

Abstract
Rabies remains an important worldwide health problem. Newcastle disease virus (NDV) was developed as a vaccine vector in animals by using a reverse genetics approach. Previously, our group generated a recombinant NDV (LaSota strain) expressing the complete rabies virus G protein (RVG), named rL-RVG. In this study, we constructed the variant rL-RVGTM, which expresses a chimeric rabies virus G protein (RVGTM) containing the ectodomain of RVG and the transmembrane domain (TM) and a cytoplasmic tail (CT) from the NDV fusion glycoprotein to study the function of RVG's TM and CT. The RVGTM did not detectably incorporate into NDV virions, though it was abundantly expressed at the surface of infected BHK-21 cells. Both rL-RVG and rL-RVGTM induced similar levels of NDV virus-neutralizing antibody (VNA) after initial and secondary vaccination in mice, whereas rabies VNA induction by rL-RVGTM was markedly lower than that induced by rL-RVG. Though rL-RVG could spread from cell to cell like that in rabies virus, rL-RVGTM lost this ability and spread in a manner similar to the parental NDV. Our data suggest that the TM and CT of RVG are essential for its incorporation into NDV virions and for spreading of the recombinant virus from the initially infected cells to surrounding cells.
Keywords: Newcastle disease virus, antibody response, chimeric rabies glycoprotein, rabies virus, viral vector
Introduction
Rabies is a zoonotic disease that causes severe nerve damage in humans and mammals [8]. Rabies virus (RV) is an enveloped virus from the genus Lyssavirus of the Rhabdoviridae family. The RV virion is composed of five structural proteins [9,11,15,16,24,25], and its G protein binds to receptors at the cell surface and undergoes a pH-dependent conformational change to mediate membrane fusion and endocytosis, allowing RV to enter cells [1]. Rabies virus G protein (RVG) is also the main protein responsible for inducing and reacting with virus-neutralizing antibody (VNA), which is useful as a marker of productive immunization [35].
Newcastle disease virus (NDV) causes severe disease symptoms in all species of birds [7]. Some NDV strains were developed as live-virus vectors by using a reverse genetics approach [3,17,18,20,27,29,32]. In order to find an effective, safe, and affordable RV vaccine, our group has generated rL-RVG, which is an NDV vector (avirulent NDV LaSota strain) expressing RVG [19]. Our results showed that RVG is efficiently incorporated into the NDV virion, and rL-RVG can spread on BHK-21 cells in a manner similar to that of RV but different from NDV's manner of spreading. Safe in cats and dogs, L-RVG can induce a substantial and protective rabies virus neutralization antibody in intramuscular-vaccined animals [19]. To determine the function of the transmembrane domain I and cytoplasmic tail (CT) of RVG, we generated a recombinant chimeric RGV-NDV variant, in which the ectodomain of RVG has been fused with the TM and CT of the NDV F protein. In the present study, we evaluated the influence of the RVG TM and CT on immune responses and incorporation into NDV.
Materials and Methods
Cells and viruses
BHK-21 cells were grown in Dulbecco's modified Eagle medium containing 5% fetal bovine serum. Our group generated the parental NDV vector of the LaSota vaccine strain (rL) and rL-RVG, which is a recombinant NDV strain expressing RVG [19]. A modified vaccinia virus expressing T7 RNA polymerase [36] was grown and ittered in primary chicken embryo fibroblasts. Recombinant RV and NDV expressing enhanced green fluorescent protein were generated by our laboratory group.
Construction of recombinant NDV expressing chimeric rabies virus G protein (RVGTM)
A RVGTM gene containing sequences encoding the TM domain and CT of the NDV F protein was constructed by gene synthesis. We introduced the modified G gene into the vector pLa through a unique PmeI site and rescued the virus as described previously [17]. The rescued recombinant rL-RVGTM was confirmed by assessing hemagglutination activity, and the nucleotide sequences of the incorporated RVGTM gene in the rescued virus were confirmed by performing reverse transcription-polymerase chain reaction (RT-PCR) and nucleotide sequencing.
Laser confocal microscopy
BHK-21 cells were grown on coverslips in 35 mm diameter dishes and infected with rL, rL-RVG, or rL-RVGTM at a multiplicity of infection (MOI) of 0.1. At 24 h post-infection, productive infection levels were evaluated by using a 1:100 dilution of dog serum against RV or a 1:3,000 dilution of mouse monoclonal IgG1 against NDV HN antibody (sc53563; Santa Cruz Biotechnology, USA) along with a 1:200 dilution of fluorescein isothiocyanate (FITC)-conjugated rabbit anti-dog antibody (F7884; Sigma, USA) or a tetramethyl rhodamine isocyanate (TRITC)-conjugated goat anti-mouse antibody (ZF-0313; ZSGB-BIO, China), as described previously [26].
