.005 and 0.037 ± 0.006 min–1 (average of four or more experiments ± SD) in the presence or absence of 3AB, respectively. The decrease in the presence of 3AB suggested an enhancement of polymerase binding or stability. However, binding was tight even without 3AB, consistent with the highly processive (at least several hundred nucleotides) nature of 3Dpol. The results support a mechanism whereby 3AB enhances the ability of 3Dpol to form a productive complex with the primer-template. Once formed, this complex is very stable resulting in highly processive synthesis.
INTRODUCTION
Picornaviruses are responsible for several diseases including the common cold and chronic hepatitis in humans and foot and mouth disease in animals. These viruses contain plus sense single-stranded RNA that is directly translated into a long polyprotein upon entry into the cytoplasm. Approximately 10 different proteins are produced by cleavage of the polyprotein at specific locations (1,2). Among these are the RNA-dependent RNA polymerase (RdRP), which is responsible for converting the plus sense genome to a minus sense complement then using the complement to produce new plus sense genomes. Because of the unique nature of its activity, RdRP represents a potentially important therapeutic target. The RdRP enzymes from several picornaviruses have been purified either from infected cells or more recently by over-expression in bacteria (3–8). The most heavily studied of these polymerases is the poliovirus RdRP, also referred to as 3Dpol. In addition, three other viral proteins have been shown to participate in poliovirus genome replication. The viral proteins are termed 3CD, 3AB and 3B with the latter being a cleavage product of 3AB and 3CD the precursor of 3Dpol (9–13). Presumably these viral factors and possibly a host protein form a complex by interacting with a cloverleaf structure that forms near the 5′-end of the viral RNA (9,13–15). The exact sequence of events leading to release of 3B and 3Dpol from the precursor proteins is not clear although 3C, or the 3C portion of 3CD, is likely to carry out the cleavage. Protein 3B (more commonly called VPg) is covalently linked to the 5′-end of newly synthesized viral RNAs and is believed to serve as a primer for RNA synthesis by 3Dpol (16–18). It has been shown in vitro that 3Dpol catalyzes uridylation of a specific tyrosine residue on 3B and can subsequently use this complex to prime poly(rU) synthesis (19). The role of 3AB is not as clearly defined although results suggest that it stimulates RNA synthesis by 3Dpol perhaps by enhancing primer utilization (11,12,20,21). In addition, the 3A domain of 3AB may act to anchor the replication complex to membranes (22). Although a basic understanding of the activities of the various viral proteins in RNA replication has been garnered, in no case is there a detailed mechanistic understanding of the individual proteins or the interactions between them.
Work on 3Dpol has been the most extensive and has resulted in an understanding of some biochemical properties including among others, cation and pH preference, Kd and Km measurements, substrate preference determinations and measurements of fidelity (23–26). Recently the crystal structure of the polymerase has also been solved (27). Interestingly, with homopolymeric templates, 3Dpol shows greater activity with Mn2+ as opposed to Mg2+ (the likely divalent cation in vivo) as divalent cation. The enzyme can also catalyze both primer-dependent and de novo (template-dependent synthesis in the absence of a primer) synthesis and both activities are optimal with Mn2+. Despite the improved activity with Mn2+, fidelity is markedly reduced and the enzyme is capable of incorporating dNTPs and ddNTPs and using DNA as a template (25). In addition to polymerase activity, 3Dpol also has strand displacement and terminal transferase activities. The former could conceivably function to release single-stranded genomes or antigenomes from their templates while the role of the latter activity is not clear (28). It has also been reported that 3Dpol binds cooperatively to nucleic acids and can multimerize. It has been suggested that multimerization may be required for efficient nucleic acid binding and polymerization (29).
Results indicate that 3AB enhances RNA synthesis by 3Dpol (see above). Among the activities associated with 3AB are non-specific RNA binding (20), direct binding to 3CD and 3Dpol (21,30), stimulation of cleavage of 3CD to 3C plus 3Dpol (30), and association with membranes as an integral membrane protein (22). The exact mechanism by which 3AB stimulates 3Dpol is not clear although a number of insights have been made. Since 3AB binds both RNA and 3Dpol (and 3CD) it could serve as a protein cofactor that promotes association of 3Dpol to the template (12). Such cofactors have been found in other systems, for example the proliferating cellular nuclear antigen (PCNA) and herpes UL42 protein, which enhance the binding of eukaryotic polymerase δ and herpes DNA polymerase, respectively, to their templates (31–34). Consistent with this is the finding that 3Dpol alone is a relatively poor nucleic acid binding protein and may require a cofactor to carry out synthesis (29,35). 3AB may also serve to localize the polymerase to the appropriate position to initiate RNA synthesis. During viral replication this would presumably be near the termini of the genome or antigenome, perhaps associated with specific cloverleaf structures near the termini that have been shown to be important in replication (9,36). Although 3AB may be part of such a replication complex, the non-specific nature of its RNA binding activity suggests that other factors are required to impart specificity on the replication complex (20). During primer-dependent synthesis in vitro or to reinitiate aborted synthesis in vivo (for example to carry out recombination), the site of initiation would be a recessed 3′ RNA terminus (RNA primer). 3AB has been shown to enhance primer usage in vitro. One report indicated that 3AB enhances de novo RNA synthesis and synthesis on weakly bound primer–templates but does not stimulate synthesis when the primer and template form a stable complex (12). This implies that 3AB may stabilize the primer–template complex in addition to interacting with 3Dpol.
To better understand the mechanism of viral replication and the interactions between the various viral and host proteins, a reliable in vitro system will be required. In this report we have sought to maximize 3Dpol and 3AB activities in vitro by examining optimal conditions for primer extension. We also studied the processivity (average number of nucleotides added to the primer in a single binding event between the primer-template and polymerase) and dissociation rate (koff) of 3Dpol in the presence and absence of 3AB using virus-derived RNA templates. To our surprise very low divalent cation concentrations (<1 mM Mg2+) greatly stimulated primer extension while having no apparent effect on processivity. Although 3AB stimulated primer extension under all tested cation and salt concentrations, synthesis was maximal in the presence of 3AB with low cation and salt levels. Once again no change in processivity was observed. 3AB stimulation was apparently due at least in part to a stabilization of 3Dpol binding to the primer as the off-rate (koff) in the presence of 3AB was significantly lowered. However, even in the absence of 3AB a relatively low off-rate was measured. Consistent with the low off-rate, the processivity of 3Dpol was high averaging at least several hundred nucleotides. The results are consistent with the inability of 3Dpol to form a high proportion of productive complexes with the primer. Once formed, such complexes are highly stable and even more so in the presence of 3AB. After primer extension is initiated the polymerase generally continues extension for at least several hundred nucleotide additions. This suggests that the role of 3AB and perhaps other factors are to enhance initiation and perhaps reutilization or 3Dpol after completion of synthesis. The highly processive nature of 3Dpol implies only a single initiation event may be required to complete genome synthesis.
MATERIALS AND METHODS
Materials
Restriction enzymes, calf intestinal alkaline phosphatase (CIP), T3 RNA polymerase, DNase I (RNase free), ribonucleotides and deoxyribonucleotides were obtained from Roche. RNasin was obtained from Promega. Thrombin was obtained from Sigma. DNA oligonucleotides used as primers were purchased from Genosys. RNA oligonucleotides used as primer were purchased from Dharmacon Research and Cyber Syn. Radiolabeled compounds were from New England Nuclear. Nonidet P-40 was obtained from Calbiochem. Protein concentrators, Centricon 30 and 10, were obtained from Amicon. HEPES (N-2-hydroxyethylpiperazine-N′-2-ethanesulfonic acid) and glycerol, enzyme grade were obtained from Fisher Scientific. Antibiotics, molecular biology grade were obtained from Kodak International Biotechnology. Dithiothreitol (DTT) was obtained from Shelton Scientific. All other chemicals were obtained from Sigma.
