

Abstract
Natural Killer (NK) cells are large granular lymphocytes that promote the antitumor response via communication with other cell types in the tumor microenvironment. Previously, we have shown that NK cells secrete a profile of immune stimulatory factors (e.g., IFNγ, MIP-1α, TNFα) in response to dual stimulation with the combination of antibody (Ab)-coated tumor cells and cytokines, such as interleukin (IL) 12. We now demonstrate that this response is enhanced in the presence of autologous monocytes. Monocyte enhancement of NK cell activity was dependent on cell-to-cell contact as determined by a transwell assay. It was hypothesized that NK cell effector functions against Ab-coated tumor cells were enhanced via binding of MICA on monocytes to NK cell NKG2D receptors. Strategies to block MICA-NKG2D interactions resulted in reductions in IFNγ production. Depletion of monocytes in vivo resulted in decreased IFNγ production by murine NK cells upon exposure to Ab-coated tumor cells. In mice receiving trastuzumab and IL12 therapy, monocyte depletion resulted in significantly greater tumor growth in comparison to mock-depleted controls (P < 0.05). These data suggest that NK cell-monocyte interactions enhance NK cell antitumor activity in the setting of monoclonal Ab therapy for cancer.
Keywords: NK cell, monocyte, cytokine, antibody, immunotherapy
Introduction
Natural Killer (NK) cells are innate immune cells that target and lyse virally infected cells as well as malignant cells. NK cells contain cytolytic granules and express an array of cellular adhesion molecules and cytokine receptors [1–4]. They produce a variety of immunostimulatory cytokines including IFNγ, TNFα, MIP-1α, RANTES and GM-CSF [5, 6]. Constitutive expression of multiple cytokine receptors facilitates their response to inflammatory events [7, 8]. NK cell effector functions are regulated by a balance between activating and inhibitory receptors, as well as the expression of target cell MHC class I antigens [8]. NK cells express an activating receptor for the Fc portion of immunoglobulin G (FcγRIIIa) that enables recognition of antibody (Ab)-coated targets. Support for the concept that antitumor activity of therapeutic monoclonal antibodies (mAbs) is in part dependent on FcR-bearing cells stems from experiments conducted in FcR-deficient mice [9] and the observation that polymorphisms in the human FcR gene influence the clinical efficacy of therapeutic mAbs [10, 11]. Studies from our group have shown that cytokine stimulation of NK cells enhances the immune response to Ab-coated tumor cells in vitro, in murine models, and in phase I clinical trials [12–15]. Costimulation via the IL12 receptor (IL12R) and FcR has been shown to activate multiple cellular signaling pathways (e.g., JAK/STAT and PI3-kinase/Akt) and lead to maximal stimulation of NK cell effector functions [15].
NK cells express an activating, lectin-like receptor known as NKG2D [16]. Human NKG2D binds to ligands MICA, MICB, and ULBPs that are expressed by a variety of tumor cell types [17, 18]. Engagement of NKG2D on NK cells results in activation of the adapter protein DAP10, recruitment of the DAP10 binding partner Grb2 and signaling through Vav1, and also activation of PI3-kinase and the downstream serine threonine kinase Akt [19, 20]. NK-cell interactions with NKG2D ligand-expressing tumor cells lead to NK cell-mediated cytotoxicity [21].
The role of reciprocal activation between monocytes and NK cells in immune responses is gaining increased recognition [22–25]. NK cells function as one of the early sources of immunoregulatory cytokines [1]. Upon activation, NK cells produce IFNγ, the prototypic macrophage activating factor, which optimizes monocyte/macrophage function [26, 27]. Activated monocytes then produce cytokines like IL12 and IL15 that stimulate NK cells [28, 29]. This positive feedback system supports the immune response against invading organisms and enables the rapid and effective clearance of infectious agents by monocytes/macrophages. We hypothesized that NK cell effector functions against Ab-coated tumor cells would be enhanced upon interactions with monocytes. We now report the ability of MICA-expressing monocytes to engage NKG2D on NK cells and promote NK-cell activity against Ab-coated tumor targets.
Materials and Methods
Cytokines and antibodies
Recombinant human IL12 (rhuIL12) was provided by Genetics Institute Inc. (Cambridge, MA). Recombinant human IL15 and IL18 were purchased from Peprotech, Inc. (Rocky Hill, NJ). The anti-HER2 mAb trastuzumab was provided by Genentech Inc. (San Francisco, CA). Antibodies utilized for flow cytometry include APC anti-hNKG2D (BD Biosciences, 558071), PE anti-hIFNγ (BD Biosciences, 559326), PE anti-hMICA (R&D systems, FAB1300P), and PE anti-hMICA/B (Biolegend, 320906). The anti-MICA human F(ab’)2 Ab and human IgG F(ab’)2 control utilized for neutralization studies were generated as described [30]. sMICA used in monocyte coculture experiments was generated as described [31].
Cell lines
The SK-BR-3 and MDA-MB-468 human breast adenocarcinoma cell lines were obtained from American Type Culture Collection (Manassas, VA). The C1R and C1R-MICA cell lines were provided by Dr. Veronika Groh. The identity of these cell lines was not authenticated. Cells were maintained in RPMI-1640 medium containing 10% FBS, 100 U/mL penicillin, 100 μg/mL streptomycin, and C1R-MICA cultures were supplemented with G418 sulfate (Life Technologies Inc., Rockville, MD). All cell lines used in this study were routinely tested for mycoplasma infection using the MycoAlert Mycoplasma Detection Kit (Lonza) and Plasmocin prophylactic (Invivogen) was added to all growth media at a concentration of 2.5 μg/mL. Cell lines were maintained in culture for no more than 10 passages.
Isolation of human NK cells and monocytes
PBMC and NK cells were isolated from donor leukopacks (American Red Cross, Columbus, OH) as described [15]. CD14+ monocytes were isolated using anti-CD14 magnetic beads (Miltenyi Biotec, Auburn, CA). Immune cells were cultured in 10% HAB medium.
In vitro coculture with tumor cells
SK-BR-3 and MDA-MB-468 cells were cultured in 96-well plates. Cells were treated the following day with 100 μg/mL trastuzumab for 1h. Immune cells were added to wells (2×105 NK cells/well) in 10% HAB medium supplemented with IL12 (10 ng/mL). Cocultures were plated at a 2:1 NK cell to monocyte/PBMC ratio. Supernatants were harvested and analyzed for cytokines by ELISA (R&D Systems) as described [12].
Intracellular staining for IFNγ
NK-cell IFNγ production was analyzed using an IFNγ PE-conjugated mAb and a CD56 APC-conjugated mAb as described [13].
Real-Time PCR
Cellular RNA was isolated using RNeasy Mini-kits (Qiagen, Valencia, CA). cDNA was generated with random hexamer primers and MMLV-RT according to manufacturer’s recommendations (Life Technologies Inc., Rockville, MD). Real-time PCR for huIFNγ transcript was performed using an ABI prism 7700 sequence detector (Applied Biosystems, Carlsbad, CA) with an 18S rRNA internal control (PE Applied Biosystems, Foster City, CA).
Transwell cocultures
SK-BR-3 cells were cultured in 24-well plates and treated the next day with 100 μg/ml trastuzumab for 1h. A transwell insert with a 0.4 micron filter was placed in each well, and NK cells and/or monocytes were added in media ± IL12 (10 ng/ml). Supernatants were analyzed for IFNγ content by ELISA.
MICA siRNA and monocyte transfection
Monocytes were transfected using the Amaxa Nucleofector apparatus (Amaxa Biosystem, Cologne, Germany). Cells were resuspended in Nucleofector Solution T and nucleofected with scrambled or human MICA siRNA from Ambion (Austin, TX). Cells were transferred to RPMI media supplemented with 10% FBS and M-CSF (20 ng/mL). Cy3-labeled GAPDH siRNA was used to analyze transfection efficiency.