Western blotting
BHK-21 cells were incubated with rL, rL-RVG, or rL-RVGTM at an MOI of 5.0, and cell sediments were prepared for analysis at 60 h post-infection. To compare the incorporation of RVG and RVGTM into NDV virions, allantoic fluid was harvested from specific pathogen-free (SPF) chicken eggs at 72 h post-infection, and virus particles were purified as described previously [19]. We determined the amounts of cell-associated protein and purified virion protein by assessing the thickness of β-actin and NDV bands on western blots. Equal amounts of protein were analyzed by western blotting with SDS-12% PAGE. After being incubated with chicken serum against NDV or rabbit serum against RV, horseradish peroxidase-conjugated rabbit anti-chicken IgG (A9046; Sigma) or goat anti-rabbit IgG (ZB-2301; ZSGB-BIO) was used to detect chicken or rabbit serum binding.
Pathogenicity of recombinant virus
To assess the pathogenicity of rL-RVGTM, the mean death time (MDT), intracerebral pathogenicity index (ICPI), and intravenous pathogenicity index (IVPI) of White Leghorn chickens were determined [17]. Values for rL-RVG and rL virus were determined simultaneously for comparison purposes. Two groups of 12 mice (6-week-old mice) were inoculated intramuscularly (i.m.) in the gastrocnemius muscle with 5 × 107 times the 50% egg infective dose (EID50) of rL-RVG or rL-RVGTM. Mouse weight change was determined to assess viral pathogenicity in mammals.
Care of animals and animal experimentation in our laboratory were performed according to approved animal ethics guidelines and protocols. All animal studies were approved by the Animal Ethics Committee of Harbin Veterinary Research Institute of the Chinese Academy of Agricultural Sciences (SYXK (H) 2006-032).
Growth kinetics of rL-RVGTM in cells or eggs
To determine whether RVGTM expression alters the growth properties of rL, 104.5 EID50 of rL, rL-RVG, or rL-RVGTM were inoculated into SPF chicken eggs. Allantoic fluid from five eggs was harvested every 12 h (12–108 h) post-infection, and the TCID50 of each virus was determined by assessing immunofluorescence.
The three viruses at MOIs of 0.1 were inoculated into BHK cells on 24-well plates, and the supernatant harvested at 24, 36, 48, 60, 72, and 84 h post-infection. The viruses were titrated by determining both their immunofluorescence and EID50. Cells were evaluated by examining immunofluorescence in order to investigate the manner of spread of rL-RVGTM in cells after the supernatant was collected. Part of the collected supernatant was used to determine, via immunofluorescence, whether the incorporation of RVGTM alters the trypsin-dependent infectivity of the NDV vector in mammalian cells.
ELISA to detect the incorporation of RVGTM
Purified viruses were assessed by enzyme-linked immunosorbent assay (ELISA) with dog serum against RV or chicken antibody against NDV along with horseradish peroxidase-conjugated rabbit anti-dog IgG (A6792; Sigma) or rabbit anti-chicken IgG (A9046; Sigma). We determined the protein concentrations to standardize the amount of loading (10 µg, 5 µg, 2.5 µg, 1.25 µg, 0.625 µg, 0.3125 µg, 0.15625 µg, and 0.078125 µg) of duplicate samples of the purified virions on the ELISA plate. Data was collected by using an enzyme standard instrument (Model 650; Bio-Rad Laboratories, USA).
Immunization comparison
Two groups of 12 mice were inoculated i.m. with rL-RVG or rL-RVGTM as described in section “Pathogenicity of recombinant virus”. After 4 weeks, the mice received second doses. For serological assessment, blood was obtained from the retro-orbital sinus under isoflurane inhalation anesthesia at 3 and 4 weeks after the first dose and at additional times (1, 2, 3, 5, 7, 9, 13, 17, and 21 weeks) after the second dose. With a World Health Organization standard as a reference, titers of RV VNA were expressed in international units per milliliter of serum (IU/mL) [19].
Results
Generation of recombinant NDVs expressing the RVGTM gene
To determine the effects of replacing the TM and CT regions of RVG with corresponding domains of the NDV F protein, we prepared a synthetic RVGTM and inserted it into the unique PmeI site of pLa to produce the resulting plasmid prL-RVGTM (Fig. 1). The rescued recombinant rL-RVGTM was confirmed by its hemagglutination activity results, and the nucleotide sequences of the incorporated RVGTM gene in the rescued virus were confirmed by the RT-PCR and nucleotide sequencing results (data not shown).
Fig. 1. Construction of a viral vector for expressing the chimeric NDV/RV protein RVGTM. (A) NDV has two transmembrane glycoproteins; HN protein and F protein. The NDV F protein and rabies virus G protein (RVG) are type I transmembrane proteins. (B) Schematic of the generation of recombinant NDV rL bearing the RVGTM gene. RVGTM is a chimera of the RVG ectodomain fused with the sequence for the transmembrane domain and cytoplasmic tail of the NDV protein. The RVGTM sequence was inserted into the vector PBRN-rL at the PmeI site to produce prL-RVGTM. NDV, Newcastle disease virus; RV, rabies virus; RVGTM, chimeric rabies virus G protein.
