


Abstract
Expression of functional TRA-2 protein in the male germline of Drosophila is regulated through a negative feedback mechanism in which a specific TRA-2 isoform represses splicing of the M1 intron in the TRA-2 pre-mRNA. We have previously shown that the mechanism of M1 splicing repression is conserved between distantly related Drosophila species. Using transgenic fly strains, we have examined the effects on regulation of mutations in two conserved features of the M1 intron. Our results show that TRA-2-dependent repression of M1 splicing depends on the presence of a suboptimal non-consensus 3′ splice site. Substitution of this 3′ splice site with a strong splice site resulted in TRA-2 independent splicing, while substitution with an unrelated weak 3′ splice site was compatible with repression, implying that reduced basal splicing efficiency is important for regulation. A second conserved element internal to the intron was found to be essential for efficient M1 splicing in the soma where the intron is not normally retained. We show that the role of this element is to enhance splicing and overcome the reduction in efficiency caused by the intron’s suboptimal 3′ splice site. Our results indicate that antagonistic elements in the M1 intron act together to establish a context that is permissive for repression of splicing by TRA-2 while allowing efficient splicing in the absence of a repressor.
INTRODUCTION
A number of RNA binding proteins have now been identified that regulate the cell-specific splicing of specific pre-mRNAs (1,2). Several of these factors also autoregulate their own expression by directing alternative splicing of the pre-mRNAs that encode them (3–7). For example, the splicing regulator TRA-2 controls the splicing of pre-mRNAs of genes such as doublesex (dsx) and fruitless (fru) (8–14) as well as that from the tra-2 gene itself (4). Autoregulation of splicing in this case serves to limit the expression of a specific isoform of TRA-2 (TRA-2226) that is required in male germ cells for normal spermatogenesis (15). Negative feedback has an important biological role because expression of excessive levels of TRA-2226 in the germline results in formation of abnormal spermatids and sterility in males (16).
The focal point of tra-2 autoregulation is the M1 intron which splits the initiation codon of the TRA-2226 open reading frame. M1 splicing is repressed by the TRA-2226 protein resulting in the formation of an alternative mRNA encoding TRA-2179, a protein isoform initiating translation at a downstream start codon (15,17,18). Unlike TRA-2226, expression of TRA-2179 is neither sufficient nor required in the germline for spermatogenesis. In flies homozygous for point mutations blocking expression of functional TRA-2226, the M1 intron is efficiently spliced and M1-containing RNAs are not detected in the germline. In contrast, in wild-type males expressing this protein the M1 intron is retained in >50% of mature germline tra-2 mRNA. Thus repression of M1 splicing serves to limit the amount of TRA-2226-encoding mRNA that is produced (16).
In addition to germ cells, TRA-2 mRNAs are produced in a wide variety of somatic tissues in both sexes. Although a low level of M1 retention is observed in the soma, accumulation of somatic M1-containing RNA is not dependent on functional TRA-2 protein (4). The basis of the germline specificity of TRA-2 autoregulation is unknown, but it is unlikely that TRA2226 is a limiting factor as significant levels of mRNAs encoding this isoform have been shown to be present both in the soma and germline.
The mechanism responsible for TRA-2-dependent repression of M1 splicing in male germ cells is likely to differ significantly from the well characterized mechanism by which TRA-2 activates female-specific splicing of dsx RNA (19–25). The latter activity occurs only in the soma rather than the germline, requires a cofactor (TRA) that is not involved in repression of M1 splicing and can be functionally separated from the repression of M1 splicing by specific mutations in the tra-2 gene (26). Moreover, in dsx splicing, TRA-2226 and other TRA-2 isoforms associate with the dsx pre-mRNA and enhance splice site recognition by recruiting general splicing factors, but in the case of the M1 intron, TRA-2 protein acts to repress splicing.
We have taken an evolutionary approach to identify cis-elements affecting M1 splicing. In a previous study we showed that the negative feedback strategy used by the Drosophila melanogaster tra-2 gene is conserved in Drosophila virilis, a species that diverged >60 million years ago (27). Although the intron sequences of these two species are highly divergent, two short conserved sequence features were identified within M1. In this study we have investigated the possible regulatory roles of these sequences. Our results show that these sequences have opposing activities that are necessary for the correct tissue-specific repression of M1 splicing.
MATERIALS AND METHODS
Construction of transgenes
P[tra2/ftz], in which the ftz intron is substituted for the M1 intron of tra-2, was introduced into the Drosophila genome using the plasmid pCSPtra2/ftzIVS. To generate this plasmid, primer-directed mutagenesis of sequences flanking both introns was performed to introduce ClaI sites. After ClaI digestion, the complete ftz intron was then ligated in place of the M1 intron in a 1.4 kb Apa-H3 genomic fragment of the tra-2 gene contained in the plasmid pSK Apa-H3 DACla to produce the plasmid pSK Apa-H3-tra2/ftz. A fragment containing the substitution and flanking regions was then inserted into a 3.8 kb genomic tra-2 fragment in the plasmid pTZ3.9BΔSac. The entire insert fragment was then transferred to the P element transformation vector pCaSpeR4 (28) to generate pCSPtra2/ftzIVS. Ligation junctions and mutations were confirmed by DNA sequencing. The 5′ splice site of the hybrid gene is ATCGATG/gtaggcat and at the 3′ splice site is cuuacag/ATCGAT where the lower case bases represent ftz intron sequences and underlined bases encode the ClaI restriction site.
Other transgenes were produced by primer-directed mutagenesis of the plasmid P[tra2+1] and then insertion of the altered fragments into pCaSpeR (16). All P[tra2+1] derived constructs contain a 7 nt insertion in exon 4 that produces a translation frameshift, blocking production of functional TRA-2 protein. The 7 nt insertion also makes it possible to distinguish transcripts derived from the wild-type and transgene RNAs in RT–PCR experiments. Sequences of oligonucleotides used in primer-directed mutagenesis are as follows: tra2-VCsub, 5′-TAAGAACCCACCACCTGGATATCACCTATTCTACGTCCA-3′; tra2-VCdel, 5′-TGGACGTAGAATTCGGTTCTTACA-3′; tra2-MHCE3′, 5′-GTCGACCTTGTTTGCAAGGGGATAAGTTCAAATTATCTTGCG-3′; tra2-CAG3′, 5′-GTCAGACCTGGATTCGATG-3′ and tra2-ftz3′, 5′-GTAGTCAGACCTGTAAGCATAAGCAAAAGAAAAAAAATAAGATTATGTTG-3′.
All constructs were injected directly into Drosophila embryos of the genotype w1118/BsY; tra-2B/CyO in combination with the plasmid pΔ2-3, which provided transposase. Individuals bearing stable P insertions were identified in the G1 generation by changes in eye pigmentation due to the mini-w+ gene contained in pCaSpeR.