Murine Studies
Mice received two intraperitoneal (i.p.) injections of F4/80 Ab (250 μg), isotype control Ab (250 μg), or PBS. CT-26HER2/neu cells were incubated in PBS plus 10% FBS with 4D5 (100 μg/ml) [6]. 4×106 cells in PBS containing 1 μg muIL12 were injected i.p. Serum was harvested at 24h and analyzed by ELISA. Splenocytes were collected to assess monocyte depletion by flow cytometry. For ex vivo studies, wild-type BALB/c splenocytes were cocultured with CT-26HER2/neu tumor cells. Culture supernatants were analyzed for muIFNγ by ELISA. NKG2D knockout mice were provided by Dr. David Raulet.
For the murine tumor study, mice received i.p. injections of control or clodronate-containing liposomes (1 mg/kg in 100 uL PBS) on days 0 and 4 with respect to tumor inoculation and every fourth day thereafter [32]. On day 0, mice were inoculated with 8×105 EMT6HER2/neu cells in the mammary fat pad [33]. On day 7 and every third day, mice received i.p. injections of trastuzumab and IL12 (10 mg/kg and 2.5 ng, respectively). Tumor volume = 0.5 × [(large diameter) × (small diameter)2]. Upon completion of the study, mice were sacrificed and tumors were collected. Cells were labeled with F4/80 PE-conjugated Ab and CD11b APC-conjugated Ab to evaluate monocyte depletion.
Statistics
Statistical analyses of cytokine levels were performed using Student’s t-tests. Changes in tumor volume over time were assessed via a longitudinal model. Tumor data was log transformed, and a linear mixed effects model was applied to account for correlations of observations from the same mouse.
Results
NK cells secrete immune stimulatory cytokines in response to IL12 and tumor cells
To investigate the ability of monocytes to enhance NK-cell interactions with Ab-coated tumor cells, we first evaluated NK-cell cytokine production and lytic activity in vitro in response to a stimulatory strategy: a therapeutic mAb coupled with the cytokine originally referred to as NK-cell stimulatory factor, IL12. The HER2 over-expressing breast cancer cell line SK-BR-3 or the HER2 negative MDA-MB-468 cell line were cocultured with NK cells in the presence or absence of trastuzumab and IL12. NK-cell production of IFNγ in response to trastuzumab-coated SK-BR-3 cells was enhanced in the presence of IL12, as compared to control conditions; however, this was not observed with the HER2-negative cell line (Fig. 1A). The capacity of total peripheral blood mononuclear cells (PBMC) to respond to IL12 and Ab-coated tumor cells also was tested. PBMC (plated at the same cell density as pure NK cells) secreted more IFNγ compared to NK cells alone in response to dual stimulation via Ab-coated tumor cells and IL12. This relationship held true for other NK cell-derived cytokines including TNFα and MIP-1α (Fig. 1B). On average, PBMC IFNγ production was 200% higher than that of pure NK cells. The number of NK cells within the PBMC population added to each well was approximately 4-fold less than the number of pure NK cells plated (2 × 105 cells per well). Next, PBMC IFNγ production in response to various stimuli (e.g., IL1β, IL12, IL15, IL18, and/or trastuzumab-coated cells) was compared to that of NK cells. PBMC IFNγ production was greater than that of purified NK cells for all conditions tested (Fig. 1C). The most potent individual cytokine stimulus for NK-cell IFNγ production in response to Ab-coated tumor cells was IL12. Subsequent investigation sought to uncover which cellular compartment within total PBMC could be responsible for providing a stimulatory signal to increase NK cell antitumor activity in the presence of what was considered to be a strong stimulatory strategy, namely FcR and IL12R co-activation.
Figure 1. Cytokine production is enhanced in the presence of PBMC.
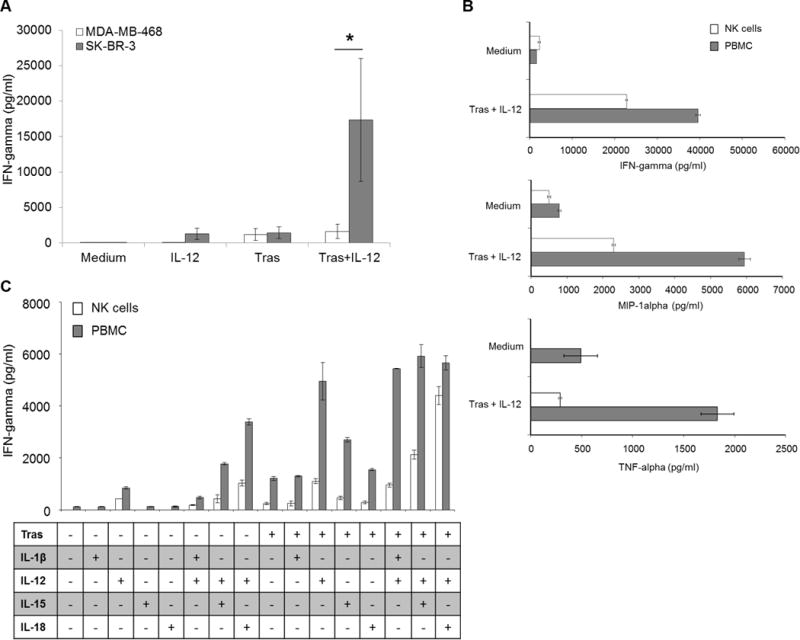
(A). NK cells were cocultured with SK-BR-3 or MDA-MB-468 tumor cells in the presence of medium, rhuIL12 (IL12), trastuzumab (Tras), or the combination. (B). NK cells or PBMC were cultured in medium alone or with trastuzumab-coated SK-BR-3 cells and IL12. (C). NK cells or PBMC were cultured in the presence of rhuIL1β, IL12, IL15 or IL18 and/or trastuzumab-coated SK-BR-3 cells. Bars represent the mean concentration ± SD of cytokine (IFNγ for A, B, C, MIP-1α for B, or TNFα for B) content from culture supernatants analyzed by ELISA. Data shown are representative of three independent experiments with similar results (a) or two experiments (b and c). *P < 0.05
Monocytes within a PBMC population enhance the NK-cell cytokine response
The impact of different immune cell populations on the NK-cell IFNγ response was investigated. Depletion of monocytes from PBMC, but not B cells or T cells, resulted in decreased IFNγ production following exposure to IL12 and trastuzumab-coated tumor cells (Fig. 2A, Supplementary Fig. S1). The ability of monocyte-depleted PBMC to induce IFNγ production in response to IL12 and Ab-coated tumor cells was restored in a dose-dependent fashion by the reintroduction of autologous monocytes (Fig. 2B). To confirm the effect of monocytes on NK-cell cytokine production, NK cells were cocultured with purified autologous monocytes. NK cells cocultured with monocytes in the presence of Ab-coated tumor cells and IL12 produced more IFNγ than NK cells alone (Fig. 2C). Monocytes produced no measureable amounts of of IFNγ. Similar results were obtained with IL15 as the cytokine stimulus (Fig. 2D). Coculture with suppressive monocyte-derived cells such as tumor-associated macrophages (TAM) or myeloid-derived suppressor cells (MDSC) generated in vitro from autologous monocytes led to a reduction in NK-cell IFNγ production in this system (Figure 2E). Thus, it appeared that monocytes, although not the direct source of IFNγ, were important for enhancement of NK-cell IFNγ production. The effect of monocyte addition was dose-dependent, as the introduction of increasing numbers of monocytes led to NK-cell IFNγ production that was equivalent to the IFNγ output of unmanipulated PBMC (Fig. 2F). The effect of monocytes on NK-cell IFNγ production was confirmed by intracellular flow cytometry (Fig. 2G). Also, IFNγ transcript levels were evaluated in dual-stimulated NK cells in the presence and absence of monocytes. Addition of monocytes to coculture enhanced NK-cell IFNγ transcript levels (Fig. 2H). These results suggest that monocytes within the PBMC population are capable of enhancing NK-cell cytokine production in response to Ab-coated tumor cells and IL12.