Preparation of polymerase (3Dpol) and protein 3AB by expression in Escherichia coli
The polymerase of poliovirus type 1 (Mahoney strain) used for these studies was expressed in E.coli using plasmid pT7pol (4,23). This plasmid codes for kanamycin resistance and contains the gene for 3Dpol under control of a T7 RNA polymerase promoter. The plasmid was used to transform E.coli strain BL21 DE3(LysS) which contains a chloramphenicol resistant gene and a chromosomal copy of the gene coding for T7 RNA polymerase under control of an IPTG-inducible promoter. The viral polymerase, 3Dpol was expressed after addition of IPTG to the growth medium to induce production of T7 polymerase. Polymerase 3Dpol was purified as described previously (4,23). The recovered enzyme was homogenous as determined by Coomassie blue staining and had a specific activity of ∼1.5 × 106 U/mg [1 U is defined as 1 pmol of UMP incorporated into trichloroacetic acid-precipitable poly(U) in 30 min at 30°C using oligo(U)-primed poly(rA) as template]. The 3AB viral protein of poliovirus type 1 (Mahoney strain) was expressed in E.coli using plasmid pGEX-3AB. Expression and purification were performed as described previously (21). Purified 3AB was stored at –70°C in buffer containing 50 mM Tris–HCl pH 8, 1 mM DTT, 0.05% Triton X-100 and 10% glycerol (3AB buffer). The final concentration of 3AB was ∼840 nM.
Preparation of plasmids containing poliovirus sequences
The constructs developed to test the biochemical properties of 3Dpol were prepared by inserting fragments of the poliovirus sequence derived from pT7PV1-5 (a gift from E.Wimmer, SUNY Stony Brook), which contains a cDNA copy of the poliovirus genome, inserted in pBSM13+ (Stratagene). Plasmid pBSM13-228 was constructed by inserting a 228 bp HindII fragment (nucleotides 6776–7003 of poliovirus type 1, Mahoney strain) from pT7PV1-5 into the HindII restriction site at position 914 of pBSM13+. Plasmid pBSM13-1320 was constructed by inserting a 1320 bp AflIII fragment (nucleotides 4577–5896 of poliovirus type 1, Mahoney strain) from pT7PVM1-5 into the HindII restriction site at position 914 of pBSM13+.
Preparation of RNA templates
Plasmids pBSM13-228 and pBSM13-1320 were cleaved with PvuII and T3 RNA was used to generate run-off transcripts ∼407 and 1499 nt in length, respectively, according to the enzyme manufacturer’s protocol. After transcription, 1 U of DNase I (RNase free) was added to digest the DNA plasmid and the incubation was continued for 15 min. The transcription reactions were extracted with phenol:chloroform:isoamyl alcohol (25:24:1) and precipitated with ethanol. The RNA was further purified on a 6% denaturing polyacrylamide gel, located by ultraviolet shadowing, and recovered from the excised gel slice (37). The RNA was quantified by spectrophotometric analysis. To calculate the molecular weight of the RNA the following equation was used: [(A × 328.2.2) + (G × 344.2) + (C × 304.2) + (U × 305)]. Then the molecular weight was used to determine the molar concentration of the RNA transcripts using the standard conversion of 1 OD260 ≈ 40 µg/ml for single-stranded RNA.
RNA–RNA hybridization
For the reactions performed, a 5′-32P-labeled 20 nt RNA primer (5′-GGGCGAAUUCGAGCTCGGUA-3′) was hybridized to the RNA template by mixing primer–RNA transcript at a 3:1 ratio in buffer containing 50 mM HEPES pH 7, 10 mM KCl and 5 mM DTT. The mixture was heated at 65°C for 5 min then slowly cooled to room temperature.
Standard RNA synthesis reaction with 3Dpol
The 407 or 1499 nt RNA transcripts (described above) were hybridized to a 5′-32P-labeled 20 nt RNA primer (as described above). Full extension of the primer produced 296 or 1388 nt RNA products for the 407 and 1499 nt templates, respectively. Primer–template complexes (5 nM in template) were pre-incubated with 3Dpol (13 nM) in 10.5 µl in a buffer containing 50 mM HEPES pH 7, 10 mM KCl, 0.8 mM MgCl2, 5 mM DTT and 0.4 U/µl RNasin for 5 min at 30°C. In some reactions viral protein 3AB was added at the indicated concentration during the pre-incubation. Reactions were initiated by addition of 50 µM (final concentration in reaction) rNTPs in 2 µl of the above buffer and incubations were continued for 1 h at 30°C. In some assays specific components of the reaction mixture were varied (as indicated) to test the effect of reaction conditions on RNA synthesis. Reactions were stopped with 12.5 µl of 2× loading buffer [90% (v/v) formamide, 10 mM EDTA pH 8, bromophenol blue and xylene cyanol, 0.1% (w/v) each] and the products were analyzed on 5 or 6% denaturing polyacrylamide gels, for the 1499 and 407 nt RNA templates, respectively. Dried gels were used for autoradiography or quantified using a GS-525 phosphoimager (Bio-Rad).
Polyacrylamide gel electrophoresis
Denaturing polyacrylamide gels [5 or 6% (w/v) polyacrylamide (19:1 acrylamide:bisacryamide), 8 M urea] were prepared and electrophoresed as described (37).
Conditions for reactions to test processivity
Reactions to determine the processivity of 3Dpol in the presence or absence of 3AB were performed as described above for standard RNA synthesis reactions with the following changes. In order to achieve a better visual analysis of RNA synthesis the 3Dpol concentration was increased to 26 nM and the 407 or 1499 nt RNA template was used with 5′-32P-lableled RNA primer at a final concentration of 5 nM (unless otherwise indicated). Reactions were performed in the presence or absence of 3AB (134 nM final concentration). The reactions were initiated by a supplement containing rNTPs and heparin (1 µg/µl final concentration). Heparin was included to sequester 3Dpol molecules after dissociation from primer-template. This limited the synthesis to a single round of binding between the enzyme and primer-template. To determine the effects on processivity of changes in the reaction conditions, variation of salt and magnesium concentrations were also performed as indicated.
Reactions to determine the dissociation rate constant (koff) of 3Dpol
Reactions were performed using the 407 nt RNA template used in standard 3Dpol reactions described above. Primer-template was pre-incubated at 30°C with 3Dpol for 5 min in 40 µl of the buffer described for the standard reactions. Reactions were performed in the presence or absence of 3AB in the pre-incubation. After pre-incubation, 10 µl of standard buffer containing heparin was added. Aliquots (5 µl) were removed 5, 10, 15, 20, 25, 30 and 35 min after heparin addition. The aliquots were added to a tube containing rNTPs in the same buffer and incubated for 10 min. The final concentrations of primer-template, 3Dpol, 3AB, heparin and rNTPs were 10 nM (in template), 26 nM, 134 nM, 1 µg/µl and 50 µM, respectively. Samples were terminated and processed as described for the standard reactions above. The amount of extended primer was determined using a phosphoimager. The dissociation rates were calculated by constructing a graph of total primer extension versus time. The data was fit to an equation for single exponential decay: f(x) = ae–bx, where a =1 and b is the dissociation rate. The graph was prepared using Sigma Plot (Jandel Scientific) to construct the graphs and determine the koff.