Verification of rL-RVGTM expression
To verify the expression of RVGTM, we performed laser confocal microscopy. The rL-RVG and rL-RVGTM were stained by both mouse monoclonal IgG1 against NDV HN antibody and dog serum against RV, while the rL was only stained by mouse monoclonal IgG1 against NDV HN (panel A in Fig. 2). Western blot analysis confirmed the expression levels of RVG and RVGTM in BHK-21 cells were similar (panel B in Fig. 2).
Fig. 2. The rL-RVGTM expresses RV and NDV components. (A) Detection of RVGTM expression by immunofluorescence. BHK-21 cells were infected with rL, rL-RVG, or rL-RVGTM at a multiplicity of infection of 0.1. At 24 h post-infection, the cells were fixed and stained with chicken anti-NDV and dog anti-RV sera followed by fluorescein isothiocyanate (FITC)-conjugated rabbit anti-dog and tetramethyl rhodamine isocyanate (TRITC)-labeled goat anti-chicken antibodies. The cells were permeabilized with 0.02% Triton X-100 and cell nuclei were stained with DAPI. (B) Western blot analysis of recombinant NDV expressing RVGTM. Lysates of BHK-21 cells infected with rL, rL-RVG, or rL-RVGTM were separated by 12% SDS-PAGE and incubated with dog serum against RV, chicken serum against NDV, or anti-β-actin monoclonal antibody as a loading control. After incubation with peroxidase-conjugated secondary antibody, the proteins were visualized with 3,3-diaminobenzidine reagent. The locations of marker proteins are indicated on the left, and the antiserum or antibody used is indicated on the right. RVGTM, chimeric rabies virus G protein; RV, rabies virus; NDV, Newcastle disease virus; RVG, rabies virus G protein; MK, protein molecular marker.
Viral growth kinetics
To further assess the ability of RVGTM to produce virus, we measured viral titers after infection of SPF chicken eggs. The rL, rL-RVG, and rL-RVGTM grew to similar levels in the eggs (panel A in Fig. 3). The rL-RVGTM reached a peak titer of 8.375 log TCID50/mL at 72 h post-infection, which was similar to the titer level of rL and slightly higher than that of rL-RVG. These results confirm that rL-RVGTM produces virus at levels equivalent to or greater than those produced by rL and rL-RVG.
Fig. 3. Comparison of viral growth kinetics and viral pathogenicity of rL, rL-RVG, and rL-RVGTM. (A) Ten-day-old embryonated eggs were inoculated with rL, rL-RVG, or rL-RVGTM (0.1 mL of 100 EID50), and the allantoic fluid from each group was harvested at different times (12, 24, 36, 48, 60, 72, 84, 96, or 108 h post-infection). The fluid from five eggs per time was pooled for TCID50 determination in BHK cells. The data shown are means of the results from five experiments, and the error bars indicate SD. (B) Pathogenicity of the recombinant viruses was determined in chicken embryos and chicks by determining the mean death time (MDT) in embryonated eggs, the intracerebral pathogenicity index (ICPI) in 1-day-old chickens, and the intravenous pathogenicity index (IVPI) in 6-week-old chickens. (C) Weight changes of mice inoculated with rL-RVG or rL-RVGTM. Two groups of 12 mice were inoculated intramuscularly (i.m.) in the thigh muscle with 5 × 107 EID50 rL-RVG or rL-RVGTM and were observed and weighed daily for 14 days. All mice survived and body weight changes of each group are shown as ratios of the body weight at day 0 (set as 100). RVG, rabies virus G protein; RVGTM, chimeric rabies virus G protein.
Comparison of the virulence of different recombinant NDVs
Determination of MDT, ICPI, and IVPI (panel B in Fig. 3) levels was performed to obtain measures of virulence. The MDT of rL-RVGTM was more than 120 h while its ICPI and IVPI values were 0, indicating that expression of the RVGTM gene does not alter the pathogenicity of the NDV vector in poultry.
A previous report indicated no differences between the body weight of rL and rL-RVG infected mice. To determine the effects of RVGTM on pathogenicity in mice, we inoculated mice with rL-RVG and rL-RVGTM; all mice survived and body weight changes between two groups were similar (panel C in Fig. 3). Therefore, RVGTM expression does not alter the pathogenicity of the recombinant NDV vector in mice.
RVGTM incorporates into the NDV virion at reduced levels
To determine whether RVGTM incorporates into the NDV virion, we collected viral particles 72 h post-infection of SPF chicken eggs and assessed the contents by western blot analysis. RVG was incorporated more efficiently than RVGTM into the NDV virions, and RVGTM was undetectable by western blotting (panel A in Fig. 4). These results indicate that RVGTM incorporates into the NDV virion at reduced levels.