Figure 2. Monocytes promote NK-cell IFNγ production.
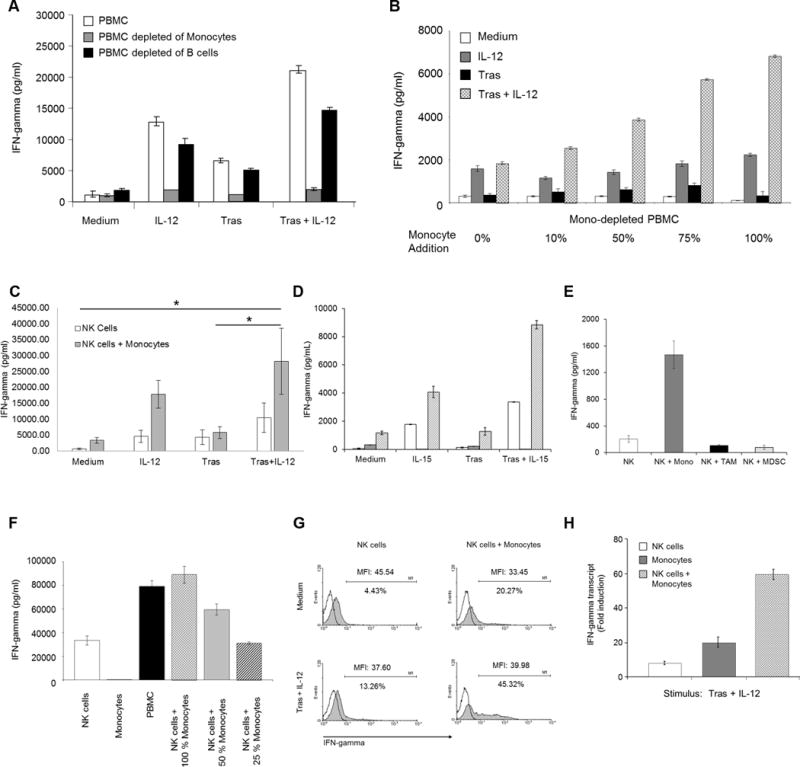
(A). PBMC (~14% CD14+ cells), PBMC depleted of monocytes (<0.03% CD14+ cells), or PBMC depleted of B cells (>98% CD19− cells) were cocultured with SK-BR-3 cells in medium, IL12, trastuzumab, or the combination. (B). Monocyte-depleted PBMC were mixed with increasing numbers of autologous monocytes and cultured with SK-BR-3 cells in the presence of medium, IL12, trastuzumab, or the combination. (C). NK cells, or NK cells plus monocytes were cultured with SK-BR-3 cells in the presence of medium, IL12, trastuzumab, or the combination. (D). NK cells were mixed with monocytes and cocultured with SK-BR-3 cells in the presence of medium, IL15, trastuzumab, or the combination. (E). NK cells were cultured in the presence of trastuzumab-coated SK-BR-3 cells and IL12, and monocytes and/or in vitro-generated autologous TAM or MDSC at a 2:1 ratio were added to the coculture [48, 49]. (F). NK cells were cultured with varying numbers of autologous monocytes in the presence of trastuzumab-coated SK-BR-3 cells and IL12. (G). NK cells alone or in the presence of monocytes were cocultured with trastuzumab-coated SK-BR-3 cells and IL12. Cells were harvested and intracellular IFNγ was examined by flow cytometry. (H). NK cells, monocytes, or both were cultured in the presence of trastuzumab-coated SK-BR-3 cells and IL12. Bars represent the mean concentration ± SD of IFNγ content from culture supernatants analyzed by ELISA (a, b, c, d, e, f) or IFNγ transcript as measured by PCR (h). Data shown are representative of two (a, b, d, f, g) or three (e) independent experiments with similar results, or the mean concentration ± SD of n = 8 donors (c). *P < 0.05
Monocytes enhance the lytic activity of NK cells
The effect of monocytes on the ability of IL12-activated NK cells to lyse Ab-coated tumor cells was examined (Fig. 3). At the 50:1 effector to target ratio, IL12-activated NK cells mediated approximately 48% lysis of trastuzumab-coated SK-BR-3 target cells (Fig. 3, top). Monocytes alone exhibited negligible cytotoxic activity (Fig. 3, middle). It has been demonstrated that monocytes are capable of ADCC 8–14 hours after priming with IFNγ; however, this was not observed in our standard 4–h chromium release assay conditions. Exposure of NK cells to monocytes (Fig. 3, bottom) prior to the addition of Ab-coated target cells and IL12 facilitated an increase in NK cell-mediated cytotoxicity to nearly 98% at the 50:1 effector to target ratio (P < 0.05).
Figure 3. Monocytes enhance NK cell ADCC.
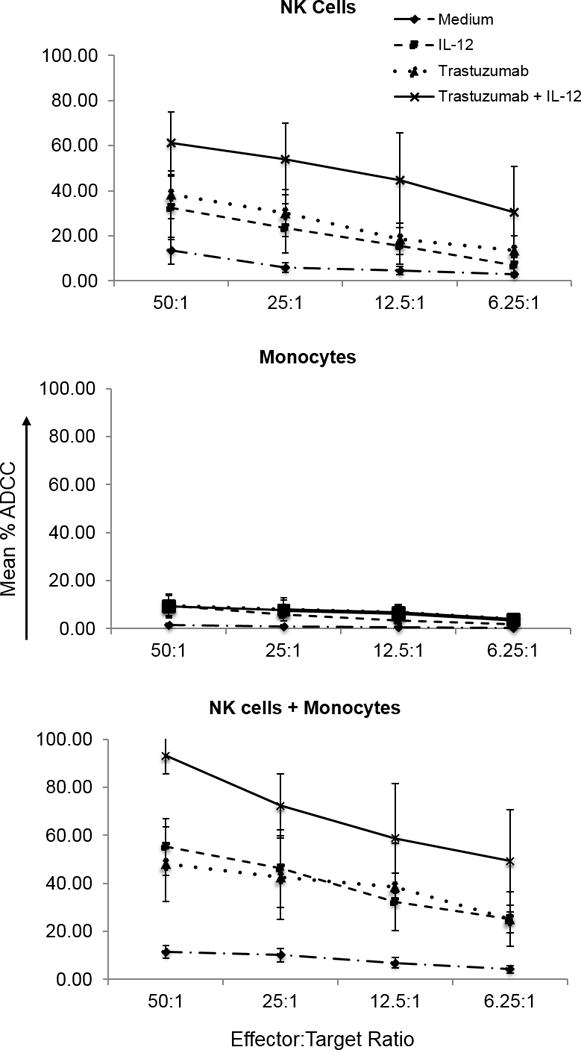
NK cells, monocytes, or both were incubated overnight in medium alone or medium containing IL12. NK cell ADCC was assessed in a standard 4-h chromium release assay with trastuzumab-coated SK-BR-3 target cells. Symbols represent target cell mean percent lysis ± SD for three independent experiments.
NK cell activation is enhanced by direct contact between NK cells and monocytes
Increased NK-cell IFNγ production upon interaction with monocytes could be due to direct cell-to-cell contact or the diffusion of soluble mediators. To determine whether cell-to-cell contact was required for stimulation, a transwell coculture experiment was performed. When NK cells and monocytes were separated by a cell-impermeable membrane, NK cells exposed to trastuzumab-coated tumor cells and IL12 produced normal amounts of IFNγ. However, coculture of NK cells and monocytes in the same compartment (permitting contact) led to significantly greater IFNγ production (Fig. 4A, P = 0.03). Figure 4B demonstrates the same experiment with a full set of controls, which was challenging to perform due to the requirement for a large number of NK cells from a single donor. The effect of monocytes on NK-cell cytokine production in Figure 4B, although substantial and reproducible, was less than in previous experiments possibly due to the physical characteristics of larger transwell cultures. In summary, monocyte enhancement of NK-cell IFNγ production in response to Ab-coated tumor cells and IL12 requires cell-to-cell contact, and this effect is lost when monocytes are separated from NK cells by a cell-impermeable membrane.
