


Abstract
In the budding yeast Saccharomyces cerevisiae the Srs2/RadH DNA helicase promotes survival after ultraviolet (UV) irradiation, and has been implicated in DNA repair, recombination and checkpoint signalling following DNA damage. A second helicase, Sgs1, is the S.cerevisiae homologue of the human BLM and WRN proteins, which are defective in cancer predisposition and/or premature ageing syndromes. Saccharomyces cerevisiae cells lacking both Srs2 and Sgs1 exhibit a severe growth defect. We have identified an Srs2 orthologue in the fission yeast Schizosaccharomyces pombe, and have investigated its role in responses to UV irradiation and inhibition of DNA replication. Deletion of fission yeast srs2 caused spontaneous hyper-recombination and UV sensitivity, and simultaneous deletion of the SGS1 homologue rqh1 caused a severe growth defect reminiscent of that seen in the equivalent S.cerevisiae mutant. However, unlike in budding yeast, inactivation of the homologous recombination pathway did not suppress this growth defect. Indeed, the homologous recombination pathway was required for maintenance of normal fission yeast viability in the absence of Srs2, and loss of homologous recombination and loss of Srs2 contributed additively to UV sensitivity. We conclude that Srs2 plays related, but not identical, roles in the two yeast species.
INTRODUCTION
Eukaryotic cells engage a variety of protective responses following DNA damage. These include the activation of DNA repair and intracellular checkpoint signalling pathways, which serve to delay cell cycle progression while DNA damage persists. Genetic strategies, principally using the budding yeast Saccharomyces cerevisiae and the distantly related fission yeast Schizosaccharomyces pombe, have identified a large number of radiation sensitive (rad) mutants that are defective in one or more of these protective pathways. Such studies have provided an increasingly detailed view of the roles of the corresponding Rad proteins in DNA damage responses. Many of the rad gene products are required for DNA repair processes, specifically nucleotide excision repair (NER), recombinational repair, non-homologous end joining (NHEJ) and post-replication gap-filling. Other Rad proteins have primary roles in checkpoint responses. The DNA damage checkpoint components partially overlap with those of the S-M checkpoint, which delays progression into or through mitosis when chromosomal DNA is not fully replicated. The extent of this overlap is clearest in S.pombe where, following either DNA damage or inhibition of replication, a common set of ‘checkpoint Rad’ proteins transduce checkpoint signals, which are presumed to be generated following the detection of abnormal DNA structures (1,2). Defects in the checkpoint Rad pathway allow progression into mitosis in the absence of complete DNA replication or in the presence of DNA strand breaks, giving rise to a characteristic ‘cut’ (cell untimely torn) phenotype.
For several gene products the distinction between functions in DNA repair, in checkpoint arrest or in recovery from such arrest is not straightforward. This is the case for the product of the SGS1 gene in S.cerevisiae, and that of rqh1 (also known as hus2/rad12), its homologue in S.pombe. These non-essential genes encode DNA helicases that are of particular interest because their human counterparts BLM and WRN are mutated in Bloom’s and Werner’s syndromes, disorders characterised in part by increased cancer incidence and premature ageing (3). Sgs1 and Rqh1 are required for maintenance of normal levels of resistance to the DNA replication inhibitor hydroxyurea (HU), to UV light and to ionizing radiation (4–7). The radiation sensitivity of sgs1/rqh1 mutants can be interpreted in terms of a role for the corresponding proteins in DNA repair (5,7). In S.pombe this repair pathway appears also to involve the checkpoint Rad proteins, together with their effector, the protein kinase Cds1, and the homologous recombination machinery (5). Sgs1, Rqh1 and BLM are also required to prevent unscheduled mitotic recombination, either in the presence of DNA replication inhibitors or in the absence of any perturbation (4,8,9).
In S.cerevisiae, a second DNA helicase, Srs2, appears to be functionally related to Sgs1. Like sgs1 mutants, cells lacking Srs2 are sensitive to DNA damaging agents and display elevated levels of mitotic recombination (10). Furthermore, simultaneous deletion of SRS2 (also known as RADH) and SGS1 results in a dramatic reduction in cell viability (7,11). This has been attributed to a role for these helicases in DNA replication, a function also recently ascribed to the Xenopus BLM protein (11,12). An alternative, but related, view is that simultaneous loss of Sgs1 and Srs2 results in lethal levels of aberrant recombination. This view is supported by the observation that inactivation of the homologous recombination pathway by deletion of the RAD51 gene substantially suppresses the growth defect seen in sgs1 srs2 strains (7,13).
Although Srs2 is structurally related to bacterial DNA helicases (14), orthologues of S.cerevisiae SRS2 have not so far been described in other eukaryotes. Here, we describe the identification and characterisation of the S.pombe gene most closely related to SRS2. Our data lead us to conclude that this fission yeast gene is required for maintenance of normal levels of resistance to DNA damaging agents, and that in some aspects its function resembles S.cerevisiae SRS2. Nonetheless, genetic interactions between S.pombe srs2 and the recombination pathway differ from those of SRS2 in S.cerevisiae, suggesting significant divergence of activity.
MATERIALS AND METHODS
Database searches
Searches to identify Srs2-related sequences in S.pombe were performed using the Sanger Centre BLAST server (http://www.sanger.ac.uk/Projects/S_pombe/blast_server.shtml). Ψ-BLAST (http://www.ncbi.nlm.nih.gov/cgi-bin/BLAST/nph-psi_blast) searches were used to identify similarities between Srs2 and proteins in the Swiss-Prot database. The protein sequence alignment in Figure 1A was created using the PILEUP program (University of Wisconsin Genetics Computer Group) and displayed using MacBoxshade (written by Michael Baron: michael.baron@bbsrc.ac.uk).
Figure 1.

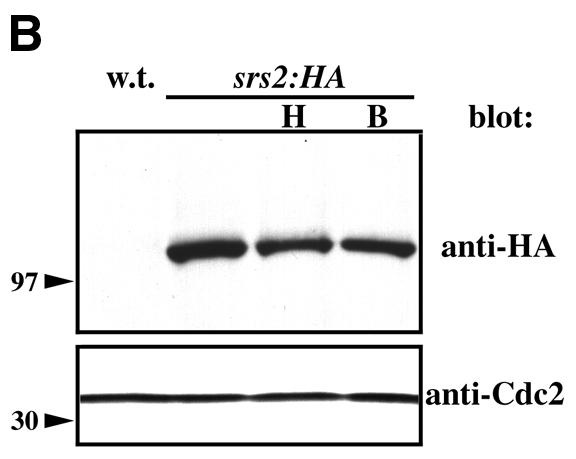
Srs2 is structurally conserved between S.cerevisiae and S.pombe. (A) An alignment of the amino acid sequences of budding yeast Srs2 (Srs2Sc; residues 1–782) and the fission yeast Srs2 protein described here (Srs2Sp; residues 1–698). White text on a black background indicates amino acid identity, black text on a shaded background indicates conservative amino acid substitutions. (B) Whole cell protein extracts were prepared from an untagged control (w.t.) strain or a genomic HA-tagged srs2 strain (srs2:HA; see Materials and Methods) grown to mid-exponential phase, and exposed to 10 mM hydroxyurea (H) for 4 h or 200 µg/ml bleomycin (B) for 2 h. The extracts were separated by SDS–PAGE and subjected to immunoblotting using anti-HA or anti-Cdc2 (loading control) antibodies, as indicated. The positions at which 97 and 30 kDa size markers migrated are also indicated.
Fission yeast strains and methods
Conditions for growth, maintenance and genetic manipulation of fission yeast were as described previously (15). A complete list of the strains used in this study is given in Table 1. Except where otherwise stated, strains were grown at 30°C in YPD or EMM2 medium with appropriate supplements. Where necessary, gene expression from the nmt1 promoter was repressed by the addition of 5 µM thiamine to the growth medium.