Fig. 4. RVGTM incorporates poorly into the NDV virion. (A) To determine whether the RVGTM incorporates into the NDV virion, the recombinant rL-RVG and rL-RVGTM viruses were purified through sucrose gradients. Equivalent amounts of rL, rL-RVG, and rL-RVGTM virions were analyzed by SDS-PAGE. (B and C) Top: ELISA analysis of the incorporation of RVGTM and RVG into the NDV virion was performed by testing for G protein. Bottom: ELISA analysis of NDV demonstrates that the amount of the two recombinant viruses is equivalent despite differences in G protein incorporation. RVGTM, chimeric rabies virus G protein; NDV, Newcastle disease virus; RVG, rabies virus G protein; MK, protein molecular marker; NP, nucleuo protein; M, matrixprotein; GP, Glycoprotein.
To verify these results, we performed ELISA testing with anti-RV and anti-NDV antibodies. ELISA performed with dog anti-RV demonstrated that rL-RVG levels were notably higher than rL-RVGTM levels at low dilutions, but the two levels tended to be similar at high dilutions. In contrast, ELISA with chicken anti-NDV yielded similar results at every dilution for both rL-RVG and rL-RVGTM, suggesting that the rL-RVG and rL-RVGTM virion amounts were similar (panel C in Fig. 4). These results confirm that native G protein in rL-RVG incorporates more effectively than the G protein in rL-RVGTM.
Chimeric rL-RVGTM virus has altered ability to spread
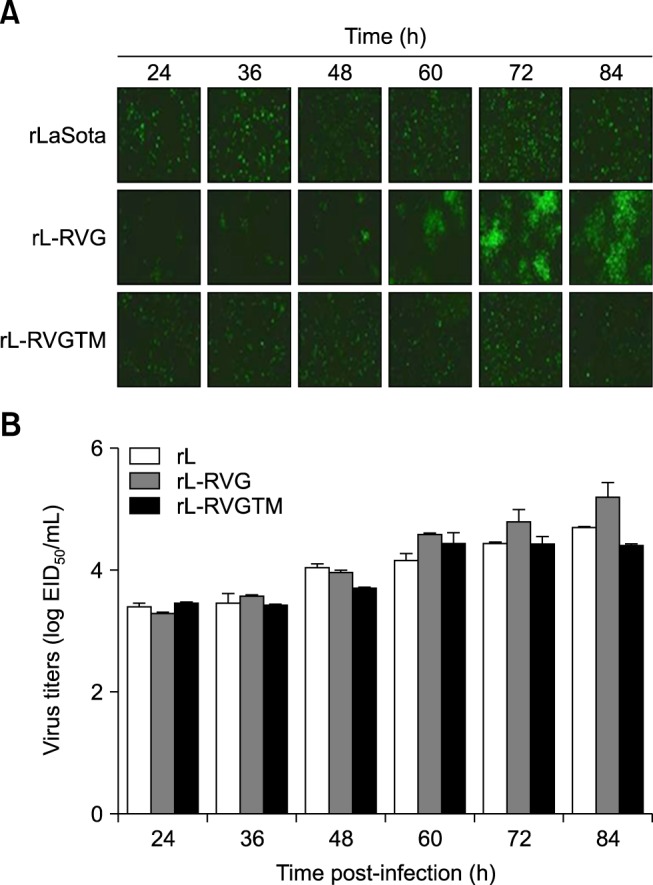
To assess the ability of the rL-RVGTM virus to spread, we performed immunofluorescence assays. The percentages of cells infected by rL and rL-RVGTM were similar and did not change significantly during the 24 to 96 h post-infection time course in the absence of tosylsulfonyl phenylalanyl chloromethyl ketone (TPCK) trypsin. However, with elapsing time, rL-RVG fluorescence spread notably in the absence of TPCK trypsin (panel A in Fig. 5). These results are in contrast to the results obtained in the presence of TPCK trypsin, in which both RVG and RVGTM spread over time, which suggests that the reduced infection level of rL-RVGTM is due to a reduced ability to spread rather than due to effects on the number of viral particles. These findings were verified by analyzing the EID50 of supernatant collected from the three progeny viruses; the three viruses grew to similar levels in eggs, but the titer of rL-RVG was a little higher than those of rL and rL-RVGTM (panel B in Fig. 5).
Fig. 5. rL-RVGTM shows a pattern of spread that is similar to the pattern of rL. (A) Immunofluorescence was used to examine the spread of the viruses. BHK-21 cells were infected with rL, rL-RVG, or rL-RVGTM for 1 h at a multiplicity of infection of 0.1, followed by washing in phosphate belanced solution (PBS) and incubation at 37℃. The infected cells were fixed at the indicated times (24, 36, 48, 60, 72, or 84 h) for immunofluorescent staining with chicken serum against NDV. As shown, rL-RVGTM lost the ability to spread from the initial infected cell to adjacent cells. (B) Supernatants were collected at the same times as in panel A to measure virus titers of 10-day-old embryonated chicken eggs. As shown, the peak titer of rL-RVGTM was similar to that of rL but was approximately one-fifth of a log lower than that of rL-RVG. RVGTM, chimeric rabies virus G protein; RVG, rabies virus G protein.