Preparation of D.melanogaster RNA
In the range of 50–100 flies were homogenized using a Tekmar Tissuemizer in 2 ml phenol combined with 2 ml 2× NETS (200 mM NaCl, 20 mM EDTA, 20 mM Tris pH 8.0, 0.2% SDS). Phases were separated by centrifugation and the aqueous phase was collected. After 2–3 additional phenol extractions, the RNA was precipitated with 2 vol ethanol. The RNA was resuspended in RNase-free water and quantified by absorbance. Following treatment with DNase at 37°C for 1 h samples were heat-inactivated for 10 min at 70°C, phenol extracted and precipitated with 2 vol ethanol. Poly(A) RNA was isolated by binding to oligo(dT)-cellulose.
Detection of alternatively spliced RNAs
In most experiments the dependence of M1 splicing on tra-2 was determined by RT–PCR performed in parallel on RNA from adult flies with a functional tra-2+ allele (tra-2B/CyO) and homozygous tra-2 mutants (tra-2B/tra-2B). First strand cDNA synthesis was performed on poly(A) RNA or total RNA using the SuperScript Preamplification system (Gibco BRL). Aliquots of 0.8 µg poly(A) RNA or 5 µg total RNA with 0.5 µg Oligo(dT)12–18 (New England Biolabs) were denatured at 70°C for 10 min in a final volume of 12 µl and incubated on ice for 1 min. The subsequent annealing, cDNA synthesis, reaction termination, and removal of RNA steps were performed according to manufacturers instructions. An aliquot of 7 µl first strand cDNA was amplified directly using Taq polymerase and the following primers: X3S, 5′-CTCAGCCGATTCAGCTGGTGCTCTTG-3′; X5/X6, 5′-CGCTGTGTTTGTGCGTCAATCA-3′ and X2S, 5′-GACAACTTCATCCGGACATAAG-3′.
PCR reactions were carried out in a final volume of 100 µl with 10 mM Tris–HCl pH 8.3, 50 mM potassium chloride, 1.5 mM magnesium chloride, by cycling 20 times at 94°C for 1 min, followed by 60°C for 30 s for primer pair X3S and X5/X6 or 55°C for primer pair X2S and X5/X6, and 72°C for 30 s. Cycling at these temperatures was followed by a single final incubation at 72°C for 5 min. In all experiments the primers were positioned so that PCR products derived from M1-containing RNAs could be readily distinguished from products derived from contaminating genomic DNA based on gel mobility. The above PCR conditions produced ratios of endogenous spliced and unspliced M1 intron consistent with those determined in nuclease protection experiments and were therefore judged to be representative of the relative ratios of transgenic products as well.
PCR products were separated on a 1.2% agarose gel and blotted overnight in 1.5 M sodium chloride, 0.1 M sodium citrate. The tra-2 transcripts were detected after blotting with a 32P-end-labeled oligonucleotide specific for either endogenous tra-2 (Nae-WT, 5′-TGGTGCCGGCGACTG-3′) or transgene-derived RNAs (Nae+1, 5′-GACCGCTGGTGCCGATCCCCG-3′). These oligonucleotides are complementary to sequences at the position of the 7 nt insertion placed in exon 4. Blots were hybridized with the Nae-WT probe at 42°C in 1 M sodium chloride, 0.1 M PIPES pH 7.0, 0.2% BSA, 0.2% Ficoll, 0.2% polyvinylpyrrolidone, 10% dextran sulfate for 12–16 h and washed at 58°C with 6× SSC, 0.1% SDS. In experiments with the Nae +1 probe, incubations with probe were carried out at 60°C, for 12–16 h and washed at 68°C in 6× SSC, 0.1% SDS. Northern blots were performed as previously described (27) using 2 µg adult poly(A) RNA.
RESULTS
Sequences in the M1 intron are required for splicing repression
To determine if sequences from the M1 intron are necessary for splicing repression we substituted the entire M1 intron with the 150 nt intron from the Drosophila ftz gene which is not subject to tra-2-dependent regulation in its normal context. A restriction fragment containing the entire tra-2/ftz hybrid gene as well as the tra-2 promoter and its other flanking sequences was then introduced into the P element transformation vector pCaSper4 to produce the plasmid pCSPtra-2/ftzIVS (Fig. 1A). In these and all subsequent constructs we included a 7 nt insertion in exon 4 that served both to block translation of TRA-2226 protein from the transgene and as a means of distinguishing the transgene-derived RNAs from those of the endogenous tra-2 gene (Fig. 1A). Transcripts from the transgene were specifically detected by RT–PCR using an oligonucleotide (Nae+1) that includes sequences homologous to the 7 nt insertions in exon 4.
Figure 1.
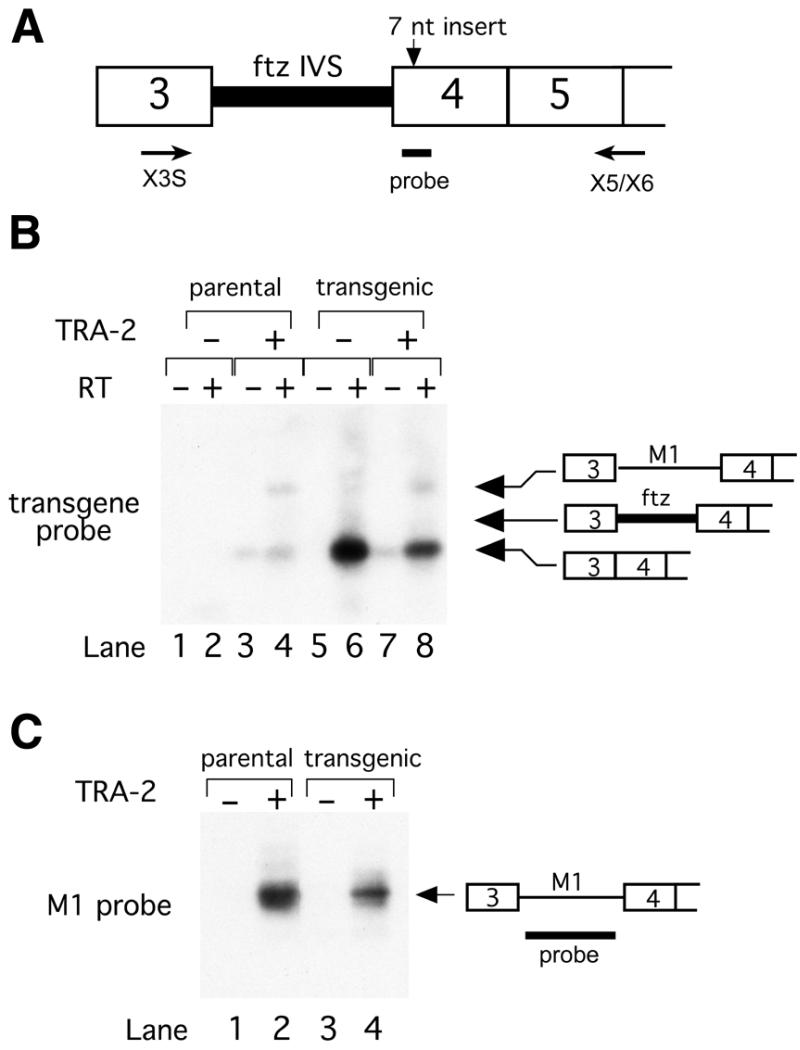
Replacement of the tra-2 M1 intron with the intron from the ftz gene. (A) A schematic showing the predicted 5′ end of male germline tra-2 pre-mRNA in which M1 intron of tra-2 has been replaced with the constituitively spliced intron of the ftz gene (ftz IVS). Also shown is the relative location of the 7 nt insertion downstream of the 3′ splice site, the transgene probe and the positions of the PCR primers (X3S and X5/X6). Male germline specific transcripts initiate within exon 3 of the tra-2 gene. (B) A Southern blot of low-cycle RT–PCR products from male tra-2/ftzIVS transgenic flies hybridized with a 32P-end-labeled transgene-specific oligonucleotide containing the 7 nt insertion is shown (lanes 2, 4, 6, 8). Positions of products corresponding to different spliced forms are indicated on the right. Products from PCR with material for mock reverse transcriptase reactions (RT) in which enzyme was omitted are also shown (lanes 1, 3, 5, 7). Samples denoted ‘–’ for TRA-2 are derived from males homozygous for a strong loss of function allele (tra-2B) that contains a nonsense mutation located within the TRA-2 RNA binding domain. Samples denoted ‘+’ for TRA-2 are derived from sibling heterozygous mutant males (tra-2B/+). The lanes marked ‘parental’ derive from the strain used for microinjection (w1118/BsY; tra-2B/CyO) which contains no transgene. (C) RT–PCR products from the same RNA samples hybridized with a probe from the M1 intron to confirm that endogenous transcripts undergo tra-2-dependent M1 splicing repression.