Table 1. Schizosaccharomyces pombe strains used in this study.
 |
Construction of the ade6-M26/ade6-L469 stable diploid strain and measurement of mitotic recombination rates were performed exactly as described elsewhere (4).
Gene disruption and related techniques
The one-step disruption method was used, following PCR-mediated generation of the entire ura4+ gene or kanMX cassette flanked by 80 bp segments from the 5′ and 3′ regions of the gene to be disrupted (16). Oligonucleotides used to generate disruption cassettes for srs2, rad22 and rad51 (SRS2A and B, R22A and B and R51A and B, respectively) are listed in Table 2. Following transformation of a diploid strain 428/429, ura+ progeny were screened for the desired integration pattern by diagnostic PCR using primer pairs spanning the presumptive recombination sites (details of the additional primers used for this purpose are available from the authors on request). Frequencies of homologous recombination (i.e. successful targeted gene disruption) ranged from 9 to 80%. Meiosis and sporulation were induced by plating onto malt extract agar, and tetrad dissection was performed with an MSM micromanipulator (Singer Instruments, UK) as described by Moreno et al. (15). Construction of the nmt1 promoter replacement and chromosomally HA epitope-tagged srs2 strains (nmt1:HAsrs2 and srs2:HA) was accomplished by an analogous method using primers NMTA and B and TAGA and B, respectively (Table 2). Deletion of rqh1 was described previously (17).
Table 2. Oligonucleotides used in this study.
 |
Antibodies and immunoblotting
Immunoblotting was performed essentially as described elsewhere (18) using Mini-Protean electrophoresis equipment (Bio-Rad, Hercules, CA) and a semi-dry transfer apparatus (Hoefer) in conjunction with Hybond ECL membranes (Amersham Pharmacia, UK). Proteins were detected using enhanced chemiluminescence (ECL, Amersham) following 1 h incubations at room temperature with the respective primary and horseradish peroxidase-conjugated anti-mouse antibodies (Sigma, Poole, UK). The mouse anti-influenza haemagglutinin (HA) monoclonal HA-11 (BAbCO, Berkeley, CA) was used at 1 µg/ml for detection of HA-tagged Srs2. Cdc2 was detected using the mouse monoclonal antibody Y100 (generated by Dr J. Gannon and kindly provided by Dr H. Yamano).
Microscopy and flow cytometry
Cells fixed in 70% ethanol were re-hydrated and stained with 4′,6-diamidino-2-phenylindole (DAPI) before examination by fluorescence microscopy. Images were acquired using a Zeiss Axioskop equipped with an Axiocam cooled CCD camera and Axiovision software (Carl Zeiss Ltd, Welwyn Garden City, UK) and were assembled using Adobe Photoshop. For flow cytometric analysis, cells were fixed at a density of 107/ml in 70% ethanol at 4°C overnight. Fixed cells were washed once with 50 mM sodium citrate buffer (pH 7), resuspended in the same buffer containing 100 µg/ml DNase-free RNase and incubated at 37°C overnight. After staining with propidium iodide (4 µg/ml final concentration), cells were analysed by flow cytometry (FACScan, Becton Dickinson, Oxford, UK) using a 488 nm laser. Red fluorescence (DNA) and forward light scatter (cell size) data were collected on linear scales for 10 000 cells for each strain and were analysed using CellQuest software.
RESULTS
Identification of the S.pombe srs2 gene and protein
BLAST searches of the near-complete S.pombe genome database using the 1175 amino acid S.cerevisiae Srs2 protein sequence identified a single novel S.pombe gene (designated SPAC4H3.05) encoding a protein (Swiss-Prot accession number Q10213) significantly related to the query sequence throughout most of its length (27% amino acid identity, 40% similarity over a 698 amino acid region; Fig. 1A). Among all the putative ATP-dependent DNA helicases in the Swiss-Prot database, the hypothetical S.pombe protein sequence was most closely related to that of S.cerevisiae Srs2. We consequently refer to this SRS2-like S.pombe gene as srs2+, in deference to the established S.cerevisiae nomenclature. The C-terminal regions of the two Srs2 sequences lacked significant similarity either to each other or to other known proteins.
The srs2+ gene encodes a predicted protein of 887 amino acid residues, with a calculated molecular weight of 101 061. Homologous recombination with a PCR-generated fragment was used to place an HA epitope tag sequence in-frame with the 3′ end of the srs2 open reading frame. Western blotting demonstrated the expression of the HA-tagged Srs2 protein, and indicated that there was no obvious change in the abundance or electrophoretic mobility of this protein after exposure of S.pombe cells to HU or the radiomimetic drug bleomycin (Fig. 1B). Consistent with the calculated molecular weight, HA-tagged Srs2 had a migration appropriate to a protein of just over 100 kDa.
Deletion of srs2 causes hyper-recombination and sensitivity to DNA damage