Induction of anti-NDV and anti-RVG serum antibody responses in mice immunized with recombinant NDVs
To determine whether rL-RVG and rL-RVGTM differ in their immunogenic properties, we immunized mice twice (at weeks 0 and 4) and performed neutralization tests over a 21-week period using serum collected from the mice. The NDV VNA levels of the rL-RVG and rL-RVGTM group both increased sharply and then gradually declined, with similar levels in both viruses after both the first and second immunization and at all times tested (panel A in Fig. 6). In contrast, after the first immunization, the mean titer of RV VNA for rL-RVG was 0.77, while the mean titer of RV VNA for rL-RVGTM was only 0.21 (p = 0.01). After the second immunization, the RV VNA levels were induced to a much greater extent and remained high for a longer period for rL-RVG than for rL-RVGTM (panel B in Fig. 6). Additionally, more than 83.3% of the mice in the rL-RVG group were protected, while only 33.3% of the mice in the rL-RVGTM group were protected after a single dose (data not shown). These results suggest that substitution of the TM and CT regions of RVG greatly diminishes its ability to induce a VNA response.
Fig. 6. rL-RVGTM elicits reduced levels of RV VNA. (A) Induction of serum antibodies specific to NDV and RV were assessed following intramuscularly (i.m.) immunization of mice with equivalent levels of rL-RVG and rL-RVGTM. Twelve mice from each group were given the first immunization at week 0 and a second dose 4 weeks later. Serum samples were collected at the third and fourth weeks after the first immunization and at appropriate times after second dose. Neutralization test of NDV VNA. Values represent the dilution ratio of NDV specific antibodies that can protect half of the cells from being infected by NDV. (B) Neutralization test of RV VNA. Values are presented as IU/mL. *p < 0.05, **p < 0.01. RVGTM, chimeric rabies virus G protein; RV, rabies virus; VNA, virus-neutralizing antibody; NDA, Newcastle disease virus; RVG, rabies virus G protein.
Discussion
Previously, our group generated a recombinant NDV expressing RVG and evaluated its potential as a novel vectored vaccine against rabies in animals [19]. In this study, we replaced the TM domain and CT of RVG with that of the NDV F protein by performing gene synthesis. We generated a recombinant NDV expressing a chimeric RV glycoprotein and tested its characteristics. The expression of RVGTM was verified by undertaking laser confocal microscopy and western blotting, and the results showed that the expression levels of native RVG and RVGTM were similar in BHK-21 cells. Furthermore, MDT, IVPI, and ICPI measurements from mouse experiments indicated that the expression of RVGTM did not increase the virulence of the NDV vector; rL-RVGTM was shown still to comprise a lentogenic strain that is safe for poultry and mice.
Foreign glycoproteins can be incorporated into recombinant NDV virions during their envelopment from the host cell plasma membrane [2,12,13,26]. Western blotting and ELISA tests with equal amounts of purified rL, rL-RVG, and rL-RVGTM viral particles indicated that the incorporation of RVGTM into NDV virions decreased compared to the incorporation of RVG. Mouse experiments showed that the NDV VNA levels of the two groups were similar after the first and second immunization, whereas the level of RV VNA induced by rL-RVGTM was obviously lower than that of rL-RVG. This result is consistent with the result of a study of recombinant NDVs expressing native gD protein and a chimeric gD from bovine herpes virus-1 [22], but is contrary to the result of a study of recombinant NDV expressing native HA protein and a chimeric HA protein of avian H7 influenza [3,29]. Therefore, whether chimeric proteins are incorporated into virions may depend on specific structural criteria of different viral glycoproteins.
RVG is a typical type I glycoprotein that is anchored in the envelope, forming spikes as a trimer, which is the functional unit of its assembly into virions, and mediating virus entry. The basis for the highly efficient incorporation of the RVG into the NDV virion is still not fully described, and the reason for the reduced incorporation when the TM and CT of RVG are replaced is unknown. According to a previous study [22], one possibility is that some amino acid sequence characteristics of the TM domain or CT of the native G protein contribute to their inclusion in particles. TM and CT replacement of G proteins results in slower transportation from the endoplasmic reticulum to the Golgi complex, and the rate at which G protein is transported from the endoplasmic reticulum to the Golgi complex influences the rate of accumulation on the plasma membrane [14,34]. Thus, it is possible that native RVG might accumulate in higher molar amounts at the cell surface leading to efficient incorporation.
