

Leaf thickness in desert-adapted tomato is characterized by the anatomic and transcriptional alterations that are uncovered by QTL analysis of introgression lines.
Abstract
Thicker leaves allow plants to grow in water-limited conditions. However, our understanding of the genetic underpinnings of this highly functional leaf shape trait is poor. We used a custom-built confocal profilometer to directly measure leaf thickness in a set of introgression lines (ILs) derived from the desert tomato Solanum pennellii and identified quantitative trait loci. We report evidence of a complex genetic architecture of this trait and roles for both genetic and environmental factors. Several ILs with thick leaves have dramatically elongated palisade mesophyll cells and, in some cases, increased leaf ploidy. We characterized the thick IL2-5 and IL4-3 in detail and found increased mesophyll cell size and leaf ploidy levels, suggesting that endoreduplication underpins leaf thickness in tomato. Next, we queried the transcriptomes and inferred dynamic Bayesian networks of gene expression across early leaf ontogeny in these lines to compare the molecular networks that pattern leaf thickness. We show that thick ILs share S. pennellii-like expression profiles for putative regulators of cell shape and meristem determinacy as well as a general signature of cell cycle-related gene expression. However, our network data suggest that leaf thickness in these two lines is patterned at least partially by distinct mechanisms. Consistent with this hypothesis, double homozygote lines combining introgression segments from these two ILs show additive phenotypes, including thick leaves, higher ploidy levels, and larger palisade mesophyll cells. Collectively, these data establish a framework of genetic, anatomical, and molecular mechanisms that pattern leaf thickness in desert-adapted tomato.
Leaves are the primary photosynthetic organs of land plants. Quantitative leaf traits have important connections to their physiological functions and, ultimately, to whole-plant productivity and survival. While few aspects of leaf morphology have been determined unambiguously as functional (Nicotra et al., 2011), clear associations between leaf traits and variations in climate have been drawn (Wright et al., 2004). Leaf thickness, the distance between the upper (adaxial) and lower (abaxial) leaf surfaces, has been shown to correlate with environmental variables such as water availability, temperature, and light quantity. Thus, on a global scale, across habitats and land plant diversity, plants adapted to arid environments tend to have thicker leaves (Wright et al., 2004; Poorter et al., 2009).
Leaf thickness is a continuous, rather than a categorical, trait. Thus, it is important to distinguish between thickness in the context of typical leaf morphology, generally possessing clear dorsiventrality (adaxial/abaxial flattening), in comparison with extremely thick leaves, described as succulent, which are often more radial. While the definition of succulence is ecophysiological rather than morphological (Ogburn and Edwards, 2010), at the cellular level it is broadly associated with increased cell size and relative vacuole volume (Gibson, 1982; von Willert, 1992). These cellular traits promote the capacity to store water and to survive in dry environments (Becker, 2007). Allometric studies across land plant families have shown that leaf thickness scales specifically with the size of palisade mesophyll cells: the adaxial layer of photosynthetic cells in leaves (Garnier and Laurent, 1994; Roderick et al., 1999; Sack and Frole, 2006; John et al., 2013). Increased palisade cell height leads to an increased area of contact with the intercellular space and, thereby, to improved uptake of carbon dioxide (CO2) into mesophyll cells (Oguchi et al., 2005; Terashima et al., 2011), possibly offsetting the increased CO2 diffusion path in thicker leaves. At the organismal level, thicker leaves present a tradeoff between rapid growth versus drought and heat tolerance (Smith et al., 1997). This idea is supported by global correlations between leaf mass per area (LMA), a proxy for leaf thickness, and habits associated with slower growth (Poorter et al., 2009).
Although leaf thickness is a highly functional trait, mechanistic understanding of how it is patterned during leaf ontogeny is poor. The main cellular events that underpin leaf development are the establishment of adaxial/abaxial polarity, followed by cell division, directional expansion, and differentiation (Efroni et al., 2008). Changes in the relative timing (heterochrony) and duration of these events can impact leaf morphology, including thickness. Several mutants have been identified that show clear alterations in leaf thickness. These include the Arabidopsis (Arabidopsis thaliana) angustifolia and rotundifolia3 (Tsuge et al., 1996) as well as argonaute1, phantastica, and phabulosa (Bohmert et al., 1998), which have aberrations in the polarity of cell elongation and the establishment of adaxial/abaxial polarity, respectively, as well as Nicotiana sylvestris fat and lam-1 (McHale, 1992, 1993), which affect the extent of periclinal cell division in leaves. However, these developmental mutants do not necessarily inform us of the mechanisms by which natural selection acts to pattern quantitative variation in leaf thickness.
Efforts to understand the genetic basis of leaf thickness in the context of natural variation face several important challenges. First, direct measurement of leaf thickness at a scale that would allow the investigation of quantitative trait loci (QTL) for the trait is not trivial. Because of the difficulty in measuring leaf thickness directly, LMA is used most often as a proxy for this trait (Poorter et al., 2009; Muir et al., 2014). Second, in addition to genetic components, leaf thickness is environmentally plastic: it is responsive to both the quantity and quality of light (Pieruschka and Poorter, 2012). Finally, because leaf thickness varies on a continuous spectrum and is not associated with any particular phylogenetic lineage or growth habit, mechanistic questions regarding its patterning need to be addressed in a taxon-specific manner.
With these considerations in mind, we used two members of the tomato clade (Solanum sect. Lycopersicon) that are closely related, morphologically distinct, and occupy distinct environments (Nakazato et al., 2010) to study the genetic basis and developmental patterning of leaf thickness. The domesticated tomato species Solanum lycopersicum inhabits a relatively wide geographic range characterized by warm, wet conditions with little seasonal variation. By contrast, the wild species Solanum pennellii is endemic to coastal regions of the Atacama Desert of Peru, a habitat characterized by extremely dry conditions (Nakazato et al., 2010). The leaves of S. pennellii plants, therefore, exhibit morphological and anatomical features that are likely adaptations to dry conditions (McDowell et al., 2011; Haliński et al., 2015), including thick leaves (Koenig et al., 2013). Moreover, a set of homozygous introgression lines (ILs) harboring defined, partially overlapping segments of the S. pennellii genome in an otherwise S. lycopersicum background (Eshed and Zamir, 1995) has been used to successfully map a number of QTL, including fruit metabolite concentrations (Fridman et al., 2004; Schauer et al., 2006), yield (Semel et al., 2006), and leaf shape (Chitwood et al., 2013). Here, we used a custom-built dual confocal profilometer to obtain precise measurements of leaf thickness across the IL panel and identified QTL for this trait in tomato. Leaf thickness correlates with other facets of leaf shape as well as a suite of traits associated with desiccation tolerance and lower productivity. We investigated the anatomical manifestations of thickness in tomato and found a prominent increase in palisade cell height in many thick ILs. Finally, we inferred comparative gene regulatory networks of early leaf development (plastochron stages P1–P4) in two thick lines using organ-specific RNA sequencing (RNA-Seq) and identified molecular networks that pattern S. pennellii-like desert-adapted leaves.
RESULTS
Complex Genetic Architecture of Leaf Thickness across S. pennellii ILs
To investigate the genetic architecture and patterning of leaf thickness in the S. pennellii IL panel, we used a custom-built dual confocal profilometer device (Supplemental Fig. S1), which generates precise thickness measurements throughout the leaflet lamina at a range of resolutions (0.1–1 mm2) and at high throughput. The device makes use of two confocal lasers positioned on either side of the sample and calculates thickness by measuring the distance between each of the sample’s surfaces and the corresponding laser probe. Finally, we visualize thickness as a heat map of thickness values across the surface of the leaf lamina (Fig. 1A).
Figure 1.

Desert-adapted tomato plants have thicker leaves than domesticated tomato and are resistant to drought. A, Thickness across leaflet blades of domesticated (S. lycopersicum M82) and desert-adapted (S. pennellii) tomatoes measured with a custom-built dual confocal profilometer device (Supplemental Fig. S1). The median thickness of the S. lycopersicum leaflet shown here is 211 μm, and that for S. pennellii is 294 μm. B, Confocal images of propidium iodide-stained leaflet cross sections. Bars = 200 μm. C, Total shoot area normalized by taking the square root of pixels (px) from top-view phenotyping images over 16 d in three water treatments (n = 8). Gray shading reflects se.
We first compared leaflet thickness in S. lycopersicum variety M82 and its desert relative S. pennellii LA0716. Our confocal profilometer measurements showed that S. pennellii leaflets are thicker than those of domesticated tomato, as reported previously (Fig. 1; Koenig et al., 2013), demonstrating the capacity of this device to quantitatively detect fine differences in leaf lamina thickness. We compared dynamic growth patterns of the two species under water-limited conditions and show that, unlike the domesticated species, S. pennellii is unaffected by drought (Fig. 1C). This observation highlights the importance of understanding the patterning of developmental traits in this species, such as leaf thickness, which may contribute to drought tolerance. We proceeded to measure leaf thickness across the S. pennellii IL panel in field conditions.
We used mixed linear regression models to compare each of the ILs with the domesticated parent M82 (Supplemental Data Set S1) and found that 31 ILs had significantly thicker leaflets than the M82 parent, while five had transgressively thinner leaflets. The overall broad-sense heritability for leaflet thickness is 39.1% (Fig. 2). The lines with thickest leaflets are IL5-4, IL5-3, IL8-1, IL4-3, IL8-1-1 (contained within IL8-1), and IL2-5, while IL4-1-1, IL2-6-5, IL9-1-3, IL12-4-1, and IL2-1 have thinner leaves than the M82 parent.
Figure 2.

QTL for leaf thickness in tomato. A, Leaflet thickness values across the S. pennellii IL panel. Colors indicate the level of significance in comparisons of each IL with M82 (arrow). B, Significant correlations (Spearman’s rho) between leaf thickness or LMA and a suite of other traits across the S. pennellii IL panel (q < 0.05). Traits are grouped by type: ION, elemental profile; MOR, morphological; DEV, developmental; ENZ, enzyme activity; SED, seed metabolite content (Supplemental Data Sets S3 and S4).
