


Abstract
How the 5′-terminus of the template affects RNA synthesis by viral RNA replicases is poorly understood. Using short DNA, RNA and RNA–DNA chimeric templates that can direct synthesis of replicase products, we found that DNA templates tend to direct the synthesis of RNA products that are shorter by 1 nt in comparison to RNA templates. Template-length RNA synthesis was also affected by the concentration of nucleoside triphosphates, the identity of the bases at specific positions close to the 5′-terminus and the C2′-hydroxyl of a ribose at the third nucleotide from the 5′-terminal nucleotide. Similar requirements are observed with two bromoviral replicases, but not with a recombinant RNA-dependent RNA polymerase. These results begin to define the interactions needed for the viral replicase to complete synthesis of viral RNA.
INTRODUCTION
Complete copying of viral genetic information is essential to ensure virus propagation. This may be particularly challenging in RNA viruses with linear genomes where initiation and termination of RNA synthesis must deal with the ends of the template. In (+)-sense RNA viruses of the alphavirus-like superfamily, a viral replicase composed of the RNA-dependent RNA polymerase (RdRp) and other viral and host proteins generally initiates genomic minus-strand synthesis by a primer-independent (de novo) mechanism from the 3′-end of a linear plus-strand RNA. The minus-strand RNA then directs the synthesis of complementary full-length genomic, and sometimes subgenomic, plus-strand RNA (1,2).
De novo initiation of viral RNA synthesis at the 3′-terminus of the template has received increasing attention in the past few years (3). However, what occurs at the 5′-terminus of the template near the termination of RNA synthesis is poorly understood, in a large part because the ends of the template are difficult to manipulate. RNA synthesis at the 5′-terminus of the template is likely a complex process, requiring specific interactions within the transcription elongation complex (TEC) composed of the replicase, the template and nascent RNA. Synthesis of nascent RNA less than template length would generate a prematurely terminated and likely defective viral RNA. Based on precedence with DNA-dependent RNA polymerases (DdRps), premature termination should involve dissociation of the TEC and/or steric changes within the TEC before the polymerase reaches the end of the template (4,5). To elucidate the template determinants that contribute to full-length synthesis, we have studied the replicases of brome mosaic virus (BMV) and cucumber mosaic virus (CMV), members of the bromovirus group of plant viruses in the alphavirus-like superfamily of (+)-sense RNA viruses.
The BMV replicase and several RdRps can use DNA templates for RNA synthesis (6,7). RNA synthesis from DNA templates is usually reduced in comparison to RNA templates, allowing the identification of several features in RNA that contribute to efficient RNA synthesis by RNA replicases (6,7). Also, the ease of manipulating DNA has led to insights into the recognition of RNA templates by viral replicases (8,9).
In this work, we found that DNA templates are less efficient in directing the completion of RNA synthesis. We used a combination of DNA and chimeric DNA–RNA templates to identify the template moieties in a minimal-length template that are required for full-length RNA synthesis. A ribose at the third nucleotide from the 5′-terminus was found to be crucial for template-length RNA synthesis. We further demonstrate that template base composition and a ribose 4 nt from the 5′-end of the template can contribute significantly to complete RNA synthesis. These findings provide initial insight into viral RNA synthesis at the 5′-end of RNA templates.
MATERIALS AND METHODS
DNA, RNA and chimeric templates
Synthetic DNAs were purchased from Operon Technology Inc. (Alameda, CA). Oligoribonucleotides and chimeric templates were synthesized by Dharmacon Inc. (Boulder, CO). The chemically synthesized RNAs and chimeras were purified after denaturing PAGE and eluted from the gel fragments by soaking in 0.3 M sodium acetate overnight. The purified RNA and chimeras were quantified by spectrophotometry, adjusted to the desired concentration and visually inspected after staining with Toluidine blue after denaturing PAGE.
RdRp activity assays
BMV RdRp was prepared from BMV-infected barley as described (10). The CMV replicase was prepared as described in Sivakumaran et al. (10). RdRp activity assays and electrophoresis conditions were as described in Sun et al. (11). All gels were exposed to film at –70°C and the amount of label incorporated into newly synthesized RNAs was determined with a PhosphorImager (Molecular Dynamics).
Replicase product lengths were generally determined by comparison to products generated by the bovine viral diarrhea virus (BVDV) polymerase from chemically synthesized templates. To ensure the correct assignment of replicase product length, an RNA of the same sequence as the product from d(–1/13) (5′-GUAUUAAUAACC-3′) was synthesized chemically and 100 pmol was electrophoresed next to some replicase products. The RNA marker was visualized after Toluidine blue staining and its position marked by excising the stained band with a scalpel to leave a shadow in the autoradiogram.