Fly strains containing random insertions of the P[tra-2/ftz] transgene were produced by microinjection of tra-2B/CyO embryos. The tra-2B allele is a nonsense mutation in the region encoding the TRA-2 RNA binding domain that eliminates all autoregulatory activity but produces normal amounts of mutant tra-2 mRNA. Both homozygous tra-2B mutants and normal heterozygous tra-2B/CyO individuals were sorted from transgenic strains and used to isolate RNA.
Transcripts produced from the P[tra-2/ftz] transgene were efficiently spliced and no ftz intron-containing RNA was detected in either tra-2B/CyO males (Fig. 1B, lanes 7 and 8) or their tra-2B homozygous mutant siblings (Fig. 1B, lanes 5 and 6). Control experiments confirmed that the M1 intron containing RNA from the endogenous tra-2 gene accumulated normally (Fig. 1C). M1-containing tra-2 transcripts were readily detected in transgenic males with a wild-type endogenous tra-2 allele (Fig. 1C, lane 4) but not in flies from tra-2B homozygous males (Fig. 1C, lane 3). Together these results show that tra-2/ftzIVS transcripts are not subject to splicing repression by products from the endogenous tra-2 gene. We conclude from this that sequences within the M1 intron are necessary for splicing repression.
A suboptimal 3′ splice site is essential for repression of M1 splicing in vivo
Examination of the sequence of the M1 3′ splice site region revealed that it differs significantly in several respects from the Drosophila 3′ splice site consensus. An adenosine is present at the –3 position rather than the consensus pyrimidine that is found in ∼96% of all Drosophila introns (29). In addition, the 3′ splice site was not preceded by the usual long polypyrimidine tract, but instead only by several short runs of pyrimidines each <5 nt in length (Fig. 2A). Finally, there was no good match to the Drosophila consensus branchpoint in the distal region of the intron (not shown). We previously found that the 3′ splice site region of the M1 intron of the D.virilis tra-2 gene shares these characteristics and that this intron is also repressed by TRA-2 (27). This led us to hypothesize that these atypical sequences are suboptimally recognized by the general splicing machinery and that poor recognition of the 3′ splice site/polypyrimidine tract facilitates TRA-2-dependent repression of M1 splicing. To test this idea, we produced transgenes with two mutations, in which sequences from the 3′ splice site region of M1 were replaced with sequences more closely matching the Drosophila consensus (29). In P[ftz 3′] the M1 3′ splice site and polypyrimidine tract were replaced with the analagous sequences from the ftz intron (Fig. 2A). In the other construct, P[CAG 3′], the non-consensus A residue at position –3 was changed to C as in the Drosophila consensus. Transgenic fly strains were generated using both of these mutations and assayed for the ability of transgene-derived RNA to undergo M1 repression in the presence and absence of functional endogenous TRA-2. Splicing of the ‘improved’ M1 intron in transcripts from P[ftz 3′] was not repressed in the presence of TRA-2 protein (Fig. 2B, lanes 5–8). This indicates that splicing of an intron containing a consensus 3′ splice site cannot be repressed by TRA-2. In contrast, transcripts from P[tra-2/CAG] were repressed by endogenous TRA-2 protein (Fig. 2B, lanes 9–12) to a similar extent to endogenous tra-2 RNAs (Fig. 2B, lanes 1–4). This shows that a non-consensus nucleotide at the –3 position is not essential for tra-2-dependent repression of M1 splicing.
Figure 2.
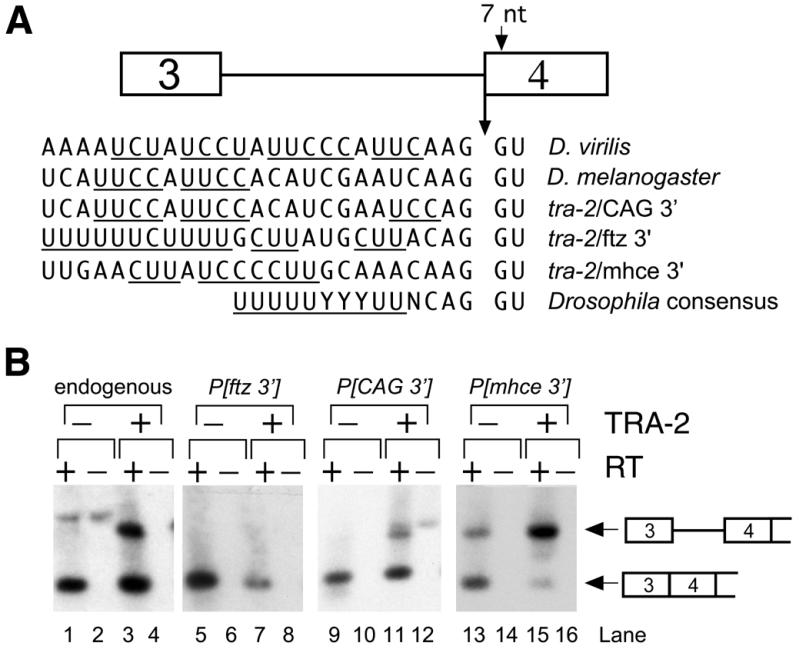
Substitutions of sequences near the M1 3′ splice site. (A) Schematic showing the M1 intron (line) and sequences of relevant 3′ splice sites. The 3′ intron sequences present in the ftz3′, CAG3′ and mhce 3′ transgenes are shown. Runs of three or more consecutive pyrimidines are underlined. (B) Southern blots of low-cycle RT–PCR products of RNAs from males of various fly strains. The primers used and the labeling of lanes are as described in Figure 1. Products in lanes 1–4 are derived from RNA of tra-2B/tra-2B and tra-2B/+ of the non-transgenic parental strain and were hybridized with an oligonucleotide probe specific for the endogenous tra-2 gene lacking the 7 nt insertion in exon 4. Products in lanes 5–16 are derived from the various transgenic strains indicated and were hybridized with a transgene-specific oligonucleotide. Expected mobilities of products from spliced and unspliced RNA are indicated by arrows on the right. The faint lower mobility band appearing in lanes 1, 2, 11 and 12 is not dependent on reverse transcriptase and corresponds in size to the products expected from contaminating genomic DNA.
