


Abstract
In the yeast Saccharomyces cerevisiae, DNA polymerase ζ (Polζ) is required in a major lesion bypass pathway. To help understand the role of Polζ in lesion bypass, we have performed in vitro biochemical analyses of this polymerase in response to several DNA lesions. Purified yeast Polζ performed limited translesion synthesis opposite a template TT (6-4) photoproduct, incorporating A or T with similar efficiencies (and less frequently G) opposite the 3′ T, and predominantly A opposite the 5′ T. Purified yeast Polζ predominantly incorporated a G opposite an acetylaminofluorene (AAF)-adducted guanine. The lesion, however, significantly inhibited subsequent extension. Furthermore, yeast Polζ catalyzed extension DNA synthesis from primers annealed opposite the AAF-guanine and the 3′ T of the TT (6-4) photoproduct with varying efficiencies. Extension synthesis was more efficient when A or C was opposite the AAF-guanine, and when G was opposite the 3′ T of the TT (6-4) photoproduct. In contrast, the 3′ T of a cis–syn TT dimer completely blocked purified yeast Polζ, whereas the 5′ T was readily bypassed. These results support the following dual-function model of Polζ. First, Polζ catalyzes nucleotide incorporation opposite AAF-guanine and TT (6-4) photoproduct with a limited efficiency. Secondly, more efficient bypass of these lesions may require nucleotide incorporation by other DNA polymerases followed by extension DNA synthesis by Polζ.
INTRODUCTION
DNA can be damaged by a variety of physical and chemical agents, such as UV radiation and acetylaminofluorene (AAF). DNA repair constitutes an important defense system by removing the lesions from DNA. However, some DNA lesions can persist in the genome during replication due to limited cellular repair and/or newly formed damage at the S phase of the cell cycle. Since many lesions block replicative DNA polymerases, cells have evolved a damage tolerance response to enable replication of the damaged DNA templates. Lesion bypass represents one of the damage tolerance mechanisms, and requires a DNA polymerase to copy the damaged DNA template. DNA synthesis (nucleotide incorporation) opposite a template lesion is also referred to as translesion synthesis. Depending on the outcome, translesion synthesis is further divided into error-free and error-prone translesion syntheses. While the former predominantly incorporates the correct nucleotide opposite the lesion, thus is a mutation-avoiding mechanism, the latter frequently incorporates an incorrect nucleotide opposite the lesion, thus is a mutation-generating mechanism.
In Escherichia coli, DNA polymerase V is a major translesion synthesis polymerase, which functions in the UmuDC mutagenesis pathway (1,2). In vitro, DNA polymerase V is capable of error-prone translesion synthesis opposite several lesions (1–3). In eukaryotes, DNA polymerase η (Polη) is involved in error-free lesion bypass of TT dimers (4–6). Thus, Polη functions as an important anti-mutagenesis mechanism in response to UV radiation in humans. Deficiency in Polη activity will lead to the human hereditary disease xeroderma pigmentosum variant (XPV) (6,7), which is characterized by photosensitivity and a predisposition to skin cancer (8). Error-prone translesion synthesis by purified Polη has also been observed in vitro in response to certain lesions such as (+)-trans-anti-benzo[a]pyrene-N2-dG adducts (9). Most likely, Polη functions as both an error-free and error-prone translesion synthesis polymerase in eukaryotic cells, depending on the specific lesion (10). Consistent with its translesion synthesis activity, Polη is an extraordinarily low fidelity polymerase with a low processivity and lacks 3′→5′ proofreading exonuclease activity (10–12).
Polζ is another eukaryotic DNA polymerase required in a major error-pone lesion bypass pathway (13), which is also referred to as the Polζ mutagenesis pathway. Rev3 is the catalytic subunit of Polζ (13). Rev7 forms a complex with Rev3 protein and is considered a subunit of Polζ (13). Additionally, the Polζ mutagenesis pathway also requires Rad6, Rad18 and Rev1 (14–18). Rad6 is a ubiquitin conjugating enzyme (E2) and forms a complex with Rad18 protein (19,20). Rev1 is a dCMP transferase capable of incorporating a C opposite a template G or an abasic site (21). The human RAD6 (HHR6A and HHR6B), RAD18, REV1, REV3 and REV7 genes have been isolated (22–29). The human REV3 protein is about twice the size of its yeast counterpart, as a result of additional sequences at the N-terminal 2/3 regions of the human protein (25,27). The significance, if any, of this size difference between the yeast and the human REV3 proteins is not known. The RAD6–RAD18 and REV3–REV7 interactions, and the dCMP transferase activity of REV1 are all conserved in humans (23,24,26,29). Furthermore, UV-induced mutagenesis requires both REV1 and REV3 gene expression in cultured human cells (27,28). Thus, the Polζ mutagenesis pathway is probably functional in humans. In addition to Polζ and Polη, it appears that Polκ and Polι are also translesion synthesis polymerases in humans (30–36). Homologs of these two DNA polymerases are not found in Saccharomyces cerevisiae.
It has been proposed that Polζ is generally responsible for error-prone translesion synthesis step during lesion bypass, and Rev1 is responsible for nucleotide incorporation opposite template apurinic/apyrimidinic (AP) sites (13,21,37). However, two in vitro studies with yeast Polζ have yielded conflicting results. Nelson et al. (13) reported that purified yeast Polζ is able to bypass a template cis–syn TT dimer, whereas Johnson et al. (34) reported that yeast Polζ is unable to incorporate a nucleotide opposite this lesion. In an earlier study we found that, following one nucleotide incorporation by purified yeast Polη opposite a template AP site, yeast Polζ is able to extend DNA synthesis from opposite the lesion (10). Thus, AP site bypass was achieved in vitro by the combined activities of Polη nucleotide incorporation and subsequent Polζ DNA extension (10). Based on this two-polymerase two-step model of Yuan et al. (10), Johnson et al. (34) later observed in vitro bypass of an AP site and a TT (6-4) photoproduct by combined actions of human Polι and yeast Polζ. Johnson et al. (34) further concluded that Polζ is an ‘extender’ rather than an ‘inserter’ during lesion bypass. These limited studies did not yield a clear understanding of Polζ in lesion bypass. To define the precise role of Polζ in lesion bypass, many more biochemical analyses of this polymerase in response to additional DNA lesions are needed.
To help understand the role of Polζ in lesion bypass, we have performed in vitro biochemical analyses of this polymerase in response to several DNA lesions. In this report, we show that (i) purified yeast Polζ is able to perform error-prone translesion synthesis opposite a template TT (6-4) photoproduct and an AAF-adducted guanine (AAF-G) to a limited extent, but it is unresponsive to a template cis–syn TT dimer due to Polζ blockage by the modified 3′ T and (ii) yeast Polζ is capable of extension DNA synthesis from primers annealed opposite these lesions. These results led to a dual-function model of Polζ, in which Polζ functions both as a nucleotide incorporation polymerase opposite some lesions and as an extension DNA synthesis polymerase during lesion bypass by the two-polymerase two-step mechanism.