RESULTS
DNA templates can result in incomplete RNA synthesis
We had previously observed that a number of DNA templates could be used to direct de novo initiation of RNA synthesis by the BMV replicase (7,8). However, products directed by DNA templates have slightly different mobilities in comparison to products from RNA templates (7). To further characterize this difference, we tested several RNA and DNA templates of minimal lengths that contain a 3′-guanylate that is required for RNA synthesis (8), a penultimate initiation cytidylate and 12 additional nucleotides that are derived from the BMV subgenomic template RNA. These templates should allow the BMV replicase to transition from initiation to elongation, a change which takes place after the synthesis of 8–10 nt of nascent RNA (12,13; Fig. 1A). If the BMV replicase completes RNA synthesis on these templates, a 13-nt product is expected. Two independent RNA templates named r(–1/13L) and r(–1/13) produced predominantly 13-nt RNA, with small amounts of 14-nt products, which are due to the addition of one non-templated nucleotide on the newly synthesized RNA (Fig. 1B, lanes 4 and 5, and 9 and 10). In contrast, DNA template d(–1/13), which has the same base sequence as r(–1/13) except for the C5-methyl moiety present in thymines but absent in uracils, directed the synthesis of a mixture of 13- and 12-nt RNA products (Fig. 1B, lanes 6 and 7, and 11 and 12). RNAs produced from d(–1/13) initiated correctly from the penultimate cytidylate (+1C) since a mutation of the +1C to a guanylate abolished RNA synthesis (Fig. 1B, lanes 3 and 8). Incomplete synthesis is also not due to the presence of thymines in d(–1/13) because a chemically synthesized DNA containing deoxyuridines yielded a similar ratio of 13- to 12-nt RNAs (Fig. 1B, compare lanes 11 and 12 to 13 and 14).
Figure 1.

Template length RNA synthesis by the BMV replicase. (A) Sequences of the templates used in this figure. The prototype RNA and DNA are named r(–1/13) and d(–1/13), respectively. In templates d(–1/13dU) and the initiation-incompetent In–, only the nucleotides that differed from r(–1/13) are shown. (B) Autoradiograms of the BMV replicase products directed by RNA and DNA templates. The molecular weight standards are the products generated from template r(–1/13) by the RdRp of BVDV in the presence of all four rNTPs, including [α-32P]CTP (lane 1), and only in the presence of [α-32P]CTP (lane 2). In a complete RNA synthesis reaction the BVDV RdRp generated a 13-nt RNA that terminated at the 5′-end of the template (N) and 14- and 15-nt RNA that contained one or two non-templated nucleotides, N+1 and N+2, respectively. The 15-nt product in lane 2 is the result of terminal nucleotide addition by the BVDV RdRp, which adds a radiolabeled CMP to the 3′-terminus of the 14-nt –1/13 (31). The products identified as N, N–1 and N+1 are shown to the right of the autoradiogram. (C) Schematic of the RNAs produced by the BMV replicase. Within a reaction, the replicase could terminate synthesis 1 nt from the 5′-terminus of the template, producing the N–1 RNA, at the end of the template, generating the N RNA, and adding one nucleotide to the nascent RNA, producing the N+1 RNA.
We used the ratio of full-length RNA (N) to RNA 1 nt shorter than full-length (N–1) as a quantitative measurement to compare termination competency of d(–1/13). This ratio was found to vary slightly depending on factors other than the template, such as the reaction temperature and especially the commercial lot of NTPs (R.Tayon, data not shown). It is likely that some NTPs may contain contaminating amounts of CTP. To eliminate this variability, all comparisons of termination competency made in this work are based on assays run in parallel and quantified on the same gel. Also, due to the position of incorporation of radiolabeled CMP, only the TEC that has reached at least the penultimate guanylate at the 5′-terminus of the template will be observed in autoradiograms.
Several features may increase the synthesis of less than full-length nascent RNA. One may be the limitation of the substrate NTP to be incorporated opposite the 5′-terminal nucleotides. In bromoviruses, template-dependent minus-strand RNA synthesis terminates with two cytidylates while plus-strand synthesis may terminate with one or two guanylates (14). Both r(–1/13) and d(–1/13) contain two guanylates at the 5′-terminus and our RNA synthesis reactions use [α-32P]CTP at a potentially limiting concentration of 0.24 µM, while the other NTPs are present at >200 µM. To determine if increasing CTP concentration in the reaction will increase the ratio of the N relative to the N–1 product using DNA d(–1/13), unlabeled CTP was added to the normal reaction to increase the final CTP concentration to 1.24 µM. This addition decreased the labeling of the replicase product, but increased the amount of the N product relative to N–1 reproducibly by 1.7-fold (Fig. 2A, compare lanes 1–3 to 4–6). In contrast, the ratio of the N+1 to the N products produced by r(–1/13L) was unchanged by the addition of unlabeled CTP (Fig. 2B), as would be expected since incorporation of the N+1 nucleotide is not directed by the template. The ability to incorporate CMP is differentially affected by whether the replicase uses a DNA or an RNA template.