Figure 4. Monocytes provide contact-dependent NK cell activating signals.
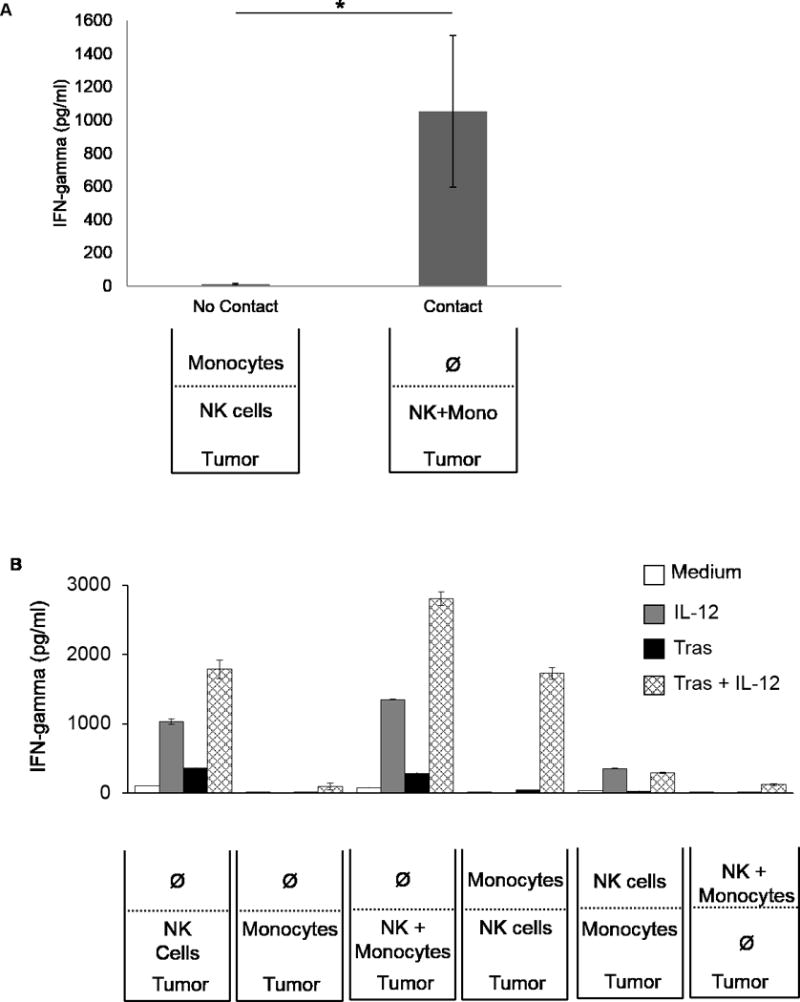
(A). NK cells and monocytes from three separate leukopacks were cultured in transwell plates. (B). NK cells and monocytes from one representative donor were cultured in various combinations in a transwell plate. Bars represent the mean concentration ± SD of supernatant IFNγ. *P < 0.05
Ligand interactions essential for monocyte enhancement of NK-cell IFNγ production
Bauer et al. reported that interactions between NK cell NKG2D and its ligands MICA/B or ULBPs, expressed by tumor cells, enhances NK cell cytokine production [18]. However, NKG2D ligands also may be expressed by cells of the innate immune system [34, 35]. Thus, we hypothesized that monocytes could activate NK cells via expression of NKG2D ligands. Monocyte MICA surface expression was measured by flow cytometry. Though variability exists in NKG2D ligand expression among healthy donors, substantial MICA expression was detected in freshly isolated monocytes (Supplementary Fig. S2). Further, it was shown that MICA expression could be upregulated following stimulation of monocytes with IFNα or LPS (Fig. 5A). LPS treatment of monocytes enhanced their ability to augment NK-cell IFNγ production (Fig. 5B). To further explore the interaction between NKG2D and MICA, the MICA overexpressing cell line C1R-MICA was employed and was confirmed to selectively over-express MICA but not other NKG2D ligands (Supplementary Fig. S3). Coculture of NK cells with C1R-MICA cells in the presence of trastuzumab-coated SK-BR-3 cells and IL12 led to increased NK-cell IFNγ production that was comparable to that observed with the addition of monocytes. Coculture of NK cells with the parental cell line C1R (which lacks MICA expression) did not modify NK-cell IFNγ production (Fig. 5C). Introduction of an anti-MICA neutralizing Ab abrogated the ability of C1R-MICA cells to enhance NK-cell IFNγ production (Fig. 5D).
Figure 5. Monocyte expression of the NKG2D ligand MICA promotes NK cell antitumor activity.
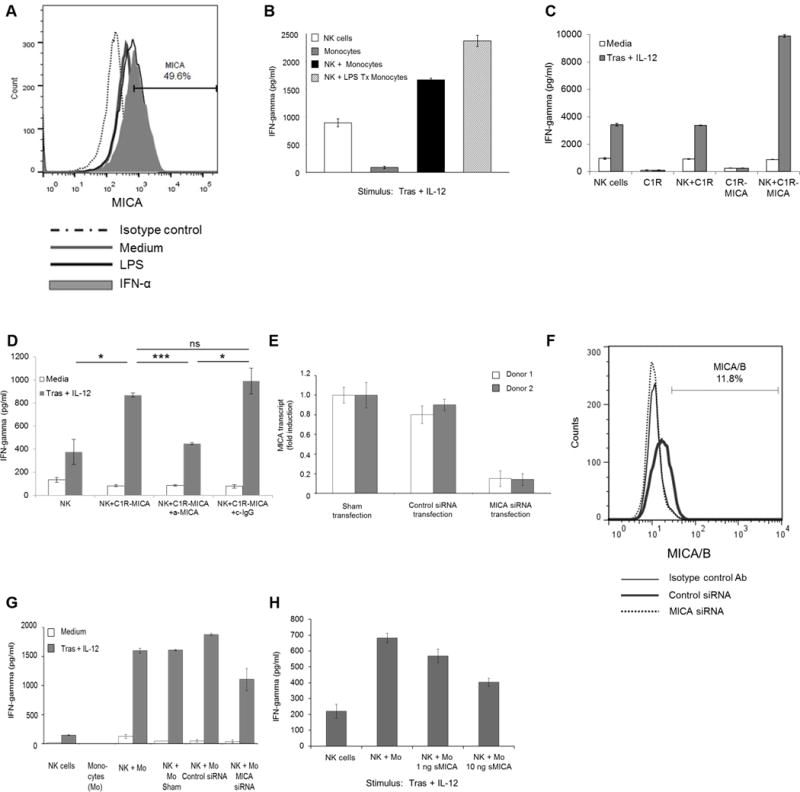
(A). Monocyte MICA expression at baseline (medium) and following treatment with LPS or IFNα was evaluated by flow cytometry. (B). Untreated and LPS-treated monocytes were cocultured with NK cells, trastuzumab-coated SK-BR-3 cells, and IL12. (C). NK cells were cocultured with C1R or C1R-MICA cells in the presence of trastuzumab-coated SK-BR-3 cells and IL12. (D). C1R-MICA cells were incubated with control IgG or anti-MICA Ab. C1R-MICA cells then were cocultured with NK cells, trastuzumab-coated SK-BR-3 cells, and IL12. (E). Monocytes were transfected with MICA siRNA or scrambled control siRNA. Transfected cells were analyzed for MICA transcript by real-time PCR or (F) MICA/B surface expression was examined by flow cytometry. (G). Monocytes with downregulated levels of MICA were cocultured with NK cells in the presence of trastuzumab-coated SK-BR-3 cells and IL12. (H). NK cells were pre-treated with soluble MICA (sMICA) prior to coculture with monocytes in the presence of trastuzumab-coated SK-BR-3 cells and IL12. Bars represent the mean concentration ± SD of IFNγ content from supernatants analyzed by ELISA (b, c, d, g, h). Data shown for each panel are representative of two (b, c, g), three (a, h) independent experiments with similar results or the mean concentration ± SD of n = 3 donors (d). *P < 0.05, ***P < 0.001, ns - not significant.