Based on the observation that the heritability value for leaf thickness is 39.1%, we reasoned that environmental factors are likely to play a role in modulating leaf thickness. We thus compared our field experiment with leaf thickness data for vegetative leaves of greenhouse-grown plants. We selected 20 ILs, which were highly significant for leaf thickness differences from M82 in field conditions (P < 0.001), and observed that only some of these lines also are significantly thicker than the domesticated parent in greenhouse conditions (P < 0.05; Supplemental Fig. S2A). Finally, our observations suggest that leaf thickness varies across the shoot of a number of our select thick leaf ILs, with post-flowering leaves having thicker leaves than vegetative leaves (Supplemental Fig. S2B).
For each leaflet in our field experiment, we also quantified LMA, which reflects both thickness and density and is traditionally used as a proxy for leaf thickness. Although the heritability for LMA is similar to that for thickness (33.2% and 39.1%, respectively), significant QTL for these two traits do not overlap consistently (Supplemental Data Set S1).
Leaf Thickness and LMA Are Correlated with Distinct Suites of Traits in Tomato
We generated pairwise correlations between leaflet thickness, LMA, and a suite of other previously published traits, including metabolite, morphological, enzymatic activity in fruit pericarp, seed-related, developmental, and elemental profile-related traits (Supplemental Data Sets S2–S4; Chitwood et al., 2013, and refs. therein). Spearman’s correlation coefficients with significant q values (q < 0.050) are reported in Figure 2B. Leaf thickness and LMA are correlated (rho = 0.423, q = 0.003). Leaf thickness also correlates with leaf shape parameters, such as roundness (rho = 0.328, q = 0.044), aspect ratio (rho = −0.327, q = 0.045), and the first two principal components of the elliptical Fourier descriptors of leaflet shape (EFD.PC1 rho = 0.414, q = 0.004 and EFD.PC2 rho = 0.406, q = 0.005). Thickness is negatively correlated with several reproductive traits, including yield (rho = −0.337, q = 0.037), seed weight (rho = −0.342, q = 0.033), and seed number per plant (rho = −0.339, q = 0.036). Moreover, leaf thickness is negatively correlated with leaf stomatal ratio, the relative density of stomata on the abaxial and adaxial sides of the leaf (rho = −0.352, q = 0.031), and positively correlated with Glu dehydrogenase activity (rho = 0.367, q = 0.017) and seed galactinol content (rho = 0.342, q = 0.048).
LMA is associated with a distinct suite of traits from leaf thickness. In addition to a positive correlation with the content of some enzymes (GAPDH and Shikimate DE) and metabolites (Glu), LMA is significantly negatively correlated with the accumulation of Na and Mg in all leaflets tested. LMA, but not leaf thickness, also is significantly positively correlated with total plant weight, reflecting vegetative biomass accumulation.
Thick IL Leaves Have Elongated Palisade Parenchyma Cells
Leaf cross sections of field-grown M82 and select ILs with increased leaf thickness, as well as greenhouse-grown S. pennellii leaves, were stained with propidium iodide to assess the anatomical changes that lead to increased leaf thickness. We observed that, relative to the M82 parent, the S. pennellii parent and several ILs have an elongated palisade mesophyll cell layer corresponding to the adaxial layer of photosynthesizing cells in tomato leaves (Fig. 3). Palisade parenchyma elongation is especially dramatic for IL1-3, IL2-5, IL4-3, and IL10-3. Both leaf thickness and palisade elongation phenotypes are attenuated for vegetative leaves of greenhouse-grown plants (Supplemental Figs. S2 and S3A).
Figure 3.

Anatomical manifestations of thicker leaves. A, Confocal images of propidium iodide-stained cross sections of field-grown M82, select ILs, and S. pennellii grown in greenhouse conditions. Bars = 50 μm. B and C, Representative leaf thickness plots (B) and leaflet binary images of field-grown plants (C) as for A.
Anatomy and Early Leaf Development in Select ILs with Thick Leaves
To capture an overall view into the core mechanisms of leaf thickness patterning, we further analyzed lines IL2-5 and IL4-3. We selected IL2-5 due to its dramatic anatomy in field conditions (Fig. 3) and its lack of other characterized leaf morphology phenotypes (Chitwood et al., 2013), while IL4-3 leaflets are both significantly thicker and less serrated than those of the domesticated parent (Fig. 2; Supplemental Data Set S1; circularity, the ratio between leaflet area and the square of its perimeter, reflects lobing and serration). To further investigate the relationships between genetic determinants of leaf thickness in these ILs, we generated a double homozygous line combining the entire S. pennellii segments of IL2-5 and IL4-3.
Double homozygotes (IL2-5/IL4-3) have significantly thicker leaves than M82 at both vegetative (Fig. 4A; P = 0.019) and postflowering stages (Supplemental Fig. S2B) in greenhouse conditions. Additionally, IL2-5/IL4-3 plants have significantly smoother margins than either of the IL parents (Fig. 4B), suggesting additive genetic interactions for both of these traits. We next compared the dimensions of the mesophyll cell layers in each IL and the double homozygote line to determine the contribution that each cell layer makes to the observed increase in leaflet thickness. We found that palisade mesophyll cells are significantly larger in IL2-5/IL4-3 than in M82 leaves (Supplemental Fig. S4). Furthermore, the ratios of palisade cell length to both total leaf thickness and the length of the spongy mesophyll are significantly larger in IL2-5/IL4-3 than in M82 leaves (Supplemental Fig. S4). IL2-5 shows similar, albeit less pronounced, trends to the double homozygote line, while in IL4-3, both spongy and palisade mesophyll cell layers are longer than in M82, with the spongy mesophyll layer making the most significant contribution to leaf thickness.
Figure 4.

Leaf morphology and ploidy of IL2-5/IL4-3 double homozygote plants. A, Representative propidium iodide-stained leaflet cross sections (left) and thickness measurements (right) for the seventh leaf of greenhouse-grown M82 and double homozygous IL2-5/IL4-3 plants (n = 10). **, P < 0.01. Bars = 200 μm. B, Circularity (ratio of area to the square of the perimeter) of distal lateral leaflets as in A. Silhouettes of representative M82 and IL2-5/IL4-3 leaflets are shown above the bars. Letters indicate statistical significance in each pairwise genotype comparison (P < 0.05). C, Distribution of relative nuclear sizes reflecting endoreduplication in leaflets as in A and B (n = 5). Letters denote statistical significance between pairwise genotype comparisons at each ploidy level. Sp, S. pennellii. D, Leaf plastochron P3 dimensions calculated from 3D surface reconstructions of vegetative shoot apices (n = 9; *, P < 0.05 and **, P < 0.01 relative to M82).
Since increases in cell size often are driven by endopolyploidy, we performed flow cytometry on fully expanded vegetative leaves of each genotype and observed increased ploidy profiles in all lines relative to the domesticated parent (Fig. 4C). Notably, the double homozygote line exhibited higher ploidy levels than both single ILs and the S. pennellii parent (Fig. 4C; Supplemental Fig. S4). Notably, we also observed a trend to increased ploidy in several greenhouse-grown thick ILs (IL7-4-1 and IL8-1; Supplemental Fig. S3B).
To understand if alterations in leaf size occur during early stages of leaf ontogeny in these lines, we quantified P3 organ dimensions and compared them with the M82 parental line. For this, we assembled 3D confocal reconstructions of vegetative shoot apices, calculated the surface mesh, extracted P3 leaf primordia, and quantified their total volume, length, and mean diameter. We found that IL4-3 P3 leaf primordia are significantly larger than M82 primordia in terms of overall volume (P = 0.0179) as well as both length (P = 0.0035) and diameter (P = 0.0230). In IL2-5 P3, volume (not statistically significant) and diameter (P = 0.0116) are increased, while length is comparable to that in M82. Although P3 primordia of double homozygote plants were statistically indistinguishable from those of M82 plants except for shorter arc length (P = 0.0411; Fig. 4D), our observations also suggest that double homozygote leaves increase in size dramatically between P3 and P4 stages (Supplemental Fig. S5).
Transcriptomic Signatures of Early Leaf Development in Thicker ILs
To investigate the molecular events that define the patterning of IL2-5 and IL4-3 leaves, we isolated leaf primordia from each IL and the two parents (M82 or S. lycopersicum and S. pennellii) at four successive stages of development: P1 (containing the shoot apical meristem [SAM] and the youngest leaf primordium), P2 and P3 (characterized by leaflet emergence), and P4 (typically the onset of cell differentiation; Fig. 5A). For S. pennellii, P1 samples were composed of the SAM, P1, and P2, since these organs were not separable by hand dissection. Thus, the S. pennellii transcriptomic data set includes samples designated as P1, P3, and P4. Principal component analysis of the resulting RNA-Seq data, after normalization and filtering, shows that samples group clearly by organ stage (Fig. 5B, PC2). In addition, PC1 separates S. pennellii samples from all other genotypes. To investigate how IL leaves are similar to the S. pennellii parent, we looked for genes that are differentially expressed (DEGs) between corresponding stages of each IL and the M82 parent while also being differentially expressed between M82 and S. pennellii. In other words, we identified the set of DEGs for each organ stage that is common to each IL and S. pennellii relative to M82. For P2, we considered only the comparison with M82, as our S. pennellii data set did not include independently dissected P2 stage primordium samples (Supplemental Fig. S6; Supplemental Data Set S5).
Figure 5.

Comparative transcriptomics of leaf development in two thick ILs and their parents. A, Successive stages of leaf development (plastochrons P1–P4 colored as in the legend to B) were dissected from M82, S. pennellii (Sp), and thick IL2-5 and IL4-3. B, Principal component analysis of normalized RNA-Seq read counts. C, Venn diagrams (not to scale) depict an overview of DEGs (q < 0.05) that are shared in each IL and the S. pennellii parent relative to M82. The number of DEGs unique to each organ is shown within the ellipses, and those common to all organs are shown in the center. The total number of DEGs at each plastochron stage is shown outside the ellipses.