Progeny of RV can spread from the initial infected cells to contiguous or noncontiguous cells surrounding the interstitial space to form large plaques [10,19]. However, rL is a low-pathogenicity NDV strain that can infect individual cells but cannot spread to adjacent BHK-21 cells without TPCK trypsin (1 mg/mL). The rL-RVG acquired the ability to spread from cell to cell in BHK-21 cells [19], whereas rL-RVGTM lost this ability, as assessed by immunofluorescence. This change may be explained by several considerations. The vesicular stomatitis virus (VSV) G protein lost its fusion properties when its TM segment was replaced with a glycophosphatidylinositol anchor [28], and a similar phenomenon could explain the lost spreading ability of rL-RVGTM. Mutations in the glycine residues of the TM segment of VSV G protein block fusion at the hemifusion stage [6]. These results suggest that the TM segment of the G protein has a major role in the fusion process of Rhabdoviridae. Furthermore, the 13 membrane-proximal amino acids close to the TM are reported to influence cell-to-cell spreading [21,23]. Therefore, the replacement of the TM and CT may influence the interaction between the 13 membrane-proximal amino acids of RVG and the NDV F protein. Refolding of the C-terminal portion of the molecule also facilitates the switch between the pre- and post-conformations of the G protein [33], a switch that is essential for the fusion process, and the loss of the native C-terminal portion may prevent the structural transition of RVGTM. An additional consideration is that the F protein is a class I fusion protein, while RV G protein is class III fusion protein, and their C-terminal segments have different roles in the pre- and post-fusion states [31]. Thus the TM and CT from the F protein may not be able to compensate for the roles of G protein's TM and CT. The HA protein of avian H7 influenza and the NDV F protein are both class I fusion proteins. Although the trimer structure of NDV F glycoprotein is different from that of HA, in which the orientation of the heptad repeat A is opposite to that of the central, coiled trimer observed in HA [4,5], they undergo a similar conformational change during the fusion process. Amino acid alignment has revealed that all rhabdovirus glycoproteins can be said to share the same fold as VSV; on that basis, the RV G is a class III fusion protein [1]. Although the recognition of G protein domains is similar to that of class II fusion proteins, G protein's refolding process from pre- to post-fusion is similar to that of class I proteins, which form 6-helix bundles by first forming the trimeric central core of the post-fusion conformation. However, in the pre-fusion state, G proteins show a unique property that differs from those of both class I and class II fusion proteins. Their fusion loops are not buried in an oligomeric interface, but instead they point toward the viral membrane [31]. These differences between F, HA, and G proteins may lead to different results. Finally, there are estimated to be at least 15 G protein spikes in the fusion complex that is involved in achievement of the fusion reaction [30,31], and the reduced incorporation of RVGTM may contribute the reduced fusion of RVGTM. We speculate that the chimeric TM and CT results in the loss of the fusion properties of RVGTM.
In summary, we have demonstrated that RVG incorporates more efficiently into recombinant virions than RVGTM does. The replacement of the TM and CT of RVG with that of the NDV F protein may change the conformation of RVGTM and influence the effective antigen epitopes of the G protein. The decreased incorporation of RVGTM into NDV virions and the loss of fusion function of RVGTM further contribute to the reduction of the RV VNA levels induced by rL-RVGTM directly. Our results demonstrate that the C-terminal portion of RVG has an important role in the spreading of the virus from the initially infected cells to surrounding cells.
Acknowledgments
We thank all our laboratory members for their help and Bernard Moss for providing the modified vaccinia virus strain Ankara expressing the T7 RNA polymerase. This research was supported by the Chinese National S&T Plan (2012ZX10004214).
Footnotes
Conflict of Interest: The authors declare no conflicts of interest.
References