MATERIALS AND METHODS
Materials
A mouse monoclonal antibody against the His6 tag was obtained from Qiagen (Valencia, CA). Alkaline phosphatase conjugated anti-mouse IgG was obtained from Sigma Chemical Co (St Louis, MO). Taq DNA polymerase was purchased from BRL (Bethesda, MD). N-acetoxy-N-2-acetylaminofluorene (AAAF, the activated form of AAF) was obtained from the Midwest Research Institute (Kansas City, MO).
Damaged DNA templates
A 49-nt DNA template containing a site-specific cis–syn TT dimer or a TT (6-4) photoproduct was prepared as previously described (38). Its sequence was 5′-AGCTACCATGCCTGCACGAATTAAGCAATTCGTAATCATGGTCATAGCT-3′, where the modified TT is underlined. AAF-adducted DNA template was prepared by incubating 2 nmol of the oligonucleotide 5′-CCTTCTTCATTCGAACATACTTCTTCTTCC-3′ with 200 nmol of AAAF at 37°C in the dark for 3 h in 100 µl of TE buffer (10 mM Tris–HCl, pH 7.5, 1 mM EDTA) containing 20% ethanol, followed by purification as previously described (31).
Overexpression plasmids of the yeast REV3 and REV7 genes
The yeast REV3 gene was obtained by polymerase chain reaction (PCR) amplification from S.cerevisiae DNA using Taq DNA polymerase and two primers, 5′-CGGGATCCATGTCGAGGGAGTCGAACGAC-3′ and 5′-CGCGTCGACCCAATCATTTAGAGATATTAATGCTTCTTCC-3′. The resulting 4.5 kb PCR product was then cloned into the BamHI and SalI sites of the vector pEGTh6, yielding pEGTh6-REV3. The yeast REV7 gene was obtained by PCR amplification from S.cerevisiae DNA using Taq DNA polymerase and two primers, 5′-GCTCTAGAATGAATAGATGGGTAGAGAAGTGGCTG-3′ and 5′-CCGCTCGAGCCATGGCTGCAGAAACAAAGATCCAAAAATGCTCTC-3′. The resulting 740 bp PCR product was then cloned into the XbaI and PstI sites of the vector pEGUh6, yielding pEGUh6-REV7. The cloned yeast REV3 and REV7 genes in the expression constructs were confirmed by functional complementation of the deficient UV-mutagenesis of yeast rev3 and rev7 deletion mutant strains, respectively. Yeast expression vectors pEGTh6 and pEGUh6 were derived from Yeplac112 and Yeplac195, respectively (39), which contained the yeast TRP1 gene and the yeast URA3 gene, respectively, for plasmid selection. Additionally, both vectors contained the 2 µm origin for multicopy plasmid replication, the yeast GAL1/10 promoter for inducible gene expression and six histidine codons for N-terminal protein tagging.
Purification of yeast DNA Polζ
Yeast Polζ is composed of the Rev3 (the catalytic subunit) and Rev7 proteins in a complex (13). Yeast cells harboring both pEGTh6-REV3 and pEGUh6-REV7 were grown to stationary phase in minimal medium containing 2% sucrose. Induction of Polζ expression was achieved by diluting the culture 10-fold in 16 l of YPG (2% bacto-peptone, 1% yeast extract and 2% galactose) medium supplemented with 0.5% sucrose and growth for 15 h at 30°C. Cells were collected by centrifugation and washed in water. After resuspending in an extraction buffer containing 50 mM Tris–HCl, pH 7.5, 1 M KCl, 10% sucrose, 5 mM β-mercaptoethanol, and protease inhibitors (23), cells were disrupted by Zirconium beads in a bead-beater (Biospec Products, Bartlesville, OK) for 15 pulses of 30 s each on ice. The clarified extract (∼120 ml) was loaded onto two connected HiTrap chelating columns (5 ml each) charged with NiSO4 (Amersham Pharmacia Biotech, Piscataway, NJ), followed by washing the columns sequentially with 100 ml Ni buffer A (20 mM phosphate buffer, pH 7.4, 0.5 M NaCl, 10% glycerol, 5 mM β-mercaptoethanol, and protease inhibitors) containing 10 mM imidazole and 100 ml Ni buffer A containing 35 mM imidazole. Bound proteins were eluted with a linear gradient (250 ml) of 35–108 mM imidazole in Ni buffer A. Pooled Polζ fractions were concentrated by dialysis against solid polyethylene glycol 10 000 overnight and desalted through five connected HiTrap Sephadex G-25 columns (5 ml each) (Amersham Pharmacia Biotech) in FPLC buffer A (50 mM Tris–HCl, pH 7.5, 10% glycerol, 5 mM β-mercaptoethanol) containing 80 mM KCl. The resulting sample (∼50 ml) was loaded onto an FPLC Mono S HR5/5 column (Amersham Pharmacia Biotech) equilibrated with FPLC buffer A containing 80 mM KCl. Bound proteins were eluted with 30 ml linear gradient of 80–500 mM KCl in FPLC buffer A. Pooled Mono S fractions (∼5 ml) were concentrated by polyethylene glycol 10 000 and loaded onto an FPLC Superdex 200 gel filtration column equilibrated with FPLC buffer A containing 300 mM KCl. During Polζ purification, the yeast Rev3 and Rev7 proteins were identified by western blot analyses using a mouse monoclonal antibody against the His6 tag.
DNA polymerase assays
The standard DNA polymerase reaction (10 µl) contained 25mM KH2PO4 pH 7.0, 5 mM MgCl2, 5 mM dithiothreitol, 100 µg/ml bovine serum albumin, 10% glycerol, 50 µM each dATP, dCTP, dTTP and dGTP, 50 fmol of a primed DNA template and purified yeast Polζ. DNA primers were labeled at their 5′ ends by [γ-32P]ATP and T4 polynucleotide kinase. After incubation at 30°C for 30 min, reactions were terminated with 7 µl of a stop solution (20 mM EDTA, 95% formamide, 0.05% bromophenol blue and 0.05% xylene cyanol). Reaction products were separated by electrophoresis on a 20% polyacrylamide gel containing 8 M urea and visualized by autoradiography.
Kinetic measurements of primer extension by yeast Polζ
Kinetic analysis of primer extension by yeast Polζ was performed using a previously described method (30,40,41). Briefly, DNA polymerase assays were performed using 50 fmol of a primed DNA template, 10 ng (50 fmol) of purified yeast Polζ and increasing concentrations of dGTP (0.1–300 µM). Four DNA primers, 5′-GGAAGAAGAAGTATGTTC-3′, 5′-GGAAGAAGAAGTATGTTA-3′, 5′-GGAAGAAGAAGTATGTTT-3′ and 5′-GGAAGAAGAAGTATGTTG-3′, were labeled at their 5′ ends with 32P and separately annealed to the AAF-adducted DNA template with the primer 3′ nucleotide opposite the template AAF-G. The next template base was a C. After incubation for 10 min at 30°C under standard DNA polymerase assay conditions, reaction products were separated by electrophoresis on a 20% denaturing polyacrylamide gel. The percentage of primers extended by the polymerase was calculated following scanning densitometry of the extended DNA band and the remaining primer band on the autoradiogram. Product formed (P) was derived from the calculation P = fraction of primer extension × 50 fmol. Observed enzyme velocity (v) was obtained from the calculation v = P/10 min. Then, the observed enzyme velocity was plotted as a function of dGTP concentration. The plotted data reflected the Michaelis–Menton kinetics of enzyme reaction, and thus were fitted by a non-linear regression curve to the Michaelis–Menton equation, v = (Vmax × [dNTP])/(Km + [dNTP]), using the SigmaPlot software. Vmax and Km values for the primer extension were obtained from the fitted curves. Relative efficiency of extension (fext) of matched primer versus mismatched primer from opposite the AAF-G was finally calculated from the equation fext = (Vmax/Km)matched/(Vmax/Km)mismatched.