Figure 2.
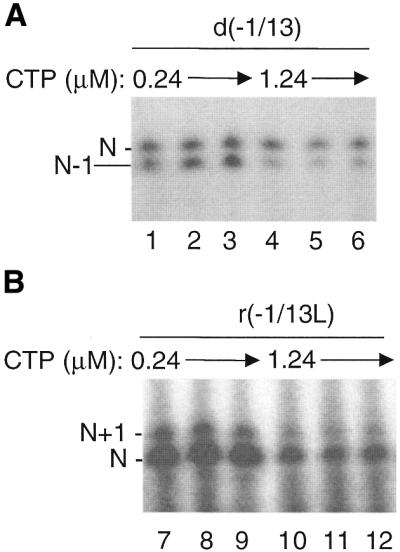
Production of the N–1, but not N+1, RNA is in part due to limiting substrate nucleotides. (A) Autoradiogram demonstrating that the ratio of N to N–1 RNA increases with addition of unlabeled CTP. The template used and the concentration of CTP are indicated above the autoradiogram. N and N–1 RNAs are indicated to the left of the autoradiogram. (B) Autoradiogram demonstrating that the ratio of the N+1 to N RNAs does not change with addition of unlabeled CTP. Template and concentration of CTP and the identity of the RNA products are shown above and to the left of the autoradiogram, respectively.
Template sequence affects the ratio of N to N–1 products
Template sequence is known to affect the TEC by contributing to the stability of the duplex between the template and nascent RNA (15,16). A G-C base pair contributes more stability to the TEC than an A-U or A-T base pair by affecting the template–nascent RNA duplex (15). This hypothesis was tested using d(–1/13) as a prototype. A series of eight templates that contain a substitution to three consecutive cytidylates that scanned along positions –11 to –2 from the 5′-end of the template was made and tested for RNA synthesis by the BMV replicase (Fig. 3). Products from all of the templates will contain three more guanylates compared to those from d(–1/13), which we observed to affect the mobility of the replicase products slightly in comparison to those from d(–1/13). Also, some of the changes near the initiation cytidylate could have resulted in lower overall amounts of RNA synthesis. We have previously reported that sequence near the initiation site will modulate the amount of RNA synthesis (9). In this work, we will primarily be concerned with the ratio of the N to N–1 products generated by each template with limiting CTP concentration. Three consecutive cytidylates within positions –11 to –4 in templates –11/9C to –6/4C all resulted in N to N–1 ratios ranging from 0.51 and 0.78, a level similar to the ratio of products from d(–1/13) (Fig. 3). Templates –11/9C, –10/8C and –9/7C also appear to have initiated synthesis at one of the substituted cytidylates in addition to +1C. Cytidylates at positions –5 to –3 and –4 to –2 in templates –5/3C and –4/2C, respectively, produced significantly more of the 13-nt products. With –4/2C, the ratio of the N to N–1 products increased by more than 6-fold relative to d(–1/13). The bases at positions –4 to –2 from the 5′-terminus of the DNA template appear to significantly affect the amount of full-length RNAs made by the BMV replicase.
Figure 3.

The effects of three consecutive template cytidylates at various positions on the ratio of the N to N–1 RNAs produced by the BMV replicase. The qualitative effects of the cytidylate substitutions are shown in the autoradiogram. The autoradiogram is oriented with the poles of electrophoresis shown at the top and identification of the N–1 and N RNAs at the bottom. The templates tested were named according to the positions of the cytidylate substitutions from the 5′-terminus, and sequences are shown to the right of the names. The relative positions of nucleotides –10, –5 and –1 are noted on top of the sequence. The ratios of the N to N–1 products are derived from at least three independent assays.
To further analyze the role of the nucleotide sequence at positions –4 to –2, 10 templates with different nucleotide combinations at these positions were tested for RNA synthesis by the BMV replicase (Fig. 4). None of the combinations of triplets contained guanylate, thus preventing complications that can arise due to the use of CTP as the radiolabel. Templates that had triplet combinations consisting of all adenylates and/or thymidylates resulted in N to N–1 ratios of less than 0.52 (Fig. 4). When one cytidylate was introduced into this nucleotide triplet, in ACA and CAT, the ratios became 0.7 and 1.6, respectively. The ratio of N to N–1 further increased to above 2.0 when the triplets contained two or three cytidylates.
Figure 4.