We identified a total of 812 DEGs across the P1 to P4 stages in IL2-5, and of these, 544 are up-regulated in at least one organ stage while 269 are down-regulated (Fig. 5C). In IL4-3, we detected 632 DEGs, 361 of which are up-regulated and 271 are down-regulated in the IL (Fig. 5C). Many of the DEGs are differentially expressed at more than one stage (Fig. 5C; Supplemental Data Set S5). Additionally, based on tomato transcription factor (TF) annotation by Suresh et al. (2014), we identified putative TF-encoding genes among each IL’s DEG sets. Myb-related, Ethylene Responsive, MADS, and WRKY are the abundant classes of TF-encoding DEGs in IL2-5, while in IL4-3, TFs belonging to bZIP and Myb-related are highly represented families (Supplemental Fig. S7).
We identified differentially expressed TF-encoding genes that are common to the two ILs and the S. pennellii parent (Fig. 6), reasoning that some of these can be regulators of leaf thickness. Five of the seven shared TF-encoding genes are up-regulated in the ILs relative to M82. A MADS-box TF (Solyc12g087830) is up-regulated at all stages in both ILs, while two additional inflorescence meristem-related TFs, LFY-like (Solyc03g118160) and AP2-like (Solyc07g049490), are differentially expressed at corresponding stages in both ILs. The SHORTROOT-like (SHR-like) GRAS TF Solyc08g014030 is up-regulated at P2 in both ILs, while its expression increases at each progressive stage and peaks at P4 in all genotypes. A putative JASMONATE ZIM-domain protein (JAZ1; Solyc12g009220) also is up-regulated at P2 in both ILs, while a LIM domain protein (Solyc04g077780) is up-regulated in the ILs at P3 (in IL4-3) and P4 (both ILs; Fig. 6A).
Figure 6.

Comparative expression profiles of genes in three functional categories across leaf development (P1–P4) in thick IL2-5 and IL4-3. A, Transcription factors common to both ILs. B, Genes involved in leaf development in tomato (as in Ichihashi et al., 2014). C, Genes annotated to encode components of the cell cycle or ubiquitin proteasome pathway (they contain one of the terms cell cycle, cyclin, ubiquitin, E2F, mitosis, mitotic, or SKP). Plastochron stages with statistically significant DEGs (q < 0.05) relative to M82 are marked with asterisks. Genes that are differentially expressed in at least one stage in both ILs are marked in boldface.
Next, we compared the expression profiles of genes known to be involved in tomato leaf development (Ichihashi et al., 2014). We selected only genes that are differentially expressed in the same direction in each IL and S. pennellii relative to the domesticated parent M82 and highlighted genes that are common to both thick ILs to arrive at a set of entities that may be core to the patterning of leaf thickness (Fig. 6B). A gibberellin 20-oxidase-encoding gene (GA 20-ox; Solyc03g006880) is up-regulated at P3 in both ILs and throughout the P1-P3 interval in IL4-3. A set of two closely related ULTRAPETALA1 genes (Solyc12g010360 and Solyc12g010370) is down-regulated at all leaf developmental stages in both ILs. A number of leaf development regulators are additionally differentially expressed in either of the ILs. Some noteworthy classes include entities related to auxin metabolism or transport (auxin efflux carrier, IAA-carboxymethyltransferase, and YUCCA-like monooxygenase), leaf complexity, lobing, and serrations (three BEL1-like TFs, CUC2-like, and BOP2-like), meristem maintenance or patterning (two BAM1-like receptor kinases and an AP2-like TF), and cell division and expansion (GRF1 and ROT3-like TFs).
Similarly, we also queried DEG sets for entities annotated as cell cycle or endoreduplication to assess whether these two thick ILs share a common trajectory of cellular events during leaf ontogeny (Fig. 6C). Overall, we observed distinct expression profiles for these genes in IL2-5 and IL4-3.
Finally, to broadly characterize the types of processes that may regulate the molecular networks of early leaf development in the ILs, we applied Gene Ontology (GO) enrichment analysis (agriGO; Du et al., 2010; Supplemental Data Set S6) and identified statistically enriched promoter motifs among the organ-specific DEG sets (Supplemental Data Set S7). Importantly, we observed that, at P4, the set of up-regulated genes in IL2-5 is enriched for biological process terms relating to photosynthesis (GO:0015979) and translation (GO:0006412), while down-regulated genes at this stage are enriched for terms relating to DNA binding (GO:0003677). Our promoter motif analysis showed that motifs associated with regulation by abiotic factors such as light, circadian clock, water availability, and temperature are prominent among IL2-5 DEGs. In addition, binding sites for developmental regulators, hormone-associated promoter motifs, and a cell cycle regulator are among the list of significant motifs. Among development-associated motifs, CArG (MADS-box), BEL1-like, and SBP-box TF-binding sites also are enriched significantly in both IL2-5 and IL4-3 DEG sets. (Supplemental Fig. S8; Supplemental Data Set S7).
Gene Regulatory Networks of Early Leaf Development in Thick ILs
To detect regulators of early leaf development that each IL (IL2-5 and IL4-3) shares with the S. pennellii parent, we inferred dynamic Bayesian networks using the IL and S. pennellii overlapping DEG sets described in the previous section (de Luis Balaguer et al., 2017). Additionally, we only allowed putative TF-encoding genes (Suresh et al., 2014) as source nodes (genes that control the expression of other coexpressed genes). First, we constructed individual networks for each leaf developmental stage, for which an overlap with S. pennellii data is available (P1, P3, and P4) and then combined the results to visualize the overall S. pennellii-like leaf developmental networks (Fig. 7; Supplemental Data Set S8). The IL2-5 network (Fig. 7A) contains two major regulators, which are central to more than one developmental stage: a SQUAMOSA promoter-binding protein-like domain gene (SBP-box 04g, Solyc04g064470) and a CONSTANS-like zinc finger (Zn-finger CO-like 05g, Solyc05g009310; Supplemental Data Set S8). Similarly, the IL4-3 network (Fig. 7C) features two central regulators: a BEL1-like homeodomain TF gene (BEL1 04g, Solyc04g080780) and a MADS-box domain-containing gene (MADS-box 12g, Solyc12g087830; Supplemental Data Set S8). Importantly, few nodes are shared between the organ-specific networks of IL2-5 and IL4-3. We surveyed each network for shared differentially expressed leaf development genes and found that GA 20-ox 03g (Solyc03g006880) is present in both networks but is regulated by different sets of TFs in each IL (Fig. 7, B and D).
Figure 7.

Select leaf development gene regulatory subnetworks for IL2-5 (A–C) and IL4-3 (D–F). Subnetworks for regulators central to more than one plastochron stage are shown in A and D. A GA 20-ox gene (Solyc03g006880) and its regulators in each IL are shown in B and E. Subnetworks of dynamic gene regulatory networks, showing interactions of an AUX/IAA TF (AUX/IAA 12g, Solyc12g096980) with other source nodes, are shown in C and F. Gene identifiers of the highlighted nodes are as follows: SBP-box 04g, Solyc04g064470; BEL1-like 04g, Solyc04g080780; MADS-box 12g, Solyc12g087820; MYB TF 05g, Solyc05g007710; bHLH TF 04g, Solyc04g074810; GRAS 08g, Solyc08g014030; Myb 07g, Solyc07g052490; WRKY 05g, Solyc05g015850; AUX/IAA 06g, Solyc06g008580; Myb 03g, Solyc03g005570; Myb 08g, Soly08g005870; and WRKY 02g, Solyc02g080890. Nodes and edges are colored according to the legend.
We also inferred a second set of networks for each of the ILs by identifying DEGs using similar criteria to those described above. However, in contrast to the previous set of networks, where genes were separated into organ stages based on differential expression at each discrete stage, we used a clustering approach to group regulators and select coexpressed gene sets according to expression profiles. For these analyses, we also included P2 DEGs (IL versus M82) to ensure the continuity of expression profiles (Supplemental Data Set S9). This approach allowed us to examine a more dynamic view of early developmental processes. The resulting networks (Supplemental Data Set S9) feature a putative auxin-responsive TF, AUX/IAA 12g (Solyc12g096980), for both ILs (Fig. 7, E and F). Moreover, the AUX/IAA 12g subnetwork or IL2-5 includes the SHR-like GRAS domain TF that is up-regulated during leaf development in both ILs (GRAS 08g, Solyc08g014030; Figs. 6A and 7E).
DISCUSSION
Leaf Thickness Has a Complex Genetic Architecture in Desert-Adapted Tomato and Is Associated with Overall Leaf Shape, Desiccation Tolerance, and Decreased Yield
While extensive progress has been made dissecting the molecular-genetic patterning of two-dimensional leaf morphology, relatively little is known about the third dimension of leaf shape: thickness. Here, we used a custom-built dual confocal profilometer to obtain direct measurements of leaf thickness across the S. pennellii × S. lycopersicum IL panel (Eshed and Zamir, 1995; Fig. 1; Supplemental Fig. S1) and identified QTL for this trait (Fig. 2A). We found that nearly half of the ILs have significantly thicker leaves than the domesticated parent M82, while a small number have transgressively thinner leaves. The broad-sense heritability for leaf thickness in this experiment is moderate (39%). Collectively, these observations point to a complex genetic basis for this trait. A previous quantitative genetic analysis of a suite of desert-adaptive traits in the same S. pennellii IL panel found fewer significantly thicker lines and lower heritability (12%) for this trait (Muir et al., 2014). However, the previous study estimated thickness as the ratio of LMA to leaflet dry matter content, while we measured thickness directly. Furthermore, our study was conducted in field conditions, while Muir et al. (2014) measured the trait using greenhouse-grown plants. Given that environment significantly affects the magnitude of this trait (Supplemental Fig. S2), it is not surprising that these studies report only partially overlapping outcomes.
In order to understand how variation in leaf thickness relates to other traits, particularly to LMA, we calculated pairwise correlation coefficients among all leaf shape and elemental profile traits as well as a collection of previously published traits (summarized by Chitwood et al., 2013; Supplemental Data Sets S3 and S4). As expected, leaf thickness and LMA are correlated significantly across the IL panel. However, the two traits have distinct sets of significant trait correlations (Fig. 2B). Taken together with our finding that only a proportion of ILs harbor QTL for both leaf thickness and LMA (Supplemental Data Set S1), these data suggest that thickness and LMA are likely patterned by partially distinct mechanisms and that direct measurements of leaf thickness are necessary to further dissect the genetic basis of this trait.