RESULTS
Purification of yeast DNA Polζ
To facilitate protein purification, we tagged both Rev3 and Rev7 with six histidine residues at their N-termini and co-overexpressed both proteins in yeast cells. Plasmids coding for the His-tagged Rev3 and Rev7 fully complemented the yeast rev3 and rev7 mutants, respectively, for UV-induced mutagenesis (data not shown), indicating that the His6 tag did not affect Rev3 and Rev7 functions. Polζ was purified as the Rev3–Rev7 protein complex by liquid chromatography through an affinity Ni–agarose column, an FPLC Mono S column and an FPLC gel filtration column. The most pure fraction of the Rev3–Rev7 complex was analyzed by electrophoresis on a 10% SDS–polyacrylamide gel. As shown in Figure 1A, two major bands of ∼175 and ∼30 kDa, respectively, were detected by silver staining, consistent with the calculated molecular weight of yeast Rev3 (173 kDa) and Rev7 (29 kDa), respectively. Western blotting analysis using a monoclonal antibody against the His6 tag confirmed that the two bands were Rev3 and Rev7, respectively (Fig. 1B). As expected, a DNA polymerase activity was detected with the purified yeast Polζ (Fig. 1C). We consistently observed that the last template base was inefficiently copied by yeast Polζ (Fig. 1C, 30mer DNA band).
Figure 1.

Analyses of purified yeast Polζ. (A) Purified yeast Polζ (300 ng) was analyzed by electrophoresis on a 10% SDS–polyacrylamide gel and visualized by silver staining. Protein size markers (lane M) are indicated on the left. (B) Purified yeast Polζ (300 ng) was analyzed by a western blot using a mouse monoclonal antibody against the His6 tag. (C) A DNA polymerase assay was performed with purified yeast Polζ (26 ng, 129 fmol) using the 30mer DNA template 5′-CCTTCTTCATTCGAACATACTTCTTCTTCC-3′ annealed with the 5′-32P-labeled 17mer primer 5′-GGAAGAAGAAGTATGTT-3′. DNA size markers in nucleotides are indicated on the left.
Lesion bypass of a template TT (6-4) photoproduct by yeast Polζ
Cyclobutane pyrimidine dimers (CPDs) and (6-4) photoproducts are the major lesions induced by UV radiation (42). To ask whether Polζ can replicate UV-damaged DNA, we first examined translesion synthesis activity of purified yeast Polζ opposite a template TT (6-4) photoproduct. A 32P-labeled 15mer primer was annealed to a 49mer DNA template containing a TT (6-4) photoproduct, right before the 3′ T of the lesion (Fig. 2). DNA synthesis assays were then performed with purified yeast Polζ at 30°C for 30 min. As shown in Figure 2 (lane 1), some TT (6-4) photoproducts were bypassed by yeast Polζ. In a separate experiment, we found that purified human Polκ was completely blocked by the TT (6-4) photoproduct in this same DNA template (31), confirming that the DNA substrates used were not contaminated by unmodified templates. To identify the nucleotide incorporated opposite the 3′ T of the TT (6-4) photoproduct, we performed DNA synthesis assays in the presence of only one deoxyribonucleoside triphosphate, dATP, dCTP, dTTP or dGTP individually. As shown in Figure 2 (lanes 2–5), yeast Polζ incorporated A or T with similar efficiencies opposite the 3′ T of the TT (6-4) photoproduct. Less frequently, G was also incorporated (Fig. 2, lane 4). In the presence of dATP alone, translesion synthesis stopped opposite the undamaged template A 5′ to the TT (6-4) photoproduct (Fig. 2, lane 2). The misincorporation of A opposite the undamaged template A suggests that yeast Polζ is not a highly accurate polymerase, consistent with its ability to perform translesion synthesis.
Figure 2.
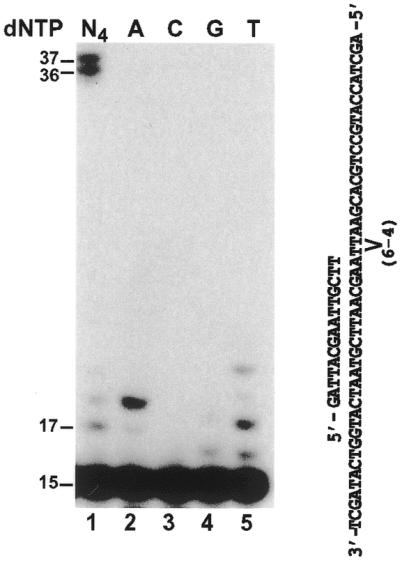
Bypass of a template TT (6-4) photoproduct by yeast Polζ. A 15mer primer was labeled with 32P at its 5′ end and annealed to a DNA template containing a TT (6-4) photoproduct, right before the lesion. DNA polymerase assays were performed with 39 ng (193 fmol) of purified yeast Polζ in the presence of all four dNTPs (lane 1), or a single deoxyribonucleoside triphosphate, dATP (lane 2), dCTP (lane 3), dGTP (lane 4) or dTTP (lane 5). DNA size markers in nucleotides are indicated on the left.
To identify the nucleotide incorporated opposite the 5′ T of the TT (6-4) photoproduct, three 16mer primers were synthesized, labeled with 32P at their 5′ ends, and separately annealed to the damaged DNA template right before the 5′ T of the lesion (Fig. 3A). DNA synthesis assays were then performed with purified yeast Polζ at 30°C for 30 min. As shown in Figure 3B (lanes 1, 6 and 11), the primers were extended by yeast Polζ regardless of whether the primer 3′ end was an A, G or T opposite the 3′ T of the TT (6-4) photoproduct. Since C incorporation by yeast Polζ was not detected opposite the 3′ T of this lesion (Fig. 2, lane 3), extension DNA synthesis from C-terminated primer was not examined. Extension from G opposite the 3′ T of the TT (6-4) photoproduct was most efficient (Fig. 3B, compare lanes 1, 6 and 11). It is also apparent that nucleotide incorporation by yeast Polζ opposite the 5′ T was more efficient than that opposite the 3′ T of the TT (6-4) photoproduct (compare Fig. 2, lane 1 with Fig. 3B, lanes 1, 6 and 11). Furthermore, purified yeast Polζ predominantly incorporated the correct A opposite the 5′ T of the TT (6-4) photoproduct with all three primers (Fig. 3B, lanes 2, 7 and 12). These results indicate that yeast Polζ is capable of bypassing a template TT (6-4) photoproduct with a low efficiency, and that nucleotide incorporation by this polymerase is more error-prone and less efficient opposite the 3′ T compared to the 5′ T of the lesion.