As a first step towards determining the function of srs2+ in S.pombe, one copy of the gene was replaced with the ura4+ marker in a ura4-D18 diploid strain and, after induction of meiosis, tetrads were dissected. Haploid ura+ progeny segregated 2:2, indicating that the srs2 gene is not essential for growth. The ura+, srs2-deleted (srs2Δ) progeny had plating efficiencies and growth rates similar to those of their srs2+ (ura–) counterparts and to rqh1-deleted (rqh1Δ) cells (Fig. 2A and Table 3).
Figure 2.
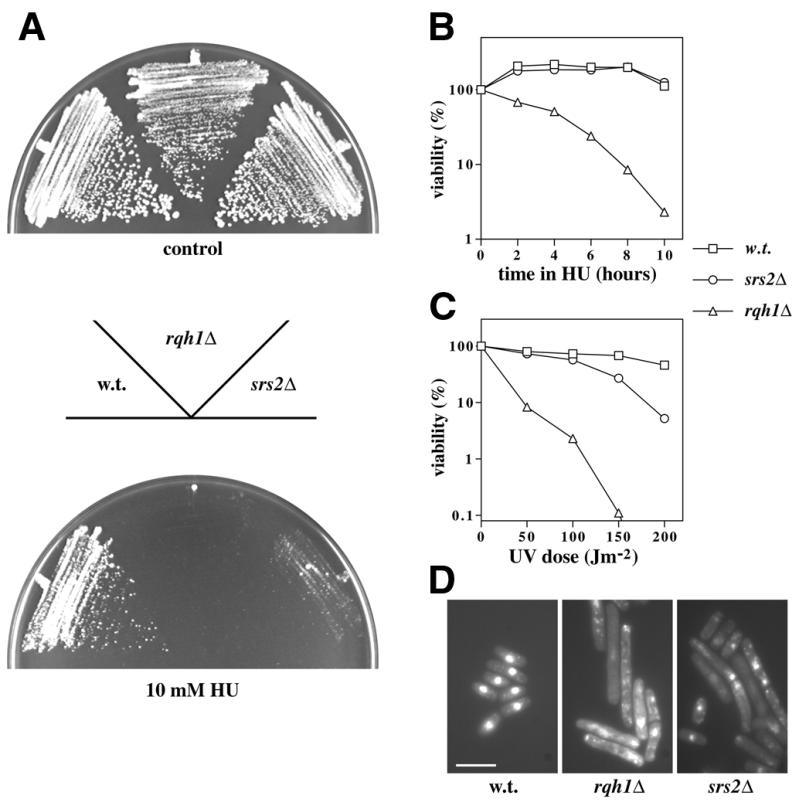
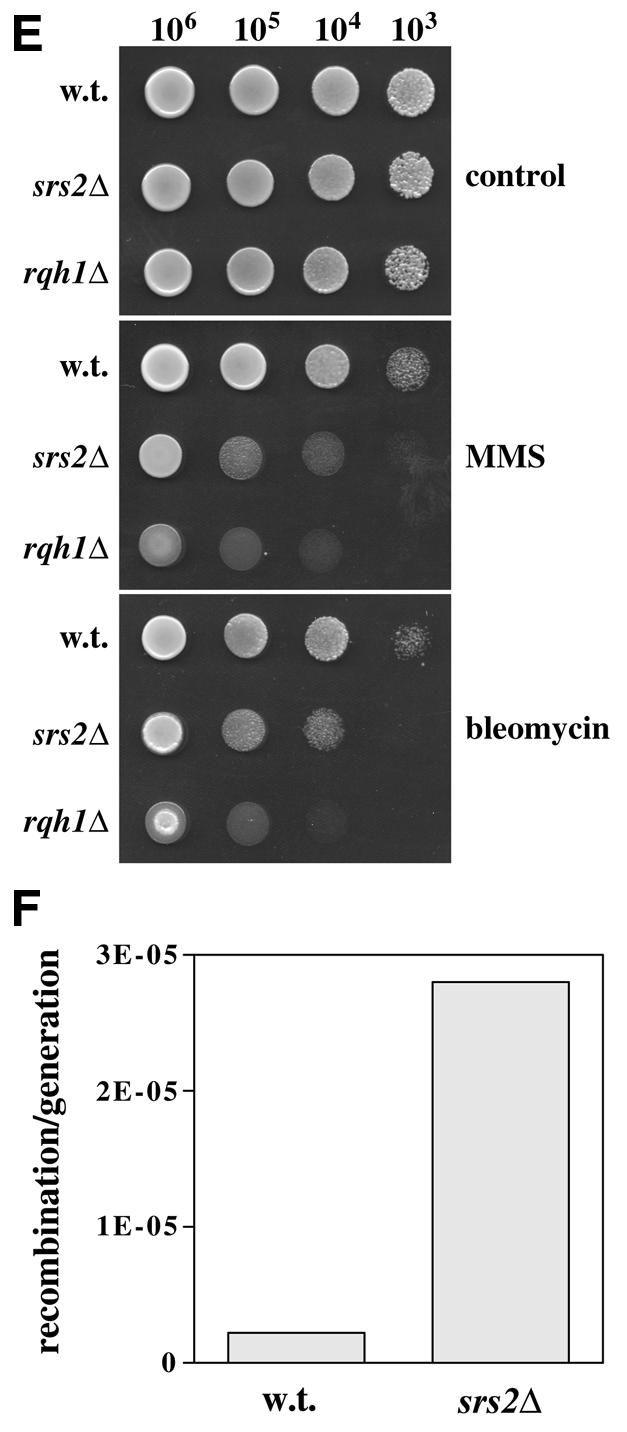
Characterisation of the srs2Δ phenotype; sensitivity to HU, UV, MMS or bleomycin, and elevated recombination rate. (A) A wild-type strain (HM123) and the single mutants rqh1Δ and srs2Δ were streaked in the positions shown onto YPD agar (control) or YPD agar containing 10 mM HU, as indicated. Plates were photographed after 4 days growth at 30°C. (B) The same strains were exposed to 10 mM HU in liquid culture for up to 10 h. Viability relative to that seen before addition of HU was measured by plating aliquots of each culture onto YPD agar plates and counting colony numbers after 4 days growth at 30°C. (C) Sensitivity of the same strains to UV was measured similarly by counting colony numbers after exposure to the indicated UV doses on YPD agar plates. (D) Cell and nuclear morphologies of rqh1+ srs2+ (HM123), rqh1Δ and srs2Δ strains were determined by fluorescence microscopy of DAPI-stained cells 14 h after exposure to 200 Jm–2 UV. Scale bar: 10 µm. (E) Ten-fold serial dilutions of these strains spanning the range from 106 to 103 cells, as indicated, were spotted onto YPD agar plates containing 0.005% MMS, 5 µg/ml bleomycin or neither drug (control). Plates were photographed after 3–5 days incubation at 30°C. (F) Recombination rates were measured in srs2+ (HM123) and srs2Δ strains as described in Materials and Methods. Data are expressed as mean recombination frequency per generation.
Table 3. Growth characteristics of srs2Δ, rqh1Δ, rhp51Δ, rad22Δ and rhp54Δ strains.
| Strain |
Doubling time (h) |
Plating efficiency (%) |
| HM123 (wild-type) | 2.3 | 98 |
| srs2Δ | 2.6 | 85 |
| rqh1Δ | 2.7 | 87 |
| srs2Δ rqh1Δ | 6.3 | 36 |
| rhp51Δ | 3.5 | 73 |
| rhp51Δ rqh1Δ | 3.8 | 58 |
| rhp51Δ srs2Δ | 4.8 | 43 |
| rhp51Δ srs2Δ rqh1Δ | 7.2 | 21 |
| rad22Δ | 4.0 | 48 |
| rad22Δ rqh1Δ | 7.5 | 10 |
| rhp54Δ | 3.8 | 64 |
| rhp54Δ rqh1Δ | 4.0 | 31 |
We next compared the sensitivites of srs2Δ and rqh1Δ cells to agents that inhibit DNA replication or induce DNA damage. Schizosaccharomyces pombe srs2Δ cells were mildly sensitive to HU, although not as sensitive as rqh1Δ cells, as judged by continuous growth on agar containing HU (Fig. 2A). The rqh1Δ strain lost >90% cell viability by 8 h exposure to HU in liquid culture, as judged by colony formation (Fig. 2B). In contrast, there was no significant loss of viability in the srs2Δ strain after short-term HU treatment. The sensitivity of srs2Δ cells grown on agar containing HU therefore presumably reflects a requirement for Srs2 during continuous exposure to HU.
There was no significant loss of viability in the srs2Δ strain after exposure to 50 J/m2 UV, a dose sufficient to cause a 90% loss of viability of rqh1Δ cells (Fig. 2C). At higher doses, srs2Δ cells were significantly UV sensitive (∼90% loss of viability at 200 J/m2) in comparison with a srs2+ control, but were far less sensitive than rqh1Δ cells. Microscopic examination of cells 14 h after UV treatment (200 J/m2) showed that wild-type controls had regained a normal morphology by this time (Fig. 2D). In comparison, UV-treated srs2Δ or rqh1Δ cells displayed a variety of defects, including cell elongation, aberrant nuclear morphology and unequal chromosome segregation. In some cases the highly elongated cells contained a single mass of chromatin bisected by a septum. In the case of rqh1 mutants, this ‘late cut’ phenotype reflects failure to resume cell cycle progression faithfully after UV-induced cell cycle arrest, which explains the cell elongation (4). Both rqh1Δ and srs2Δ strains showed significant sensitivity to the alkylating agent methyl methanesulphonate (MMS) and to the radiomimetic drug bleomycin (Fig. 2E). In each case the srs2Δ strain displayed a sensitivity intermediate to those of the wild-type and rqh1Δ strains.
Sensitivity to DNA damage in rqh1 and sgs1 mutants is associated with elevated levels of mitotic recombination. In order to address whether or not the same is true for srs2Δ cells, srs2+ and srs2Δ stable diploid strains were constructed containing the ade6-M26 and ade6-L469 heteroalleles. Reciprocal recombination or gene conversion between these alleles can lead to the generation of ade+ progeny, and the frequency with which these arise therefore provides a convenient index of mitotic recombination rate (4). Using this approach, we found that the spontaneous recombination rate in the srs2Δ diploid strain was elevated ∼12-fold with respect to the wild-type control (Fig. 2F).