A possible explanation for the above findings is that non-consensus sequences near the 3′ splice site are necessary to lower the basal rate of M1 splicing to a level that permits repression by TRA-2226. A second possibility is that the substituted sequences are directly and specifically recognized by TRA-2 or another regulatory factor involved in repression. To distinguish these possibilities we used sequences from the Drosophila myosin heavy chain intron E (mhce) (30), which also has a non-consensus 3′ splice site, to substitute for those of the M1 3′ splice site/polypyrimidine tract region. The mhce 3′ splice site sequences differ from those of the native M1 intron (Fig. 2A) and thus, would not be expected to bind similar regulatory factors. Transcripts produced from a transgene with this substitution P[mhce 3′] are shown in Figure 2B (lanes 13 and 15). Unlike endogenous tra-2 transcripts, a large fraction of transgene-derived RNAs retained the M1 intron in the absence of functional TRA-2 protein (lane 13). This suggests that recognition of the 3′ splice site in P[mhce 3′] by the basal splicing machinery is less efficient than splicing of transcripts from the endogenous tra-2 gene. Significantly, in the presence of a functional endogenous TRA-2 protein, splicing of the M1 intron from P[mhce 3′] was further reduced (lane 15). These results indicate that substitution of mhce sequences near the 3′ end of the intron are compatible with TRA-2-dependent splicing repression. Because repression occurs with either the mhce or the native M1 sequences, we conclude that a critical feature of this region necessary for splicing repression is a suboptimal (but functional) polypyrimidine tract.
Mutation of a conserved sequence internal to the M1 intron affects splicing in somatic tissues
Comparison of the D.virilis and D.melanogaster tra-2 genes also revealed a sequence element (VC) internal to the M1 intron of both genes that is conserved at 15 out of 18 positions (27). To determine if the VC element plays an important role in M1 repression we generated two transgenes with different mutations disrupting the element in the D.melanogaster M1 intron (Fig. 3A). In the transgene P[VC-sub] we replaced the VC element with random nucleotides at 14 out of 18 positions. In the transgene P[VCdel] we deleted 14 nt from the element. Both transgenes produced M1-containing RNAs in a TRA-2 independent manner, suggesting that the VC element is necessary for efficient splicing of the intron (Fig. 3B). Northern blot analysis of M1-containing RNAs from a transgenic strain carrying P[VC-sub] revealed the appearance of a novel M1-containing RNA that is independent of the presence of functional TRA-2 protein and is larger than the normal male germline M1-containing tra-2 transcript (Fig. 3C). The size of the novel RNA suggests that it might be derived from the soma where an upstream transcription start site is utilized (Fig. 4A) and where the M1 intron is normally not retained at significant levels (31). RNAase protection assays confirmed that these flies expressed novel transcripts with the somatic form of exon 3 and retained the entire M1 intron (data not shown). To further determine that these novel transcripts accumulate in the soma, we utilized primers from exons 2 and 4 in RT–PCR experiments with RNA extracted from gonad-free carcasses of adult male flies. M1-containing sequences made up a large fraction of the products deriving from both the P[VC-sub] and P[VC-del] transgenes but were only a minor fraction of products from the endogenous tra-2 gene (Fig. 4B). As expected based on previous observations (4,31), neither endogenous nor transgenic M1 containing RNAs found in the soma were dependent on the activity of the TRA-2 protein. We conclude that somatic splicing of the M1 intron is reduced in the absence of the VC element.
Figure 3.
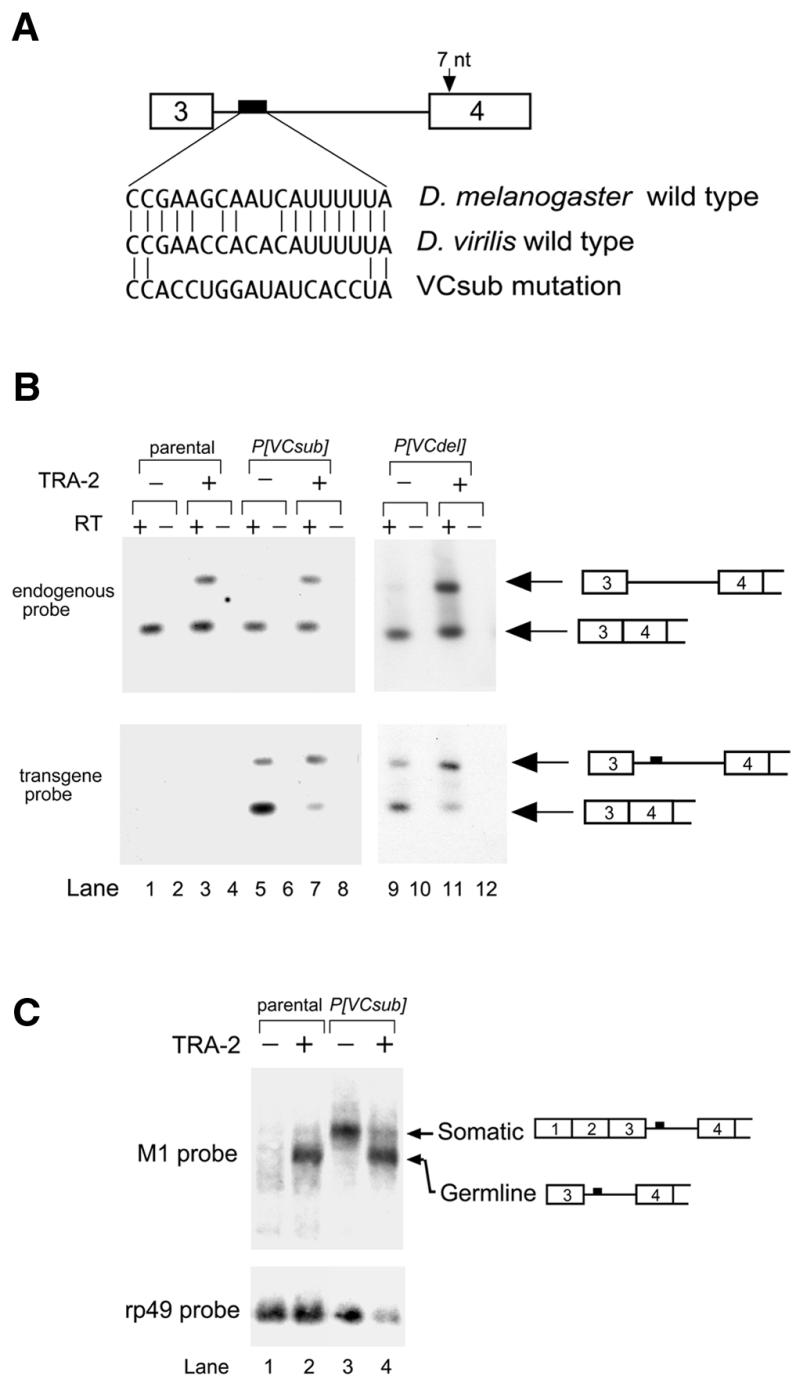
Mutation of a conserved intron sequence decreases M1 splicing in the soma. (A) Schematic showing the sequence and location of the conserved VC element (black box) in the M1 intron (line). The sequence of the VC element and the substitution mutation (VCsub) is shown. (B) Low cycle RT–PCR was performed as in Figure 1B. RT–PCR products were hybridized with oligonucleotides specific for either endogenous or transgene-derived RNA. Expected mobilities and the structure of spliced and unspliced products are shown to the right of the panels. After hybridization with the probe for endogenous transcripts the same blot was stripped and used to detect products from the transgene. Endogenous M1 containing RNA is observed only in wild-type males (upper panel, lanes 7 and 11), but M1-containing RNA derived from the transgenes is observed in both wild-type (lower panel, lanes 7 and 11) and tra-2B mutant males (lower panel, lanes 5 and 9). (C) Northern blot analysis of poly(A)+ RNA from flies expressing the VCsub mutation. M1 intron containing transcripts were detected using a 32P-labeled in vitro transcribed antisense M1 RNA. The smaller, TRA-2-dependent RNA corresponds to the normal germline M1 containing RNA and is observed in both the parental (lanes 1 and 2) and transgenic flies (lanes 3 and 4). The larger RNA which is independent of TRA-2 function, is observed at high levels in transgenic flies and at much lower levels in the parental strain. This RNA corresponds in size to transcripts initiating at the somatic transcription initiation site as indicated on the right of the figure. As a loading control the blot was rehybridized with a probe for transcripts from the ribosomal protein 49 (rp49) gene. Note that lane 4 is underloaded.