Figure 3.

Nucleotide incorporation by yeast Polζ opposite the 5′ T of the TT (6-4) photoproduct. (A) Three 16mer primers that differed by 1 nt at the 3′ end were labeled with 32P at their 5′ ends and separately annealed to the indicated DNA template containing a TT (6-4) photoproduct, right before the 5′ T of the lesion. (B) DNA polymerase assays were performed with 39 ng (193 fmol) of purified yeast Polζ in the presence of all four dNTPs (N4), or a single deoxyribonucleoside triphosphate, dATP (A), dCTP (C), dGTP (G) or dTTP (T), as indicated. DNA size markers in nucleotides are indicated on the left.
Strong blockage of yeast Polζ by the 3′ T of a template cis–syn TT dimer
Following UV radiation, cis–syn TT dimers are among the CPDs formed in DNA. To examine translesion synthesis activity of purified yeast Polζ opposite a template cis–syn TT dimer, we annealed a 32P-labeled 17mer primer to a 49mer DNA template, 8 nt before the 3′ T of the dimer (Fig. 4A). DNA synthesis assays were then performed with purified yeast Polζ at 30°C for 30 min. As shown in Figure 4A, DNA synthesis by yeast Polζ stopped right before the 3′ T of the TT dimer (25mer DNA band). Additionally, nucleotide incorporation opposite the undamaged template base 3′ to the TT dimer was also inhibited by the lesion, as evidenced by accumulation of the 24mer DNA band (Fig. 4A). DNA synthesis opposite or beyond the TT dimer was not detected even when excessive amounts (150 ng, 743 fmol) of yeast Polζ was used (Fig. 4A). Translesion synthesis was also not detected using a primer annealed right before the lesion (Fig. 4B). In separate experiments, the TT dimer in this same DNA template was efficiently bypassed by purified yeast and human Polη as expected (9,10), confirming the integrity of the template used.
Figure 4.

Response of yeast Polζ to a cis-syn TT dimer in template DNA. (A) A 17mer primer was labeled with 32P at its 5′ end and annealed 8 nt before a template TT dimer as indicated. A DNA polymerase assay was performed with 150 ng (743 fmol) of purified yeast Polζ in the presence of all four dNTPs. (B) A 15mer primer was labeled with 32P at its 5′ end and annealed right before the TT dimer as indicated. DNA polymerase assays were performed with 39 ng (193 fmol) of purified yeast Polζ in the presence of all four dNTPs (lane 1), or a single deoxyribonucleoside triphosphate, dATP (lane 2), dCTP (lane 3), dGTP (lane 4) or dTTP (lane 5). DNA size markers in nucleotides are indicated on the left.
To examine if the 5′ T of the cis–syn TT dimer also blocks yeast Polζ, we annealed a 32P-labeled 16mer primer right before the 5′ T of the dimer (Fig. 5). DNA synthesis assays were then performed with purified yeast Polζ. As shown in Figure 5 (lane 1), yeast Polζ efficiently bypassed the 5′ T of the cis–syn TT dimer. To identify the nucleotide incorporated opposite the 5′ T of the cis–syn TT dimer, we performed DNA synthesis assays in the presence of only one deoxyribonucleoside triphosphate: dATP, dCTP, dTTP or dGTP individually. As shown in Figure 5 (lanes 2–5), yeast Polζ predominantly incorporated the correct A opposite the 5′ T of the cis–syn TT dimer. These results show that yeast Polζ is unable to perform translesion synthesis opposite the 3′ T of a template cis–syn TT dimer, leading to DNA synthesis stop right before the lesion.
Figure 5.
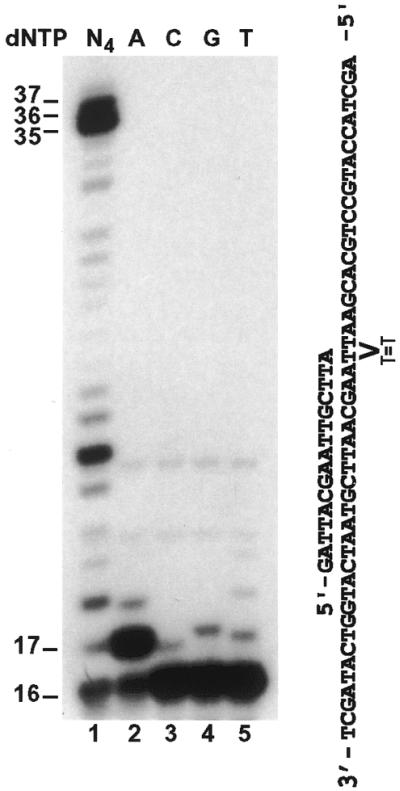
Efficient bypass of the 5′ T of the cis–syn TT dimer by yeast Polζ. A 16mer primer was labeled with 32P at its 5′ end and annealed right before the 5′ T of the TT dimer as indicated. DNA polymerase assays were performed with 39 ng (193 fmol) of purified yeast Polζ in the presence of all four dNTPs (lane 1), or a single deoxyribonucleoside triphosphate, dATP (lane 2), dCTP (lane 3), dGTP (lane 4) or dTTP (lane 5). DNA size markers in nucleotides are indicated on the left.
Translesion synthesis opposite AAF-adducted guanine by yeast Polζ
AAF-Gs are bulky lesions in DNA, which block many DNA polymerases (10,43). To examine whether yeast Polζ is able to perform translesion synthesis opposite the bulky AAF-G, we annealed a 32P-labeled 13mer primer 4 nt before the template AAF-G (Fig. 6A). DNA synthesis assays were then performed with purified yeast Polζ. As shown in Figure 6A (lane 2), some AAF-damaged DNA templates were bypassed by yeast Polζ. However, a significant amount of DNA synthesis stopped right before the lesion (17mer DNA band) (Fig. 6A, lane 2). Major DNA synthesis stop at this position was not observed with the undamaged control template (Fig. 6A, lane 1). Thus, nucleotide incorporation opposite the AAF adduct by yeast Polζ was inhibited by the lesion. Translesion synthesis was also observed by using a 17mer primer annealed right before the template AAF-G (Fig. 6B, lane 1). Apparently, following 1 nt incorporation opposite the lesion, the subsequent extension by yeast Polζ was also significantly inhibited by the AAF adduct, as indicated by the accumulation of the 18mer DNA band (Fig. 6B, lane 1). In a separate experiment, this same AAF-damaged template completely blocked yeast Polα (10), confirming that the DNA substrates used were not contaminated by unmodified templates.
Figure 6.

Translesion synthesis by yeast Polζ opposite an AAF-G. (A) A 13mer primer was labeled with 32P at its 5′ end and annealed 4 nt before the template G without (lane 1) or with (lane 2) an AAF adduct. DNA polymerase assays were performed with 39 ng (193 fmol) of purified yeast Polζ in the presence of all four dNTPs. Nucleotide incorporation opposite the template AAF-G would form an 18mer DNA band. (B) A 17mer primer was labeled with 32P at its 5′ end and annealed right before the template AAF-G as indicated. DNA polymerase assays were performed with 39 ng (193 fmol) of purified yeast Polζ in the presence of all four dNTPs (lane 1), or a single deoxyribonucleoside triphosphate, dATP (lane 2), dCTP (lane 3), dGTP (lane 4) or dTTP (lane 5). DNA size markers in nucleotides are indicated on the left.