Simultaneous deletion of srs2 and rqh1 causes a dramatic growth defect
As simultaneous mutation of budding yeast SGS1 and SRS2 results in lethality, we were interested to see whether there might be an analogous interaction between the corresponding genes in S.pombe. A diploid strain was constructed in which one allele of srs2 was disrupted by the ura4+ marker and one allele of rqh1 was disrupted by the kanMX6 cassette, conferring resistance to the neomycin analogue G418. Dissection of ascospores from this strain following induction of meiosis showed that, as expected, haploid segregants bearing either the srs2::ura4+ or the rqh1::kanMX6 allele were able to grow and form colonies normally. From 90 tetrads dissected, the numbers of srs2+ rqh1+, srs2Δ rqh1+ and srs2+ rqh1Δ segregant colonies visible after 3 days growth at 30°C were 78, 76 and 80, respectively. These numbers are close to the 90 that would be expected for independently segregating markers, and indicate that there was only a slight overall loss of spore viability attributable to heterozygosity at the srs2 and rqh1 loci in the diploid. In contrast, only 14 srs2Δ rqh1Δ segregant colonies were visible after 7 days growth, and these formed only extremely slowly growing microcolonies. Microscopic examination showed that many of the cells in these slowly growing colonies were highly elongated, suggesting that one or more aspects of cell cycle progression are defective when both Rqh1 and Srs2 are absent (Fig. 3A). DAPI staining of these cells showed that cell and nuclear morphologies were relatively normal in the srs2Δ and rqh1Δ single mutants, although some of the cells in each of these strains were somewhat elongated (Fig. 3B). In line with their poor overall viability, the srs2Δ rqh1Δ double mutants exhibited a variety of aberrant morphologies, including highly elongated cells, ‘cut’ cells and others with apparently much reduced nuclear DNA content. The overall appearance of these cells was broadly similar to that of UV-treated srs2Δ or rqh1Δ single mutants (Fig. 2D). Flow cytometric analysis confirmed that the DNA content and overall size of srs2Δ rqh1Δ cells were very variable in comparison with those of the corresponding single mutants (Fig. 3C). Some 17% of the srs2Δ rqh1Δ cells had little or no nuclear DNA as judged by this assay, compared with 0.17, 0.63 and 6.5% for wild-type, srs2Δ and rqh1Δ cells, respectively.
Figure 3.
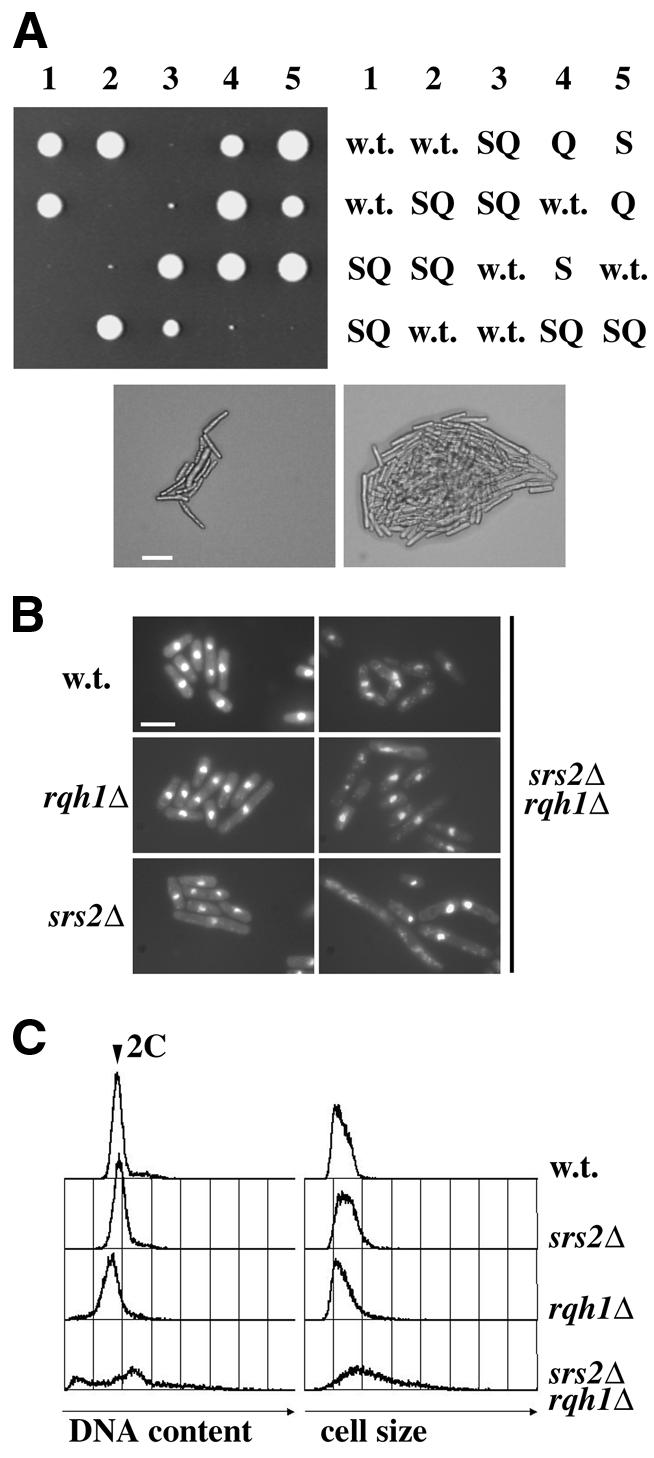
Genetic interaction between srs2 and rqh1. (A) An h+/h– srs2::ura4+/srs2+ rqh1::kanMX6/rqh1+ diploid [srs2Δ × rqh1Δ(kanMX6)] was induced to undergo meiosis and sporulation, and tetrads were microdissected onto YPD agar. Colonies resulting from five such tetrads were photographed after 7 days growth at 30°C (upper left). The genotypes of these segregants were determined by replica plating and are indicated schematically (upper right: w.t., srs2+ rqh1+; S, srs2Δ rqh1+; Q, srs2+ rqh1Δ; SQ, srs2Δ rqh1Δ). Micrographs of two representative srs2Δ rqh1Δ colonies, taken after 7 days growth at 30°C, are shown below (scale bar: 20 µm). (B) Fluorescence micrographs of DAPI-stained srs2+ rqh1+ (w.t.), rqh1Δ, srs2Δ and srs2Δ rqh1Δ cells (scale bar: 10 µm). (C) Flow cytometric analyses of propidium iodide-stained cells of the strains indicated. Histograms of DNA content (red fluorescence, left) and cell size (forward light scatter, right) derived from 10 000 cells are shown. The position corresponding to a 2C (G2/M) DNA content is indicated (arrow). Note that the brevity of G1 and the coincidence of S phase with cytokinesis result in the absence of a detectable 1C population in exponentially growing S.pombe cultures.
Deletion of rhp51 does not suppress the growth defect of srs2 rqh1 double mutants
A recent study showed that the loss of viability in S.cerevisiae sgs1 srs2 strains can be partially suppressed by simultaneous loss of the homologous recombination function encoded by RAD51 (7). With this in mind, we investigated the effect of deletion of rhp51, the S.pombe orthologue of RAD51 (19), on the viability of srs2Δ rqh1Δ cells. A diploid strain was constructed that was heterozygous for deletions of srs2 (replaced in this case with the LEU2 marker) rqh1 (kanMX6) and rhp51 (ura4+). Following sporulation and dissection of tetrads, identification of the genotypes of the progeny showed that deletion of rhp51 did not suppress the growth defect characteristic of srs2Δ rqh1Δ cells (Fig. 4A). On the contrary, srs2Δ rqh1Δ rhp51Δ cells were barely viable at all, with a longer doubling time and lower plating efficiency than those of an otherwise isogenic srs2Δ rqh1Δ mutant (Fig. 4B and Table 3). From 116 tetrads dissected, only five slowly growing srs2Δ rqh1Δ rhp51Δ microcolonies (and nine srs2Δ rhp51Δ colonies) were visible after 7 days growth at 30°C. In contrast, the numbers of srs2+ rqh1+ rhp51+ and srs2Δ segregants (50 and 63, respectively) were close to the 58 expected in the case of three independently segregating markers (116 × 4/23). The additional growth defect conferred by rhp51 deletion was ascribed largely to a genetic interaction between rhp51 and srs2. Deletion of rqh1 in a rhp51Δ background resulted in a slightly increased doubling time and decreased plating efficiency in comparison with the rhp51Δ single mutant, but the effect of srs2 deletion in this background was much more severe (Fig. 4B and Table 3). srs2Δ rhp51Δ double mutants grew at a similarly slow rate to that seen in srs2Δ rqh1Δ double mutants.