- 1.Albertini AA, Baquero E, Ferlin A, Gaudin Y. Molecular and cellular aspects of rhabdovirus entry. Viruses. 2012;4:117–139. doi: 10.3390/v4010117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Bukreyev A, Huang Z, Yang L, Elankumaran S, St Claire M, Murphy BR, Samal SK, Collins PL. Recombinant Newcastle disease virus expressing a foreign viral antigen is attenuated and highly immunogenic in primates. J Virol. 2005;79:13275–13284. doi: 10.1128/JVI.79.21.13275-13284.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Bukreyev A, Skiadopoulos MH, Murphy BR, Collins PL. Nonsegmented negative-strand viruses as vaccine vectors. J Virol. 2006;80:10293–10306. doi: 10.1128/JVI.00919-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Chen L, Colman PM, Cosgrove LJ, Lawrence MC, Lawrence LJ, Tulloch PA, Gorman JJ. Cloning, expression, and crystallization of the fusion protein of Newcastle disease virus. Virology. 2001;290:290–299. doi: 10.1006/viro.2001.1172. [DOI] [PubMed] [Google Scholar]
- 5.Chen L, Gorman JJ, McKimm-Breschkin J, Lawrence LJ, Tulloch PA, Smith BJ, Colman PM, Lawrence MC. The structure of the fusion glycoprotein of Newcastle disease virus suggests a novel paradigm for the molecular mechanism of membrane fusion. Structure. 2001;9:255–266. doi: 10.1016/s0969-2126(01)00581-0. [DOI] [PubMed] [Google Scholar]
- 6.Cleverley DZ, Lenard J. The transmembrane domain in viral fusion: essential role for a conserved glycine residue in vesicular stomatitis virus G protein. Proc Natl Acad Sci U S A. 1998;95:3425–3430. doi: 10.1073/pnas.95.7.3425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Della-Porta AJ, Spencer T. Newcastle disease. Aust Vet J. 1989;66:424–426. doi: 10.1111/j.1751-0813.1989.tb13567.x. [DOI] [PubMed] [Google Scholar]
- 8.Dietzschold B, Schnell M, Koprowski H. Pathogenesis of rabies. Curr Top Microbiol Immunol. 2005;292:45–56. doi: 10.1007/3-540-27485-5_3. [DOI] [PubMed] [Google Scholar]
- 9.Dietzschold B, Wang HH, Rupprecht CE, Celis E, Tollis M, Ertl H, Heber-Katz E, Koprowski H. Induction of protective immunity against rabies by immunization with rabies virus ribonucleoprotein. Proc Natl Acad Sci U S A. 1987;84:9165–9169. doi: 10.1073/pnas.84.24.9165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Dietzschold B, Wiktor TJ, Trojanowski JQ, Macfarlan RI, Wunner WH, Torres-Anjel MJ, Koprowski H. Differences in cell-to-cell spread of pathogenic and apathogenic rabies virus in vivo and in vitro. J Virol. 1985;56:12–18. doi: 10.1128/jvi.56.1.12-18.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Dietzschold B, Wunner WH, Wiktor TJ, Lopes AD, Lafon M, Smith CL, Koprowski H. Characterization of an antigenic determinant of the glycoprotein that correlates with pathogenicity of rabies virus. Proc Natl Acad Sci U S A. 1983;80:70–74. doi: 10.1073/pnas.80.1.70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.DiNapoli JM, Kotelkin A, Yang L, Elankumaran S, Murphy BR, Samal SK, Collins PL, Bukreyev A. Newcastle disease virus, a host range-restricted virus, as a vaccine vector for intranasal immunization against emerging pathogens. Proc Natl Acad Sci U S A. 2007;104:9788–9793. doi: 10.1073/pnas.0703584104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.DiNapoli JM, Ward JM, Cheng L, Yang L, Elankumaran S, Murphy BR, Samal SK, Collins PL, Bukreyev A. Delivery to the lower respiratory tract is required for effective immunization with Newcastle disease virus-vectored vaccines intended for humans. Vaccine. 2009;27:1530–1539. doi: 10.1016/j.vaccine.2009.01.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Doms RW, Keller DS, Helenius A, Balch WE. Role for adenosine triphosphate in regulating the assembly and transport of vesicular stomatitis virus G protein trimers. J Cell Biol. 1987;105:1957–1969. doi: 10.1083/jcb.105.5.1957. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Faber M, Li J, Kean RB, Hooper DC, Alugupalli KR, Dietzschold B. Effective preexposure and postexposure prophylaxis of rabies with a highly attenuated recombinant rabies virus. Proc Natl Acad Sci U S A. 2009;106:11300–11305. doi: 10.1073/pnas.0905640106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Fu ZF. Rabies and rabies research: past, present and future. Vaccine. 1997;15(Suppl):S20–S24. doi: 10.1016/s0264-410x(96)00312-x. [DOI] [PubMed] [Google Scholar]
- 17.Ge J, Deng G, Wen Z, Tian G, Wang Y, Shi J, Wang X, Li Y, Hu S, Jiang Y, Yang C, Yu K, Bu Z, Chen H. Newcastle disease virus-based live attenuated vaccine completely protects chickens and mice from lethal challenge of homologous and heterologous H5N1 avian influenza viruses. J Virol. 2007;81:150–158. doi: 10.1128/JVI.01514-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Ge J, Tian G, Zeng X, Jiang Y, Chen H, Bua Z. Generation and evaluation of a Newcastle disease virus-based H9 avian influenza live vaccine. Avian Dis. 2010;54(1 Suppl):S294–S296. doi: 10.1637/8731-032509-ResNote.1. [DOI] [PubMed] [Google Scholar]