To identify the nucleotide incorporated opposite the AAF-guanine, we performed DNA synthesis assays in the presence of only one deoxyribonucleoside triphosphate. As shown in Figure 6B (lanes 2–5), yeast Polζ most frequently incorporated a G opposite the AAF-guanine. These results show that yeast Polζ is capable of error-prone nucleotide incorporation opposite an AAF-G, although the lesion is inhibitory to this polymerase at both the nucleotide incorporation and subsequent extension steps.
Accurate extension of DNA synthesis from opposite the AAF-adducted guanine by yeast Polζ
Recently, we observed that yeast Polη and human Polι are able to insert a C opposite the template AAF-G before aborting DNA synthesis (10,32). It is possible that the aborted DNA synthesis may be continued by another DNA polymerase to achieve bypass of AAF adducts in cells. We hypothesize that Polζ could play such a role as the second DNA polymerase, as we have observed for AP site bypass by the combined activities of yeast Polη and Polζ (10). To test this possibility, we asked whether purified yeast Polζ can extend matched and mismatched DNA primers from opposite the AAF-G. Four 18mer primers were labeled with 32P at their 5′ ends and annealed separately to the AAF-damaged DNA template, generating four DNA substrates containing primer 3′ C, A, T and G, respectively, opposite the AAF-G (Fig. 7A). DNA synthesis assays were then performed with purified yeast Polζ. As shown in Figure 7B (lanes 1, 6, 11 and 16), yeast Polζ extended the primers from opposite the AAF-G with different efficiencies. Surprisingly, extension from the mismatched A:GAAF was most efficient, followed by the matched C:GAAF primer (Fig. 7B, lanes 1, 6, 11 and 16). Extensions from mismatched T:GAAF and G:GAAF were least efficient among the four substrates (Fig. 7B, lanes 1, 6, 11 and 16). To identify the nucleotide incorporated opposite the undamaged template C 5′ to the AAF-G, we performed DNA synthesis assays in the presence of only one deoxyribonucleoside triphosphate. As shown in Figure 7B (lanes 4, 9, 14 and 19), yeast Polζ incorporated the correct G during extension from opposite the AAF-G to the next template base C, regardless of which nucleotide had resided opposite the lesion.
Figure 7.

Primer extension from opposite the AAF-G by yeast Polζ. (A) Four 18mer primers that differed by 1 nt at the 3′ end were labeled with 32P at their 5′ ends and separately annealed to the indicated DNA template, with the primer 3′ end opposite the template AAF-G. (B) DNA polymerase assays were performed with 39 ng (193 fmol) of purified yeast Polζ in the presence of all four dNTPs (N4), or a single deoxyribonucleoside triphosphate, dATP (A), dCTP (C), dGTP (G) or dTTP (T) as indicated. DNA size markers in nucleotides are indicated on the left.
To quantitatively compare the relative extension efficiencies among the four different primers, we measured the kinetic parameters of primer extension by yeast Polζ from opposite the AAF-G to the next template C. Based on the Vmax/Km values (Table 1), primer extension by purified yeast Polζ followed the order, from most efficient to most inefficient, A:GAAF > C:GAAF > T:GAAF > G:GAAF. This conclusion agreed with that obtained from the experiment shown in Figure 7B. These results show that yeast Polζ is capable of accurate extension DNA synthesis from opposite the AAF-G, especially when the primer 3′ end is an A or a C opposite the lesion.
Table 1. Kinetic measurements of primer extension from opposite the GAAF to the next template C by yeast Polζ.
| Primer: template |
Vmax (fmol/min) |
Km (µM) |
Vmax/Km |
fexta |
| A:GAAF | 1.10 ± 0.06 | 2.3 ± 0.6 | 0.480 | 1.3 |
| C:GAAF | 0.77 ± 0.07 | 2.1 ± 1.1 | 0.370 | 1.0 |
| T:GAAF | 0.71 ± 0.07 | 6.0 ± 2.6 | 0.120 | 3.2 × 10–1 |
| G:GAAF | 0.78 ± 0.02 | 36.0 ± 4.3 | 0.022 | 5.9 × 10–2 |
afext = (Vmax/Km)matched primer/(Vmax/Km)mismatched primer. The correct nucleotide G was incorporated opposite the template C 5′ to the lesion. Thus, dGTP was used for the kinetic measurements of extension.
DISCUSSION
Yeast genetic studies revealed that Polζ is required for UV- and AAF-induced mutagenesis in vivo (37,44,45). In this study, we have examined the ability of purified yeast Polζ in translesion synthesis opposite an AAF-G, a TT (6-4) photoproduct and a cis–syn TT dimer. Yeast Polζ is able to perform translesion synthesis opposite the first two DNA lesions in vitro. However, both lesions significantly inhibited the DNA synthesis activity of yeast Polζ (compare Fig. 1C with Figs 2 and 6B, lane 1). Thus, the efficiency of translesion syntheses by yeast Polζ is low opposite both the TT (6-4) photoproduct and the AAF-G.
Recently, several lesion bypass DNA polymerases have been described in eukaryotes. We have tested purified human Polη, Polι and Polκ for in vitro bypass of TT (6-4) photoproduct. Remarkably, none of the polymerases alone is able to incorporate two nucleotides opposite the two modified Ts (5,9,31,32). Thus, yeast Polζ is the only eukaryotic DNA polymerase that can perform translesion synthesis opposite both the modified Ts of the TT (6-4) photoproduct and subsequently extend DNA synthesis further downstream, at least under our in vitro reaction conditions.
Opposite the 3′ T of the TT (6-4) photoproduct, yeast Polζ incorporates either an A or a T with similar efficiencies, and less frequently a G. Opposite the 5′ T of the lesion, yeast Polζ incorporates the correct A much more frequently than other nucleotides. Therefore, the in vitro results predict that the 3′ T is more mutagenic than the 5′ T of the TT (6-4) photoproduct during its bypass by yeast Polζ. When A is incorporated opposite the damaged 3′ T, subsequent extension DNA synthesis by yeast Polζ occurs more efficiently than when T is incorporated opposite the damaged 3′ T. Together, the incorporation and extension results predict that bypass of TT (6-4) photoproduct by yeast Polζ would mainly result in two correct A incorporations opposite the lesion. The most efficient extension DNA synthesis by yeast Polζ occurs when a G is first incorporated opposite the 3′ T of the TT (6-4) photoproduct, which is expected to enhance T→C mutations at the damaged 3′ T. These biochemical properties of Polζ and the resulting predictions are consistent with and can provide explanations for the in vivo results of TT (6-4) photoproduct bypass in yeast cells reported by Nelson et al. (46). After replicating a plasmid containing a site-specific TT (6-4) photoproduct in yeast cells, the majority of bypassed products (69%) contained two correctly inserted As opposite the damaged Ts, and the mutagenic bypass events occurred mainly as G (25%) and T (4%) misinsersions opposite the 3′ T of the lesion (46). Based on the agreement between the in vitro and the in vivo results, we conclude that Polζ directly participates in the bypass of TT (6-4) photoproducts in yeast cells in that this polymerase catalyzes both nucleotide incorporations opposite the lesion and subsequent extension DNA synthesis.