Effects of different nucleotide combinations in positions –2 to –4 of DNA templates. The autoradiogram is displayed in the same manner as explained in the legend to Figure 3. Except for template 9TTT, only the nucleotides that are different from template 9TTT are shown. The ratios of the N to N–1 products were calculated from three independent assays.
We wanted to determine whether the differences in the N/N– 1 ratio could be correlated with changes in the template sequence. One likely factor is the interaction between the nascent RNA and the template. Calculations of the thermodynamics of the TEC would require information about the length of the RNA–RNA duplex, which is currently unavailable. The RNA–DNA duplex in elongating complexes of DdRps has been determined to be ∼3 or 8 bp (4,17,18). Assuming a nascent strand–template duplex of 3–8 bp for the viral replicase, the ΔH of the RNA–DNA interaction was calculated using the thermodynamic tables of Turner et al. (19). A poor correlation (r2 < 0.3) exists between ΔH and the ratio of N to N–1 (R.Tayon, data not shown). However, a more straightforward analysis of the number of potential base pairing H-bonds at positions –4 to –2 (three per G-C and two per A-T or A-U) revealed a better correlation with the N to N–1 ratio, with an r2 correlation value of 0.89. We tested further whether a correlation exists between the N to N–1 ratio and the number of H-bonds in positions –2 to –4 by testing inosine bases in two DNA templates that had the sequence 3′-ITT-5′ and 3′-AIT-5′ at positions –4 to –2. Inosine can form a wobbled base pair with adenine, cytosine and uracil. In each of these base pairs, two H-bonds are formed, the same number formed at the comparable position in d(–1/13). The templates containing inosine gave nearly identical N to N–1 ratios as d(–1/13) (C.Kao, data not shown), a result consistent with the hypothesis that the number of H-bonds affects the ratio of the N to N–1 RNAs.
Positions –4 to –2 from the 5′-terminus affect the completion of RNA synthesis
The effects of positions –4 to –2 on termination of RNA synthesis was examined using 13-nt templates. This length raises a concern. The BMV replicase is known to transition from initiation to elongation ∼8 to 10 nt after initiation, which is coincident with positions –4 to –2 (12,13,17). Thus, we wanted to determine whether the effects of positions –4 to –2 are on transition of the replicase to elongation and/or proper termination by using templates of 17 and 21 nt. Each series of templates contains either TTT or CCC at positions –4 to –2 from the respective 5′-termini (Fig. 5). The sequences of TTT and CCC were selected because they generate lowest and highest N to N–1 ratios, respectively, in the context of a 13-nt template (Fig. 4). The 17- and the 21-nt templates with a –4 to –2 sequence of CCC produced several-fold higher N to N–1 ratios in comparison to the templates of the same length with TTT (Fig. 5). As another control, 17-nt templates with TTT at positions –8 to –6 produced identical N to N–1 ratios as compared to a template with CCC at this position (Fig. 5). These results clearly indicate that with three sets of templates of different lengths, positions –4 to –2 from the 5′-terminal nucleotide determine the efficiency of complete RNA synthesis by the BMV replicase, independent of the transition of the replicase to elongation.
Figure 5.

Positions –2 to –4 for templates of different lengths had the same effect on the ratio of N to N–1 products. There are two sequences for each of the three sets of templates tested, one of which contains TTT at position –2 to –4 (black bar) while the other contains CCC at the same position (light bar). The 13-nt templates compared are 9TTT and 9CCC (see Fig. 4). The 17- and 21-nt templates have the sequences 3′-GCATAATTATTAAXXXGG and 3′-GCATATTTACATAATTAXXXGG, where the XXX represents either TTT or CCC. For the 17mer templates, two additional changes that had CCC or TTT at positions –8 to –6 were tested. The key to the bars is shown in the inset. Lines above each bar denote one standard deviation from each mean.
A ribose 2′-hydroxyl contributes to N-length synthesis
Templates r(–1/13) produced primarily N-length replicase products, despite having the same base sequence as d(–1/13dU) (Fig. 1A and B). This suggests that one or more C2′-hydroxyls of the RNA template increased termination competency. To locate the minimal template ribonucleotide(s) required for predominantly N-length synthesis, we tested a series of DNA–RNA chimeras containing the same base sequence as in d(–1/13), but with increasing length of ribonucleotides from the 5′-terminus (Fig. 6). Chimeras with one or three ribonucleotides at the 5′-terminus, c(d12r1) and c(d9r3), resulted in synthesis of predominantly N–1 products, just like d(–1/13) (Fig. 6, lanes 2–3, and R.Tayon, data not shown). However, chimeras with five or seven 5′-terminal ribonucleotides, c(d8r5) and c(d6r7), respectively, yielded predominantly N products (Fig. 6, lanes 4 and 5). These results indicate that one or more riboses within positions –4 and –2 of the template contribute to N-length RNA synthesis by the BMV replicase. This result was tested with chimera c(r–3/4), which has ribonucleotides only at positions –4 and –3. c(r–3/4) generated predominantly N-length RNA (Fig. 6, lane 6). We then tested two chimeras, each of which contains a single ribose at either position –4 or –3, c(r–4) and c(r–3), respectively. Only c(r–3) yielded a predominantly N-length product (Fig. 6, lanes 7 and 8), indicating that a C2′-hydroxyl at position –3 contributes most significantly to N-length RNA synthesis.