RESULTS
Optimal conditions for 3Dpol synthesis
To determine optimal conditions for primer extension a 407 nt RNA template was primed with a 32P-labeled 20 nt RNA primer such that synthesis to the end of the template resulted in a 296 nt product. The plasmid used to make the RNA template contained a 228 nt insert derived from a poliovirus cDNA clone (see Materials and Methods). Optimal conditions for extension were determined using standard reaction conditions (see Materials and Methods) but varying one of the components of the reactions. An optimal pH of 7 was determined (Fig. 1). The level of primer extension relative to pH 7 was ∼50% at pH 8 while <10% extension was observed at pH 6. No extension occurred at pH 4 or 5. Values between 7 and 8 were also tested without improvement relative to pH 7 (data not shown). An optimal temperature of 30°C was observed for extension during a 1 h reaction (Fig. 1). Decreasing the temperature to 25°C had little effect on extension while reactions were severely inhibited at 37 and 45°C although some extension was observed. Note that some lanes in the gel are over-exposed so that small levels of synthesis occurring at higher temperatures could be observed. Extending the reactions beyond 1 h did not increase the level of primer extension (data not shown).
Figure 1.

Determination of optimal conditions for primer extension by 3Dpol. Shown are autoradiograms from experiments testing primer extension while changing various reaction parameters (as indicated). The 407 nt template and the 20 nt 5′-32P-labeled primer were used for all experiments. Standard RNA synthesis reaction conditions (see Materials and Methods) were employed except that one parameter (as indicated) was varied in each set of reactions. Fully extended (FL) products are 296 nt in length. Lanes labeled C show reactions in the absence of enzyme. For the MgCl2 titration the concentrations used were (left to right after lane C) 0.05, 0.1, 0.2, 0.4, 0.8, 1.6, 2.0, 2.5, 3.0, 3.5, 4.0, 4.5 and 5.0 mM. For the KCl titration the concentrations used were (left to right after lane C) 1, 4, 10, 20, 40 and 80 mM.
Optimal divalent cation, salt and nucleotide concentrations were also investigated. Surprisingly the optimal MgCl2 concentration was very low. Values between 0.1 and 0.8 mM gave approximately equal levels of primer extension (Fig. 1). Note that these values are significantly lower than the 3–5 mM levels of divalent cation generally used in 3Dpol reactions (4,11,12,20,25). The level of primer extension was substantially decreased at ≥1.6 mM although products were observed. The results were essentially identical using magnesium acetate (data not shown). Salt (KCl) concentrations of <10 mM supported optimal primer extension while inhibition was observed at ≥20 mM (Fig. 1). A similar result was noted using NaCl in place of KCl (data not shown). Consistent with recent reports from others (25), MnCl2 also supported primer extension. The level of primer extension at various MnCl2 concentrations was typically only slightly greater than at the same level with MgCl2 (data not shown). Primer extension on several other primer-templates including heteropolymers with different sequences and homopolymers was also tested. In all cases, low divalent cation concentrations lead to the greatest level of primer extension and reactions were inhibited by high salt (data not shown). It should be noted that even under the best conditions the proportion of primers extended by 3Dpol was low. Given the concentration of 3Dpol employed it was clear that the majority of enzymes do not carry out primer extension in the assays. In general, less than one in four primed templates (see below) were used for synthesis despite the fact that the enzyme was in stoichiometric excess. Others have also observed this phenomenon (24,35,38,39) (see below).
Further analysis was carried out to examine the discrepancy between the optimal levels reported by others and those found here. Reactions with 0.8 or 4 mM MgCl2 were examined at several rNTP concentrations (Fig. 2). Increasing the rNTP concentration beyond 100 µM inhibited reactions with 0.8 mM MgCl2. In contrast, reactions with 4 mM MgCl2 were stimulated by higher rNTP levels with maximum levels of primer extension observed at 500 µM rNTPs, a point where essentially no activity was observed with low MgCl2. The highest level of primer extension observed with 4 mM MgCl2 was ∼40–50% of the level observed with 0.8 mM MgCl2. Other experiments showed that between 3 and 5 mM MgCl2 was optimal at 500 µM rNTP concentrations (data not shown). This is consistent with results from other studies (25). Overall the results show that high activity can be obtained using conditions reported by others; however, maximal activity requires low divalent cation and rNTP conditions.
Figure 2.

Optimal nucleotide concentrations for reactions with 0.8 or 4 mM MgCl2. The 407 nt template and 20 nt 5′-32P-labeled RNA primer were used for reactions with 0.8 or 4 mM MgCl2 and various amounts of rNTPs. A plot of the relative level of primer extension versus rNTP concentration, [rNTPs], is shown. The highest level of primer extension (0.8 mM MgCl2, 100 µM rNTPs) was set to a value of 1.0 and the levels of primer extension in other reactions are expressed relative to this lane. A repeat of this experiment yielded similar results (data not shown).
Enhanced activity with low MgCl2 and rNTP concentrations results primarily from an increase in the initiation of primer extension, apparently resulting from an enhanced binding of active enzyme to the primer
As was noted above, the proportion of primers extended by 3Dpol even under optimal conditions was low. Especially considering that standard reactions contained excess polymerase relative to the level of primer-template. Most primer-dependent polymerases would extend essentially all template-bound primers under such conditions. In contrast, 3Dpol extends relatively low levels as was noted in this work and by several others. One likely reason is the high Kd value of 3Dpol for primer-template. Measurements of the Kd value for 3Dpol on small heteropolymeric and homopolymeric primer-templates indicated values in the 1 µM range, while polymerases in general are in the nanomolar range (24,35). Other reasons include the slow formation of active polymerase-primer–template complexes and the association of enzyme with the template at positions other than the 3′ primer terminus (24). To investigate this further, reactions were performed with a constant level of enzyme (26 nM) and various levels of primer-template ranging from 2.5 to 80 nM (Fig. 3). These reactions were performed in the presence and absence of a ‘trap’ that can limit extension to a single round of binding between the polymerase and primer-template. This was accomplished by pre-binding enzyme to the primer-template then initiating reactions in the presence or absence of heparin (see Materials and Methods). Heparin effectively sequestered 3Dpol that was not bound to or had dissociated from the primer-template. Several concentrations of trap were tested to determine an amount that could trap all the polymerase molecules yet was inert with respect to influencing polymerase molecules bound to or traversing along the template. Heparin at 1 µg/µl final concentration satisfied these criteria. No synthesis products were observed in reactions in which 3Dpol was premixed with this amount of heparin and incubated for 1 h (Fig. 3, lane T). Also, even much higher concentrations of heparin did not change the distribution of synthesis products in reactions with 3Dpol (data not shown). These results indicate that heparin at 1 µg/µl is an effective and inert trap. Increasing the level of primer-template substantially elevated the level of primer extension in reactions with or without trap. Phosphoimager analysis indicated that ∼20% of the primed templates were used for extension in reactions with 20 nM primer-template and no trap. In the presence of trap about half as many primers were extended in the same reactions. Higher concentrations resulted in an increase in overall extension and a decrease in percentage extension as expected (Fig. 3). Overall, the substantial increase in primer extension in trap reactions indicated that the enhancement resulted at least in part from an initial increase in the amount of active enzyme that associated with the primer on primer-templates. Clearly, increasing the level of primer-template may also increase the rate of primer extension by enhancing the rate at which enzyme molecules recycle after a round of synthesis in non-trap reactions. However, the dramatic increase in the presence of trap strongly suggests that a major factor in the low proportion of primers extended by 3Dpol is the low level of active enzyme that associates with the primer.