Figure 4.
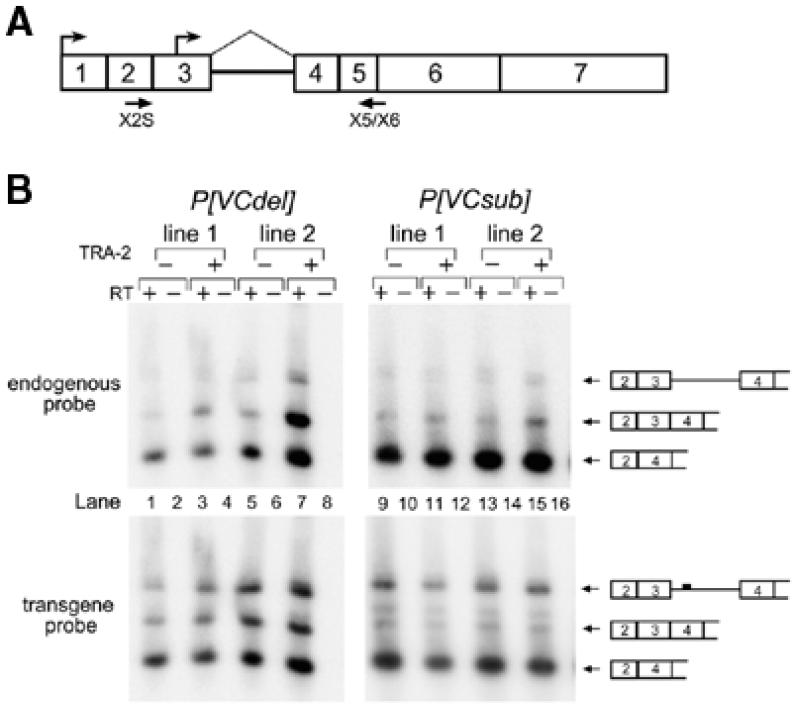
The VC element is necessary for the efficient removal of the M1 intron in the soma. (A) Schematic diagram of an M1-containing somatic tra-2 RNA is shown. Boxes indicate exons and arrows above indicate both the somatic transcription start site at the beginning of exon 1 and the germline transcription start site within exon 3. PCR primers designed to specifically recognize cDNA generated from the longer somatic tra-2 RNAs are indicated below the RNA. Primer X2S is upstream of the germline-specific transcription start site and primer X5/X6 spans the splice junction between exons 5 and 6. (B) Low cycle RT–PCR analysis of somatic RNA extracted from male fly carcasses from which the testes had been removed by dissection are shown. The flies examined expressed RNA from either the P[VCdel] or the P[VCsub] tra-2 transgenes. Endogenous and transgene RT–PCR products were distinguished with specific oligonucleotide probes as in previous experiments. Note that in all samples the band corresponding to somatic M1-containing RNA comprises a significantly higher fraction of the transgene transcripts compared to endogenous gene transcripts. As observed in previous studies, somatic M1 splicing is independent of the endogenous tra-2 genotype.
The VC element is necessary to overcome suboptimal splicing signals near the 3′ end of the M1 intron
The low levels of M1-containing transcripts present in the soma suggest that the intron is normally efficiently spliced in these tissues despite the presence of suboptimal 3′ signals. This led us to hypothesize that a function of the VC element is to overcome these suboptimal splicing signals. If so, then an intron with consensus 3′ splicing signals would not be expected to require the VC element in the soma for efficient splicing. We therefore produced transgenic flies that expressed transcripts carrying a combination of mutations in both VC and the 3′ splice site region P[tra-2/VCsub, ftz 3′]. This transgene contains a combination of the substitutions used in P[VC-sub] and P[tra-2/ftz 3′]. As predicted by our hypothesis, the M1 intron was efficiently spliced from transcripts containing this transgene despite the absence of a functional VC element (Fig. 5). These results indicate that the VC element is required for M1 splicing only in the presence of the suboptimal, native 3′ splicing signals.
Figure 5.
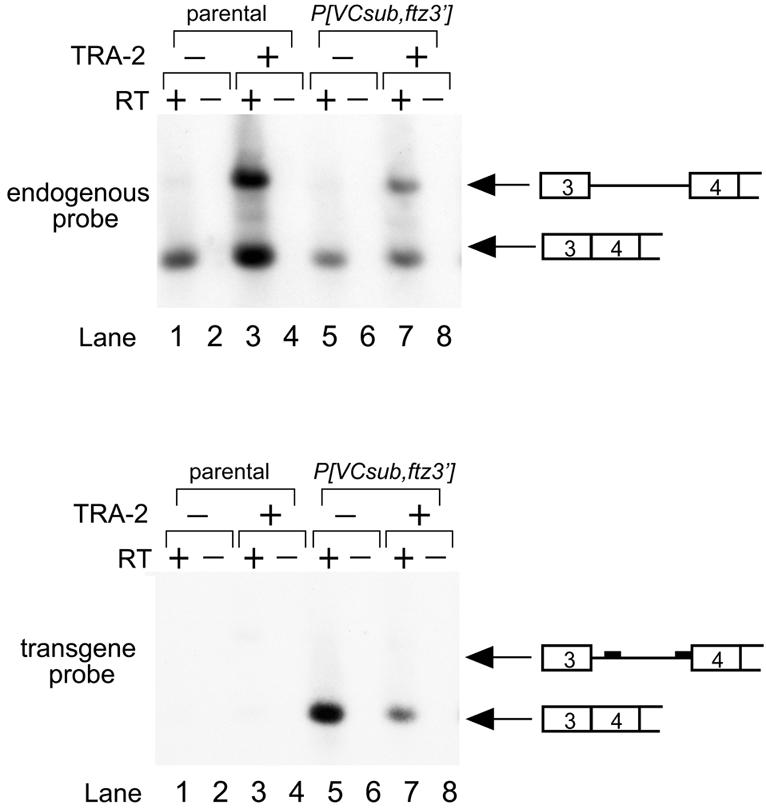
Substitution of a strong polypyrimidine tract/3′ splice site into the M1 intron alleviates the somatic requirement for the VC element. M1 splicing in flies expressing P[VCsub,ftz3′] transgene is analyzed by low cycle RT–PCR and Southern blotting. In this transgene the VCsub mutation is combined with a substitution of the strong ftz 3′ splice site. Products from endogenous and transgene transcripts were detected using specific oligonucleotide probes as in other experiments. Expected mobilities of spliced and unspliced products are shown schematically on the right of the panels. Intron-containing transcripts were detected from the endogenous gene (upper panel, lanes 3 and 7) but not from the transgene (lanes 5 and 7) in the presence or absence of functional TRA-2 protein.