Leaf thickness is correlated significantly with leaf shape traits such as aspect ratio and the first two principal components of elliptical Fourier descriptors of overall shape. However, our data do not establish whether this correlation reflects a common patterning mechanism or developmental and/or mechanical constraints among these traits. Alternatively, the relatively modest correlations (rho values between 0.33 and 0.41) could reflect independent variation in these traits resulting in considerable flexibility in final leaf morphology, as suggested by Muir et al. (2017).
Leaf thickness is negatively correlated with yield-related traits, which suggests a tradeoff between investments in vegetative and reproductive biomass that is further substantiated by the positive correlation between LMA and plant weight (Fig. 2B). Some studies support the hypothesis of a tradeoff between LMA and rapid growth (Smith et al., 1997; Poorter et al., 2009), while others find poor coordination between growth rate and LMA (Muir et al., 2017). Finally, leaf thickness is correlated significantly with leaf stomatal ratio, Glu dehydrogenase activity, and galactinol content in seeds, a suite of traits associated with desiccation tolerance in plants (Taji et al., 2002; Lightfoot et al., 2007). We also observed negative correlations between LMA and the accumulation of several elements in leaves, most notably Na and Mg (Fig. 2C). This finding supports the idea that LMA and thickness are distinct traits, and that LMA reflects the material composition of leaves, while leaf thickness is a developmentally patterned trait.
Thicker S. pennellii IL Leaves Have Elongated Palisade Mesophyll Cells
The observed elongated palisade mesophyll cells in the leaves of several field-grown ILs with significantly thicker leaves (Fig. 3A), as well as in the desert-adapted S. pennellii parent, suggest that the dorsiventral expansion of palisade mesophyll cells contributes most prominently to increased leaf thickness. This hypothesis is supported by the fact that palisade cell height increases more significantly than the total height of the spongy mesophyll in thick leaves of double homozygous IL2-5/IL4-3 lines (Supplemental Fig. S4). Palisade cell height is positively correlated with photosynthetic efficiency (Niinemets et al., 2009; Terashima et al., 2011) and water storage capacity in succulent Crassulacean acid metabolism plants (Nelson et al., 2005). Our data also indicate that the magnitudes of palisade cell elongation, as well as overall leaf thickness, are modulated by environmental inputs (Fig. 2; Supplemental Fig. S2). High light has been shown to mediate increased leaf thickness (Poorter et al., 2009; Wuyts et al., 2012; Kalve et al., 2014) as well as specifically palisade cell elongation (Kozuka et al., 2011) in Arabidopsis, while elongated palisade cells promote a more efficient distribution of direct light throughout the photosynthetic mesophyll compared with shorter cells (Brodersen et al., 2008; Brodersen and Vogelmann, 2010). Thus, thicker leaves composed of elongated palisade cells may be an adaptation to desert-like dry, direct light environments, whereby the magnitude of these traits is responsive to these environmental cues. Consistent with this hypothesis, we observed that IL2-5 DEG promoters are enriched in motifs that reflect sensitivity to abiotic stimuli, prominently light and water status (Supplemental Fig. S8; Supplemental Data Set S7).
Mechanisms of Cell Enlargement in Thick ILs: Increased Ploidy and Alterations in Cell Cycle-Related Gene Expression
We compared the size of palisade mesophyll cells in leaf cross sections of thick IL2-5, IL4-3, and a homozygous line combining both introgression segments and observed larger palisade cells compared with M82 (Supplemental Fig. S4), suggesting a link between leaf thickness and cell size in tomato. Furthermore, we showed significantly higher ploidy levels in the leaves of these lines relative to the domesticated parent (Fig. 4C), indicating that increased endoreduplication may underpin larger cells and, ultimately, thicker leaves. A partially overlapping series of cell division, cell expansion, and cell differentiation events underlie leaf development (Efroni et al., 2008). These processes are tightly coordinated to buffer perturbations in overall organ shape and size (Beemster et al., 2003; Tsukaya, 2003). Thus, the relative timing and duration of any of these events can impact leaf size and morphology. Additionally, different tissue types in the leaf can have distinct schedules of cellular events during leaf ontogeny; for example, in Arabidopsis, palisade mesophyll cells have a shorter window of cell division compared with epidermal cells, and thus an earlier onset of cell expansion and endoreduplication, resulting in differences in cell volumes and geometry (Wuyts et al., 2012; Kalve et al., 2014). Given the prominent contribution of specific cell types to leaf thickness (palisade mesophyll cells in IL2-5, for example, versus both palisade and spongy mesophyll cells in IL4-3; Supplemental Fig. S4), kinematic studies to capture the timing and extent of tissue-specific cell division and endoreduplication are needed to fully address the dynamic cellular basis of leaf thickness patterning. The observed increase in P3 organ volume and thickness in IL4-3 and, to a lesser extent, IL2-5 relative to M82 (Fig. 4D) support the notion that differences in the trajectory of cellular events during early leaf ontogeny may underpin leaf thickness.
Comparative gene expression profiles of early leaf ontogeny in IL2-5 and IL4-3 show evidence of S. pennellii-like alterations in cell proliferative activity in these thick ILs. Specifically, among a small set of shared differentially expressed genes, the GRAS-domain TF GRAS 08g (Solyc08g014030) is up-regulated at P2 in both lines (Fig. 6A; Supplemental Data Set S5). This gene is closely related to the Arabidopsis gene encoding SHR (Huang et al., 2015), which together with another GRAS-domain TF, SCARECROW (SCR), regulates the duration of cell proliferation in leaves (Dhondt et al., 2010). Moreover, consistent with previous reports, IL2-5 and IL4-3 DEGs are enriched for E2F binding site motifs (Supplemental Data Set S7; Ranjan et al., 2016). E2F TFs act downstream of SHR and SCR to regulate progression through the S-phase of the cell cycle (Dhondt et al., 2010). These data support the notion that the extent and/or duration of cell proliferation underpin increased thickness in these lines. Another set of DEGs that distinguish the thick ILs and the S. pennellii parent from domesticated tomato include three genes with predicted functions in regulating the cell cycle and cell expansion activities: a LIM-domain protein (Solyc04g077780), a JAZ1 TF (Solyc12g009220), and a GA 20-ox (Solyc03g006880; Fig. 6). LIM-domain proteins have been implicated in a variety of functions, including regulation of the cell cycle and organ size in Arabidopsis (Li et al., 2008). GA 20-ox encodes a key GA biosynthetic enzyme, which acts to promote cell elongation (Hisamatsu et al., 2005; de Lucas et al., 2008) and, thus, determinacy during leaf morphogenesis of compound leaves, such as those of tomato (Hay et al., 2002). Moreover, JAZ proteins act as transcriptional repressors and are a central hub in the signaling circuit that integrates environmental cues, such as light quality, to balance growth and defense (for review, see Hou et al., 2013). Finally, it is noteworthy that abiotic cues such as light quality and abscisic acid have been shown to interact and modulate the activity of GA 20-ox and JAZ, and the Arabidopsis LIM-domain protein DA1, respectively, thereby establishing a conceptual means of environmental regulation of leaf thickness patterning. Taken together with higher endopolyploidy levels, the shared expression patterns for these genes between both thick ILs and the S. pennellii parent suggest that leaf thickness may be modulated by changes in the trajectory of cellular events during leaf ontogeny, specifically, the duration of cell proliferation, and the timing and extent of cell expansion. Further validation of this hypothesis is necessary to evaluate the contribution of dynamic, environmentally responsive changes in cell division and endoreduplication to leaf thickness.
Gene Expression Networks Point to Distinct Leaf Ontogeny in IL2-5 and IL4-3
Since we observed a set of shared DEGs in lines IL2-5 and IL4-3, we hypothesized that general patterns of leaf ontogeny also may be shared between these lines, suggesting a core shared trajectory of leaf thickness patterning. However, we found that dynamic Bayesian networks of gene coexpression in IL2-5 and IL4-3 are largely distinct (Fig. 7, A and D; Supplemental Data Sets S8 and S9).
For example, central to the organ-specific network of IL2-5 is an SBP-box domain gene, SBP 04g (Solyc04g064470), which is highly expressed throughout leaf development in IL2-5 (Fig. 7, A and B; Supplemental Fig. S6). SBP TFs regulate various aspects of plant growth by controlling the rate and timing of developmental events, including leaf initiation rate (for review, see Preston and Hileman, 2013). Furthermore, the promoters of IL2-5 DEGs are enriched for SBP motifs (Supplemental Data Set S7), supporting the central role of this group of TFs during IL2-5 leaf ontogeny. Interestingly, GO terms for photosynthesis and translation are enriched among P4 up-regulated genes. This observation suggests that processes associated with cell differentiation (i.e. photosynthetic gene function and protein translation) are precociously activated in IL2-5 relative to domesticated tomato and supports a hypothesis whereby the overall schedule of leaf developmental events may be hastened in IL2-5.
In contrast, a central node in the IL4-3 coexpression network is a BEL1-like 04g (Solyc04g080780). BEL1-like homeodomain proteins interact with class I KNOX TFs to pattern the SAM and lateral organs, including leaf complexity (Kimura et al., 2008; Hay and Tsiantis, 2010) and the extent of lobing and serrations (Kumar et al., 2007). Like S. pennellii, IL4-3 leaflets have significantly smoother margins (fewer serrations) than M82, as reflected in increased circularity (Fig. 4B; Holtan and Hake, 2003; Chitwood et al., 2013).
These distinct dynamic patterns of leaf ontogeny that each IL shares with the desert-adapted parent may reflect aspects of leaf development unrelated to the patterning of leaf thickness, such as the patterning of leaf complexity and leaflet shape in IL4-3. Alternatively, it is also possible that the core mechanism of leaf thickness patterning is achieved by regulation of the timing and extent of cellular activities, such as the balance between cell proliferation and the onset of cell expansion and endoreduplication, with a number of potential molecular networks needed to accomplish these roles. An observation supporting this model is the fact that IL2-5 and IL4-3 have nonoverlapping sets of cell cycle-related DEGs. This hypothesis is consistent with the additive phenotypes of IL2-5/IL4-3 double homozygotes (Fig. 4; Supplemental Fig. S4), whereby IL-specific regulators may converge and act additively to modulate the expression of a common set of targets (such as GA 20-ox and SHR-like) that regulate cell size and shape and, eventually, leaf thickness. Extending these analyses to a broader set of thick lines will yield further insight into the validity of this hypothesis.