Figure 3.

Primer extension reactions in the presence or absence of trap with various template concentrations. Shown is an autoradiogram of a primer extension reaction performed in the presence or absence of heparin (trap). The 407 nt template and the 20 nt 5′-32P-labeled primer were used for all experiments. Conditions were as described in Materials and Methods (‘Conditions for reactions to test processivity’) except that the concentration of template was varied as indicated. Lane C, no enzymes added; lane T, trap control (enzyme mixed with heparin trap and rNTPs before mixing with primer-template). The concentrations of primers extended were 0.2, 0.6, 1.7, 5.8, 8.9 and 14.1 nM for reactions without trap and 2.5, 5, 10, 20, 40 and 80 nM template, respectively. The concentrations of primers extended were 0.2, 0.4, 1.1, 2.6, 3.3 and 3.7 nM for reactions with trap and 2.5, 5, 10, 20, 40 and 80 nM template, respectively. All other markings were as described in Figure 1.
Further analysis using various conditions in reactions with or without trap lead to the conclusion that, relative to non-optimal conditions, optimal conditions enhanced binding of active enzyme to the primer. As shown in Figure 4, 3Dpol was preincubated with primer-template under various conditions in the presence of either MgCl2 or rNTPs, then reactions were initiated by adding a supplement containing the component missing in the preincubation (either MgCl2 or rNTPs) in the presence or absence of heparin and continued for 1 h. The level of primer extension in reactions without heparin accesses overall extension under the final reaction conditions while those with heparin reflect the amount of 3Dpol that associated with the primer-template under the preincubation conditions (see Fig. 4 for relative levels of extension). The general conclusions from this experiment were that the inclusion of MgCl2 in the preincubation-enhanced association of 3Dpol to primer-template in comparison to including rNTPs [Fig. 4, compare lane 4 with lane 6 (0.8 mM MgCl2 or 50 µM rNTPs, respectively) and lane 8 with lane 10 (4 mM MgCl2 or 500 µM rNTPs, respectively)]. In addition, the lower MgCl2 and rNTP concentrations improved association relative to the higher ones (Fig. 4, compare lane 4 with lane 8 and lane 6 with lane 10). In the case of MgCl2, the enhancement in reactions with heparin was ∼10-fold indicating that 10 times as much active enzyme was initially bound to the primer with the lower divalent cation concentration (Fig. 4, compare lane 4 with lane 8). Also noteworthy was the level of enzyme recycling under the various conditions. Assuming that reactions with trap represent the level of enzyme initially bound to the primer-template, increased extension in the absence of trap would represent recycling of the enzyme and/or initiation by other enzyme molecules during the course of the reaction. The level of primer extension doubled in the absence of trap under optimal conditions (Fig. 4, compare lane 3 with lane 4) while increasing >10-fold under high Mg2+ and rNTPs conditions (compare lane 7 with lane 8). This suggests higher levels of divalent cation and dNTPs, although decreasing the initial binding of active 3Dpol to the primer-template may enhance recycling or stabilize the enzyme when it is not bound to substrate. In agreement with the latter explanation, others have found that 3Dpol is more stable in the presence of high concentrations of nucleotide (24). The results are consistent with the improvement in overall primer extension under optimal conditions resulting from enhanced binding of 3Dpol to the primer-template rather than enhanced turnover or stabilization of the enzyme.
Figure 4.
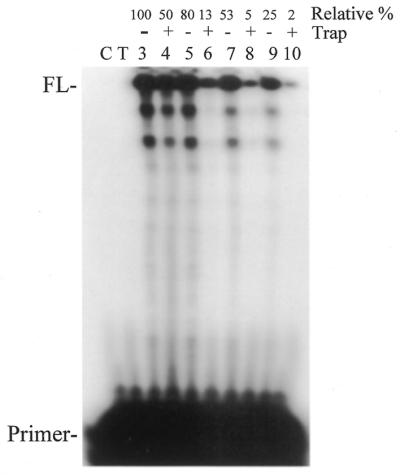
Evaluation of reaction conditions on the initial association and turnover of 3Dpol with primer-template in reactions with or without trap. Shown is an autoradiogram of a primer extension reaction performed in the presence or absence of heparin (trap) as indicated. The 407 nt template and the 20 nt 5′-32P-labeled primer were used for all experiments at a concentration of 10 nM template. Conditions were as described in Materials and Methods (‘Conditions for reactions to test processivity’). Enzyme was preincubated with primer-template in 10 µl in a buffer containing 50 mM HEPES pH 7, 10 mM KCl, 5 mM DTT, 0.8 U/µl RNasin and the following: lanes 3 and 4, 0.8 mM MgCl2; lanes 5 and 6, 50 µM rNTPs; lanes 7 and 8, 4.0 mM MgCl2; lanes 9 and 10, 500 µM rNTPs. Preincubations were for 5 min at 30°C. The reactions were initiated by adding 2.5 µl of supplement in the above buffer including the following: lanes 3 and 4, 0.8 mM MgCl2 and 250 µM rNTPs (final concentration 50 µM in reaction); lanes 5 and 6, 50 µM rNTPs and 4.0 mM MgCl2 (final concentration 0.8 mM in reactions); lanes 7 and 8, 4.0 mM MgCl2 and 2.5 mM rNTPs (final concentration 500 µM in reactions); lanes 9 and 10, 500 µM rNTPs and 20 mM MgCl2 (final concentration 4.0 mM in reactions). The supplement also included heparin (trap) as indicated above the lanes. Reactions were continued for 1 h after supplement addition. The percentage of primer extended in the reactions relative to lane 3 (100%) is shown above each lane. All other markings were as described in Figure 3.
One possible explanation for the findings that was ruled out was that binding to the primer decreased with the higher divalent cation concentration because a greater proportion of the enzyme distributes to the single-stranded region of the template. Filter binding assay showed that 3Dpol bound better to single-stranded RNA under the low Mg2+ conditions. In addition, competition assays indicated that ∼3-fold excess of single-stranded template was required to compete 50% of the enzymes off the primer under both low and high divalent cation conditions (data not shown). Essentially this means that most of the ‘active’ enzyme associates with the primer portion of the template under both conditions. These results show that enhanced primer extension with low Mg2+ does not result from a simple redistribution of the enzyme.
Viral protein 3AB decreases the off-rate (koff) of 3Dpol from primer-template
The effect of viral protein 3AB on primer extension was also studied. Consistent with results from others, 3AB increased the level of primer extension using both optimal and sub-optimal condition and various KCl concentrations (data not shown). The level of increase varied (generally between 2- and 10-fold) depending on the conditions used and was dependent on the concentration of 3AB added to the reactions up to the 136 nM concentration used in the experiments shown in Figures 5 and 6 (2 µl of the 3AB preparation). Further increases lead to inhibition although this may have resulted from Triton X-100 in the 3AB preparation. This reagent, which is required for the stability of 3AB, is known to inhibit the interaction between 3AB and 3Dpol (21). We also observed some inhibition of 3Dpol in the presence of 2 µl 3AB buffer indicating that Triton X-100 also inhibits 3Dpol synthesis.
Figure 5.