Experiments involving an anti-MICA neutralizing Ab were difficult to optimize with donor monocytes and NK cells due to FcR expression. An F(ab’)2 against MICA was designed and utilized in an attempt to block NKG2D-MICA interactions; however, unintended NK cell activation persisted. An anti-MICA siRNA was employed to study the interaction between NK cell NKG2D and monocyte MICA. MICA expression levels were down-regulated in monocytes using this siRNA approach. Reduced MICA expression at the transcript level and on the surface of monocytes was confirmed by real time PCR and flow cytometry (Fig. 5E and F).
Down-regulation of MICA impaired the ability of monocytes to augment NK-cell cytokine production and led to reduction in IFNγ levels in response to Ab-coated tumor cells and IL12 (Fig. 5G). The interaction between NK-cell NKG2D and monocyte MICA also was disrupted by pre-treating NK cells with soluble MICA (sMICA) to block NKG2D receptor availability prior to coculture with monocytes [36]. This strategy resulted in a dose-dependent decrease in IFNγ production, supporting the hypothesis that direct NKG2D-MICA interactions between NK cells and monocytes promote NK-cell activity (Fig. 5H). No correlation was found between MICA expression levels on donor monocytes and IFNγ production from NK cells indicating that although MICA expression on monocytes is necessary for enhanced NK cell activation, the level to which MICA is expressed is not as important (Supplementary Fig. S4).
Modulation of NKG2D expression on NK cells
It was confirmed that NK cells express NKG2D, and expression may be upregulated upon NK-cell stimulation via IL12 or IL15 (Fig. 6A) [37]. Pretreatment of NK cells with IL15 (followed by an overnight rest) increased NKG2D expression and enhanced NK-cell IFNγ production in the presence of monocytes (Fig. 6B). Additionally, NK cell NKG2D expression was not decreased following interactions with MICA expressing cells in vitro (Supplementary Fig. S5). Further, an anti-NKG2D neutralizing Ab was employed to block activation of this receptor [38]. NKG2D blockade reduced NK-cell IFNγ production in response to Ab-coated tumor cells and IL12 in the presence of C1R-MICA cells (Fig. 6C) as well as monocytes (Fig. 6D).
Figure 6. NK cells interact with MICA on monocytes via the NKG2D receptor.
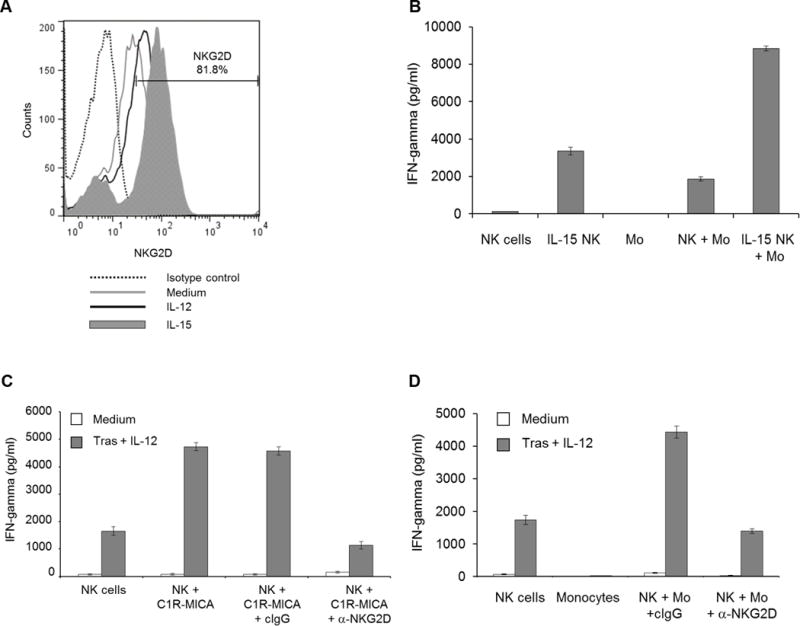
(A). NK cell NKG2D expression was evaluated at baseline (medium) and following stimulation via IL12 or IL15. (B). NK cells were treated with IL15, rested overnight, then added to coculture with monocytes, trastuzumab-coated SK-BR-3 cells, and IL12. (C). NK cells were incubated with a control IgG or anti-NKG2D Ab. NK cells then were cultured in the presence trastuzumab-coated SK-BR-3 cells, IL12, and C1R-MICA cells or (D) monocytes. Bars represent the mean concentration ± SD of IFNγ content from supernatants analyzed by ELISA. Data shown for each panel are representative of two independent experiments with similar results.
Murine NK-cell IFN-γ production is enhanced in the presence of monocytes
Next, experiments were conducted using murine models. Though mice do not express MIC-derived molecules, they do express similar MHC class I-like molecules including RAE-1, H60, and MULT-1 that engage murine NKG2D and promote NK-cell activation [39]. Guerra et al. showed in in murine models that tumor immunosurveillance is inhibited in NKG2D-deficient mice [40]. Given that in vitro experiments in our laboratory demonstrated a relationship between NK cells and monocytes, we sought to evaluate whether interruption of this interaction would impact the NK-cell response to mAb therapy in murine models as well.
Splenocytes from wild-type mice were cultured in the presence of IL12 and 4D5 (a murine anti-HER2 mAb)-coated HER2-positive murine tumor cells. Splenocytes depleted of NK cells produced less IFNγ, identifying NK cells as the source of IFNγ, as described by our group previously (Fig. 7A, [6]). Clodronate-containing liposomes were employed as a means of depleting mice of monocytes in vivo. Splenocytes isolated from mice treated with control liposomes retained normal monocyte numbers and produced IFNγ ex vivo in response to IL12 and 4D5-coated tumor cells. In contrast, treatment with clodronate-containing liposomes led to a 60% reduction in monocyte numbers and a reduction in NK-cell IFNγ production (Fig. 7B). An F4/80 antibody was employed as an additional strategy to deplete monocytes. Control mice treated with an isotype Ab retained normal monocyte numbers, and the amount of IFNγ in the serum increased in response to i.p. administration of IL12 and 4D5-coated tumor cells. However, treatment with an F4/80 Ab led to a 61% reduction in monocytes in the spleen and resulted in a 2-fold reduction in serum IFNγ levels in response to dual stimulation (Fig. 7C). Next, mice rendered genetically deficient in NKG2D were employed. Splenocytes were harvested from wild-type and NKG2D knock-out mice and cultured in the presence of murine IL12 and 4D5-coated CT26HER2/neu tumor cells. IFNγ production was inhibited in mice lacking the NKG2D receptor (Fig. 7D). Lastly, a study was conducted in which mice bearing HER2-positive tumors were treated with trastuzumab and IL12 along with systemic administration of clodronate-containing liposomes or control liposomes. Treatment of mice with clodronate-containing liposomes led to significantly larger tumors as compared to mice receiving control liposomes (P < 0.05, Fig. 7E).
Figure 7. Monocytes enhance NK cell function in vivo.