DISCUSSION
The alternative splicing of tra-2 appears to be conserved during evolution. TRA-2 protein variants that are generated by alternative splicing in humans are strikingly similar to those produced in the fly (26,32). Although conservation of the regulatory mechanisms between human and insects appears unlikely, examination of the splicing of tra-2 RNA from D.virilis, a species that diverged from D.melanogaster over 60 million years ago, revealed that it undergoes similar regulation in these two species (27). Testes-specific splice variants in D.virilis retain an intron analogous to M1 that interrupts the first translation initiation codon. When introduced into D.melanogaster, the D.virilis tra-2 gene acted in trans to regulate splicing of the endogenous tra-2 pre-mRNA. Using a defective D.virilis transgene it was found that the endogenous D.melanogaster TRA-2 protein could likewise repress splicing of D.virilis tra-2 pre-mRNA. Thus, the cis-regulatory sequences affecting tra-2 splicing are likely to be very similar in the two species.
Comparison of the D.virilis and D.melanogaster tra-2 pre-mRNA sequences revealed conserved features within the M1 intron. In both species the 3′ splice site of the M1 intron is similarly divergent from the Drosophila 3′ splice site consensus, with both introns containing a non-consensus A at position –3, as well as interrupted polypyrimidine tracts. Sequences near the M1 5′ splice site (AT/GTAAGA) are also conserved in the M1 intron. However, in contrast to the 3′ splice site, this conservation is likely to be due only to the canonical requirement for these sequences. The conserved 5′ splice site closely matches the Drosophila 5′ splice site consensus (AG/GTRAGT) both in its extent and sequence and is thus similar to 5′ splice sites found in many other introns (29). A final conserved feature of M1 identified in the comparison between Drosophila species is an intron-internal sequence (the VC element) that is identical at 15 out of 18 positions but does not correspond to any known canonical splicing signals.
To determine if these conserved sequences are needed for regulation of M1 splicing we generated transgenic fly strains expressing altered tra-2 pre-mRNAs. Mutations of the M1 3′ splice site region suggest that the non-consensus sequences at this position are critical for normal splicing regulation. Substitution of the wild-type M1 3′ splice site region with sequences from a different non-consensus site supported TRA-2-dependent splicing repression but substitutions that more closely match the Drosophila consensus did not. One possibility is that a poor 3′ splice site limits the rate of splicing allowing regulatory factors an opportunity to intervene before splicing is initiated. This idea is supported by in vitro splicing experiments showing that substitution of the ftz 3′ splice site signals into M1 dramatically increases the rate of splicing relative to RNAs with the wild-type 3′ splice site (D.S.Chandler and W.Mattox, manuscript in preparation).
Although the non-consensus sequences in the wild-type 3′ splice site appear to confer a relatively slow rate of splicing on M1, the intron is removed from all germline tra-2 mRNA produced in the absence of functional TRA-2226 protein (4,16). Thus, the inherent ‘strength’ of 3′ splicing signals in the native M1 intron appears to be precisely tuned to a level that permits intron retention in the presence of repressor and complete splicing in the absence of repressor.
Non-consensus 3′ splice sites appear to be a common, but not universal, feature of introns where splicing is repressed. The 3′ splice sites of several other Drosophila RNAs with similarly regulated introns are shown in Figure 6 (3,33–36). Four of the five introns deviate significantly from the Drosophila 3′ splice site consensus (29), and in the fifth case (the Drosophila P element) splicing is known to be repressed through interactions of regulators near the 5′ splice site. We speculate that a reduced rate of basal splicing caused by non-consensus sequences near the 3′ splice sites in many such introns is essential for regulated intron retention in vivo.
Figure 6.

Comparison of 3′ splice site regions of introns that undergo splicing repression. The 3′ splice site regions of five introns which are retained in mature mRNA under specific developmental conditions or in response to known regulatory factors are shown in comparison to the D.melanogaster consensus sequences. The splice junctions are indicated by a forward slash and runs of three or more pyrimidines are underlined. Three of the five sequences (cyclin B and two introns from the SWAP gene) lack a polypyrimidine tract of >4 nt near the 3′ splice site. A fourth sequence (msl-2) has a polypyrimidine tract starting 16 nt upstream of the splice site and this spacing has been shown to reduce recognition of the splice site by U2AF in a manner important for splicing repression (46). Splicing of the intron from P element pre-mRNA is known to be repressed by complexes formed near the 5′ splice site of that intron (35).
Analysis of the VC element, the most highly conserved sequence within the M1 introns of D.melanogaster and D.virilis, showed that this sequence acts to facilitate M1 splicing. This effect appears to be specific to the somatic tissues of adults, where TRA-2-dependent repression of M1 splicing does not occur. When the VC element was deleted or substituted, adult flies accumulated high levels of somatic RNAs containing M1. Thus, the VC sequence appears to be necessary for efficient splicing of M1 in the soma.
The above results raise the issue of why the tra-2 pre-mRNA contains sequences that both positively and negatively affect M1 splicing. We hypothesized that positive elements, such as VC, might be required to overcome the weak 3′ splice site and allow efficient M1 splicing in somatic tissues. In support of this idea we found that substitution of the M1 3′ splice site with a strong 3′ site alleviated the need for the VC element in the soma. Thus, the VC element allows for efficient M1 splicing in the presence of the intron’s weak 3′ splice site, which is itself a necessary feature of germline splicing repression.
Neither of the sequences examined here appears to be a direct target of TRA-2226 or a co-repressor of M1 splicing. The VC element clearly has a positive rather than repressive role and alterations in it do not affect repression of M1 splicing in the germline. The weak M1 3′ splice site is required for germline splicing repression, but the ability of another weak 3′ splice site to functionally replace it suggests that it does not contain sequences recognized specifically by factors involved in M1 splicing repression. Therefore, it seems most likely that these conserved sequences affect only the basal level of M1 splicing in the soma and germline. The evolutionary conservation of such ‘context elements’ and the observation that alteration of these elements leads to qualitative changes in M1 splicing indicates that the basal splicing efficiency is critical to normal regulation. Increased rates of basal M1 splicing overcome the ability of TRA-2226 to repress it in the germline, while a decrease in basal splicing in the soma leads to accumulation of M1 containing transcripts where they are normally not found.