Figure 6.

Identification of the ribose(s) that contributes to N-length RNA synthesis by the BMV replicase. The autoradiograms are oriented with the poles of electrophoresis shown at the top and identification of the N–1 and N RNAs at the bottom. DNA and RNA chimeras are identified with a name that starts with a C and the positions of the deoxy- or ribonucleotides relevant to this experiment in parentheses. The sequence of DNA is in capitals, while RNA is in capital italic. In lanes 2–12, only the nucleotides that are different than those in d(–1/13) are shown. Φ denotes a reaction preformed in the absence of template.
To further confirm the importance of the ribose at the –3 position, we designed a chimeric template, c(dN–3), that contains all ribonucleotides except for one deoxyribonucleotide at the –3 position (Fig. 6). The control r(–1/13) produced the predominantly N-length RNA, as expected, while c(dN–3) produced an equal ratio of N and N–1 RNAs (Fig. 6, lanes 9–12). This result confirms that the ribose at the N–3 position in an RNA template contributes to N-length RNA synthesis by the BMV replicase.
RNA synthesis by the CMV replicase
The BMV and CMV replicases have different modes of recognition for the core promoters for the syntheses of subgenomic and genomic minus-strand RNAs (11,20). While promoter recognition is expected to be species specific, it is generally assumed that the ternary complex is likely to be quite similar from species to species. To test this hypothesis, the panel of chimeric RNAs and relevant controls were tested with the CMV replicase. CMV replicase used d(–1/13) 2–3-fold less efficiently relative to r(–1/13) in comparison to the BMV replicase. However, the N and N–1 ratio (0.64 ± 0) was similar to that for the BMV replicase (Fig. 7A). Chimeric template c(d8r5), with five ribonucleotides at the 5′-terminus, and c(r–3/4), with ribonucleotides only at positions –3 and –4, produced essentially all N-length products. Notably, synthesis by the CMV replicase from c(r–4) produced far more N–1 product relative to d(–1/13) than did the BMV replicase (compare Fig. 7, lane 6 and Fig. 6, lane 7). Finally, the key ribonucleotide for N-length RNA synthesis by the CMV replicase was found to reside at N–3 since c(r–3) produced primarily N-length RNAs while c(r–4) produced primarily N–1 RNAs (Fig. 7A). Therefore, despite some differences in the relative amounts of RNA synthesis from the various templates, both the BMV and CMV replicases require a ribose at the N–3 position for N-length RNA synthesis.
Figure 7.

RNA synthesis by the CMV replicase. (A) The templates tested are the same as the one used to elucidate the importance of the N–3 position for the BMV replicase in Figure 6. The autoradiogram is oriented with the poles of electrophoresis shown at the top and identification of the N–1 and N RNAs at the bottom. (B) Effects of the sequence between positions –2 and –4 on the ratio of N to N–1 RNAs produced by the CMV replicase.
We also used the CMV replicase to test the correlation between the ratios of the N to N–1 RNAs and the bases at the –2 to –4 positions. The change in the relative amounts of N to N–1 RNAs was less dramatic in comparison to the BMV replicase when completely DNA templates were used. Also, while the overall level of N-length RNA synthesis increased when positions –4 to –2 had one or more C, we did not find a linear correlation, as we did with the BMV replicase (Fig. 7B). For example, template d9CAT generated an N to N–1 ratio of 1.0 while d9CCC generated a ratio of 0.9 (Fig. 7B). Altogether, these results suggest that the interactions between the replicase and the 5′-terminus of the template can differ between two RNA replicases within the same viral family.
RNA synthesis by the BVDV RdRp
RdRp, the catalytic subunit of the viral replicase, lacks the other viral and cellular proteins found in the replicase complex. However, it is able to initiate RNA synthesis de novo from single-stranded RNA and DNA templates (Fig. 8; 6). We wanted to see whether RNA synthesis by the recombinant BVDV RdRp is affected by the sequence near the 5′-terminus of the template. With all of the DNA and chimeric templates tested, the BVDV RdRp did not produce N–1 RNAs; all of the reactions produced RNAs of length N or longer (Fig. 8 and M.-J.Kim, data not shown). In fact, N+1 and N+2 RNAs dominated the reactions (Fig. 8). With the chimeric templates c(r–3/4), c(r–4) and c(r–3), the products synthesized were not significantly affected by the presence or absence of ribonucleotides within the 5′-terminal 7 nt.