Determination of the koff value for 3Dpol on primer-template in the presence or absence of 3AB. Off-rates (koff) were determined as described in Materials and Methods. (A) An autoradiogram from a typical experiment performed in the presence or absence of 3AB (as indicated). The number above each lane indicates the time (min) between the addition of the heparin trap and the rNTPs. Lanes C, no enzymes added; lanes T, trap control (as described in Fig. 4); lanes E, control for primer extension in the absence of heparin. Reactions for lanes C, T and E were incubated for 35 min. Other markings are as described in Figure 2. (B) Plot of a koff experiment: volume (phosphoimager units) versus time from a typical koff experiment is shown. The values for volume were derived from phosphoimager analysis of the above autoradiogram. The curve was fit to an equation for single exponential decay in order to calculate koff (see Materials and Methods). Values for koff were 0.011 ± 0.005 min–1 and 0.037 ± 0.006 min–1 (averages of at least four experiments ± SD), in the presence or absence of 3AB, respectively.
Figure 6.
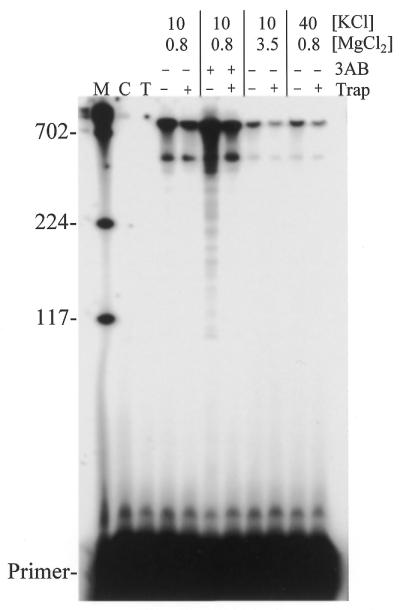
Polymerase 3Dpol is highly processive. Reactions to evaluate the processivity of 3Dpol were performed using the 1499 nt template and a 20 nt 5′-32P-labeled RNA primer as described in Materials and Methods (‘Conditions for reactions to test processivity’). Full extension of the primer would result in a 1388 nt product although the gel system used was unable to resolve products greater than ∼800 nt. An autoradiogram from a primer extension experiment with assays performed using various MgCl2 and KCl concentrations (as indicated) is shown. Some reactions were initiated in the presence of heparin (as indicated) and 3AB was included in some reactions (as indicated). The level of stimulation by 3AB was determined to be ∼3-fold by phosphoimager analysis. Lane M, DNA markers of the indicated sizes in nucleotides; lane C, no enzyme added; lane T, trap control (as described in Fig. 3).
Others have suggested that 3AB may stabilize the interaction between polymerase and primer-template although this has not been directly tested (12,21,40,41). To test this we performed experiments to determine the dissociation rate constant (koff) for 3Dpol bound to primer-template in the presence and absence of 3AB. Reactions were performed by first incubating 3Dpol in the presence or absence of 3AB with the primer-template but without rNTPs. After preincubation the trap was added and at various times after trap addition aliquots were removed and incubated with rNTPs. The amount of extension observed is proportional to the number of enzyme molecules productively bound to the primer-template. This amount decreases over time due to dissociation and, in the presence of trap molecules, the enzyme cannot rebind after dissociating. A typical experiment is shown in Figure 5. The koff value can be obtained by plotting the level of primer extension versus time (see Materials and Methods). Consistent with 3AB stabilizing the interaction of 3Dpol with the primer-template, an ∼3-fold decrease (from 0.037 ± 0.006 to 0.011 ± 0.005 min–1, average of four or more experiments ± SD) was observed in the presence of 3AB. It was also clear that 3AB enhanced the initial association of 3Dpol with primer-template given that the level of primer extension was increased at time zero in a reaction containing 3AB relative to those without. Notable, however, is the tight binding of the polymerase to primer-template even in the absence of 3AB. The 0.037 min–1 corresponds to a complex half-life of ∼18.7 min. Such a stable interaction is generally indicative of a highly processive polymerase (see below).
3Dpol is a highly processive polymerase
The processivity of 3Dpol was also examined in the presence and absence of 3AB. Because 3Dpol was highly processive a longer template was used to study processivity (see Materials and Methods). Full extension on this primer-template would result in a 1388 nt product. Primer extension on this template basically mimicked extension on the shorter template with respect to the conditions that enhanced extension (data not shown). In these reactions 3Dpol was pre-incubated with primer-template and reactions were initiated with rNTPs or with rNTPs and heparin trap. The reactions containing trap can be used to evaluate processivity since extension by 3Dpol is limited to a single binding event with the primer-template. The major product in the presence (and absence) of trap ran at the top of the gel, consistent with products greater than ∼800 nt in length (the limit of the gels resolving power) (Fig. 6). There were low levels of smaller products also present (especially evident in reactions without trap). These could result from the polymerase falling off the template before reaching the end or they could be due to low levels of truncated RNAs in the gel purified RNA preparation. Note that incomplete products were evident in many experiments with the smaller template but the gels were generally over-exposed to allow visualization of products produced at sub-optimal conditions. Phosphoimager analysis of products produced using the small template revealed that the fully extended product was the most prevalent (data not shown). With the larger template, products >800 nt were present in the greatest proportions. Sub-optimal conditions with higher salt or divalent cation concentrations produced a similar profile although the level of primer extension was significantly reduced. Reactions using 4 mM MgCl2 and 500 µM rNTPs showed the same basic trend with a decrease in total primer extension relative to low divalent cation and nucleotides but no change in the synthesis profile (data not shown). The results suggest that the inhibition of primer extension observed with high salt and divalent cation concentrations did not result from an effect on processivity. Overall 3Dpol was highly processive in the presence or absence of 3AB. Since most extended primers presumably reached the end of the template with or without 3AB, any effect that 3AB may have on processivity would have to be evaluated on a much longer template.
DISCUSSION
We have examined the effect of various reaction conditions and viral protein 3AB on primer-dependent RNA synthesis by 3Dpol. The observed optimal conditions for primer extension varied from those typically used in in vitro reactions with 3Dpol. Most experiments have used Mg2+ concentrations between 3 and 5 mM and 400–500 µM rNTP levels (4,11,12,20,25). We observed greater levels of primer extension with lower divalent cation and rNTP concentrations. Other work directly addressing in vitro primer-dependent synthesis by 3Dpol was consistent with results we observed at higher rNTP concentrations (500 µM) (25). In this work, with the higher rNTPs, optimal divalent cation concentrations of 5 mM were found and both Mn2+ and Mg2+ supported 3Dpol activity.
Several groups have studied 3Dpol synthesis on viral genomic RNA either primed with oligo(U) or using material from cell-free extracts (4,16,21,42–49). It is possible that RNA synthesis could be enhanced in these assays by using optimal conditions reported here; however, this would have to be determined. Perhaps higher divalent cation and/or salt concentrations are required for optimal folding of the genomic RNA or for interactions with components present in cell extracts. The role of genomic secondary structures in viral replication has been clearly demonstrated (9,13–15). The formation of these structures and the interactions of viral and host proteins with them could be sensitive to the concentrations of divalent cation and salt.
It was interesting that 3Dpol appeared to have at least two distinct sets of conditions where high levels of primer extension activity were observed. In the case of reactions with low divalent cation, very little primer extension was observed at 250 µM or greater rNTP concentrations. In contrast, using high divalent cation (3–5 mM) required high rNTPs (500 µM) for maximal activity. Despite the apparent differences, both sets of conditions support a mechanism where the association or activity of 3Dpol with the primer-template requires free divalent cation and is favored by a low concentration of free cation. In this regard the activity observed with higher Mg2+ concentrations is enhanced by higher rNTP concentrations that could act to ‘buffer’ or complex the Mg2+ resulting in a lower level of free cation. With low Mg2+ concentrations high rNTPs essentially ‘chelate’ all the available divalent cation leading to little or no free Mg2+.