Figure 4.
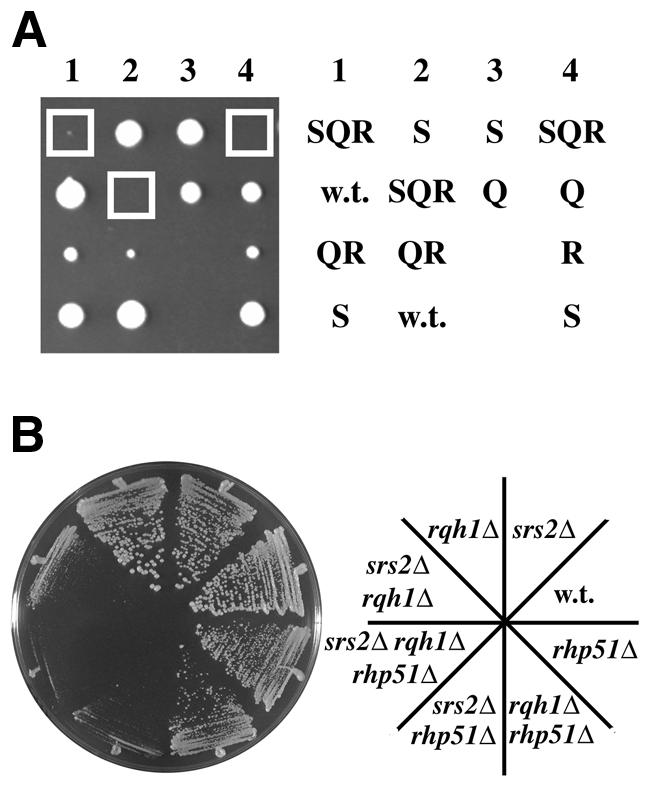
Deletion of rhp51 fails to suppress the srs2Δ rqh1Δ growth defect. (A) An h+/h– srs2::LEU2/srs2+ rqh1::kanMX6/rqh1+ rhp51::ura4+/rhp51+ diploid was induced to undergo meiosis and sporulation, and tetrads were microdissected onto YPD agar. Colonies resulting from four such tetrads were photographed after 7 days growth at 30°C (left). The genotypes of these segregants were determined by replica plating and are indicated schematically (right: w.t., srs2+ rqh1+ rhp51+; S, srs2Δ rqh1+; Q, srs2+ rqh1Δ; R, rhp51Δ; QR, rqh1Δ rhp51Δ; SQR, srs2Δ rqh1Δ rhp51Δ). The positions occupied by srs2Δ rqh1Δ rhp51Δ mutants are boxed (left). (B) The strains indicated (right) were streaked onto YPD agar and photographed (left) after 4 days growth at 30°C.
Homologous recombination is required for maintenance of viability in srs2 and rqh1 mutants
The genetic interaction between srs2 and rhp51 described above was investigated in further detail by examining the effect of combining srs2Δ with deletions of rad22 or rhp54 (S.pombe homologues of S.cerevisiae RAD52 and RAD54, respectively), which like rhp51 encode components of the homologous recombination pathway (19–22). Moreover, it was of interest to determine whether or not the lethality previously observed to result from combining srs2 and rad54 mutations in S.cerevisiae (23) would also be seen in S.pombe. Diploid strains heterozygous for deletion of srs2 and for deletion of rhp51, rad22 or rhp54 were constructed (see Materials and Methods). Dissection of ascospores following induction of meiosis provided independent confirmation that there is a synthetic interaction between srs2 and rhp51 (Fig. 5A). Of 108 tetrads dissected from the srs2Δ rhp51Δ heterozygous diploid, only 13 spores gave rise to viable haploid srs2Δ rhp51Δ colonies (in contrast to the 108 that would be expected for a cross involving two unlinked markers), and all of these grew very slowly. The severity of this synthetic effect with srs2 was if anything even more extreme in the cases of rad22 and rhp54 (Fig. 5B and C), although spore viability was only 64% even in rhp54Δ single mutants. Of 108 tetrads dissected from the srs2Δ rad22Δ heterozygous diploid, and 126 dissected from the srs2Δ rhp54Δ heterozygous diploid, one and zero spores, respectively, gave rise to doubly mutant haploid microcolonies. Thus, S.pombe cells lacking Srs2 are dependent on the homologous recombination pathway for maintenance of viability.
Figure 5.

Genetic interactions between srs2 and homologous recombination genes. (A–C) Tetrads derived from diploid strains of the following genotypes were microdissected onto YPD agar: h+/h– srs2::LEU2/srs2+ rhp51::ura4+/rhp51 (A), h+/h– srs2::ura4+/srs2+ rad22::kanMX6/rad22+ (B) and h+/h– srs2::LEU2/srs2+ rhp54::ura4+/rhp54+ (C). Colonies resulting from four tetrads in each case were photographed after 7 days growth at 30°C. The genotypes of the segregants were determined by replica plating and are indicated schematically [right: w.t., srs2+ rqh1+ rhp51+ rad22+ rhp54+; S, srs2Δ; R, rhp51Δ (A), rad22Δ (B) or rhp54Δ (C); SR, srs2Δ rhp51Δ (A), srs2Δ rad22Δ (B) or srs2Δ rhp54Δ (C)]. Boxes (left) indicate the positions of srs2 double mutants with rhp51, rad22 and rhp54, respectively. (D) The strains indicated (right) were streaked onto YPD agar and photographed (left) after 4 days growth at 30°C.
Genetic interactions between rqh1 and homologous recombination genes were also examined in further detail. An earlier study demonstrated that the UV sensitivities of rqh1 and rhp51 or rhp54 mutants were not additive (5). Similarly, we found that rqh1Δ rhp51Δ and rqh1Δ rhp54Δ mutants grew only slightly more slowly than the single rhp51Δ mutant (Fig. 5D and Table 3), although their plating efficiencies were somewhat reduced. A much more pronounced synthetic interaction was seen on combining rqh1Δ with rad22Δ, however. Thus, the doubling time of a rqh1Δ rad22Δ strain was almost twice that of a rad22Δ single mutant, and the plating efficiency was decreased almost 5-fold (Fig. 5D and Table 3).
The epistatic relationship between rqh1 and rhp51/rhp54 with respect to UV sensitivity (5) has been interpreted in terms of an involvement of the products of these genes in a common pathway of DNA damage tolerance. The modest UV sensitivity of srs2Δ cells (Fig. 2C) and the synthetic interactions between srs2 and homologous recombination genes in the absence of exogenous DNA damage (Fig. 5A–C) suggested that srs2 is unlikely to participate in such a pathway, at least not to the same extent as rqh1. In line with this interpretation, the UV sensitivity conferred by srs2 deletion was additive with that resulting from deletion of rhp51, required for recombinational repair, or rad13, which is essential for the NER pathway (Fig. 6). By these criteria it would appear that srs2 is required for a pathway of UV damage repair that is neither the recombination pathway nor NER.
Figure 6.