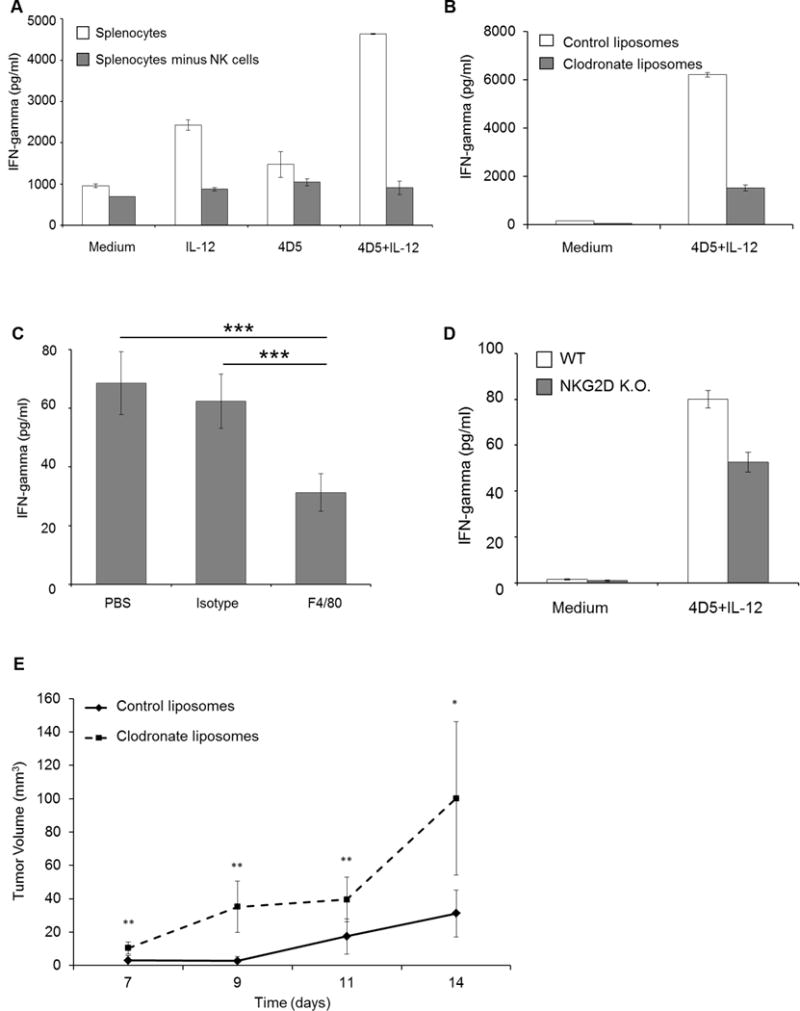
(A). Murine splenocytes were depleted of NK cells and cocultured with CT26Her2/neu tumor cells in the presence of medium, IL12, 4D5, or the combination. Bars represent the mean concentration ± SD of IFNγ content from supernatants analyzed by ELISA, n = 2 mice. (B). Splenocytes from wild-type BALB/c mice treated with clodronate-containing or control liposomes were cocultured with 4D5-coated CT26Her2/neu tumor cells and IL12. Bars represent the mean concentration ± SD of IFNγ content, n = 2 mice. (C). BALB/c mice were treated with PBS, an isotype control Ab, or an F4/80 Ab and were injected i.p. with 4D5-coated CT26Her2/neu tumor cells and IL12. Serum was collected 24h later and analyzed for IFNγ content by ELISA. The means ± SD are shown (n = 5 for each condition). (D). Splenocytes were isolated from wild-type (WT) and NKG2D-deficient mice (NKG2D K.O.) and cocultured with 4D5-coated CT26HER2/neu tumor cells and IL12. Bars represent the mean concentration ± SD of IFNγ content, n = 3. (E). Trastuzumab and IL12 were administered to EMT6HER2/neu tumor-bearing mice that were treated with clodronate-containing or control liposomes. Average tumor volume ± SEM is shown (n = 5 mice for each group). *P < 0.05, **P < 0.02 and ***P < 0.001.
Discussion
Here we investigated the role of NK cell-monocyte interactions in the setting of mAb therapy for cancer. We have demonstrated previously that NK cell-derived IFNγ serves as a key mediator of the immune response to cancer. NK cells release IFNγ following costimulation via Ab-coated tumor cells and IL12, a therapeutic strategy proven to elicit NK cell activity in vitro and in vivo. Now, it has been shown that monocytes enhance NK-cell IFNγ production in response to a stimulus. This stimulatory interaction requires direct cell-to-cell contact between NK cells and monocytes and is mediated primarily by monocyte MICA engagement of the NKG2D receptor on NK cells. Lastly, it was demonstrated in murine models that the immune response to combination therapy with trastuzumab and IL12 is enhanced through NK cell-monocyte interactions. Taken together, these results suggest that NK cells and monocytes in the tumor microenvironment participate in crosstalk to optimize Ab-mediated antitumor activity, and mechanisms to promote these interactions may enhance the response to tumor-specific mAbs.
Interactions between NK cells and monocytes promote the immune response against invading pathogens and transformed cancer cells. However, the role of monocytes in modulating NK-cell activity in the setting of mAb therapy is not well understood. We noted that exposure of PBMC to various NK stimulatory conditions led to more IFNγ production than that of NK cells alone. Monocytes were the cells within the PBMC population responsible for this boost in NK-cell cytokine production. Others have demonstrated that reciprocal activation between NK cells and monocytes occurs via contact in the setting of infection as well as chronic arthritis [41, 42]. However, the molecules responsible for this crosstalk were not determined. In the present study, a key interface between NK cells and monocytes, the NKG2D-MICA axis, was shown to be responsible for monocyte-mediated enhancement of NK cell antitumor activity. These results suggest that NK-cell NKG2D and monocyte MICA interactions play an important role in the immune response to mAb therapy.
Few studies have delved into the relationship between NK cells and monocytes in regard to cell-mediated cytotoxicity against tumor cells [43]. We now show that interactions with monocytes promote NK cell-mediated cytotoxic activity. Bhatnagar et al. described reciprocal crosstalk between NK cells and monocytes in the setting of viral infection, concluding that monocytes enhanced NK cell cytotoxicity against HIV-1 infected cells via direct contact in an FcγRII (CD32)-dependent manner [44]. The role of NK cell recognition and lysis of NKG2D-ligand expressing tumor cells has been described previously [45]. Pende et al. analyzed tumor expression levels of the NKG2D ligands MICA and ULBPs in melanoma, leukemia, neuroblastoma, and various carcinoma cell lines and correlated expression levels to increased tumor cell susceptibility to NK cell-mediated lysis. The impact of NKG2D ligand expression by circulating immune cells was not explored. Wang et al. evaluated IFNγ pre-treated monocytes and determined that this strategy induced monocyte expression of MIC molecules and membrane-bound IL15 [46]. These molecules promoted NK-cell IFNγ production and cytotoxicity against lymphoblastoid cells. This was observed only with IFNγ pre-treated monocytes in vitro, and NK-cell activity was not enhanced in the presence of freshly isolated monocytes, as was the case in the present study. Also, Wang et al. utilized allogeneic NK cell-monocyte cocultures at a 5:1 NK cell to monocyte ratio, whereas the current study investigated autologous NK cells and monocytes cultured at a 2:1 ratio, modeling what might be found in vivo. Kloss et al demonstrated that MICA-positive, LPS-activated monocytes enhanced NK cell cytokine production in vitro. The presence of monocytes led to increased NK cell proliferation, regardless of whether monocytes were untreated or primed with LPS [35]. The current study demonstrates that in the setting of mAb therapy for cancer, interactions between MICA-positive monocytes and NKG2D-expressing NK cells facilitate NK-cell cytokine production and ADCC against Ab-coated tumor cells both in vitro and in murine models.
A recent study by Crane et al. described that myeloid cells, including monocytes isolated from glioblastoma multiforme (GBM) patients, expressed NKG2D ligands [47]. It was shown that glioma cell line-derived lactate dehydrogenase-containing supernatants had the capacity to upregulate monocyte NKG2D ligand expression and promote NK-cell degranulation and cytokine production in vitro. The authors proposed that immune evasion in GBM patients may be due to NKG2D-dependent NK cell exhaustion. In contrast, this study demonstrated that interruption of NK cell-monocyte crosstalk in vivo inhibited the NK cell effector response against Ab-coated breast cancer cells. These contrasting results emphasize the need to further explore the possible effects of NKG2D-ligand expression by myeloid cells.
This study has provided evidence that monocytes promote NK-cell cytokine production and tumor cell lysis in response to FcR and IL12R stimulation, an effective dual stimulation regimen with proven antitumor activity. The antitumor effects of mAb therapy may be enhanced by strategies aimed at maximizing monocyte expression of NK-cell activating molecules.