It is difficult to speculate which in vitro conditions more closely reflect cellular conditions. Estimates of rNTP concentrations in the cytoplasm of cells vary widely depending on the cell type among other factors. Most measurements in non-transformed cells suggest very high concentrations of ATP (>2 mM), ∼10-fold less GTP and UTP, and CTP concentrations in the 100 µM range. Estimates for Mg2+ suggest a total concentration of ∼9 mM of which the vast majority is complexed to various cellular compounds (8 mM) leaving only a small amount of free Mg2+ (reviewed in 50). These results imply that the total rNTP concentration in cells is more closely reflected by the higher rNTP amounts in in vitro reactions (500 µM) for some nucleotides (ATP) but the lower amounts for others (CTP). The level of free Mg2+ in the cell more closely reflects the conditions for optimal synthesis reported in this work although the total Mg2+ in cells is closer to the 3–5 mM range used in most work. Another factor is the salt concentrations in cells, which are much higher than those required for optimal activity in vitro. In vitro reactions were inhibited by salt concentrations >20 mM regardless of the concentration of divalent cation or rNTPs used (data not shown). It seems unlikely that infections with picornaviruses have a large effect on any of the parameters noted above, especially early in infection. However, the conditions within the vesicular structures where replication is proposed to occur could be different from those in the cytoplasm. Perhaps most notable is the fact that sub-optimal conditions, although greatly affecting the level of primer extension, did not seem to significantly change the processivity of 3Dpol (Fig. 6). In effect the polymerase appears to be insensitive to conditions during the extension phase. Since 3Dpol likely has cofactors that enhance and provide specificity to the initiation step (see Introduction), sub-optimal conditions for primer-dependent initiation may not be a factor in the cell (see below).
As was noted above, results showed that the initiation of primer extension is highly sensitive to reaction conditions while polymerase extension is relatively insensitive. In addition, the high processivity values imply that 3Dpol is tightly associated with the primer-template during extension. Also consistent with high processivity were the koff values measured for 3Dpol in the presence and absence of 3AB. If it is assumed that the values measured with the primer-template are indicative of the off-rate for most regions of RNA during polymerase extension, then the off-rate and polymerization rate can be used to estimate processivity (51). The polymerization rate under optimal conditions in our assays was ∼200 nt/min (data not shown) and is similar to those found by others using heteropolymeric templates (21). The estimated processivity values can be determined by dividing the polymerization rates by the koff. This yields values of ∼5400 (200/0.037) and 18 000 (200/0.011) in the absence and presence of 3AB, respectively. These values are close to the length of the poliovirus genome suggesting that an ‘average’ polymerase molecule could complete genomic synthesis in a single binding event.
The effect of 3AB on the koff rate of 3Dpol suggested a stabilization of 3Dpol binding to primer-template. However, there is a second possibility: 3AB causes 3Dpol to be less labile. The t1/2 of 3Dpol at 42°C in the absence of nucleotides has been reported to be ∼1 min (52) while recent data indicates the presence of primer-template and nucleotides significantly stabilizes the enzyme (24). In our koff assays 3Dpol was pre-bound to primer-template and thus stabilized. Despite this it is not clear if the determined off-rates reflect dissociation of the polymerase from the primer-template or a combination of dissociation and loss of activity. If the latter is the case then the apparent decrease in off-rate in the presence of 3AB could have resulted from 3AB protecting the activity of the enzyme. Given the unstable nature of 3Dpol in the absence of primer-template it is not possible to directly differentiate between the two possibilities. The fact that enhancement of primer extension is also observed at early time points in off-rate assays with 3AB clearly indicates that 3AB initially enhances the association of 3Dpol with the primer-template (Figs 6 and 7). With this in mind stabilization of the polymerase–primer–template complex is probably, at least in part, the reason for the observed lowering of koff.
It was also noted that a relatively low proportion of available primers were extended in 3Dpol reactions (see Results). Typically a polymerase would bind the primer-template, carry out extension, dissociate and then bind other primer-templates. Since the concentration of enzyme in reactions was not limiting, this cycling should have resulted in the extension of essentially all template-bound primers, regardless of the apparent low Kd of 3Dpol for primer-template. However, the slow release (low koff) of enzyme from the primer-template may limit recycling. In addition, enzyme molecules that do not initially associate with primer-template could rapidly lose activity due to the labile nature of free 3Dpol. In essence then, the high Kd value limits the enzyme that initially binds primer-template, much of the remaining enzyme likely loses activity. Enzymes bound to the primer-template are stable but recycle slowly because of the low off-rate. With respect to this, others have reported that dissociation is ∼10-fold more rapid from the end of the template than internal regions (24). Therefore, after completion of primer extension, although the enzyme would dissociate ‘relatively’ quickly, turnover would still be of the order of several minutes on average. After dissociation, enzyme molecules are once again subjected to low rebinding due to the high Kd and rapid loss of activity. The low level of recycling under optimal conditions was evident from the results shown in Figures 3 and 4. Reactions with trap showed about half the level of primer extension relative to those without. This suggests that enzymes recycle at most once in the course of a 1 h reaction. Higher divalent cation and nucleotide concentrations seemed to favor recycling and/or stabilization of the enzyme, but primer extension was limited by the lower level of active enzyme that initially associated with the primer.
Results presented here suggest that 3Dpol is a highly processive polymerase that initiates polymerization on primers relatively poorly. The poor initiation seems to result at least in part from a low affinity (high Kd) for the primer-template. This is supported by the significant enhancement of primer extension at higher primer-template concentrations (Fig. 3). Despite the apparent high Kd, the koff values for 3Dpol were low (Fig. 5). Since Kd = koff/kon, this implies a slow association (kon) and/or slow formation of an active enzyme complex. Consistent with this, others have shown that the association of 3Dpol with primer-template and subsequent isomerization to form an active catalytic complex are relatively slow, while the catalytic complex itself is very stable (24). Given the stable nature of the catalytic complex it is likely that 3AB enhances primer extension by increasing the association and/or isomerization steps rather than stabilizing the catalytic complex. The fact that our results suggest that the complex is stabilized indicates that 3AB may also influence this step in polymerization. Such stabilization may be important in the cellular environment where the salt, divalent cation and temperature conditions would appear to be sub-optimal based on in vitro data.
The highly processive nature of 3Dpol suggest that the enzyme would require only a single binding event to complete genomic RNA synthesis. If RNA synthesis were not completed in a single binding event, rebinding would have to occur in order to complete synthesis. Clearly 3Dpol is capable of binding primer and continuing synthesis both in vitro and in vivo. The latter is strongly suggested by evidence for template switch type recombination events (53–56). However, our results suggest that the polymerase alone initiates synthesis relatively poorly. From this it appears that 3Dpol has evolved to be highly processive, yet relatively poor at initiating primer-dependent RNA synthesis. The low initiation is solved by three factors: (i) a cofactor (3AB) that promotes association with the template, (ii) specific sequence/structural information within the genome that may promote interactions with host and viral factors resulting in efficient and specific initiation in the absence of a true primer, and (iii) the high concentration of viral nucleic acids and other factors in the vesicular structures where replication occurs (24). Note that the properties of 3Dpol are different from some other RNA virus polymerases like reverse transcriptases that associate rapidly with primer-template while possessing relatively low processivity (51,57,58). The high processivity of 3Dpol and other picornavirus polymerases may have evolved due to the fact that these viruses do not replicate in a capsid complex where the polymerase molecule would be retained after dissociating from the genome. In such structures the close proximity of the genome to the polymerase likely promotes efficient rebinding, precluding the need for high processivity.