CONCLUSION
Leaf thickness is a functional trait associated with the ability of plants to occupy arid environments. Our understanding of how this trait is patterned is poor, due in part to challenges in measuring leaf thickness in a precise and high-throughput manner. Here, we combine a novel tool, a custom-built confocal profilometer designed to measure leaf thickness directly and efficiently, with anatomical and transcriptomic observations across a panel of S. pennellii ILs to assemble a broad and comprehensive framework of the genetic architecture, anatomical manifestations, and molecular patterning processes associated with thick leaves in tomato. We find a complex genetic basis for this trait and a prominent role for environmental cues in modulating it. Thick leaves are associated with elongation of the palisade parenchyma and an increase in leaf ploidy, suggesting that increased cell size underpins thick leaves in tomato. Finally, transcriptomic data show that relative gene expression trends for several putative regulators of cell elongation and meristem determinacy are shared among two thick ILs and their desert tomato parent, implicating these processes as contributors to leaf thickness pattering during development. Ultimately, given the complex genetic architecture of this trait, its responsiveness to environmental factors, and its dependence on plant age, further experiments comparing more thick lines, working with smaller introgression segments to reduce the contribution of epistasis, and explicitly addressing the roles of environment and age, are necessary to understand the mechanistic basis of leaf thickness. This work establishes the necessary foundation to further dissect this highly functional developmentally patterned trait.
MATERIALS AND METHODS
Plant Material and Growth Conditions
Seeds for 76 Solanum pennellii ILs (LA4028–LA4103; Eshed and Zamir, 1995) and the Solanum lycopersicum domesticated variety M82 (LA3475) were obtained either from Dr. Neelima Sinha (University of California, Davis) or from the Tomato Genetics Resource Center (University of California, Davis). All seeds were treated with 50% (v/v) bleach for 3 min, rinsed with water, and germinated in Phytatrays (P1552; Sigma-Aldrich). Seeds were left in the dark for 3 d, followed by 3 d in light, and finally transferred to greenhouse conditions in 50-plug trays. Hardened plants were transplanted to field conditions at the Bradford Research Station in Columbia, Missouri (May 21, 2014) with 3 m between rows and about 1 m spacing between plants within rows. A nonexperimental M82 plant was placed at both ends of each row, and an entire row was placed at each end of the field to reduce border effects on experimental plants. The final design had 15 blocks, each consisting of four rows with 20 plants per row. Each of the 76 ILs and two experimental M82 plants were randomized within each block. IL6-2 was excluded from final analyses due to seed stock contamination. For the analysis of leaf primordia by confocal microscopy and RNA-Seq, IL2-5, IL4-3, M82, and S. pennellii seeds were germinated as above and transferred to pots in controlled growth chamber conditions: irradiance at 400 μmol m−2 s−1, 23°C, and 14-h days. Growth conditions for the drought phenotyping experiment were irradiance of 200 μmol m−2 s−1 at a daytime temperature of 22°C and 18°C at night.
Whole-Plant Phenotyping under Drought
The LemnaTec Scanalyzer plant phenotyping facility at the Donald Danforth Plant Science Center (LemnaTec) was used to phenotype approximately 3-week-old S. lycopersicum and S. pennellii plants (n = 8 per genotype) subjected to one of three watering regimes: 40% field capacity, 20% field capacity, and no watering (0% field capacity). Top-view images of each plant taken every second night over 16 nights were analyzed using custom pipelines in Lemna Launcher (LemnaTec software) to extract total plant pixel area (a proxy for biomass).
Trait Measurements
After flowering (July 2014), four fully expanded adult leaves were harvested from each plant; the adaxial (upper) surfaces of distal lateral leaflets harvested from the left side of the rachis were scanned with a flatbed scanner to obtain raw JPG files. The middle portion of each leaflet was then attached on a custom-build dual confocal profilometer device (Supplemental Fig. S1), and the thickness of each leaflet was measured across the leaflet surface at a resolution of 1 mm2. Median thickness was calculated across each leaflet using values in the range 0 to 2 mm, and these median values were averaged across four leaflets per plant to arrive at a single robust metric of leaf thickness. Finally, entire leaflets were dried and their dry mass was used to calculate LMA for each leaflet. Leaflet outline scans were processed using custom macros in ImageJ (Abràmoff et al., 2004) to segment individual leaflets and to threshold and binarize each leaflet image. Shape descriptors area, aspect ratio, roundness, circularity, and solidity (described in detail by Chitwood et al., 2013) were extracted from binary images. Additionally, elliptical Fourier descriptors for leaflet outlines were determined using SHAPE (Iwata and Ukai, 2002). For this analysis, 20 harmonics with four coefficients each were used to derive principal components that describe major trends in the shape data.
Elemental Profiling (Ionomics)
Distal lateral leaflets of fully expanded young and old leaves of the same plants as above were collected from five individuals of each genotype. Whole leaflets were weighed and digested in nitric acid at 100°C for 3 h. Elemental concentrations were measured using an inductively coupled plasma mass spectrometer (Elan DRC-e; Perkin Elmer) following the procedure described by Ziegler et al. (2013). Instrument-reported concentrations were corrected for losses during sample preparation and changes in instrument response during analysis using yttrium and indium internal standards and a matrix-matched control run every 10th sample. Final concentrations were normalized to sample weight and reported in milligrams of analyte per kilogram of tissue.
Statistical Analyses and Data Visualization
All statistical analyses and visualization were carried out using R packages (R Core Team, 2013). QTL were identified using the mixed-effect linear model packages lme4 (Bates et al., 2014) and lmerTest (Kuznetsova et al., 2015) with M82 as intercept, IL genotype as a fixed effect, and field position attributes (block, row, and column) as random effects. Only effects with significant variance (P < 0.05) were included in the final models. For elemental composition data, leaf age (young and old) also was included as a random effect unless the variance due to age was the greatest source of variance; in such cases, young and old samples were analyzed separately. Heritability values represent the relative proportion of variance due to genotype. For the quantification of organ volume parameters and photosynthesis measurements, linear models were used to test the effect of genotype. All plots were generated with the package ggplot2 (Wickham, 2009).
Trait Correlations and Hierarchical Clustering
For trait correlation analyses, we included all traits reported in this study and measured on the same set of field-grown IL individuals (leaf thickness, LMA, leaflet shape traits, and elemental profiles). We also included several sets of meta-data detailed in Supplemental Data Set S3, including developmental, morphological, fruit pericarp metabolite content, enzyme activity, and seed metabolite content traits (from Chitwood et al., 2013, and refs. therein). Spearman correlation coefficients (rho) were calculated between each pair of traits using the rcorr function in Hmisc (Harrell et al., 2015), and P values for the correlations were corrected for false discovery rate using the Benjamini-Hochberg procedure (Supplemental Data Set S4). Hierarchical clustering and visualization of significant correlations (q < 0.05) of leaf thickness and LMA were clustered (hierarchical ward algorithm) and visualized using pheatmap (Kolde, 2015).
Estimation of Nuclear Size Profiles by Flow Cytometry
Distal lateral leaflets were harvested from the seventh leaf of greenhouse-grown 6-week-old plants and immediately chopped in 1 mL of ice-cold buffer LB01 as described by Doležel et al. (2007). The resulting fine homogenate was filtered through a 30-μm Partec CellTrics filter (5004-004-2326) and incubated with 50 μg mL−1 propidium iodide (Thermo Fisher; P21493) and 50 μg mL−1 RNase A (Qiagen; 19101) for 20 min on ice. Fluorescence scatter data were collected without gating using a BD Acuri CS6 instrument (BD Biosciences). Plots of event count as a function of fluorescence area were used to estimate the proportion of nuclei of sizes corresponding to 2C, 4C, and 8C in each genotype.