Supplementary Material
Acknowledgments
The authors thank The Ohio State University Comprehensive Cancer Center’s Analytical Cytometry Shared Resource for providing equipment to facilitate flow cytometry analyses.
Financial Support: This work was supported by the Susan G. Komen Breast Cancer Foundation Dissertation Research Award (RP), the OSUCCC Pelotonia Graduate Fellowship (ARC), and the NIH grants T32 GM068412 (MCD) and P01 CA95426 (WEC).
Footnotes
Conflict of Interest Statement: The authors have declared that no conflict of interest exists.
References
- 1.Bhave NS, Carson WEI. The role of natural killer cells in innate immunity. Regulation of Innate Immune Function Editors: Clay Marsh, Susheela Tridandapani and Melissa Piper. 2010:141–64. [Google Scholar]
- 2.Caligiuri MA. Human natural killer cells. Blood. 2008;112:461–9. doi: 10.1182/blood-2007-09-077438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Carson WE, Fehniger TA, Haldar S, Eckhert K, Lindemann MJ, Lai CF, et al. A potential role for interleukin-15 in the regulation of human natural killer cell survival. The Journal of clinical investigation. 1997;99:937–43. doi: 10.1172/JCI119258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Carson WE, Parihar R, Lindemann MJ, Personeni N, Dierksheide J, Meropol NJ, et al. Interleukin-2 enhances the natural killer cell response to Herceptin-coated Her2/neu-positive breast cancer cells. European journal of immunology. 2001;31:3016–25. doi: 10.1002/1521-4141(2001010)31:10<3016::aid-immu3016>3.0.co;2-j. [DOI] [PubMed] [Google Scholar]
- 5.Badgwell B, Parihar R, Magro C, Dierksheide J, Russo T, Carson WE., 3rd Natural killer cells contribute to the lethality of a murine model of Escherichia coli infection. Surgery. 2002;132:205–12. doi: 10.1067/msy.2002.125311. [DOI] [PubMed] [Google Scholar]
- 6.Jaime-Ramirez AC, Mundy-Bosse BL, Kondadasula S, Jones NB, Roda JM, Mani A, et al. IL-12 enhances the antitumor actions of trastuzumab via NK-cell IFN-γ production. J Immunol. 2011;186:3401–9. doi: 10.4049/jimmunol.1000328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Carson WE, Ross ME, Baiocchi RA, Marien MJ, Boiani N, Grabstein K, et al. Endogenous production of interleukin 15 by activated human monocytes is critical for optimal production of interferon-gamma by natural killer cells in vitro. The Journal of clinical investigation. 1995;96:2578–82. doi: 10.1172/JCI118321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Vivier E, Nunès JA, Vély F. Natural Killer Cell Signaling Pathways. Science. 2004;306:1517–9. doi: 10.1126/science.1103478. [DOI] [PubMed] [Google Scholar]
- 9.Clynes RA, Towers TL, Presta LG, Ravetch JV. Inhibitory Fc receptors modulate in vivo cytoxicity against tumor targets. Nature medicine. 2000;6:443–6. doi: 10.1038/74704. [DOI] [PubMed] [Google Scholar]
- 10.Cartron G. Therapeutic activity of humanized anti-CD20 monoclonal antibody and polymorphism in IgG Fc receptor Fc[gamma]RIIIa gene. Blood. 2002;99:754–8. doi: 10.1182/blood.v99.3.754. [DOI] [PubMed] [Google Scholar]
- 11.Weng WK, Levy R. Two immunoglobulin G fragment C receptor polymorphisms independently predict response to rituximab in patients with follicular lymphoma. J Clin Oncol. 2003;21:3940–7. doi: 10.1200/JCO.2003.05.013. [DOI] [PubMed] [Google Scholar]
- 12.Parihar R, Dierksheide J, Hu Y, Carson WE. IL-12 enhances the natural killer cell cytokine response to Ab-coated tumor cells. The Journal of clinical investigation. 2002;110:983–92. doi: 10.1172/JCI15950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Parihar R, Nadella P, Lewis A, Jensen R, De Hoff C, Dierksheide JE, et al. A Phase I Study of Interleukin 12 with Trastuzumab in Patients with Human Epidermal Growth Factor Receptor-2-Overexpressing Malignancies. Clinical Cancer Research. 2004;10:5027–37. doi: 10.1158/1078-0432.CCR-04-0265. [DOI] [PubMed] [Google Scholar]
- 14.Roda JM, Parihar R, Magro C, Nuovo GJ, Tridandapani S, Carson WE., 3rd Natural killer cells produce T cell-recruiting chemokines in response to antibody-coated tumor cells. Cancer research. 2006;66:517–26. doi: 10.1158/0008-5472.CAN-05-2429. [DOI] [PubMed] [Google Scholar]
- 15.Kondadasula SV, Roda JM, Parihar R, Yu J, Lehman A, Caligiuri MA, et al. Colocalization of the IL-12 receptor and FcgammaRIIIa to natural killer cell lipid rafts leads to activation of ERK and enhanced production of interferon-gamma. Blood. 2008;111:4173–83. doi: 10.1182/blood-2007-01-068908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Lanier LL. Up on the tightrope: natural killer cell activation and inhibition. Nature immunology. 2008;9:495–502. doi: 10.1038/ni1581. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Groh V, Bahram S, Bauer S, Herman A, Beauchamp M, Spies T. Cell stress-regulated human major histocompatibility complex class I gene expressed in gastrointestinal epithelium. Proceedings of the National Academy of Sciences. 1996;93:12445–50. doi: 10.1073/pnas.93.22.12445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Bauer S, Groh V, Wu J, Steinle A, Phillips JH, Lanier LL, et al. Activation of NK cells and T cells by NKG2D, a receptor for stress-inducible MICA. Science. 1999;285:727–9. doi: 10.1126/science.285.5428.727. [DOI] [PubMed] [Google Scholar]
- 19.Upshaw JL, Arneson LN, Schoon RA, Dick CJ, Billadeau DD, Leibson PJ. NKG2D-mediated signaling requires a DAP10-bound Grb2-Vav1 intermediate and phosphatidylinositol-3-kinase in human natural killer cells. Nature immunology. 2006;7:524–32. doi: 10.1038/ni1325. [DOI] [PubMed] [Google Scholar]
- 20.Wu J, Song Y, Bakker AB, Bauer S, Spies T, Lanier LL, et al. An activating immunoreceptor complex formed by NKG2D and DAP10. Science. 1999;285:730–2. doi: 10.1126/science.285.5428.730. [DOI] [PubMed] [Google Scholar]
- 21.Cerwenka A, Lanier LL. Ligands for natural killer cell receptors: redundancy or specificity. Immunol Rev. 2001;181:158–69. doi: 10.1034/j.1600-065x.2001.1810113.x. [DOI] [PubMed] [Google Scholar]
- 22.Amakata Y, Fujiyama Y, Andoh A, Hodohara K, Bamba T. Mechanism of NK cell activation induced by coculture with dendritic cells derived from peripheral blood monocytes. Clinical and experimental immunology. 2001;124:214–22. doi: 10.1046/j.1365-2249.2001.01550.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Fernandez NC, Flament C, Crepineau F, Angevin E, Vivier E, Zitvogel L. Dendritic cells (DC) promote natural killer (NK) cell functions: dynamics of the human DC/NK cell cross talk. European cytokine network. 2002;13:17–27. [PubMed] [Google Scholar]
- 24.Cooper MA, Fehniger TA, Fuchs A, Colonna M, Caligiuri MA. NK cell and DC interactions. Trends in immunology. 2004;25:47–52. doi: 10.1016/j.it.2003.10.012. [DOI] [PubMed] [Google Scholar]