Figure 8.

RNA synthesis by the recombinant BVDV RdRp. The templates used are named at the top of the autoradiogram. Their sequences can be found in Figures 6 and 7. The N-length RNA made by the BMV replicase (lane 2) is used as a molecular weight marker.
DISCUSSION
We seek to better understand the factors that regulate the termination of RNA synthesis by viral RNA replicases. This is an interesting process because improper termination will result in erroneous or incorrect transfer of the viral genetic information. Also, transcription termination from linear RNAs may have mechanistic features distinct from the termination of transcription from DNA templates. For example, the linear RNAs will lack downstream sequence and the non-template strand to stabilize the viral replicase TEC as it approaches the end of transcription, a situation different from the DdRps (21,22). (For reviews of termination of transcription by prokaryotic RNA polymerases and RNA PolII see 23–25.)
The availability of CTP when the replicase reaches the end of a DNA template containing two guanylates affected full-length RNA synthesis. Presumably the availability of other NTPs to be incorporated by the replicase near the 5′-terminus of the template could also affect full-length RNA synthesis by affecting dissociation of the TEC. Also, the bases at positions –4 to –2 relative to the 5′-terminus can influence the amount of N to N–1 products. The effects of the bases at positions –4 to –2 were observed with templates of different lengths, indicating that their effect is indeed on termination by the TEC, not on initiation or the transition from initiation. We also found that the 2′-hydroxyl located at position –3 relative to the terminal nucleotide plays a significant role in determining whether RNA synthesis is complete. Furthermore, these requirements are observed with both the BMV and CMV replicases, but not with the recombinant BVDV RdRp.
The observation that higher CTP concentration could increase N-length RNA synthesis suggests that the TEC may either dissociate or continue RNA synthesis in a manner dependent on substrate availability. We previously showed that template switch (transcribing a nascent RNA from a second non-covalently linked template) could be induced by limiting NTP concentrations (26). Dissociation of the TEC may be more rapid as the TEC approaches the 5′-terminus due to one or more factors, including loss of the downstream contacts, more locally limiting amounts of NTPs or an increased amount of potentially inhibitory pyrophosphates. However, several other BMV templates, including RNAs of >200 nt, were apparently able to complete RNA synthesis despite the need to incorporate a larger number of CMPs in comparison to d(–1/13) (see for example 17). This observation suggests that the effects of limiting CTP concentration are more severe when the replicase is at or near the 5′-terminus of the template. We also note that the effect of limiting CTP concentration does not affect non-templated nucleotide addition (Fig. 2B).
In addition to the interactions at the catalytic site, whether the replicase ternary complex continues synthesis may be affected by the stability of the template–nascent RNA duplex (27). The BVDV RdRp was able to synthesize efficiently through the base analog 4-thiouridine (which does not allow normal Watson–Crick base pairing) only when the RdRp had already synthesized a number of nucleotides (6). We have found that potentially more stable base pairing between –4 and –2 of a DNA template and the nascent RNA contributes to the ability of the BMV and, to a lesser degree, the CMV replicase, to produce N-length products (Figs 4 and 7). The effects of changes in the bases were more difficult to observe in RNA templates because the riboses of RNAs helped to stabilize the TEC. However, we note that positions –4 and –2 on an RNA template were also found to dramatically affect the ability of the BVDV RdRp to switch templates in vitro (26), suggesting that N-length RNA synthesis and the ability to continue synthesis on a second template may both depend on the stability of the TEC at the 5′-terminus of the template. The influence of the nascent RNA on the stability of the TEC has been documented with DdRps (5,14,27). For the Escherichia coli RNA polymerase, an ∼8 bp template DNA–nascent RNA is needed to stabilize the TEC (5,28) and manipulating the sequence of the template–nascent RNA duplex will alter the position of the TEC along the template (14). In fact, destabilizing the duplex may cause dislocation of the 3′-terminus of the nascent RNA from the catalytic site of the polymerase (4,14).
The ribose C2′-hydroxyl at position –3 relative to the 5′-terminal nucleotide contributes to N-length RNA synthesis (Fig. 6). A highly specific requirement for the C2′-hydroxyl at position –3 is found for both the BMV and CMV replicases; neighboring nucleotides are unable to compensate for the absence of the N–3 ribose. This specificity can be contrasted with the requirement for initiation, where the position of the initiation site can vary by one or a few nucleotides and retain significant amounts of RNA synthesis (8,29,30). The termination of RNA synthesis may have more rigid spatial requirements.