Acknowledgments
ACKNOWLEDGEMENTS
This work was supported in part by a National Institutes of Health Fellowship (GM18272) awarded to V.R.-W.
References
- 1.Rueckert R.R. (1990) In Fields,B.N. and Knipe,D.M. (eds), Virology, 2nd Edn. Raven Press, New York, NY, Vol. 1, pp. 507–548. [Google Scholar]
- 2.Houghton M. (1996) In Fields,B.N., Knipe,D.M. and Howley,P.M. (eds), Virology, 3rd Edn. Lippincott-Raven, Philidelphia, PA, Vol. 1, pp. 1035–1058.
- 3.Buck K.W. (1996) Comparison of the replication of positive-stranded RNA viruses of plants and animals. Adv. Virus Res., 47, 159–251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Plotch S.J., Palant,O. and Gluzman,Y. (1989) Purification and properties of poliovirus RNA polymerase expressed in Escherichia coli. J. Virol., 63, 216–225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Neufeld K.L.R., Richards,O.C. and Ehrenfeld,E. (1991) Purification, characterization and comparison of poliovirus RNA polymerase from native and recombinant sources. J. Biol. Chem., 266, 24214–24219. [PubMed] [Google Scholar]
- 6.Ishihama A. and Barbier,P. (1994) Molecular anatomy of viral RNA-directed RNA polymerases. Arch. Virol., 134, 235–258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Behrens S.E., Tomei,L. and De Francesco,R. (1996) Identification and properties of the RNA-dependent RNA polymerase of hepatitis C virus. EMBO J., 15, 12–22. [PMC free article] [PubMed] [Google Scholar]
- 8.Vazquez A.L., Martin Alonso,J.M., Casais,R., Boga,J.A. and Parra,F. (1998) Expression of enzymatically active rabbit hemorrhagic disease virus RNA-dependent RNA polymerase in Escherichia coli. J. Virol., 72, 2999–3004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Andino R., Rieckhof,G.E., Achacoso,P.L. and Baltimore,D. (1993) Poliovirus RNA synthesis utilizes an RNP complex formed around the 5′-end of viral RNA. EMBO J., 12, 3587–3598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Hope D.A., Diamond,S.E. and Kirkegaard,K. (1997) Genetic dissection of interaction between poliovirus 3D polymerase and viral protein 3AB. J. Virol., 71, 9490–9498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Paul A.V., Cao,X., Harris,K.S., Lama,J. and Wimmer,E. (1994) Studies with poliovirus 3Dpol: stimulation of poly (U) synthesis in vitro by purified poliovirus protein 3AB. J. Biol. Chem., 269, 29173–29181. [PubMed] [Google Scholar]
- 12.Richards O.C. and Ehrenfeld,E. (1998) Effects of poliovirus 3AB protein on 3D polymerase-catalyzed reaction. J. Biol. Chem., 273, 12832–12840. [DOI] [PubMed] [Google Scholar]
- 13.Xiang W., Cuconati,A., Paul,A.V., Cao,X. and Wimmer,E. (1995) Molecular dissection of the multifunctional poliovirus RNA-binding protein 3AB. RNA, 1, 892–904. [PMC free article] [PubMed] [Google Scholar]
- 14.Harris K.S., Xiang,W., Alexander,L., Lane,W.S., Paul,A.V. and Wimmer,E. (1994) Interaction of poliovirus polypeptide 3CDpro with the 5′ and 3′ termini of the poliovirus genome. Identification of viral and cellular cofactors needed for efficient binding. J. Biol. Chem., 269, 27004–27014. [PubMed] [Google Scholar]
- 15.Gamarnik A.V. and Andino,R. (2000) Interactions of viral protein 3CD and Poly(rC) binding protein with the 5′ untranslated region of the poliovirus genome. J. Virol., 74, 2219–2226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Takegami T., Semler,B.L., Anderson,C.W. and Wimmer,E. (1983) Membrane fractions active in poliovirus RNA replication contain VPg precursor polypeptides. Virology, 128, 33–47. [DOI] [PubMed] [Google Scholar]
- 17.Semler B.L., Anderson,C.W., Hanecak,R., Dorner,L.F. and Wimmer,E. (1982) A membrane-associated precursor to poliovirus VPg identified by immunoprecipitation with antibodies directed against a synthetic heptapeptide. Cell, 28, 405–412. [DOI] [PubMed] [Google Scholar]
- 18.Baron M.H. and Baltimore,D. (1982) Anti-VPg antibody inhibition of the poliovirus replicase reaction and production of covalent complexes of VPg-related proteins and RNA. Cell, 30, 745–752. [DOI] [PubMed] [Google Scholar]
- 19.Paul A.V., Boom,J.H., Filippov,D. and Wimmer,E. (1998) Protein-primed RNA synthesis by purified RNA polymerase. Nature, 393, 280–284. [DOI] [PubMed] [Google Scholar]
- 20.Lama J., Sanz,M.A. and Rodriguez,P.L. (1995) Role of 3AB protein in poliovirus genome replication. J. Biol. Chem., 270, 14430–14438. [DOI] [PubMed] [Google Scholar]
- 21.Plotch S.J. and Palant,O. (1995) Poliovirus protein 3AB form a complex with and stimulates the activity of the viral RNA polymerase, 3Dpol. J. Virol., 69, 7169–7179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Towner J.S., Ho,T.V. and Semler,B.L. (1996) Determinants of membrane association for poliovirus protein 3AB. J. Biol. Chem., 271, 26810–26818. [DOI] [PubMed] [Google Scholar]
- 23.Rodriguez-Wells V., Plotch,S.J. and DeStefano,J.J. (2001) Determination of the mutation rate of polivirus RNA-dependent RNA polymerase. Virus Res., 74, 119–132. [DOI] [PubMed] [Google Scholar]
- 24.Arnold J.J. and Cameron,C.E. (2000) Poliovirus RNA-dependent RNA polymerase (3D(pol)). Assembly of stable, elongation-competent complexes by using a symmetrical primer-template substrate (sym/sub). J. Biol. Chem., 275, 5329–5336. [DOI] [PubMed] [Google Scholar]
- 25.Arnold J.J., Kumar,S., Ghosh,B. and Cameron,C.E. (1999) Poliovirus RNA-dependent RNA polymerase (3Dpol). J. Biol. Chem., 274, 37060–37069. [DOI] [PubMed] [Google Scholar]
- 26.Ward C.D., Stokes,M.A. and Flanegan,J.B. (1988) Direct measurement of the poliovirus polymerase error frequency in vitro. J. Virol., 66, 558–562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Hansen J.L., Long,A.M. and Schultz,S.C. (1997) Structure of the RNA-dependent RNA polymerase of poliovirus. Structure, 5, 1109–1122. [DOI] [PubMed] [Google Scholar]
- 28.Neufeld K.L., Galarza,J.M., Richards,O.C., Summers,D.F. and Ehrenfeld,E. (1994) Identification of terminal adenylyl transferase activity of the poliovirus polymerase 3Dpol. J. Virol., 68, 5811–5818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Beckman M.T. and Kirkegaard,K. (1998) Site size of cooperative single-stranded RNA binding by poliovirus RNA-dependent RNA polymerase. J. Biol. Chem., 273, 6724–6730. [DOI] [PubMed] [Google Scholar]