UV sensitivity resulting from srs2 deletion is additive with that resulting from loss of homologous recombination or NER. (A and B) Sensitivity of the indicated strains to UV, relative to that seen without irradiation, was measured by counting colony numbers after 4 days growth at 30°C after exposure to the indicated UV doses on YPD agar plates.
Overexpression of srs2 cannot compensate for deletion of rqh1
If the predicted DNA helicase activity of Srs2 were functionally related to that of Rqh1, overexpression of Srs2 might be sufficient to suppress defects seen in the absence of Rqh1. To address this point, PCR-mediated gene disruption was used to replace the promoter of the srs2 gene in its normal chromosomal context with the thiamine-repressible nmt1 promoter, at the same time introducing an HA epitope tag sequence fused in-frame to the 5′ end of the srs2 open reading frame (see Materials and Methods). Removal of thiamine from the medium led to readily detectable levels of HA-Srs2 after 16 h growth at 30°C (Fig. 7A). In addition to the full-length, HA-tagged protein, a number of smaller species were detected, the most prominent of which migrated at ∼35 kDa. Since the epitope tag is at the N-terminus, these shorter species most likely represent degradation products. Comparison with levels of Srs2-HA expressed from the srs2 promoter in its normal chromosomal context indicated that the nmt1 promoter-driven gene was overexpressed ∼15-fold in comparison with endogenous srs2, but haploid HA-Srs2-expressing cells were not UV sensitive, suggesting that the overproduced protein is functional (S.W.Wang and C.Norbury, unpublished results). This level of srs2 overexpression had no effect on the UV (Fig. 7B) or HU (Fig. 7C) sensitivity of a rqh1Δ strain, however, indicating that Srs2 is incapable of performing the function of Rqh1 in response to these agents.
Figure 7.
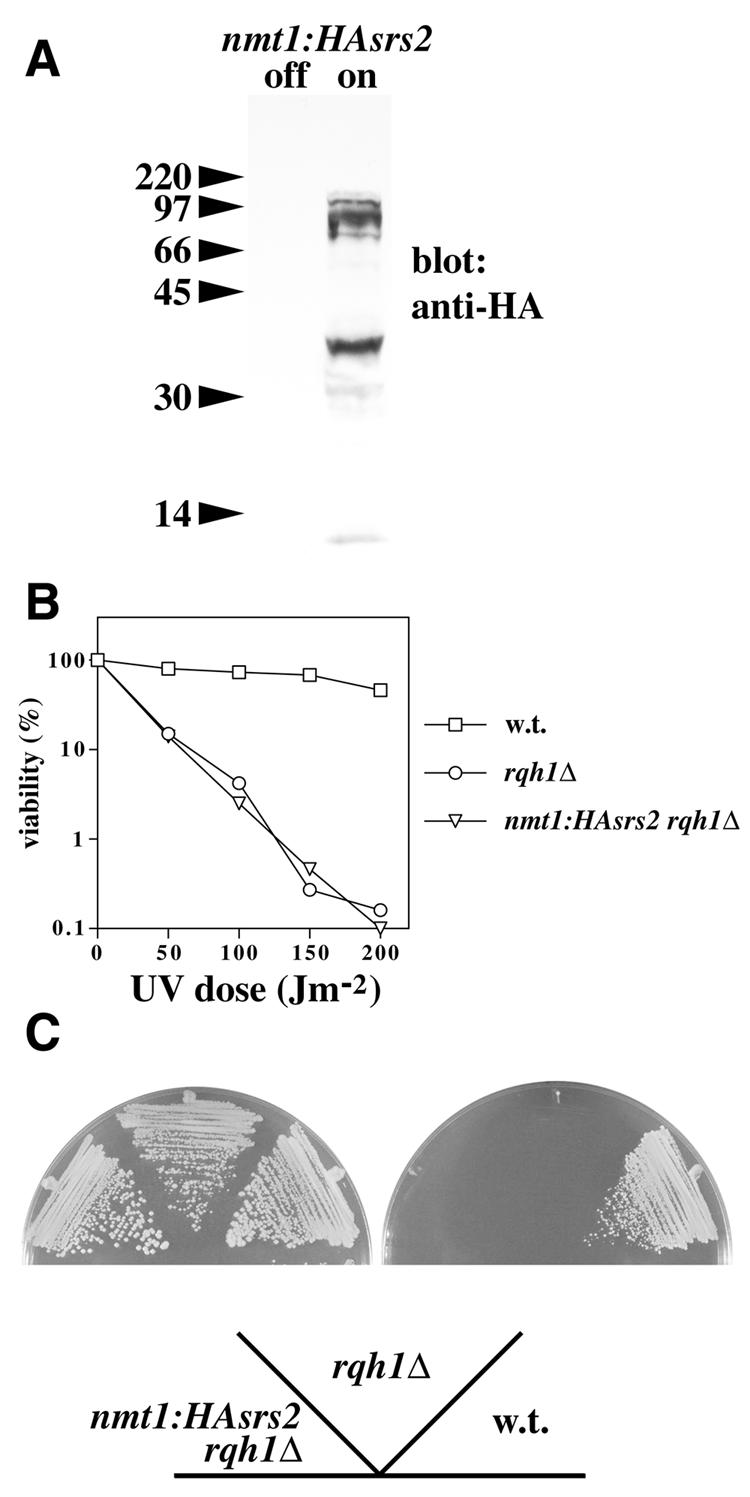
Overexpression of srs2 fails to suppress the UV or HU sensitivity of a rqh1Δ mutant. (A) A strain (nmt1:HAsrs2) was constructed in which the endogenous srs2 promoter is replaced by that of the thiamine-repressible nmt1 gene, while incorporating an HA epitope tag sequence at the 5′ end of the srs2 open reading frame (see Materials and Methods). Whole cell protein extracts were prepared from this strain grown to mid-exponential phase at 30°C in the presence of thiamine (off) or 16 h after the removal of thiamine (on), and were subjected to immunoblotting with an anti-HA antibody. (B) The UV sensitivities of the strains indicated were determined (as described in the legend to Fig. 6) 16 h after the removal of thiamine. (C) The same strains were streaked onto EMM2 agar (which lacks thiamine; plate on left) or EMM2 agar containing 10 mM HU (plate on right) and were photographed after 3–5 days growth at 30°C.
Deletion of srs2 fails to suppress the lethality of top3 deletion
We showed previously that the lethality resulting from deletion of the top3 gene encoding S.pombe topoisomerase III can be suppressed by deletion of rqh1 (17,24). Similarly, in S.cerevisiae, deletion of SGS1 suppresses the slow growth phenotype associated with TOP3 deletion (25). If srs2 were functionally related to rqh1, it might be expected that there would be an analogous genetic interaction between srs2 and top3. On dissection of meiotic progeny from a diploid strain heterozygous for both srs2 and top3 deletions, none of the top3::ura4+ spores was able to form colonies, regardless of the srs2 genotype (data not shown). This lack of suppression of the top3 phenotype by srs2 deletion provides further evidence that the functions of srs2 and rqh1 are quite distinct.
DISCUSSION
Budding yeast SRS2/RADH acts with SGS1 to suppress unscheduled recombination, and this function may be particularly important during DNA replication. Although structural and functional homologues of SGS1 have been identified in eukaryotic species as diverse as yeasts and mammals, this report is the first to describe an SRS2-like gene function in any eukaryote other than S.cerevisiae. The completion of genome sequencing projects will make it possible to establish whether or not Srs2-like proteins exist in non-fungal eukaryotic species, but at this stage it has not been possible to identify clear candidates among the available Drosophila, Caenorhabditis or human sequences (S.-W.Wang and C.Norbury, unpublished observations). The bacterial helicases most closely related to Srs2 (those of the PcrA family) are essential for DNA replication and repair (26). In S.cerevisiae an essential role for Srs2 and Sgs1 in chromosomal replication has also been proposed (11), but such a role is difficult to reconcile with the observation that the srs2 sgs1 double mutant phenotype can be suppressed by simultaneous inactivation of the homologous recombination machinery (7). BLAST searches show that the S.pombe Srs2 amino acid sequence is significantly more similar to the bacterial members of this family than is S.cerevisiae Srs2. This could indicate that the common ancestral yeast contained two diverging Srs2 isoforms, only one of which is retained in each of the modern species. Other explanations, such as horizontal gene transfer from bacteria to an S.pombe ancestor, or more rapid sequence divergence in S.cerevisiae, cannot be ruled out however.