Of all of the factors we have identified, the N–3 ribose may contribute most significantly to N-length RNA synthesis. The ribose C2′-hydroxyl at the N–3 position could be necessary either to ensure the proper conformation of the ribose (in the C3′-endo conformation) or to form an H-bond with an R group of a replicase subunit. At present, we cannot distinguish between these two possibilities. It will be interesting to identify the amino acid residue(s) in the BMV replicase that interacts with the N–3 position. Since the CMV and BMV replicases appear to have very similar requirements for the N–3 ribose, this interacting subunit in the replicase may be arranged in the same spatial position.
Lastly, RNA synthesis by the BVDV RdRp does not appear to conform to the same rules as the BMV and CMV replicases, even with identical buffer components and substrate concentrations in the reaction. The BVDV RdRp appears to stay on the template better than do the BMV and CMV replicases, generating products that are longer than length N even though CTP concentration in the reaction is limiting for the replicases. Also, the lack of a ribose at the N–3 position does not appear to affect synthesis by the BVDV RdRp. In better characterized DdRps, a number of factors, including cellular proteins, will interact with the TEC to modulate polymerase pausing and termination (for reviews see 23,24). It is tempting to speculate that the multiple subunit replicase contains sites of interaction lacking in RdRps that allow a differential response to different exogenous signals. However, we cannot rule out inherent differences in the activities of RdRps from different viral families.
Acknowledgments
ACKNOWLEDGEMENTS
R.T., M.-J.K. and C.K. gratefully acknowledge fellowships from the Ronald E. McNair Scholars Program, the Samuel Noble Foundation and the Linda and Jack Gill Fellowship, respectively. This research was funded by grants from the NSF (MCB9507344) and USDA (9702126).
References
- 1.Buck K.W. (1996) Comparison of the replication of positive-strand RNA viruses of plants and animals. Adv. Virus Res., 47, 159–251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Miller W.A. and Koev,G. (2000) Synthesis of subgenomic RNAs by positive-strand RNA viruses. Virology, 273, 1–8. [DOI] [PubMed] [Google Scholar]
- 3.Kao C., Sighn,P. and Ecker,D. (2001) De novo initiation of viral RNA-dependent RNA synthesis. Virology, in press. [DOI] [PubMed] [Google Scholar]
- 4.Nudler E., Mustaev,A., Lukhtanov,A. and Goldfarb,A. (1997) The RNA-DNA hybrid maintains the register of transcription by preventing backtracking of RNA polymerase. Cell, 89, 33–41. [DOI] [PubMed] [Google Scholar]
- 5.Wilson K.S., Conant,C.R. and von Hippel,P.H. (1999) Determinants of the stability of transcription elongation complexes: interactions of the nascent RNA with the DNA template and the RNA polymerase. J. Mol. Biol., 289, 1179–1194. [DOI] [PubMed] [Google Scholar]
- 6.Kim M.-J., Zhong,W., Hong,Z. and Kao,C. (2000) Template nucleotide moieties required for de novo initiation of RNA synthesis by a recombinant viral RNA-dependent RNA polymerase. J. Virol., 74, 10312–10322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Siegel R.W., Bellon,L., Beigelman,L. and Kao,C.C. (1999) Use of DNA, RNA and chimeric templates by a viral RNA-dependent RNA polymerase: evolutionary implications for the transition from the RNA to the DNA world. J. Virol., 73, 6424–6429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Sivakumaran K. and Kao,C. (1999) Initiation of genomic (+)-strand RNA synthesis from DNA and RNA templates by a viral RNA-dependent RNA polymerase. J. Virol., 73, 6415–6423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Sivakumaran K., Kim,C.-H., Tayon,R.,Jr and Kao,C. (1999) RNA sequence and secondary structural determinants in a minimal viral promoter that directs replicase recognition and initiation of genomic plus-strand RNA synthesis. J. Mol. Biol., 294, 667–682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Sivakumaran K., Bao,Y., Roossinck,M. and Kao,C. (2000) Recognition of the core RNA promoter for minus-strand RNA synthesis by the replicases of Brome mosaic virus and Cucumber mosaic virus. J. Virol. 74, 10323–10331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Sun J.-H., Adkins,S., Faurote,G. and Kao,C. (1996) Initiation of (–)-strand RNA synthesis catalyzed by the BMV RNA-dependent RNA polymerase: synthesis of oligonucleotides. Virology, 226, 1–12. [DOI] [PubMed] [Google Scholar]