Confocal Microscopy, 3D Reconstructions, and Organ Volume Quantification
For mature leaf cross sections, field-grown leaves were fixed in 4% (w/v) formaldehyde, 5% (v/v) glacial acetic acid, and 50% (v/v) ethanol, vacuum infiltrated, dehydrated through an ethanol series, rehydrated to 100% water, stained in 0.002% (w/v) propidium iodide (Thermo Fisher; P21493) for 2 h, dehydrated to 100% ethanol, and finally cleared in 100% methyl salicylate (Sigma; M6752) for 7 d. Hand sections were visualized with a Leica SP8 laser scanning confocal microscope using white light laser excitation at 514 nm with a 20× objective. Two partially overlapping images were captured for each cross section and merged into a single image using the Photomerge function in Adobe Photoshop CC 2014 (Adobe Systems). For the quantification of P3 leaf primordium dimensions, shoot apices (shoot apical meristem and P1–P4) of 14-d-old seedlings grown in controlled conditions were excised, fixed, processed, and stained as detailed for leaf cross sections above. Confocal stacks were obtained at software-optimized intervals and exported as TIFF files. Raw stack files were imported into MorphoGraphX (Barbier de Reuille et al., 2015). After Gaussian filtering, the marching cube surface reconstruction function was used (cube size = 5 μm and threshold = 7,000). The resulting surface mesh was smoothed, subdivided twice, and exported as a PLY file. To minimize the effects of trichomes on P3 volume, all meshes were trimmed in MeshLab (Cignoni et al., 2008). The volume, length, and diameter of processed P3 meshes were calculated using custom scripts in MatLab (MathWorks). Briefly, first, we detected the boundary of each hole and calculated its centroid point. We connected boundary points of each hole to its centroid and filled the triangle faces. After filling all the holes, 3D mesh represents the closed surface. Then, we calculated the volume based on the divergence theorem, which makes use of the fact that the inside fluid expansion equals the flux (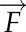 ) of the fluid out of the surface (
) of the fluid out of the surface (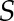 ). When the flux is
). When the flux is 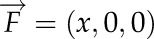 , the volume is
, the volume is 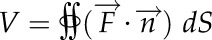 , where
, where 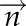 is normal vector. Thus, for each triangle, we computed the normal vector
is normal vector. Thus, for each triangle, we computed the normal vector 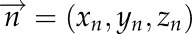 , the area
, the area 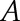 , and the centroid point
, and the centroid point 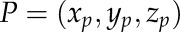 . The volume
. The volume 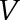 is the summation of
is the summation of 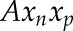 for all triangles. To estimate organ arch length, we made use of the fact that the Laplace-Beltrami eigenfunctions are deformation-invariant shape descriptors (Rustamov, 2007). We thus employed its first eigenfunction, which is associated with the smallest positive eigenvalue, and discretized the eigenfunction values into 50 sets to compute the centroid point to each set. We fit a cubic function by fixing two end-point constraints to those centroid points to get a smooth principal median axis. Note that the two end points were adjusted manually to correct for artifacts. The length of this axis is used to quantify the length of the organ. Finally, we calculated mean organ diameter as
for all triangles. To estimate organ arch length, we made use of the fact that the Laplace-Beltrami eigenfunctions are deformation-invariant shape descriptors (Rustamov, 2007). We thus employed its first eigenfunction, which is associated with the smallest positive eigenvalue, and discretized the eigenfunction values into 50 sets to compute the centroid point to each set. We fit a cubic function by fixing two end-point constraints to those centroid points to get a smooth principal median axis. Note that the two end points were adjusted manually to correct for artifacts. The length of this axis is used to quantify the length of the organ. Finally, we calculated mean organ diameter as
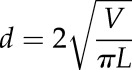 |
RNA-Seq Library Preparation and Sequencing
Apices of 14-d-old IL2-5, IL4-3, M82, and S. pennellii plants grown in a randomized design under controlled growth conditions were hand dissected using a dissecting microscope to separate plastochrons P4, P3, P2, and P1+SAM organs corresponding approximately to leaves L5 to L8. For S. pennellii plants, we were not able to separate P2 primordia from the apex, so we obtained P4, P3, and SAM+P1+P2 samples. Dissected organs were removed from the apex in less than 60 s and immediately fixed in 100% ice-cold acetone to preserve the integrity of RNA in the sample. Each biological replicate is a pool of 10 individuals, and a total of five biological replicates were obtained for each genotype/organ combination. RNA was extracted using the PicoPure RNA Isolation Kit (Thermo Fisher) according to the manufacturer’s protocol with the optional on-column DNase treatment. RNA integrity was assessed by running all samples on an Agilent RNA 6000 Pico chip (Agilent Technologies), and three biological replicates with RNA integrity > 7 were selected for further processing. Double-stranded cDNA amplified using the Clontech SMARTer PCR cDNA synthesis kit (634926; TaKaRa Bio) was fragmented for 15 min using Fragmentase (M0348; New England Biolabs) and processed into Illumina sequencing libraries as follows: the ends of 1.5× AMPure XP bead (A63880; Agencourt)-purified fragmented DNA was repaired with End Repair Enzyme Mix (E6050; New England Biolabs) and Klenow DNA Polymerase (M0210; New England Biolabs), followed by dA tailing using Klenow 3′-5′ exonuclease (M0212; New England Biolabs). The Illumina TruSeq universal adapter dimer was ligated to library fragments with rapid T4 DNA Ligase (L6030-HC-L; Enzymatics) followed by three rounds of 1× AMPure XP bead purification to remove unligated adapter. Finally, libraries were enriched and indexed by PCR using Phusion HiFi Polymerase mix (M0531; New England Biolabs). Illumina libraries were quantified using a nanodrop, pooled to a final concentration of 20 nm, and sequenced as single-end 100-bp reads on Illumina HiSeq2500 at the School of Medicine Genome Technology Access Center, Washington University in St. Louis (https://gtac.wustl.edu/).
RNA-Seq Data Analysis
Adapters and low-quality bases were removed using Trimmomatic (Bolger et al., 2014) with default parameters. Trimmed reads were mapped to the ITAG2.3 S. lycopersicum genome (https://solgenomics.net/organism/Solanum_lycopersicum/genome; Tomato Genome Consortium, 2012) using bowtie2 (Langmead and Salzberg, 2012) to obtain SAM (Sequence Alignment Map) files. After sorting and indexing of SAM files, BAM (Binary SAM) files were generated using samtools commands (Li et al., 2009). The BEDtools multicov tool (Quinlan and Hall, 2010) was then used to obtain read counts per annotated gene for each sample. Subsequent analysis was done with the R package edgeR (Robinson et al., 2010). After normalization for library size, 20,231 genes with at least one count per million reads across three samples were retained for further analysis. Lists of DEGs were generated between pairwise sample combinations with q < 0.05. For IL2-5 and IL4-3 at P1, P3, and P4 stages, we identified genes that are differentially expressed relative to M82 in both the IL and the S. pennellii parent to interrogate S. pennellii-like changes in gene expression in the ILs. For P2, the list of DEGs in each IL reflects changes relative to M82 only (Supplemental Data Set S5).
GO, MapMan, and Promoter Motif Enrichment Analyses
Lists of IL organ-specific DEGs were interrogated for enrichment of GO terms using agriGO (http://bioinfo.cau.edu.cn/agriGO/; Du et al., 2010) with default parameters (Fisher’s exact significance test and Yekutieli false discovery rate adjustment at q < 0.05). We further divided DEG gene lists into IL up-regulated and down-regulated genes and report significant terms in Supplemental Data Set S6. We tested IL organ-specific DEGs for the enrichment of annotated promoter motifs using a custom R script (Julin Maloof, personal communication). Briefly, functions in the Bioconductor Biostrings package (Pages et al., 2009) were implemented to count the frequency of 100 known motifs in the promoters of DEGs (1,000-bp upstream sequence) and calculate P values for enrichment based on these counts. We report exact matches of known motifs and motifs with up to one mismatch in IL up-regulated and down-regulated organ-specific gene sets (Supplemental Data Set S7).
IL Organ-Specific Gene Network Inference
To infer IL organ-specific networks (Fig. 7, A–D; Supplemental Data Set S8), we selected DEGs between IL2-5/M82 (IL4-3/M82) and S. pennellii/M82 for each organ (P1, P3, and P4; q < 0.05). Since coexpression analysis can inform the likelihood that genes interact, or participate in the same functional pathway, the selected genes for each IL (IL2-5 or IL4-3) and each organ were clustered based on their coexpression across genotypes. To perform clustering, the Silhouette index (Rousseeuw, 1987) followed by K-means (MacQueen, 1967) were applied. After clustering, networks were inferred as described by de Luis Balaguer et al. (2017). Briefly, for each DEG, we identified a set of potential regulators and measured the likelihood of gene-target regulation using a Bayesian Dirichlet equivalence uniform (Buntine, 1991). Genes that had the highest value of the Bayesian Dirichlet equivalence uniform were chosen as regulators, and of these, only TFs (as annotated by Suresh et al., 2014) were further considered as regulatory (source) nodes. To obtain the final IL2-5 and IL4-3 organ-specific networks, the networks for each cluster were connected. For this, we found regulations among the cluster hubs (node of each individual network with the largest degree of edges leaving the node) by using the same Bayesian Dirichlet equivalence uniform metric. In addition, we implemented a score to estimate whether the inferred interactions were activations or repressions. The score was calculated for each edge, and it measured the ratio between (1) the conditional probability that a gene is expressed given that its regulator was expressed in the prior time point and (2) the conditional probability that a gene is expressed given that its regulator was not expressed in the prior time point. If the first conditional probability is larger than the second one, then the parent was found to be an activator, and vice versa. In the case of a tie, the edge was found to have an undetermined sign. Networks for each organ were jointly visualized in Cytoscape (Shannon et al., 2003).
Dynamic IL Network Construction
To infer dynamic IL networks (Fig. 7, E and F; Supplemental Data Set S9), we selected DEGs between IL2-5/M82 or IL4-3/M82 and S. pennellii/M82 for each organ (P1, P3, and P4; q < 0.05 or fold change > 2 and q < 0.2). All DEGs in IL2-5 or IL4-3 were clustered in four groups, corresponding to the four developmental stages: each gene was assigned to the developmental stage where it showed the maximum expression. A network was then inferred for each developmental stage as described for the IL organ-specific networks. To ensure that all potential regulators of each gene were considered, genes from the preceding developmental stage were included in the inference of the network of each developmental stage. The final network for each IL was visualized in Cytoscape (Shannon et al., 2003).
Accession Numbers
RNA-Seq data have been deposited in the National Center for Biotechnology Information Short Read Archive under BioProject identifier PRJNA396585 containing 45 BioSamples.
Supplemental Data
The following supplemental materials are available.
Supplemental Figure S1. Dual confocal profilometer device used to measure leaf thickness.
Supplemental Figure S2. Comparison of leaf thickness of select ILs as a function of shoot position and field versus greenhouse conditions.
Supplemental Figure S3. Representative leaf cross sections and flow cytometry of leaf 6/7 for 10 ILs harboring leaf thickness QTLs grown in greenhouse conditions.
Supplemental Figure S4. Mean dimensions of palisade and spongy mesophyll cell layers in select thick leaf ILs and representative flow cytometry histograms of leaf 7 and post-flowering leaves from each genotype.
Supplemental Figure S5. Representative shoot apex reconstructions highlighting the appearance of early- and late-stage leaf primordia for each genotype in Figure 4.
Supplemental Figure S6. Summary of differentially expressed genes in IL2-5 and IL4-3.
Supplemental Figure S7. Expression profiles of differentially expressed putative TFs in IL2-5 and IL4-3.
Supplemental Figure S8. Summary of enriched promoter motifs among differentially expressed genes in IL2-5 and IL4-3.
Supplemental Data Set S1. Trait value estimates and heritability for leaf thickness, LMA, and leaflet shape across the IL panel.
Supplemental Data Set S2. Trait value estimates and heritability for elemental concentration across the IL panel.
Supplemental Data Set S3. Summary of all measured and meta-data traits used in the correlation matrix.
Supplemental Data Set S4. Pairwise trait correlation matrix including significance values.
Supplemental Data Set S5. List of differentially expressed genes (q < 0.05) in each organ (P1–P4) for the comparison M82/IL overlapping with M82/S. pennellii.