It is surprising that comparison of the tra-2 genes from different Drosophila species did not identify conserved sequences more directly involved in splicing repression. In a similar comparison, a critical 13 nt repeated element of the splicing enhancer from the dsx gene was found to be well conserved in the same two species (25,37). Several of the same 13 nt repeat elements are also found in the regulatory sequences within fru pre-mRNA, another target of tra-2 (12,14). Although this suggests that the 13 nt repeat element may be common to RNAs where splicing is affected by tra-2, no sequences identical to the 13 nt element were found in or near the M1 intron. In vivo studies on two related but non-identical and non-conserved repeat sequences in exon 3 upstream of the M1 intron showed that they are not required for M1 repression (D.S.Chandler and W.Mattox, unpublished). It is possible that the absence of a similar regulatory sequence in tra-2 pre-mRNA reflects differences in the role of the TRA protein, which is required in combination with TRA-2 for enhancement of dsx and fru splicing but does not function in the male germline where M1 splicing repression occurs. Thus, germline specific targets of tra-2 including the RNAs from tra-2, exuperantia and alternative-testes-transcript (38,39) may be regulated via different cis-regulatory elements than those found in dsx and fru.
The idea that regulation of alternative splicing depends on sequences that affect the efficiency of splicing is supported by observations that other regulated pre-mRNAs contain poor matches to consensus splicing signals (40–44). In several cases it has been shown that substitution of these sequences with consensus splice sites leads to constitutive use of the splice site and thus correct regulation of splicing depends on the weak splice site. In addition, some splicing enhancers have been shown to be essential for regulated splicing despite not being the direct targets of determinative tissue-specific regulators (45). Our results emphasize the sensitivity of splicing regulation to the basal rate of splicing and the importance of studying splicing regulatory elements within an appropriate context. Outside of a context where basal splicing occurs at normal efficiency, the effects of splicing regulators such as TRA-2 would not be observed.
Acknowledgments
ACKNOWLEDGEMENTS
We would like to thank Lisa Ryner, Tom Cooper, Miles Wilkinson, Sue Berget, Mitzi Kuroda and Steve Mount for their advice and useful discussions. This work was supported by NIH grant GM58625 and a Pew Scholars award to W.M. D.C. was supported by an NIH Training Grant, CA09299. DNA sequencing was supported by a Cancer Center Support (Core) Grant, CA16672.
References
- 1.Chabot B. (1996) Directing alternative splicing: cast and scenarios. Trends Genet., 12, 472–478. [DOI] [PubMed] [Google Scholar]
- 2.Graveley B.R. (2000) Sorting out the complexity of SR protein functions. RNA, 6, 1197–1211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Zachar Z., Chou,T.-B. and Bingham,P.M. (1987) Evidence that a regulatory gene autoregulates splicing of its transcript. EMBO J., 6, 4105–4111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Mattox W. and Baker,B.S. (1991) Autoregulation of the splicing of transcripts from the transformer-2 gene of Drosophila. Genes Dev., 5, 786–796. [DOI] [PubMed] [Google Scholar]
- 5.Bell L.R., Horabin,J.I., Schedl,P. and Cline,T.W. (1991) Positive autoregulation of sex-lethal by alternative splicing maintains the female determined state in Drosophila. Cell, 65, 229–239. [DOI] [PubMed] [Google Scholar]
- 6.Lejeune F., Cavaloc,Y. and Stevenin,J. (2000) Alternative splicing of intron 3 of the SR protein 9G8 gene: identification of flanking exonic splicing enhancers and involvement of 9G8 as a trans-acting factor. J. Biol. Chem., 276, 7850–7858. [DOI] [PubMed] [Google Scholar]
- 7.Sureau A., Gattoni,R., Dooghe,Y., Stevenin,J. and Soret,J. (2001) SC35 autoregulates its expression by promoting splicing events that destabilize its mRNAs. EMBO J., 20, 1785–1796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Nagoshi R.N., McKeown,M., Burtis,K.C., Belote,J.M. and Baker,B.S. (1988) The control of alternative splicing at genes regulating sexual differentiation in D.melanogaster. Cell, 53, 229–236. [DOI] [PubMed] [Google Scholar]
- 9.Ryner L.C. and Baker,B.S. (1991) Regulation of doublesex pre-mRNA processing occurs by 3′-splice site activation. Genes Dev., 5, 2071–2085. [DOI] [PubMed] [Google Scholar]
- 10.Tian M. and Maniatis,T. (1992) Positive control of pre-mRNA splicing in vitro. Science, 256, 237–240. [DOI] [PubMed] [Google Scholar]
- 11.Wu J.Y. and Maniatis,T. (1993) Specific interactions between proteins implicated in splice site selection and regulated alternative splicing. Cell, 75, 1061–1070. [DOI] [PubMed] [Google Scholar]
- 12.Ryner L.C., Goodwin,S.F., Castrillon,D.H., Anand,A., Villella,A., Baker,B.S., Hall,J.C., Taylor,B.J. and Wasserman,S.A. (1996) Control of male sexual behavior and sexual orientation in Drosophila by the fruitless gene. Cell, 87, 1079–1089. [DOI] [PubMed] [Google Scholar]
- 13.Zuo P. and Maniatis,T. (1996) The splicing factor U2AF(35) mediates critical protein–protein interactions in constitutive and enhancer-dependent splicing. Genes Dev., 10, 1356–1368. [DOI] [PubMed] [Google Scholar]
- 14.Heinrichs V., Ryner,L.C. and Baker,B.S. (1998) Regulation of sex-specific selection of fruitless 5′ splice sites by transformer and transformer-2. Mol. Cell. Biol., 18, 450–458. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Mattox W., McGuffin,M.E. and Baker,B.S. (1996) A negative feedback mechanism revealed by functional analysis of the alternative isoforms of the Drosophila splicing regulator transformer-2. Genetics, 143, 303–314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.McGuffin M.E., Chandler,D., Somaiya,D., Dauwalder,B. and Mattox,W. (1998) Autoregulation of transformer-2 alternative splicing is necessary for normal male fertility in Drosophila. Genetics, 149, 1477–1486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Amrein H., Maniatis,T. and Nothiger,R. (1990) Alternatively spliced transcripts of the sex-determining gene tra-2 of Drosophila encode functional proteins of different size. EMBO J., 9, 3619–3629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Mattox W., Ryner,L. and Baker,B.S. (1992) Autoregulation and multifunctionality among trans-acting factors that regulate alternative pre-mRNA processing. J. Biol. Chem., 267, 19023–19026. [PubMed] [Google Scholar]
- 19.Tian M. and Maniatis,T. (1993) A splicing enhancer complex controls alternative splicing of doublesex pre-mRNA. Cell, 74, 105–114. [DOI] [PubMed] [Google Scholar]
- 20.Amrein H., Hedley,M.L. and Maniatis,T. (1994) The role of specific protein–RNA and protein–protein interactions in positive and negative control of pre-mRNA splicing by Transformer 2. Cell, 76, 735–746. [DOI] [PubMed] [Google Scholar]
- 21.Tian M. and Maniatis,T. (1994) A splicing enhancer exhibits both constitutive and regulated activities. Genes Dev., 8, 1703–1712. [DOI] [PubMed] [Google Scholar]
- 22.Lynch K.W. and Maniatis,T. (1995) Synergistic interactions between two distinct elements of a regulated splicing enhancer. Genes Dev., 9, 284–293. [DOI] [PubMed] [Google Scholar]
- 23.Heinrichs V. and Baker,B.S. (1995) The Drosophila SR protein RBP1 contributes to the regulation of doublesex alternative splicing by recognizing RBP1 RNA target sequences. EMBO J., 14, 3987–4000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Lynch K.W. and Maniatis,T. (1996) Assembly of specific sr protein complexes on distinct regulatory elements of the Drosophila doublesex splicing enhancer. Genes Dev., 10, 2089–2101. [DOI] [PubMed] [Google Scholar]
- 25.Hertel K.J., Lynch,K.W., Hsiao,E.C., Liu,E.H. and Maniatis,T. (1996) Structural and functional conservation of the Drosophila doublesex splicing enhancer repeat elements. RNA, 2, 969–981. [PMC free article] [PubMed] [Google Scholar]
- 26.Dauwalder B. and Mattox,W. (1998) Analysis of the functional specificity of RS domains in vivo. EMBO J., 17, 6049–6060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Chandler D., McGuffin,M.E., Piskur,J., Yao,J., Baker,B.S. and Mattox,W. (1997) Evolutionary conservation of regulatory strategies for the sex determination factor transformer-2. Mol. Cell. Biol., 17, 2908–2919. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Pirrotta V. (1988) In Rodriguez,R.L. and Denhardt,D.T. (eds), Vectors: A Survey of Molecular Cloning Vectors And Their Uses. Butterworths, Boston, pp. 437–456.