It is possible that in vivo bypass of TT (6-4) photoproduct may be stimulated by other proteins of the Polζ mutagenesis pathway. Nevertheless, the inefficient in vitro bypass by purified yeast Polζ is also consistent with the observation that the efficiency of TT (6-4) photoproduct bypass is only 19% in vivo (46). In vitro, human Polη and Polι are able to preferentially incorporate a G and an A, respectively, opposite the 3′ T of the TT (6-4) photoproduct before aborting DNA synthesis (9,32,35). Since yeast Polζ is capable of extension DNA synthesis from opposite the 3′ T of the TT (6-4) photoproduct, Polζ may additionally function to extend DNA synthesis following Polη or Polι action. Participation by additional DNA polymerases in translesion synthesis opposite the 3′ T of the TT (6-4) photoproduct would enhance the overall efficiency of replication of this lesion in cells compared to the inefficient bypass by Polζ alone. Whether Polη and Polι indeed contribute to error-prone and error-free bypass, respectively, of TT (6-4) photoproduct in cells remains to be tested by genetic experiments.
Unlike a TT (6-4) photoproduct, a template cis–syn TT dimer completely blocked purified yeast Polζ, even when a large excess of the polymerase was used. This is in dramatic contrast to yeast Polη, which efficiently bypasses the TT dimer (4). The 3′ T of the dimer is responsible for blocking yeast Polζ. In contrast, the 5′ T can be effectively bypassed by yeast Polζ from a primer with a 3′ A annealed opposite the 3′ T of the dimer. Our results do not support the conclusion of Nelson et al. (13) that yeast Polζ is able to bypass a template cis–syn TT dimer. It is possible that the different results between our studies and those of Nelson et al. (13) may reflect different yeast Polζ preparations. While only one column chromatography was used for yeast Polζ purification in the studies of Nelson et al. (13), our Polζ was purified by three column chromatography steps. Thus, our yeast Polζ preparation may be more pure.
Our in vitro results suggest that lesion bypass activities of Polζ and Polη are complementary to each other in response to UV radiation. While Polη can bypass TT dimers but not TT (6-4) photoproducts (4–6,9), Polζ can bypass TT (6-4) photoproducts but not TT dimers. Such complementary functions of Polη and Polζ predict that both polymerases are important for translesion synthesis in response to UV radiation. Genetic studies demonstrate that this is indeed the case. Either Polη or Polζ mutant cells are sensitive to UV radiation (37,44,47,48). Furthermore, in vivo studies show that replication of DNA containing cis–syn TT dimers is not affected in the absence of Polζ in yeast cells (46). The inability of Polζ to bypass TT dimers provides a molecular basis for this in vivo observation.
With a low efficiency, yeast Polζ is able to bypass a template AAF-G in an error-prone manner. Our in vitro results agree with in vivo results of Baynton et al. (45) that lesion bypass of AAF-G in yeast cells requires Polζ and occurs with a low efficiency. However, it was observed that 96% of the bypassed products are error-free (45). Error-free bypass of AAF-G in yeast cells could be significantly contributed by Polη, since this polymerase can incorporate a C opposite the AAF-G in vitro (10). If this indeed occurs in vivo, Polζ is likely required to continue the aborted DNA synthesis following C incorporation by Polη opposite AAF-G. Supporting this model, purified yeast Polζ is capable of extension DNA synthesis from a C-terminated primer opposite a template AAF-G. Surprisingly, such extension is most efficient when the primer 3′ end is an A opposite the AAF-G. An implication of this observation is that misincorporated A opposite the AAF-G would be more likely to be fixed into mutations. Consistently, G→T transversion has been reported as a major mutation event at the AAF-G in mammalian systems (49). It should be noted that the human DINB1 protein (Polκ) can effectively incorporate an A opposite the AAF-G (31,33), which may further enhance the AAF-induced G→T transversions in mammalian cells. The yeast S.cerevisiae, however, does not contain a DINB1 homolog.
In eukaryotes, it appears that multiple enzymes are involved in translesion DNA synthesis, including Rev1, Polζ, Polη, Polι and Polκ. While some lesions such as AAF-G may be responded to by multiple polymerases, other lesions such as TT dimer may require a specific polymerase for translesion synthesis. It is perhaps this functional specificity that underscores the importance of evolving multiple DNA polymerases for translesion synthesis. Missing one lesion bypass polymerase may trigger severe health problems in humans, such is the case of XPV, a sunlight-sensitive and skin cancer-prone disease, in which Polη activity is lost (6,7). Even more dramatically, mouse embryonic development cannot be completed without Polζ (50–52).
Based on the biochemical properties of yeast Polζ in response to TT (6-4) photoproduct and AAF-guanine in vitro, we propose a dual-function model for Polζ in bypassing these two DNA lesions in cells. One function of Polζ is to directly catalyze nucleotide incorporation (translesion synthesis) opposite the lesion and subsequently extend DNA synthesis past the lesion. Since nucleotide incorporation by Polζ is inefficient opposite TT (6-4) photoproduct and AAF-G, their more efficient bypasses may require other DNA polymerases to additionally catalyze nucleotide incorporation opposite the lesion. The second function of Polζ is to catalyze the extension DNA synthesis following nucleotide incorporation opposite the lesion by other polymerases, thus serving as the second polymerase and acting at the second step in the two-polymerase two-step lesion bypass mechanism as originally postulated by Yuan et al. (10).
Acknowledgments
ACKNOWLEDGEMENTS
We thank R. Daniel Gietz for the yeast–E.coli shuttle vectors Yeplac112 and Yeplac195. We thank Yanbin Zhang for technical assistance during purification of yeast Polζ. This work was supported by a THRI grant from the Tobacco and Health Research Institute of the University of Kentucky (Z.W.), a New Investigator Award in Toxicology from Burroughs Wellcome Fund (Z.W.) and a NIH grant CA40463 (J.-S.T.). D.K.R. is a recipient of the Kentucky Research Challenge Trust Fund Fellowship and Commonwealth Research Award. D.G. is a recipient of the Kentucky Opportunity Fellowship.