Supplemental Data Set S6. List of significantly enriched (q < 0.05) GO terms for gene sets listed in Supplemental Data Set S5.
Supplemental Data Set S7. List of enriched (q < 0.05) promoter motifs for gene sets listed in Supplemental Data Set S5.
Supplemental Data Set S8. List of organ-specific (P1, P3, P4) gene interactions for IL2-5 and IL4-3.
Supplemental Data Set S9. List of dynamic gene interactions for IL2-5 and IL4-3.
Acknowledgments
S. pennellii IL panel seeds were provided by Dr. Neelima Sinha (University of California, Davis) and the Tomato Genetics Resource Center (University of California, Davis). We thank Dr. Ivan Baxter and Dr. Greg Ziegler (Donald Danforth Plant Science Center) for generating elemental profile data and Dr. Julin Maloof (University of California, Davis) for sharing custom promoter enrichment analysis scripts. We acknowledge the advice and assistance of Dr. Noah Fahlgren, Dr. Malia Gehan, and Melinda Darnell (Donald Danforth Plant Science Center) with drought phenotyping experiments. We thank Dr. Elizabeth Kellogg (Donald Danforth Plant Science Center) for insightful discussions and comments on the article.
Glossary
- LMA
leaf mass per area
- QTL
quantitative trait loci
- IL
introgression line
- RNA-Seq
RNA sequencing
- SAM
shoot apical meristem
- DEG
differentially expressed gene
- TF
transcription factor
- GO
Gene Ontology
Footnotes
This work was supported by funds from the Donald Danforth Plant Science Center. R.S. is supported by an NSF CAREER grant (MCB-1453130). M.H.F. is supported by an NSF NPGI postdoctoral fellowship (IOS-1523668).
Articles can be viewed without a subscription.
References
- Abràmoff MD, Magalhães PJ, Ram SJ (2004) Image processing with ImageJ. Biophoton Int 11: 36–42 [Google Scholar]
- Bates D, Maechler M, Bolker B, Walker S (2014) lme4: Linear mixed-effects models using Eigen and S4. R package version, 1: 1–23 [Google Scholar]
- Barbier de Reuille P, Routier-Kierzkowska AL, Kierzkowski D, Bassel GW, Schüpbach T, Tauriello G, Bajpai N, Strauss S, Weber A, Kiss A, et al. (2015) MorphoGraphX: a platform for quantifying morphogenesis in 4D. eLife 4: 05864. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Becker B. (2007) Function and evolution of the vacuolar compartment in green algae and land plants (Viridiplantae). Int Rev Cytol 264: 1–24 [DOI] [PubMed] [Google Scholar]
- Beemster GT, Fiorani F, Inzé D (2003) Cell cycle: the key to plant growth control? Trends Plant Sci 8: 154–158 [DOI] [PubMed] [Google Scholar]
- Bohmert K, Camus I, Bellini C, Bouchez D, Caboche M, Benning C (1998) AGO1 defines a novel locus of Arabidopsis controlling leaf development. EMBO J 17: 170–180 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bolger AM, Lohse M, Usadel B (2014) Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 30: 2114–2120 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brodersen CR, Vogelmann TC (2010) Do changes in light direction affect absorption profiles in leaves? Funct Plant Biol 37: 403–412 [Google Scholar]
- Brodersen CR, Vogelmann TC, Williams WE, Gorton HL (2008) A new paradigm in leaf-level photosynthesis: direct and diffuse lights are not equal. Plant Cell Environ 31: 159–164 [DOI] [PubMed] [Google Scholar]
- Buntine W. (1991) Theory refinement on Bayesian networks. In BD D'Ambrosio, P Smets, PP Bonissone, eds, Proceedings of the Seventh Conference on Uncertainty in Artificial Intelligence. Morgan Kaufmann Publishers, Los Angeles, CA, pp 52–60 [Google Scholar]
- Chitwood DH, Kumar R, Headland LR, Ranjan A, Covington MF, Ichihashi Y, Fulop D, Jiménez-Gómez JM, Peng J, Maloof JN, et al. (2013) A quantitative genetic basis for leaf morphology in a set of precisely defined tomato introgression lines. Plant Cell 25: 2465–2481 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cignoni P, Callieri M, Corsini M, Dellepiane M, Ganovelli F, Ranzuglia G (2008) Meshlab: an open-source mesh processing tool. In V Scarano, R De Chiara, U Erra, eds, Eurographics Italian Chapter Conference, 2008. pp 129–136 [Google Scholar]
- de Lucas M, Davière JM, Rodríguez-Falcón M, Pontin M, Iglesias-Pedraz JM, Lorrain S, Fankhauser C, Blázquez MA, Titarenko E, Prat S (2008) A molecular framework for light and gibberellin control of cell elongation. Nature 451: 480–484 [DOI] [PubMed] [Google Scholar]
- de Luis Balaguer MA, Fisher AP, Clark NM, Fernandez-Espinosa MG, Moller B, Weijers D, Lohmann JU, Williams C, Lorenzo O, Sozzani R (2017) An inference approach combines spatial and temporal gene expression data to predict gene regulatory networks in Arabidopsis stem cells. PNAS https://doi.org/10.1101/140269 [DOI] [PMC free article] [PubMed]
- Dhondt S, Coppens F, De Winter F, Swarup K, Merks RM, Inzé D, Bennett MJ, Beemster GT (2010) SHORT-ROOT and SCARECROW regulate leaf growth in Arabidopsis by stimulating S-phase progression of the cell cycle. Plant Physiol 154: 1183–1195 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doležel J, Greilhuber J, Suda J (2007) Estimation of nuclear DNA content in plants using flow cytometry. Nat Protoc 2: 2233–2244 [DOI] [PubMed] [Google Scholar]
- Du Z, Zhou X, Ling Y, Zhang Z, Su Z (2010) agriGO: a GO analysis toolkit for the agricultural community. Nucleic Acids Res 38: W64–W70 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Efroni I, Blum E, Goldshmidt A, Eshed Y (2008) A protracted and dynamic maturation schedule underlies Arabidopsis leaf development. Plant Cell 20: 2293–2306 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eshed Y, Zamir D (1995) An introgression line population of Lycopersicon pennellii in the cultivated tomato enables the identification and fine mapping of yield-associated QTL. Genetics 141: 1147–1162 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fridman E, Carrari F, Liu YS, Fernie AR, Zamir D (2004) Zooming in on a quantitative trait for tomato yield using interspecific introgressions. Science 305: 1786–1789 [DOI] [PubMed] [Google Scholar]
- Garnier E, Laurent G (1994) Leaf anatomy, specific mass and water content in congeneric annual and perennial grass species. New Phytol 128: 725–736 [Google Scholar]
- Gibson AC. (1982) The anatomy of succulence. In IP Ting, M Gibbs, eds, Crassulacean Acid Metabolism. American Society of Plant Physiologists, Rockville, MD, pp 1–17 [Google Scholar]
- Haliński ŁP, Kalkowska M, Kalkowski M, Piorunowska J, Topolewska A, Stepnowski P (2015) Cuticular wax variation in the tomato (Solanum lycopersicum L.), related wild species and their interspecific hybrids. Biochem Syst Ecol 60: 215–224 [Google Scholar]
- Harrell, FE Jr, Dupont, C, et al (2015). Hmisc: Harrell Miscellaneous. R package version 3.16-0. http://CRAN.R-project.org/package=Hmisc [Google Scholar]
- Hay A, Kaur H, Phillips A, Hedden P, Hake S, Tsiantis M (2002) The gibberellin pathway mediates KNOTTED1-type homeobox function in plants with different body plans. Curr Biol 12: 1557–1565 [DOI] [PubMed] [Google Scholar]
- Hay A, Tsiantis M (2010) KNOX genes: versatile regulators of plant development and diversity. Development 137: 3153–3165 [DOI] [PubMed] [Google Scholar]
- Hisamatsu T, King RW, Helliwell CA, Koshioka M (2005) The involvement of gibberellin 20-oxidase genes in phytochrome-regulated petiole elongation of Arabidopsis. Plant Physiol 138: 1106–1116 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holtan HE, Hake S (2003) Quantitative trait locus analysis of leaf dissection in tomato using Lycopersicon pennellii segmental introgression lines. Genetics 165: 1541–1550 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hou X, Ding L, Yu H (2013) Crosstalk between GA and JA signaling mediates plant growth and defense. Plant Cell Rep 32: 1067–1074 [DOI] [PubMed] [Google Scholar]
- Huang W, Xian Z, Kang X, Tang N, Li Z (2015) Genome-wide identification, phylogeny and expression analysis of GRAS gene family in tomato. BMC Plant Biol 15: 209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ichihashi Y, Aguilar-Martínez JA, Farhi M, Chitwood DH, Kumar R, Millon LV, Peng J, Maloof JN, Sinha NR (2014) Evolutionary developmental transcriptomics reveals a gene network module regulating interspecific diversity in plant leaf shape. Proc Natl Acad Sci USA 111: E2616–E2621 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iwata H, Ukai Y (2002) SHAPE: a computer program package for quantitative evaluation of biological shapes based on elliptic Fourier descriptors. J Hered 93: 384–385 [DOI] [PubMed] [Google Scholar]
- John GP, Scoffoni C, Sack L (2013) Allometry of cells and tissues within leaves. Am J Bot 100: 1936–1948 [DOI] [PubMed] [Google Scholar]
- Kalve S, Fotschki J, Beeckman T, Vissenberg K, Beemster GT (2014) Three-dimensional patterns of cell division and expansion throughout the development of Arabidopsis thaliana leaves. J Exp Bot 65: 6385–6397 [DOI] [PubMed] [Google Scholar]
- Kimura S, Koenig D, Kang J, Yoong FY, Sinha N (2008) Natural variation in leaf morphology results from mutation of a novel KNOX gene. Curr Biol 18: 672–677 [DOI] [PubMed] [Google Scholar]
- Koenig D, Jiménez-Gómez JM, Kimura S, Fulop D, Chitwood DH, Headland LR, Kumar R, Covington MF, Devisetty UK, Tat AV, et al. (2013) Comparative transcriptomics reveals patterns of selection in domesticated and wild tomato. Proc Natl Acad Sci USA 110: E2655–E2662 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kolde R. (2015) pheatmap: Pretty Heatmaps. R package version 1.0.2