- 29.Mount S. (1993) Messenger RNA splicing signals in Drosophila genes. In Maroni,G. (ed), Atlas of Drosophila Genes. Oxford University Press, New York, NY, pp. 333–358.
- 30.Wassenberg D.R.,II, Kronert,W.A., O’Donnell,P.T. and Bernstein,S.I. (1987) Analysis of the 5′ end of the Drosophila muscle myosin heavy chain gene. Alternatively spliced transcripts initiate at a single site and intron locations are conserved compared to myosin genes of other organisms. J. Biol. Chem., 262, 10741–10747. [PubMed] [Google Scholar]
- 31.Mattox W., Palmer,M.J. and Baker,B.S. (1990) Alternative splicing of the sex determination gene transformer-2 is sex-specific in the germ line but not in the soma. Genes Dev., 4, 789–805. [DOI] [PubMed] [Google Scholar]
- 32.Nayler O., Cap,C. and Stamm,S. (1998) Human transformer-2-beta gene (SFRS10): complete nucleotide sequence, chromosomal localization and generation of a tissue-specific isoform. Genomics, 53, 191–202. [DOI] [PubMed] [Google Scholar]
- 33.Dalby B. and Glover,D.M. (1992) 3′ non-translated sequences in Drosophila cyclin B transcripts direct posterior pole accumulation late in oogenesis and peri-nuclear association in syncytial embryos. Development, 115, 989–997. [DOI] [PubMed] [Google Scholar]
- 34.Siebel C.W. and Rio,D.C. (1990) Regulated splicing of the Drosophila P transposable element third intron in vitro: somatic repression. Science, 248, 1200–1208. [DOI] [PubMed] [Google Scholar]
- 35.Siebel C.W., Fresco,L.D. and Rio,D.C. (1992) The mechanism of somatic inhibition of Drosophila P-element pre-mRNA splicing: multiprotein complexes at an exon pseudo-5′ splice site control U1 snRNP binding. Genes Dev., 6, 1386–1401. [DOI] [PubMed] [Google Scholar]
- 36.Zhou S., Yang,Y., Scott,M.J., Pannuti,A., Fehr,K.C., Eisen,A., Koonin,E.V., Fouts,D.L., Wrightsman,R., Manning,J.E. et al. (1995) Male-specific lethal 2, a dosage compensation gene of Drosophila, undergoes sex-specific regulation and encodes a protein with a RING finger and a metallothionein-like cysteine cluster. EMBO J., 14, 2884–2895. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Burtis K.C. and Baker,B.S. (1989) Drosophila doublesex gene controls somatic sexual differentiation by producing alternatively spliced mRNAs encoding related sex-specific polypeptides. Cell, 56, 997–1010. [DOI] [PubMed] [Google Scholar]
- 38.Hazelrigg T. and Tu,C. (1994) Sex-specific processing of the Drosophila exuperantia transcript is regulated in male germ cells by the tra-2 gene. Proc. Natl Acad. Sci. USA, 91, 10752–10756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Madigan S.J., Edeen,P., Esnayra,J. and McKeown,M. (1996) Att, a target for regulation by Tra2 in the testes of Drosophila melanogaster, encodes alternative RNAs and alternative proteins. Mol. Cell. Biol., 16, 4222–4230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Hoshijima K., Inoue,K., Higuchi,I., Sakamoto,H. and Shimura,Y. (1991) Control of doublesex alternative splicing by transformer and transformer-2 in Drosophila. Science, 252, 833–836. [DOI] [PubMed] [Google Scholar]
- 41.Watakabe A., Tanaka,K. and Shimura,Y. (1993) The role of exon sequences in splice site selection. Genes Dev., 7, 407–418. [DOI] [PubMed] [Google Scholar]
- 42.Sun Q., Mayeda,A., Hampson,R.K., Krainer,A.R. and Rottman,F.M. (1993) General splicing factor SF2/ASF promotes alternative splicing by binding to an exonic splicing enhancer. Genes Dev., 7, 2598–2608. [DOI] [PubMed] [Google Scholar]
- 43.Humphrey M.B., Bryan,J., Cooper,T.A. and Berget,S.M. (1995) A 32-nucleotide exon-splicing enhancer regulates usage of competing 5′ splice sites in a differential internal exon. Mol. Cell. Biol., 15, 3979–3988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Zheng Z.M., Quintero,J., Reid,E.S., Gocke,C. and Baker,C.C. (2000) Optimization of a weak 3′ splice site counteracts the function of a bovine papillomavirus type 1 exonic splicing suppressor in vitro and in vivo. J. Virol., 74, 5902–5910. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Xu R., Teng,J. and Cooper,T.A. (1993) The cardiac troponin T alternative exon contains a novel purine-rich positive splicing element. Mol. Cell. Biol., 13, 3660–3674. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Merendino L., Guth,S., Bilbao,D., Martinez,C. and Valcarcel,J. (1999) Inhibition of msl-2 splicing by Sex-lethal reveals interaction between U2AF35 and the 3′ splice site AG. Nature, 402, 838–841. [DOI] [PubMed] [Google Scholar]