- 19.Ge J, Wang X, Tao L, Wen Z, Feng N, Yang S, Xia X, Yang C, Chen H, Bu Z. Newcastle disease virus-vectored rabies vaccine is safe, highly immunogenic, and provides long-lasting protection in dogs and cats. J Virol. 2011;85:8241–8252. doi: 10.1128/JVI.00519-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Huang Z, Elankumaran S, Yunus AS, Samal SK. A recombinant Newcastle disease virus (NDV) expressing VP2 protein of infectious bursal disease virus (IBDV) protects against NDV and IBDV. J Virol. 2004;78:10054–10063. doi: 10.1128/JVI.78.18.10054-10063.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Jeetendra E, Ghosh K, Odell D, Li J, Ghosh HP, Whitt MA. The membrane-proximal region of vesicular stomatitis virus glycoprotein G ectodomain is critical for fusion and virus infectivity. J Virol. 2003;77:12807–12818. doi: 10.1128/JVI.77.23.12807-12818.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Khattar SK, Collins PL, Samal SK. Immunization of cattle with recombinant Newcastle disease virus expressing bovine herpesvirus-1 (BHV-1) glycoprotein D induces mucosal and serum antibody responses and provides partial protection against BHV-1. Vaccine. 2010;28:3159–3170. doi: 10.1016/j.vaccine.2010.02.051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Maillard AP, Gaudin Y. Rabies virus glycoprotein can fold in two alternative, antigenically distinct conformations depending on membrane-anchor type. J Gen Virol. 2002;83:1465–1476. doi: 10.1099/0022-1317-83-6-1465. [DOI] [PubMed] [Google Scholar]
- 24.Morimoto K, Foley HD, McGettigan JP, Schnell MJ, Dietzschold B. Reinvestigation of the role of the rabies virus glycoprotein in viral pathogenesis using a reverse genetics approach. J Neurovirol. 2000;6:373–381. doi: 10.3109/13550280009018301. [DOI] [PubMed] [Google Scholar]
- 25.Morimoto K, Hooper DC, Spitsin S, Koprowski H, Dietzschold B. Pathogenicity of different rabies virus variants inversely correlates with apoptosis and rabies virus glycoprotein expression in infected primary neuron cultures. J Virol. 1999;73:510–518. doi: 10.1128/jvi.73.1.510-518.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Nakaya T, Cros J, Park MS, Nakaya Y, Zheng H, Sagrera A, Villar E, García-Sastre A, Palese P. Recombinant Newcastle disease virus as a vaccine vector. J Virol. 2001;75:11868–11873. doi: 10.1128/JVI.75.23.11868-11873.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Neumann G, Whitt MA, Kawaoka Y. A decade after the generation of a negative-sense RNA virus from cloned cDNA--what have we learned? J Gen Virol. 2002;83:2635–2662. doi: 10.1099/0022-1317-83-11-2635. [DOI] [PubMed] [Google Scholar]
- 28.Odell D, Wanas E, Yan J, Ghosh HP. Influence of membrane anchoring and cytoplasmic domains on the fusogenic activity of vesicular stomatitis virus glycoprotein G. J Virol. 1997;71:7996–8000. doi: 10.1128/jvi.71.10.7996-8000.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Park MS, Steel J, García-Sastre A, Swayne D, Palese P. Engineered viral vaccine constructs with dual specificity: avian influenza and Newcastle disease. Proc Natl Acad Sci U S A. 2006;103:8203–8208. doi: 10.1073/pnas.0602566103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Roche S, Gaudin Y. Characterization of the equilibrium between the native and fusion-inactive conformation of rabies virus glycoprotein indicates that the fusion complex is made of several trimers. Virology. 2002;297:128–135. doi: 10.1006/viro.2002.1429. [DOI] [PubMed] [Google Scholar]
- 31.Roche S, Rey FA, Gaudin Y, Bressanelli S. Structure of the prefusion form of the vesicular stomatitis virus glycoprotein G. Science. 2007;315:843–848. doi: 10.1126/science.1135710. [DOI] [PubMed] [Google Scholar]
- 32.Veits J, Wiesner D, Fuchs W, Hoffmann B, Granzow H, Starick E, Mundt E, Schirrmeier H, Mebatsion T, Mettenleiter TC, Römer-Oberdörfer A. Newcastle disease virus expressing H5 hemagglutinin gene protects chickens against Newcastle disease and avian influenza. Proc Natl Acad Sci U S A. 2006;103:8197–8202. doi: 10.1073/pnas.0602461103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Weissenhorn W, Hinz A, Gaudin Y. Virus membrane fusion. FEBS Lett. 2007;581:2150–2155. doi: 10.1016/j.febslet.2007.01.093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Whitt MA, Buonocore L, Prehaud C, Rose JK. Membrane fusion activity, oligomerization, and assembly of the rabies virus glycoprotein. Virology. 1991;185:681–688. doi: 10.1016/0042-6822(91)90539-n. [DOI] [PubMed] [Google Scholar]
- 35.Wiktor TJ, Macfarlan RI, Reagan KJ, Dietzschold B, Curtis PJ, Wunner WH, Kieny MP, Lathe R, Lecocq JP, Mackett M. Protection from rabies by a vaccinia virus recombinant containing the rabies virus glycoprotein gene. Proc Natl Acad Sci U S A. 1984;81:7194–7198. doi: 10.1073/pnas.81.22.7194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Wyatt LS, Moss B, Rozenblatt S. Replication-deficient vaccinia virus encoding bacteriophage T7 RNA polymerase for transient gene expression in mammalian cells. Virology. 1995;210:202–205. doi: 10.1006/viro.1995.1332. [DOI] [PubMed] [Google Scholar]