- Kozuka T, Kong SG, Doi M, Shimazaki K, Nagatani A (2011) Tissue-autonomous promotion of palisade cell development by phototropin 2 in Arabidopsis. Plant Cell 23: 3684–3695 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kumar R, Kushalappa K, Godt D, Pidkowich MS, Pastorelli S, Hepworth SR, Haughn GW (2007) The Arabidopsis BEL1-LIKE HOMEODOMAIN proteins SAW1 and SAW2 act redundantly to regulate KNOX expression spatially in leaf margins. Plant Cell 19: 2719–2735 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuznetsova A, Christensen RH, Bavay C, Brockhoff PB (2015) Automated mixed ANOVA modeling of sensory and consumer data. Food Qual Prefer 40: 31–38 [Google Scholar]
- Langmead B, Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2. Nat Methods 9: 357–359 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li H, Handsaker B, Wysoker A, Fennell T, Ruan J, Homer N, Marth G, Abecasis G, Durbin R (2009) The sequence alignment/map format and SAMtools. Bioinformatics 25: 2078–2079 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li Y, Zheng L, Corke F, Smith C, Bevan MW (2008) Control of final seed and organ size by the DA1 gene family in Arabidopsis thaliana. Genes Dev 22: 1331–1336 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lightfoot DA, Mungur R, Ameziane R, Nolte S, Long L, Bernhard K, Colter A, Jones K, Iqbal MJ, Varsa A, et al. (2007) Improved drought tolerance of transgenic Zea mays plants that express the glutamate dehydrogenase gene (gdhA) of E. coli. Euphytica 156: 103–116 [Google Scholar]
- MacQueen J. (1967) Some methods for classification and analysis of multivariate observations. In LM Le Cam, J Neyman, eds, Proceedings of the Fifth Berkeley Symposium on Mathematical Statistics and Probability. University of California Press, Berkeley, CA, pp 281–297 [Google Scholar]
- McDowell ET, Kapteyn J, Schmidt A, Li C, Kang JH, Descour A, Shi F, Larson M, Schilmiller A, An L, et al. (2011) Comparative functional genomic analysis of Solanum glandular trichome types. Plant Physiol 155: 524–539 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McHale NA. (1992) A nuclear mutation blocking initiation of the lamina in leaves of Nicotiana sylvestris. Planta 186: 355–360 [DOI] [PubMed] [Google Scholar]
- McHale NA. (1993) LAM-1 and FAT genes control development of the leaf blade in Nicotiana sylvestris. Plant Cell 5: 1029–1038 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Muir CD, Conesa MÀ, Roldán EJ, Molins A, Galmés J (2016) Weak coordination between leaf structure and function among closely related tomato species. New Phytol 213: 1642–1653 [DOI] [PubMed] [Google Scholar]
- Muir CD, Pease JB, Moyle LC (2014) Quantitative genetic analysis indicates natural selection on leaf phenotypes across wild tomato species (Solanum sect. Lycopersicon; Solanaceae). Genetics 198: 1629–1643 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakazato T, Warren DL, Moyle LC (2010) Ecological and geographic modes of species divergence in wild tomatoes. Am J Bot 97: 680–693 [DOI] [PubMed] [Google Scholar]
- Nelson EA, Sage TL, Sage RF (2005) Functional leaf anatomy of plants with Crassulacean acid metabolism. Funct Plant Biol 32: 409–419 [DOI] [PubMed] [Google Scholar]
- Nicotra AB, Leigh A, Boyce CK, Jones CS, Niklas KJ, Royer DL, Tsukaya H (2011) The evolution and functional significance of leaf shape in the angiosperms. Funct Plant Biol 38: 535–552 [DOI] [PubMed] [Google Scholar]
- Niinemets U, Díaz-Espejo A, Flexas J, Galmés J, Warren CR (2009) Role of mesophyll diffusion conductance in constraining potential photosynthetic productivity in the field. J Exp Bot 60: 2249–2270 [DOI] [PubMed] [Google Scholar]
- Ogburn R, Edwards EJ (2010) The ecological water-use strategies of succulent plants. Adv Bot Res 55: 179–225 [Google Scholar]
- Oguchi R, Hikosaka K, Hirose T (2005) Leaf anatomy as a constraint for photosynthetic acclimation: differential responses in leaf anatomy to increasing growth irradiance among three deciduous trees. Plant Cell Environ 28: 916–927 [Google Scholar]
- Pages H, Aboyoun P, Gentleman R, DebRoy S (2009) String objects representing biological sequences, and matching algorithms. R package version 2(2)
- Pieruschka R, Poorter H (2012) Phenotyping plants: genes, phenes and machines. Funct Plant Biol 39: 813–820 [DOI] [PubMed] [Google Scholar]
- Poorter H, Niinemets U, Poorter L, Wright IJ, Villar R (2009) Causes and consequences of variation in leaf mass per area (LMA): a meta-analysis. New Phytol 182: 565–588 [DOI] [PubMed] [Google Scholar]
- Preston JC, Hileman LC (2013) Functional evolution in the plant SQUAMOSA-PROMOTER BINDING PROTEIN-LIKE (SPL) gene family. Front Plant Sci 4: 80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Quinlan AR, Hall IM (2010) BEDTools: a flexible suite of utilities for comparing genomic features. Bioinformatics 26: 841–842 [DOI] [PMC free article] [PubMed] [Google Scholar]
- R Core Team (2013) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. http://www.R-project.org/
- Ranjan A, Budke JM, Rowland SD, Chitwood DH, Kumar R, Carriedo L, Ichihashi Y, Zumstein K, Maloof JN, Sinha NR (2016) eQTL regulating transcript levels associated with diverse biological processes in tomato. Plant Physiol 172: 328–340 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robinson MD, McCarthy DJ, Smyth GK (2010) edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 26: 139–140 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roderick ML, Berry SL, Noble IR, Farquhar GD (1999) A theoretical approach to linking the composition and morphology with the function of leaves. Funct Ecol 13: 683–695 [Google Scholar]
- Rousseeuw PJ. (1987) Silhouettes: a graphical aid to the interpretation and validation of cluster analysis. J Comput Appl Math 20: 53–65 [Google Scholar]
- Rustamov RM. (2007) Laplace-Beltrami eigenfunctions for deformation invariant shape representation. In Proceedings of the Fifth Eurographics Symposium on Geometry Processing. Eurographics Association, Barcelona, Spain, pp 225–233 [Google Scholar]
- Sack L, Frole K (2006) Leaf structural diversity is related to hydraulic capacity in tropical rain forest trees. Ecology 87: 483–491 [DOI] [PubMed] [Google Scholar]
- Schauer N, Semel Y, Roessner U, Gur A, Balbo I, Carrari F, Pleban T, Perez-Melis A, Bruedigam C, Kopka J, et al. (2006) Comprehensive metabolic profiling and phenotyping of interspecific introgression lines for tomato improvement. Nat Biotechnol 24: 447–454 [DOI] [PubMed] [Google Scholar]
- Semel Y, Nissenbaum J, Menda N, Zinder M, Krieger U, Issman N, Pleban T, Lippman Z, Gur A, Zamir D (2006) Overdominant quantitative trait loci for yield and fitness in tomato. Proc Natl Acad Sci USA 103: 12981–12986 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shannon P, Markiel A, Ozier O, Baliga NS, Wang JT, Ramage D, Amin N, Schwikowski B, Ideker T (2003) Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res 13: 2498–2504 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith SD, Monson RK, Anderson JE (1997) Plant processes and responses to stress. In Physiological Ecology of North American Desert Plants. Springer, Berlin, pp 45–71 [Google Scholar]
- Suresh BV, Roy R, Sahu K, Misra G, Chattopadhyay D (2014) Tomato genomic resources database: an integrated repository of useful tomato genomic information for basic and applied research. PLoS ONE 9: e86387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taji T, Ohsumi C, Iuchi S, Seki M, Kasuga M, Kobayashi M, Yamaguchi-Shinozaki K, Shinozaki K (2002) Important roles of drought- and cold-inducible genes for galactinol synthase in stress tolerance in Arabidopsis thaliana. Plant J 29: 417–426 [DOI] [PubMed] [Google Scholar]
- Terashima I, Hanba YT, Tholen D, Niinemets Ü (2011) Leaf functional anatomy in relation to photosynthesis. Plant Physiol 155: 108–116 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tomato Genome Consortium (2012) The tomato genome sequence provides insights into fleshy fruit evolution. Nature 485: 635–641 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsuge T, Tsukaya H, Uchimiya H (1996) Two independent and polarized processes of cell elongation regulate leaf blade expansion in Arabidopsis thaliana (L.) Heynh. Development 122: 1589–1600 [DOI] [PubMed] [Google Scholar]
- Tsukaya H. (2003) Organ shape and size: a lesson from studies of leaf morphogenesis. Curr Opin Plant Biol 6: 57–62 [DOI] [PubMed] [Google Scholar]
- von Willert DJ. (1992) Life Strategies of Succulents in Deserts: With Special Reference to the Namib Desert. Cambridge University Press, Cambridge, United Kingdom [Google Scholar]
- Wickham H. (2009) ggplot2: Elegant Graphics for Data Analysis. Springer, New York [Google Scholar]
- Wright IJ, Reich PB, Westoby M, Ackerly DD, Baruch Z, Bongers F, Cavender-Bares J, Chapin T, Cornelissen JH, Diemer M, et al. (2004) The worldwide leaf economics spectrum. Nature 428: 821–827 [DOI] [PubMed] [Google Scholar]
- Wuyts N, Massonnet C, Dauzat M, Granier C (2012) Structural assessment of the impact of environmental constraints on Arabidopsis thaliana leaf growth: a 3D approach. Plant Cell Environ 35: 1631–1646 [DOI] [PubMed] [Google Scholar]
- Ziegler G, Terauchi A, Becker A, Armstrong P, Hudson K, Baxter I (2013) Ionomic screening of field-grown soybean identifies mutants with altered seed elemental composition. Plant Genome 6: http://doi.org/10.3835/plantgenome2012.07.0012 [Google Scholar]