We have shown that Srs2 in S.pombe is required to maintain the normal level of resistance to UV irradiation, as is its counterpart in S.cerevisiae. The precise nature of this protective function is not yet clear. A cell cycle checkpoint role was proposed recently for S.cerevisiae Srs2 (27), but we currently have no indication that Srs2 has an analogous checkpoint function in S.pombe. Unlike the S.cerevisiae protein, S.pombe Srs2 was not phosphorylated in a checkpoint-dependent manner, as far as it was possible to judge by SDS–PAGE (Fig. 1B). In addition, srs2Δ cells, like rqh1Δ cells, became substantially elongated after UV irradiation (Fig. 2D), suggesting that establishment of the S.pombe DNA damage checkpoint is not Srs2 dependent. Instead, like Rqh1, Srs2 may be involved in a damage tolerance pathway that involves bypass of DNA damage by replication forks, reversal of cell cycle arrest and/or suppression of recombination (4,5). The substantially elevated level of spontaneous recombination seen in srs2Δ cells (Fig. 2F) would be compatible with such a role. Although elevated recombination was also seen on deletion of rqh1, this was not intrinsic, but dependent on inhibition of replication (4).
An alternative interpretation of the data would be that Srs2, like Rad6 and Rad18 in S.cerevisiae, functions in a post-replication gap-filling repair pathway. Indeed, the SRS2 locus was first identified through dominant mutations capable of suppressing the UV sensitivities of rad6 and rad18 mutants (28). In this case, the hyper-recombination phenotype seen on srs2 deletion could be attributable to an increased reliance on a recombinational pathway of post-replication repair, while simultaneous inactivation of this alternative pathway would give rise to the observed lethality or slow growth phenotype. A very recent study concluded that the synthetic lethality observed in S.cerevisiae srs2 rad54 strains was attributable to activation of the DNA damage checkpoint (29). It will be interesting to determine whether or not inactivation of this checkpoint pathway can suppress the lethality seen in the equivalent double mutant in S.pombe.
Deletion of rhp51 was reported previously to confer no additional sensitivity to UV on an rqh1 deletion strain, suggesting that the two genes might act in a common pathway of DNA damage tolerance (5). In contrast, we showed that the UV sensitivities seen on deletion of srs2 and rhp51 are additive (Fig. 6A), suggesting that the UV-protective role of srs2 does not operate through the recombinational repair pathway. Similarly, epistasis suggests that srs2 acts independently of the NER pathway (Fig. 6B). It is possible that Srs2 acts in the NHEJ pathway of DNA repair in S.pombe, as has been reported recently for S.cerevisiae Srs2 (30), but at this stage it seems at least equally likely that its principal role is in a non-recombinational gap-filling pathway, as discussed above. Examination of the relationship between Rqh1, Srs2 and the homologous recombination machinery also revealed a specific requirement for rad22, but not other homologous recombination genes, in cells lacking rqh1 (Fig. 5D and Table 3). Rad22/Rad52 homologues bind DNA ends and appear to act at an early stage in the homologous recombination pathway, interacting with Rad51 to promote homologous pairing (31–33). It is conceivable that Rqh1 and its homologues might act in concert with Rad22/Rad52 at this early stage in recombination, potentially explaining the specific genetic interaction observed here between rqh1 and rad22. The precise nature of any such interaction awaits further investigation.
While deletion of srs2 on its own resulted in a comparatively minor growth defect, simultaneous deletion of srs2 and rqh1 was barely compatible with cell survival (Fig. 3 and Table 3). Thus, the function of Srs2 is conserved between S.cerevisiae and S.pombe at the level of genetic interaction with the sole RecQ family helicase in each yeast (Sgs1 and Rqh1, respectively). The analogous growth defect seen in S.cerevisiae srs2 sgs1 mutants has been attributed to unrestrained recombination, as the phenotype can be suppressed by inactivation of the homologous recombination pathway (7). We have been unable to establish the underlying cause of the growth defect in srs2Δ rqh1Δ cells, partly because the strain itself grew so poorly, and partly because the growth defect is not suppressed by deletion of the homologous recombination gene rhp51 (Fig. 4). Thus, despite our observation that spontaneous recombination is elevated in srs2Δ cells (Fig. 2F), we cannot implicate unscheduled spontaneous hyper-recombination in the srs2Δ rqh1Δ growth defect. Interpretation of the failure of rhp51 deletion to suppress this growth defect is complicated by the fact that the rhp51Δ single mutant displays a growth defect, whereas no such defect has been described for the corresponding S.cerevisiae mutant. However, we observed a specific genetic interaction between rhp51 and srs2, and a comparative lack of any such interaction between rhp51 and rqh1 (Figs 4B and 5A, and Table 3). These observations support the notion that the relationship between srs2/SRS2 and the recombination pathway differs between S.cerevisiae and S.pombe.
In summary, we have identified the S.pombe orthologue of the S.cerevisiae SRS2 gene. Several aspects of this gene function have been conserved during the interval of ∼400 million years since the ancestors of these two yeasts diverged. In particular, srs2 mutations in each yeast confer elevated levels of mitotic recombination and sensitivity to DNA damage, and are synthetically lethal with mutations in rgh1/SGS1 and rhp54/RAD54. These functional similarities, along with the extensive sequence similarity (Fig. 1A), justify our conclusion that srs2+ is the closest fission yeast counterpart of SRS2. In other respects, the srs2/SRS2 function appears to have diverged. Thus, inactivation of the homologous recombination machinery, which is sufficient to suppress the lethality of S.cerevisiae srs2 sgs1 mutants, was not capable of similarly suppressing the srs2 rqh1 phenotype in S.pombe.
Acknowledgments
ACKNOWLEDGEMENTS
We are grateful to Tamar Enoch for providing the strains and methodology for measurement of recombination rates, to Albert Pastink and Tony Carr for providing S.pombe strains, and to Takashi Toda and members of the Molecular Oncology Laboratory for their helpful advice. This work was supported by the Imperial Cancer Research Fund.