References
- 1.Tang M., Shen,X., Frank,E.G., O’Donnell,M., Woodgate,R. and Goodman,M.F. (1999) UmuD′2C is an error-prone DNA polymerase, Escherichia coli pol V. Proc. Natl Acad. Sci. USA, 96, 8919–8924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Reuven N.B., Arad,G., Maor-Shoshani,A. and Livneh,Z. (1999) The mutagenesis protein UmuC is a DNA polymerase activated by UmuD′, RecA and SSB and is specialized for translesion replication. J. Biol. Chem., 274, 31763–31766. [DOI] [PubMed] [Google Scholar]
- 3.Tang M., Pham,P., Shen,X., Taylor,J.S., O’Donnell,M., Woodgate,R. and Goodman,M.F. (2000) Roles of E. coli DNA polymerases IV and V in lesion-targeted and untargeted SOS mutagenesis. Nature, 404, 1014–1018. [DOI] [PubMed] [Google Scholar]
- 4.Johnson R.E., Prakash,S. and Prakash,L. (1999) Efficient bypass of a thymine–thymine dimer by yeast DNA polymerase, Polη. Science, 283, 1001–1004. [DOI] [PubMed] [Google Scholar]
- 5.Masutani C., Araki,M., Yamada,A., Kusumoto,R., Nogimori,T., Maekawa,T., Iwai,S. and Hanaoka,F. (1999) Xeroderma pigmentosum variant (XP-V) correcting protein from HeLa cells has a thymine dimer bypass DNA polymerase activity. EMBO J., 18, 3491–3501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Masutani C., Kusumoto,R., Yamada,A., Dohmae,N., Yokoi,M., Yuasa,M., Araki,M., Iwai,S., Takio,K. and Hanaoka,F. (1999) The XPV (xeroderma pigmentosum variant) gene encodes human DNA polymerase η. Nature, 399, 700–704. [DOI] [PubMed] [Google Scholar]
- 7.Johnson R.E., Kondratick,C.M., Prakash,S. and Prakash,L. (1999) hRAD30 mutations in the variant form of xeroderma pigmentosum. Science, 285, 263–265. [DOI] [PubMed] [Google Scholar]
- 8.Cleaver J.E. and Kraemer,K.H. (1989) Xeroderma pigmentosum. In Scriver,C.R., Beaudet,A.L., Sly,W.S. and Valle,D. (eds), The Metabolic Basis of Inherited Disease, 6th Edn. McGraw-Hill Book Co., New York, pp. 2949–2971.
- 9.Zhang Y., Yuan,F., Wu,X., Talor,J.-S. and Wang,Z. (2000) Error-prone lesion bypass by human DNA polymerase η. Nucleic Acids Res., 28, 4717–4724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Yuan F., Zhang,Y., Rajpal,D.K., Wu,X., Guo,D., Wang,M., Taylor,J.-S. and Wang,Z. (2000) Specificity of DNA lesion bypass by the yeast DNA polymerase η. J. Biol. Chem., 275, 8233–8239. [DOI] [PubMed] [Google Scholar]
- 11.Matsuda T., Bebenek,K., Masutani,C., Hanaoka,F. and Kunkel,T.A. (2000) Low fidelity DNA synthesis by human DNA polymerase η. Nature, 404, 1011–1013. [DOI] [PubMed] [Google Scholar]
- 12.Johnson R.E., Washington,M.T., Prakash,S. and Prakash,L. (2000) Fidelity of human DNA polymerase η. J. Biol. Chem., 275, 7447–7450. [DOI] [PubMed] [Google Scholar]
- 13.Nelson J.R., Lawrence,C.W. and Hinkle,D.C. (1996) Thymine–thymine dimer bypass by yeast DNA polymerase ζ. Science, 272, 1646–1649. [DOI] [PubMed] [Google Scholar]
- 14.Reynolds P., Weber,S. and Prakash,L. (1985) RAD6 gene of Saccharomyces cerevisiae encodes a protein containing a tract of 13 consecutive aspartates. Proc. Natl Acad. Sci. USA, 82, 168–172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Cassier-Chauvat C. and Fabre,F. (1991) A similar defect in UV-induced mutagenesis conferred by the rad6 and rad18 mutations of Saccharomyces cerevisiae. Mutat. Res., 254, 247–253. [DOI] [PubMed] [Google Scholar]
- 16.Chanet R., Magana-Schwencke,N. and Fabre,F. (1988) Potential DNA-binding domains in the RAD18 gene product of Saccharomyces cerevisiae. Gene, 74, 543–547. [DOI] [PubMed] [Google Scholar]
- 17.Jones J.S., Weber,S. and Prakash,L. (1988) The Saccharomyces cerevisiae RAD18 gene encodes a protein that contains potential zinc finger domains for nucleic acid binding and a putative nucleotide binding sequence. Nucleic Acids Res., 16, 7119–7131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Larimer F.W., Perry,J.R. and Hardigree,A.A. (1989) The REV1 gene of Saccharomyces cerevisiae: isolation, sequence and functional analysis. J. Bacteriol ., 171, 230–237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Jentsch S., McGrath,J.P. and Varshavsky,A. (1987) The yeast DNA repair gene RAD6 encodes a ubiquitin-conjugating enzyme. Nature, 329, 131–134. [DOI] [PubMed] [Google Scholar]
- 20.Bailly V., Lamb,J., Sung,P., Prakash,S. and Prakash,L. (1994) Specific complex formation between yeast RAD6 and RAD18 proteins: a potential mechanism for targeting RAD6 ubiquitin-conjugating activity to DNA damage sites. Genes Dev., 8, 811–820. [DOI] [PubMed] [Google Scholar]
- 21.Nelson J.R., Lawrence,C.W. and Hinkle,D.C. (1996) Deoxycytidyl transferase activity of yeast REV1 protein. Nature, 382, 729–731. [DOI] [PubMed] [Google Scholar]
- 22.Koken M.H.M., Reynolds,P., Jaspers-Dekker,I., Prakash,L., Prakash,S., Bootsma,D. and Hoeijmakers,J.H.J. (1991) Structural and functional conservation of two human homologs of the yeast DNA repair gene RAD6. Proc. Natl Acad. Sci. USA, 88, 8865–8869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Xin H., Lin,W., Sumanasekera,W., Zhang,Y., Wu,X. and Wang,Z. (2000) The human RAD18 gene product interacts with HHR6A and HHR6B. Nucleic Acids Res., 28, 2847–2854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Tateishi S., Sakuraba,Y., Masuyama,S., Inoue,H. and Yamaizumi,M. (2000) Dysfunction of human Rad18 results in defective postreplication repair and hypersensitivity to multiple mutagens. Proc. Natl Acad. Sci. USA, 97, 7927–7932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Lin W., Wu,X. and Wang,Z. (1999) A full-length cDNA of hREV3 is predicted to encode DNA polymerase ζ for damage-induced mutagenesis in humans. Mutat. Res., 433, 89–98. [DOI] [PubMed] [Google Scholar]