- 25.Della Chiesa M, Romagnani C, Thiel A, Moretta L, Moretta A. Multidirectional interactions are bridging human NK cells with plasmacytoid and monocyte-derived dendritic cells during innate immune responses. Blood. 2006;108:3851–8. doi: 10.1182/blood-2006-02-004028. [DOI] [PubMed] [Google Scholar]
- 26.Cooper MA, Fehniger TA, Turner SC, Chen KS, Ghaheri BA, Ghayur T, et al. Human natural killer cells: a unique innate immunoregulatory role for the CD56(bright) subset. Blood. 2001;97:3146–51. doi: 10.1182/blood.v97.10.3146. [DOI] [PubMed] [Google Scholar]
- 27.DeMarco RA, Fink MP, Lotze MT. Monocytes promote natural killer cell interferon gamma production in response to the endogenous danger signal HMGB1. Molecular immunology. 2005;42:433–44. doi: 10.1016/j.molimm.2004.07.023. [DOI] [PubMed] [Google Scholar]
- 28.Carson WE, Giri JG, Lindemann MJ, Linett ML, Ahdieh M, Paxton R, et al. Interleukin (IL) 15 is a novel cytokine that activates human natural killer cells via components of the IL-2 receptor. The Journal of experimental medicine. 1994;180:1395–403. doi: 10.1084/jem.180.4.1395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Broudy VC, Kaushansky K, Segal GM, Harlan JM, Adamson JW. Tumor necrosis factor type alpha stimulates human endothelial cells to produce granulocyte/macrophage colony-stimulating factor. Proceedings of the National Academy of Sciences. 1986;83:7467–71. doi: 10.1073/pnas.83.19.7467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Groh V, Steinle A, Bauer S, Spies T. Recognition of Stress-Induced MHC Molecules by Intestinal Epithelial γδ T Cells. Science. 1998;279:1737–40. doi: 10.1126/science.279.5357.1737. [DOI] [PubMed] [Google Scholar]
- 31.Groh V, Wu J, Yee C, Spies T. Tumour-derived soluble MIC ligands impair expression of NKG2D and T-cell activation. Nature. 2002;419:734–8. doi: 10.1038/nature01112. [DOI] [PubMed] [Google Scholar]
- 32.Bacci M, Capobianco A, Monno A, Cottone L, Di Puppo F, Camisa B, et al. Macrophages are alternatively activated in patients with endometriosis and required for growth and vascularization of lesions in a mouse model of disease. Am J Pathol. 2009;175:547–56. doi: 10.2353/ajpath.2009.081011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Cho HM, Rosenblatt JD, Tolba K, Shin SJ, Shin DS, Calfa C, et al. Delivery of NKG2D ligand using an anti-HER2 antibody-NKG2D ligand fusion protein results in an enhanced innate and adaptive antitumor response. Cancer research. 2010;70:10121–30. doi: 10.1158/0008-5472.CAN-10-1047. [DOI] [PubMed] [Google Scholar]
- 34.Zwirner NW, Fernández-Viña MA, Stastny P. MICA, a new polymorphic HLA-related antigen, is expressed mainly by keratinocytes, endothelial cells, and monocytes. Immunogenetics. 1998;47:139–48. doi: 10.1007/s002510050339. [DOI] [PubMed] [Google Scholar]
- 35.Kloss M, Decker P, Baltz KM, Baessler T, Jung G, Rammensee HG, et al. Interaction of monocytes with NK cells upon Toll-like receptor-induced expression of the NKG2D ligand MICA. J Immunol. 2008;181:6711–9. doi: 10.4049/jimmunol.181.10.6711. [DOI] [PubMed] [Google Scholar]
- 36.Groh V, Wu J, Yee C, Spies T. Tumour-derived soluble MIC ligands impair expression of NKG2D and T-cell activation. Nature. 2002;419:734–8. doi: 10.1038/nature01112. [DOI] [PubMed] [Google Scholar]
- 37.Roberts AI, Lee L, Schwarz E, Groh V, Spies T, Ebert EC, et al. NKG2D receptors induced by IL-15 costimulate CD28-negative effector CTL in the tissue microenvironment. J Immunol. 2001;167:5527–30. doi: 10.4049/jimmunol.167.10.5527. [DOI] [PubMed] [Google Scholar]
- 38.Kwong KY, Baskar S, Zhang H, Mackall CL, Rader C. Generation, affinity maturation, and characterization of a human anti-human NKG2D monoclonal antibody with dual antagonistic and agonistic activity. J Mol Biol. 2008;384:1143–56. doi: 10.1016/j.jmb.2008.09.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Zafirova B, Mandarić S, Antulov R, Krmpotić A, Jonsson H, Yokoyama WM, et al. Altered NK cell development and enhanced NK cell-mediated resistance to mouse cytomegalovirus in NKG2D-deficient mice. Immunity. 2009;31:270–82. doi: 10.1016/j.immuni.2009.06.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Guerra N, Tan YX, Joncker NT, Choy A, Gallardo F, Xiong N, et al. NKG2D-deficient mice are defective in tumor surveillance in models of spontaneous malignancy. Immunity. 2008;28:571–80. doi: 10.1016/j.immuni.2008.02.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Newman KC, Korbel DS, Hafalla JC, Riley EM. Cross-talk with myeloid accessory cells regulates human natural killer cell interferon-gamma responses to malaria. PLoS Pathog. 2006;2:e118. doi: 10.1371/journal.ppat.0020118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Dalbeth N, Gundle R, Davies RJ, Lee YC, McMichael AJ, Callan MF. CD56bright NK cells are enriched at inflammatory sites and can engage with monocytes in a reciprocal program of activation. J Immunol. 2004;173:6418–26. doi: 10.4049/jimmunol.173.10.6418. [DOI] [PubMed] [Google Scholar]
- 43.Deguine J, Breart B, Lemaître F, Bousso P. Cutting edge: tumor-targeting antibodies enhance NKG2D-mediated NK cell cytotoxicity by stabilizing NK cell-tumor cell interactions. J Immunol. 2012;189:5493–7. doi: 10.4049/jimmunol.1202065. [DOI] [PubMed] [Google Scholar]
- 44.Bhatnagar N, Ahmad F, Hong HS, Eberhard J, Lu IN, Ballmaier M, et al. FcγRIII (CD16)-mediated ADCC by NK cells is regulated by monocytes and FcγRII (CD32) European journal of immunology. 2014;44:3368–79. doi: 10.1002/eji.201444515. [DOI] [PubMed] [Google Scholar]
- 45.Pende D, Rivera P, Marcenaro S, Chang CC, Biassoni R, Conte R, et al. Major histocompatibility complex class I-related chain A and UL16-binding protein expression on tumor cell lines of different histotypes: analysis of tumor susceptibility to NKG2D-dependent natural killer cell cytotoxicity. Cancer research. 2002;62:6178–86. [PubMed] [Google Scholar]
- 46.Wang H, Ruan Z, Wang Y, Han J, Fu X, Zhao T, et al. MHC class I chain-related molecules induced on monocytes by IFN-gamma promote NK cell activation. Molecular immunology. 2008;45:1548–56. doi: 10.1016/j.molimm.2007.10.007. [DOI] [PubMed] [Google Scholar]
- 47.Crane CA, Austgen K, Haberthur K, Hofmann C, Moyes KW, Avanesyan L, et al. Immune evasion mediated by tumor-derived lactate dehydrogenase induction of NKG2D ligands on myeloid cells in glioblastoma patients. Proc Natl Acad Sci U S A. 2014;111:12823–8. doi: 10.1073/pnas.1413933111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Lechner MG, Liebertz DJ, Epstein AL. Characterization of cytokine-induced myeloid-derived suppressor cells from normal human peripheral blood mononuclear cells. J Immunol. 2010;185:2273–84. doi: 10.4049/jimmunol.1000901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Rey-Giraud F, Hafner M, Ries CH. In vitro generation of monocyte-derived macrophages under serum-free conditions improves their tumor promoting functions. PLoS One. 2012;7:e42656. doi: 10.1371/journal.pone.0042656. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.