References
- 1.Carr A.M. (1997) Control of cell cycle arrest by the Mec1sc/Rad3sp DNA structure checkpoint pathway. Curr. Opin. Genet. Dev., 7, 93–98. [DOI] [PubMed] [Google Scholar]
- 2.O’Connell M.J., Walworth,N.C. and Carr,A.M. (2000) The G2-phase DNA-damage checkpoint. Trends Cell Biol., 10, 296–303. [DOI] [PubMed] [Google Scholar]
- 3.Chakraverty R.K. and Hickson,I.D. (1999) Defending genome integrity during DNA replication: a proposed role for RecQ family helicases. BioEssays, 21, 286–294. [DOI] [PubMed] [Google Scholar]
- 4.Stewart E., Chapman,C.R., Al-Khodairy,F., Carr,A.M. and Enoch,T. (1997) rqh1+, a fission yeast gene related to the Bloom’s and Werner’s syndrome genes, is required for reversible S phase arrest. EMBO J., 16, 2682–2692. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Murray J.M., Lindsay,H.D., Munday,C.A. and Carr,A.M. (1997) Role of Schizosaccharomyces pombe RecQ homolog, recombination and checkpoint genes in UV damage tolerance. Mol. Cell. Biol., 17, 6868–6875. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Frei C. and Gasser,S.M. (2000) The yeast Sgs1p helicase acts upstream of Rad53p in the DNA replication checkpoint and colocalizes with Rad53p in S-phase-specific foci. Genes Dev., 14, 81–96. [PMC free article] [PubMed] [Google Scholar]
- 7.Gangloff S., Soustelle,C. and Fabre,F. (2000) Homologous recombination is responsible for cell death in the absence of the Sgs1 and Srs2 helicases. Nature Genet., 25, 192–194. [DOI] [PubMed] [Google Scholar]
- 8.Heartlein M.W., Tsuji,H. and Latt,S.A. (1987) 5-Bromodeoxyuridine-dependent increase in sister chromatid exchange formation in Bloom’s syndrome is associated with reduction in topoisomerase II activity. Exp. Cell Res., 169, 245–254. [DOI] [PubMed] [Google Scholar]
- 9.Watt P.M., Hickson,I.D., Borts,R.H. and Louis,E.J. (1996) SGS1, a homologue of the Bloom’s and Werner’s syndrome genes, is required for maintenance of genome stability in Saccharomyces cerevisiae. Genetics, 144, 935–945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Aboussekhra A., Chanet,R., Adjiri,A. and Fabre,F. (1992) Semidominant suppressors of Srs2 helicase mutations of Saccharomyces cerevisiae map in the RAD51 gene, whose sequence predicts a protein with similarities to procaryotic RecA proteins. Mol. Cell. Biol., 12, 3224–3234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Lee S.K., Johnson,R.E., Yu,S.L., Prakash,L. and Prakash,S. (1999) Requirement of yeast SGS1 and SRS2 genes for replication and transcription. Science, 286, 2339–2342. [DOI] [PubMed] [Google Scholar]
- 12.Liao S., Graham,J. and Yan,H. (2000) The function of Xenopus Bloom’s syndrome protein homolog (xBLM) in DNA replication. Genes Dev., 14, 2570–2575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Klein H.L. (2000) A radical solution to death. Nature Genet., 25, 132–134. [DOI] [PubMed] [Google Scholar]
- 14.Aboussekhra A., Chanet,R., Zgaga,Z., Cassier-Chauvat,C., Heude,M. and Fabre,F. (1989) RADH, a gene of Saccharomyces cerevisiae encoding a putative DNA helicase involved in DNA repair. Characteristics of radH mutants and sequence of the gene. Nucleic Acids Res., 17, 7211–7219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Moreno S., Klar,A. and Nurse,P. (1991) Molecular genetic analysis of fission yeast Schizosaccharomyces pombe. Methods Enzymol., 194, 795–823. [DOI] [PubMed] [Google Scholar]
- 16.Bähler J., Wu,J.Q., Longtine,M.S., Shah,N.G., McKenzie,A.,III, Steever,A.B., Wach,A., Philippsen,P. and Pringle,J.R. (1998) Heterologous modules for efficient and versatile PCR-based gene targeting in Schizosaccharomyces pombe. Yeast, 14, 943–951. [DOI] [PubMed] [Google Scholar]
- 17.Goodwin A., Wang,S.W., Toda,T., Norbury,C. and Hickson,I.D. (1999) Topoisomerase III is essential for accurate nuclear division in Schizosaccharomyces pombe. Nucleic Acids Res., 27, 4050–4058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Ausubel F.M., Brent,R., Kingston,R.E., Moore,D.D., Seidman,J.G., Smith,J.A. and Struhl,K. (1995) Current Protocols in Molecular Biology. John Wiley & Sons, Inc., New York, NY.
- 19.Muris D.F.R., Vreeken,K., Schmidt,H., Ostermann,K., Clever,B., Lohman,P.H.M. and Pastink,A. (1997) Homologous recombination in the fission yeast Schizosaccharomyces pombe: different requirements for the rhp51+, rhp54+ and rad22+ genes. Curr. Genet., 31, 248–254. [DOI] [PubMed] [Google Scholar]
- 20.Schlake C., Ostermann,K., Schmidt,H. and Gutz,H. (1993) Analysis of DNA repair pathways of Schizosaccharomyces pombe by means of swi-rad double mutants. Mutat. Res., 294, 59–67. [DOI] [PubMed] [Google Scholar]
- 21.Ostermann K., Lorentz,A. and Schmidt,H. (1993) The fission yeast rad22 gene, having a function in mating-type switching and repair of DNA damages, encodes a protein homolog to Rad52 of Saccharomyces cerevisiae. Nucleic Acids Res., 21, 5940–5944. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Muris D.F., Vreeken,K., Carr,A.M., Murray,J.M., Smit,C., Lohman,P.H. and Pastink,A. (1996) Isolation of the Schizosaccharomyces pombe RAD54 homologue, rhp54+, a gene involved in the repair of radiation damage and replication fidelity. J. Cell Sci., 109, 73–81. [DOI] [PubMed] [Google Scholar]
- 23.Palladino F. and Klein,H.L. (1992) Analysis of mitotic and meiotic defects in Saccharomyces cerevisiae SRS2 DNA helicase mutants. Genetics, 132, 23–37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Maftahi M., Han,C.S., Langston,L.D., Hope,J.C., Zigouras,N. and Freyer,G.A. (1999) The top3+ gene is essential in Schizosaccharomyces pombe and the lethality associated with its loss is caused by Rad12 helicase activity. Nucleic Acids Res., 27, 4715–4724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Gangloff S., McDonald,J.P., Bendixen,C., Arthur,L. and Rothstein,R. (1994) The yeast type I topoisomerase Top3 interacts with Sgs1, a DNA helicase homolog: a potential eukaryotic reverse gyrase. Mol. Cell. Biol., 14, 8391–8398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Petit M.A., Dervyn,E., Rose,M., Entian,K.D., McGovern,S., Ehrlich,S.D. and Bruand,C. (1998) PcrA is an essential DNA helicase of Bacillus subtilis fulfilling functions both in repair and rolling-circle replication. Mol. Microbiol., 29, 261–273. [DOI] [PubMed] [Google Scholar]
- 27.Liberi G., Chiolo,I., Pellicioli,A., Lopes,M., Plevani,P., Muzi-Falconi,M. and Foiani,M. (2000) Srs2 DNA helicase is involved in checkpoint response and its regulation requires a functional Mec1-dependent pathway and Cdk1 activity. EMBO J., 19, 5027–5038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Lawrence C.W. and Christensen,R.B. (1979) Metabolic suppressors of trimethoprim and ultraviolet light sensitivities of Saccharomyces cerevisiae rad6 mutants. J. Bacteriol ., 139, 866–887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Klein H.L. (2001) Mutations in recombinational repair and in checkpoint control genes suppress the lethal combination of srs2Δ with other DNA repair genes in Saccharomyces cerevisiae. Genetics, 157, 557–565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Hegde V. and Klein,H. (2000) Requirement for the SRS2 DNA helicase gene in non-homologous end joining in yeast. Nucleic Acids Res., 28, 2779–2783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Parsons C.A., Baumann,P., Van Dyck,E. and West,S.C. (2000) Precise binding of single-stranded DNA termini by human RAD52 protein. EMBO J., 19, 4175–4181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Van Dyck E., Stasiak,A.Z., Stasiak,A. and West,S.C. (1999) Binding of double-strand breaks in DNA by human Rad52 protein. Nature, 398, 728–731. [DOI] [PubMed] [Google Scholar]
- 33.Benson F.E., Baumann,P. and West,S.C. (1998) Synergistic actions of Rad51 and Rad52 in recombination and DNA repair. Nature, 391, 401–404. [DOI] [PubMed] [Google Scholar]