- 26.Lin W., Xin,H., Zhang,Y., Wu,X., Yuan,F. and Wang,Z. (1999) The human REV1 gene codes for a DNA template-dependent dCMP transferase. Nucleic Acids Res., 27, 4468–4475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Gibbs P.E., McGregor,W.G., Maher,V.M., Nisson,P. and Lawrence,C.W. (1998) A human homolog of the Saccharomyces cerevisiae REV3 gene, which encodes the catalytic subunit of DNA polymerase ζ. Proc. Natl Acad. Sci. USA, 95, 6876–6880. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Gibbs P.E., Wang,X.D., Li,Z., McManus,T.P., McGregor,W.G., Lawrence,C.W. and Maher,V.M. (2000) The function of the human homolog of Saccharomyces cerevisiae REV1 is required for mutagenesis induced by UV light. Proc. Natl Acad. Sci. USA, 97, 4186–4191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Murakumo Y., Roth,T., Ishii,H., Rasio,D., Numata,S., Croce,C.M. and Fishel,R. (2000) A human REV7 homolog that interacts with the polymerase ζ catalytic subunit hREV3 and the spindle assembly checkpoint protein hMAD2. J. Biol. Chem., 275, 4391–4397. [DOI] [PubMed] [Google Scholar]
- 30.Zhang Y., Yuan,F., Wu,X. and Wang,Z. (2000) Preferential incorporation of G opposite template T by the low fidelity human DNA polymerase ι. Mol. Cell. Biol., 20, 7099–7108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Zhang Y., Yuan,F., Wu,X., Wang,M., Rechkoblit,O., Taylor,J.-S., Geacintov,N.E. and Wang,Z. (2000) Error-free and error-prone lesion bypass by human DNA polymerase κin vitro. Nucleic Acids Res., 28, 4138–4146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Zhang Y., Yuan,F., Wu,X., Taylor,J.-S. and Wang,Z. (2001) Response of human DNA polymerase ι to DNA lesions. Nucleic Acids Res., 29, 928–935. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Ohashi E., Ogi,T., Kusumoto,R., Iwai,S., Masutani,C., Hanaoka,F. and Ohmori,H. (2000) Error-prone bypass of certain DNA lesions by the human DNA polymerase κ. Genes Dev., 14, 1589–1594. [PMC free article] [PubMed] [Google Scholar]
- 34.Johnson R.E., Washington,M.T., Haracska,L., Prakash,S. and Prakash,L. (2000) Eukaryotic polymerases ι and ζ act sequentially to bypass DNA lesions. Nature, 406, 1015–1019. [DOI] [PubMed] [Google Scholar]
- 35.Tissier A., Frank,E.G., McDonald,J.P., Iwai,S., Hanaoka,F. and Woodgate,R. (2000) Misinsertion and bypass of thymine–thymine dimers by human DNA polymerase ι. EMBO J., 19, 5259–5266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Gerlach V.L., Feaver,W.J., Fischhaber,P.L. and Friedberg,E.C. (2001) Purification and characterization of polκ, a DNA polymerase encoded by the human DINB1 gene. J. Biol. Chem., 276, 92–98. [DOI] [PubMed] [Google Scholar]
- 37.Morrison A., Christensen,R.B., Alley,J., Beck,A.K., Bernstine,E.G., Lemontt,J.F. and Lawrence,C.W. (1989) REV3, a Saccharomyces cerevisiae gene whose function is required for induced mutagenesis, is predicted to encode a non-essential DNA polymerase. J. Bacteriol ., 171, 5659–5667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Smith C.A. and Taylor,J.S. (1993) Preparation and characterization of a set of deoxyoligonucleotide 49-mers containing site-specific cis-syn, trans-syn-I, (6-4) and Dewar photoproducts of thymidylyl(3′→5′)-thymidine. J. Biol. Chem., 268, 11143–11151. [PubMed] [Google Scholar]
- 39.Gietz R.D. and Sugino,A. (1988) New yeast–Escherichia coli shuttle vectors constructed with in vitro mutagenized yeast genes lacking six-base pair restriction sites. Gene, 74, 527–534. [DOI] [PubMed] [Google Scholar]
- 40.Washington M.T., Johnson,R.E., Prakash,S. and Prakash,L. (1999) Fidelity and processivity of Saccharomyces cerevisiae DNA polymerase η. J. Biol. Chem., 274, 36835–36838. [DOI] [PubMed] [Google Scholar]
- 41.Creighton S., Bloom,L.B. and Goodman,M.F. (1995) Gel fidelity assay measuring nucleotide misinsertion, exonucleolytic proofreading, and lesion bypass efficiencies. Methods Enzymol., 262, 232–256. [DOI] [PubMed] [Google Scholar]
- 42.Friedberg E.C., Walker,G.C. and Siede,W. (1995) DNA Repair and Mutagenesis. American Society of Microbiology Press, Washington, D.C.
- 43.Belguise-Valladier P. and Fuchs,R.P. (1995) N-2-aminofluorene and N-2 acetylaminofluorene adducts: the local sequence context of an adduct and its chemical structure determine its replication properties. J. Mol. Biol., 249, 903–913. [DOI] [PubMed] [Google Scholar]
- 44.Lawrence C.W., Das,G. and Christensen,R.B. (1985) REV7, a new gene concerned with UV mutagenesis in yeast. Mol. Gen. Genet., 200, 80–85. [DOI] [PubMed] [Google Scholar]
- 45.Baynton K., Bresson-Roy,A. and Fuchs,R.P. (1998) Analysis of damage tolerance pathways in Saccharomyces cerevisiae: a requirement for Rev3 DNA polymerase in translesion synthesis. Mol. Cell. Biol., 18, 960–966. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Nelson J.R., Gibbs,P.E., Nowicka,A.M., Hinkle,D.C. and Lawrence,C.W. (2000) Evidence for a second function for Saccharomyces cerevisiae Rev1p. Mol. Microbiol., 37, 549–554. [DOI] [PubMed] [Google Scholar]
- 47.McDonald J.P., Levine,A.S. and Woodgate,R. (1997) The Saccharomyces cerevisiae RAD30 gene, a homologue of Escherichia coli dinB and umuC, is DNA damage inducible and functions in a novel error-free postreplication repair mechanism. Genetics, 147, 1557–1568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Roush A.A., Suarez,M., Friedberg,E.C., Radman,M. and Siede,W. (1998) Deletion of the Saccharomyces cerevisiae gene RAD30 encoding an Escherichia coli DinB homolog confers UV radiation sensitivity and altered mutability. Mol. Gen. Genet., 257, 686–692. [DOI] [PubMed] [Google Scholar]
- 49.Shibutani S., Suzuki,N. and Grollman,A.P. (1998) Mutagenic specificity of (acetylamino)fluorene-derived DNA adducts in mammalian cells. Biochemistry, 37, 12034–12041. [DOI] [PubMed] [Google Scholar]
- 50.Wittschieben J., Shivji,M.K., Lalani,E., Jacobs,M.A., Marini,F., Gearhart,P.J., Rosewell,I., Stamp,G. and Wood,R.D. (2000) Disruption of the developmentally regulated rev3l gene causes embryonic lethality. Curr. Biol., 10, 1217–1220. [DOI] [PubMed] [Google Scholar]
- 51.Esposito G., Godindagger,I., Klein,U., Yaspo,M., Cumano,A. and Rajewsky,K. (2000) Disruption of the Rev3l-encoded catalytic subunit of polymerase ζ in mice results in early embryonic lethality. Curr. Biol., 10, 1221–1224. [DOI] [PubMed] [Google Scholar]
- 52.Bemark M., Khamlichi,A.A., Davies,S.L. and Neuberger,M.S. (2000) Disruption of mouse polymerase ζ (Rev3) leads to embryonic lethality and impairs blastocyst development in vitro. Curr. Biol., 10, 1213–1216. [DOI] [PubMed] [Google Scholar]