- 30.Molla A., Harris,K.S., Paul,A.V., Shin,S.H., Mugavero,J. and Wimmer,E. (1994) Stimulation of poliovirus proteinase 3Cpro-related proteolysis by the genome-linked protein VPg and its precursor 3AB. J. Biol. Chem., 269, 27015–27020. [PubMed] [Google Scholar]
- 31.Bravo R., Frank,R., Blundell,P.A. and Macdonald-Bravo,H. (1987) Cyclin/PCNA is the auxiliary protein of DNA polymerase-δ. Nature, 326, 515–517. [DOI] [PubMed] [Google Scholar]
- 32.Prelich G., Tan,C.K., Kostura,M., Mathews,M.B., So,A.G., Downey,K.M. and Stillman,B. (1987) Functional identity of proliferating cell nuclear antigen and a DNA polymerase-δ auxiliary protein. Nature, 326, 517–520. [DOI] [PubMed] [Google Scholar]
- 33.Tan C.K., Castillo,C., So,A.G. and Downey,K.M. (1986) An auxiliary protein for DNA polymerase-δ from fetal calf thymus. J. Biol. Chem., 261, 12310–12316. [PubMed] [Google Scholar]
- 34.Weisshart K., Chow,C. and Coen,D.,M. (1999) Herpes simplex virus factor UL42 imparts increased DNA-binding specificity to the viral DNA polymerase and decreased dissociation from primer-template without reducing the elongation rate. J. Virol., 73, 55–66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Arnold J.J. and Cameron,C.E. (1999) Polivirus RNA-dependent RNA polymerase (3Dpol) is sufficient for template switching in vitro. J. Biol. Chem., 274, 2706–2716. [DOI] [PubMed] [Google Scholar]
- 36.Parsley T.B., Towner,J.S., Blyn,L.B., Ehrenfeld,E. and Semler,B.L. (1997) Poly (rC) binding protein 2 forms a ternary complex with the 5′-terminal sequences of poliovirus RNA and the viral 3CD proteinase. RNA, 3, 1124–1134. [PMC free article] [PubMed] [Google Scholar]
- 37.Sambrook J., Fritsh,E.F. and Maniatis,T. (1989) Molecular Cloning, A Laboratory Manual, 2nd Edn. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
- 38.Lubinski J.M., Ransone,L.J. and Dasgupta,A. (1987) Primer-dependent synthesis of covalently linked dimeric RNA molecules by poliovirus replicase. J. Virol., 61, 2997–3003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Pata J.D., Schultz,S.C. and Kirkegard,K. (1995) Functional oligomerization of poliovirus RNA-dependent RNA polymerase. RNA, 1, 466–477. [PMC free article] [PubMed] [Google Scholar]
- 40.Paul A.V., Cao,X., Harris,K.S., Lama,J. and Wimmer,E. (1994) Studies with poliovirus polymerase 3Dpol. Stimulation of poly(U) synthesis in vitro by purified poliovirus protein 3AB. J. Biol. Chem., 269, 29173–29181. [PubMed] [Google Scholar]
- 41.Lama J., Paul,A.V., Harris,K.S. and Wimmer,E. (1994) Properties of purified recombinant poliovirus protein 3AB as substrate for viral proteinases and as co-factor for RNA polymerase 3Dpol. J. Biol. Chem., 269, 66–70. [PubMed] [Google Scholar]
- 42.Van Dyke T.A., Rickles,R.J. and Flanegan,J.B. (1982) Genome-length copies of poliovirion RNA are synthesized in vitro by the poliovirus RNA-dependent RNA polymerase. J. Biol. Chem., 257, 4610–4617. [PubMed] [Google Scholar]
- 43.Tuschall D.M., Hiebert,E. and Flanegan,J.B. (1982) Poliovirus RNA-dependent RNA polymerase synthesizes full-length copies of poliovirion RNA, cellular mRNA, and several plant virus RNAs in vitro. J. Virol., 44, 209–216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Toyoda H., Yang,C.F., Takeda,N., Nomoto,A. and Wimmer,E. (1987) Analysis of RNA synthesis of type 1 poliovirus by using an in vitro molecular genetic approach. J. Virol., 61, 2816–2822. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Takegami T., Kuhn,R.J., Anderson,C.W. and Wimmer,E. (1983) Membrane-dependent uridylylation of the genome-linked protein VPg of poliovirus. Proc. Natl Acad. Sci. USA, 80, 7447–7451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Flanegan J.B. and Van Dyke,T.A. (1979) Isolation of a soluble and template-dependent poliovirus RNA polymerase that copies virion RNA in vitro. J. Virol., 32, 155–161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Etchison D. and Ehrenfeld,E. (1981) Comparison of replication complexes synthesizing poliovirus RNA. Virology, 111, 33–46. [DOI] [PubMed] [Google Scholar]
- 48.Dasgupta A. (1983) Purification of host factor required for in vitro transcription of poliovirus RNA. Virology, 128, 245–251. [DOI] [PubMed] [Google Scholar]
- 49.Baron M.H. and Baltimore,D. (1982) In vitro copying of viral positive strand RNA by poliovirus replicase. Characterization of the reaction and its products. J. Biol. Chem., 257, 12359–12366. [PubMed] [Google Scholar]
- 50.Traut T.W. (1994) Physiological concentrations of purines and pyrimidines. Mol. Cell. Biochem., 140, 1–22. [DOI] [PubMed] [Google Scholar]
- 51.Reardon J.E. (1993) Human immunodeficiency virus reverse transcriptase A kinetic analysis of RNA-dependent and DNA-dependent DNA polymerization. J. Biol. Chem., 268, 8743–8751. [PubMed] [Google Scholar]
- 52.Richards O.C., Yu,P., Neufeld,K.L. and Ehrenfeld,E. (1992) Nucleotide binding by the poliovirus RNA polymerase. J. Biol. Chem., 267, 17141–17146. [PubMed] [Google Scholar]
- 53.Romanova L.I., Blinov,V.M., Tolskaya,E.A., Viktorova,E.G., Kolesnikova,M.S., Guseva,E.A. and Agol,V.I. (1986) The primary structure of crossover regions of intertypic poliovirus recombinants: a model of recombination between RNA genomes. Virology, 155, 202–213. [DOI] [PubMed] [Google Scholar]
- 54.Nagy P.D. and Simon,A.E. (1997) New insights into the mechanisms of RNA recombination. Virology, 235, 1–9. [DOI] [PubMed] [Google Scholar]
- 55.Jarvis T.C. and Kirkegaard,K. (1992) Poliovirus RNA recombination: mechanistic studies in the absence of selection. EMBO J., 11, 3135–3145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Kirkegaard K. and Baltimore,D. (1986) The mechanism of RNA recombination in poliovirus. Cell, 47, 433–443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Huber H.E., McCoy,J.M., Seehra,J.S. and Richardson,C.C. (1989) Human immunodeficiency virus 1 reverse transcriptase template binding, processivity, strand displacement synthesis, and template switching. J. Biol. Chem., 264, 4669–4678. [PubMed] [Google Scholar]
- 58.DeStefano J.J., Buiser,R.G., Mallaber,L.M., Fay,P.J. and Bambara,R.A. (1992) Parameters that influence processive synthesis and site-specific termination by human immunodeficiency virus reverse transcriptase on RNA and DNA templates. Biochim. Biophys. Acta, 1131, 270–280. [DOI] [PubMed] [Google Scholar]