- 12.Sun J. and Kao,C. (1997) RNA synthesis by the brome mosaic virus RNA-dependent RNA polymerase: transition from initiation to elongation. Virology, 233, 63–73. [DOI] [PubMed] [Google Scholar]
- 13.Sun J. and Kao,C. (1997) Characterization of RNA products associated with or aborted by a viral RNA-dependent RNA polymerase. Virology, 236, 348–353. [DOI] [PubMed] [Google Scholar]
- 14.Chapman M., Tayon,R. and Kao,C. (1999) Initiation of minus-strand RNA synthesis by the brome viral RNA replicases. Curr. Top. Virol., 1, 175–189. [Google Scholar]
- 15.Toulme F., Guerin,M., Robichon,N., Leng,M. and Rahmouni,A.R. (1999) In vivo evidence for back and forth oscillation of the transcription elongation complex. EMBO J., 18, 5052–5060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Korzheva N., Mustaev,A., Kozlov,M., Malhotra,A., Nikiforov,V., Goldfarb,A. and Darst,S.E. (2000) A structural model of transcription elongation. Science, 289, 619–625. [DOI] [PubMed] [Google Scholar]
- 17.Adkins S., Stawicki,S.S., Faurote,G., Siegel,R.W. and Kao,C. (1998) Mechanistic analysis of RNA synthesis by RNA-dependent RNA polymerase from two promoters reveals similarities to DNA-dependent RNA polymerase. RNA, 4, 455–470. [PMC free article] [PubMed] [Google Scholar]
- 18.Chamberlin M.J. (1995) New model for the mechanism of transcription elongation and its regulation. Harvey Lect., 88, 1–21. [PubMed] [Google Scholar]
- 19.Turner D.H., Sugimoto,N. and Freier,S.M. (1988) RNA structure prediction. Annu. Rev. Biophys. Chem., 17, 167–192. [DOI] [PubMed] [Google Scholar]
- 20.Chen M.-H., Roossinck,M.J. and Kao,C. (2000) Efficient and specific initiation of subgenomic RNA synthesis by cucumber mosaic virus replicase in vitro requires an upstream RNA stem-loop. J. Virol., 74, 11201–11209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Guerin M., Leng,M. and Rahmouni,A.R. (1996) High resolution mapping of the E. coli transcription elongation complex in situ reveals protein interactions with the nontranscribed strand. EMBO J., 15, 5397–5407. [PMC free article] [PubMed] [Google Scholar]
- 22.Wang D. and Landick,R. (1997) Nuclease cleavage of the upstream half of the nontemplate strand DNA in an Escherichia coli transcription elongation complex causes upstream translocation and transcriptional arrest. J. Biol. Chem., 272, 5989–5994. [DOI] [PubMed] [Google Scholar]
- 23.Conaway J.W. and Conaway,R.C. (1999) Transcription elongation and human disease. Annu. Rev. Biochem., 68, 301–319. [DOI] [PubMed] [Google Scholar]
- 24.Henkin T.M. (2000) Transcription termination control in bacteria. Curr. Opin. Microbiol., 3, 149–153. [DOI] [PubMed] [Google Scholar]
- 25.Zhao J., Hyman,L. and Moore,C. (1999) Formation of mRNA 3′ ends in eukaryotes: mechanism, regulation and interrelationships with other steps in mRNA synthesis. Microbiol. Mol. Biol. Rev., 63, 405–445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Kim M.-J. and Kao,C. (2001) RNA recombination in vitro by a viral RNA-dependent RNA polymerase and RNA replicases. Proc. Natl Acad. Sci. USA, 98, 4972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.von Hippel P.H. and Yager,T.D. (1991) Transcript elongation and termination are competitive kinetic processes. Proc. Natl Acad. Sci. USA, 88, 2307–2311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Deng L. and Shuman,S. (1997) Transcription termination by vaccinia RNA polymerase entails recognition of specific phosphates in the nascent RNA. J. Biol. Chem., 272, 695–698. [DOI] [PubMed] [Google Scholar]
- 29.Chapman M., Rao,A.L.N. and Kao,C. (1998) The 5′ end of BMV RNAs promotes initiation of (–)-strand RNA synthesis in vitro and in vivo. Virology, 252, 458–467. [DOI] [PubMed] [Google Scholar]
- 30.Stawicki S. and Kao,C. (1999) Spatial requirements for promoter recognition by a viral RNA-dependent RNA polymerase. J. Virol., 73, 198–204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Ranjith-Kumar R.T., Gajewski,J., Gutshall,L., Maley,D., Sarisky,R. and Kao,C. (2001) Terminal nucleotidyl transferase activity of recombinant RNA-dependent RNA polymerases. Implications for the initiation of viral RNA synthesis. J. Virol., in press. [DOI] [PMC free article] [PubMed] [Google Scholar]