


Abstract
In previous studies we showed that galectin-1 and galectin-3 are factors required for the splicing of pre-mRNA, as assayed in a cell-free system. Using a yeast two-hybrid screen with galectin-1 as bait, Gemin4 was identified as a putative interacting protein. Gemin4 is one component of a macromolecular complex containing approximately 15 polypeptides, including SMN (survival of motor neuron) protein. Rabbit anti-galectin-1 co-immunoprecipitated from HeLa cell nuclear extracts, along with galectin-1, polypeptides identified to be in this complex: SMN, Gemin2 and the Sm polypeptides of snRNPs. Direct interaction between Gemin4 and galectin-1 was demonstrated in glutathione S-transferase (GST) pull-down assays. We also found that galectin-3 interacted with Gemin4 and that it constituted one component of the complex co-immunoprecipitated with galectin-1. Indeed, fragments of either Gemin4 or galectin-3 exhibited a dominant negative effect when added to a cell-free splicing assay. For example, a dose-dependent inhibition of splicing was observed in the presence of exogenously added N-terminal domain of galectin-3 polypeptide. In contrast, parallel addition of either the intact galectin-3 polypeptide or the C-terminal domain failed to yield the same effect. Using native gel electrophoresis to detect complexes formed by the splicing extract, we found that with addition of the N-terminal domain the predominant portion of the radiolabeled pre-mRNA was arrested at a position corresponding to the H-complex. Inasmuch as SMN-containing complexes have been implicated in the delivery of snRNPs to the H-complex, these results provide strong evidence that galectin-1 and galectin-3, by interacting with Gemin4, play a role in spliceosome assembly in vivo.
INTRODUCTION
Pre-mRNA splicing is an essential post-transcriptional process coordinated by a macromolecular complex termed the spliceosome. This assembly is composed of five small nuclear ribonucleoprotein particles (U1, U2, U4, U5 and U6 snRNPs) and non-snRNP proteins, estimated to number between 50 and 100 (1–3). Many of these splicing factors are involved in one or more of the following steps: snRNP biogenesis, spliceosome assembly and disassembly and the catalytic events responsible for intron excision and exon ligation (4–6).
Galectins-1 and -3 belong to a family of proteins defined by their binding to galactose-containing glycoconjugates (for a recent review see 7). The nuclear localization of galectin-3, coupled with its co-sedimentation with nuclear RNPs and snRNPs, provided the initial hint that it might play a role in pre-mRNA splicing (8,9). Using a cell-free splicing assay, subsequent experiments documented: (i) saccharides that bind galectins with high affinity inhibited the splicing reaction; (ii) depletion of both galectins from the nuclear extract, either by lactose affinity chromatography or by antibody adsorption, resulted in concomitant loss of splicing activity; and (iii) either recombinant galectin-1 or recombinant galectin-3 was able to reconstitute splicing activity in a galectin-depleted extract (10,11). Finally, double immunofluorescence experiments showed that both galectin-1 and -3 can be co-localized in nuclear speckles with known splicing factors (the core polypeptides of snRNPs bearing the Sm epitope and the non-snRNP splicing factor SC35) (11,12).
In order to define the role played by galectins in the splicing process, it was important to identify and characterize their interacting partner(s) in the nucleus. To this end, we carried out a yeast two-hybrid screen using galectin-1 as bait. In the present communication we identify Gemin4, one component of nuclear complexes containing survival of motor neuron (SMN) protein, as a partner that directly binds to galectin-1 and galectin-3. We also show that fragments of either of the Gemin4–galectin-3 partners, such as the C-terminal 50 amino acids of Gemin4 [hereafter designated Gemin4(C50)] or the ∼140-residue N-terminal domain of galectin-3 (designated ND), exert a dominant negative effect on pre-mRNA splicing. The data suggest that the ND blocks an early step in spliceosome formation, consistent with the notion that SMN-containing complexes are responsible for the delivery of snRNPs to the spliceosome.
MATERIALS AND METHODS
Yeast two-hybrid screen
The cDNA of human galectin-1 (13) was kindly provided by Drs K.Kasai and J.Hirabayashi (Teikyo University, Sagamiko, Kanagawa, Japan). It was subcloned into the BamHI site of the vector pAS2-1 (Clontech) downstream of the yeast GAL4 DNA-binding domain. A HeLa cell cDNA library fused to pGAD-GH vector was purchased from Clontech and served as the prey. The screen for putative interacting partners was performed following the manufacturer’s instructions. The His+/LacZ+ clones were isolated and their DNAs were amplified by PCR. Sequence analysis was carried out by the Macromolecular Structure, Sequencing and Synthesis Facility at Michigan State University.
Protein preparations and in vitro binding assays
Procedures for the preparation of recombinant murine galectin-3 and its ND and CD (C-terminal domain, 147 amino acids) have been described (14). The purity of these protein preparations was characterized by SDS–PAGE and immunoblotting using domain-specific antibodies. In addition, the molecular weights of recombinant galectin-3, ND and CD were determined by mass spectrometry, using matrix-assisted laser desorption/ionization (Voyager-DE STR; PE PerSeptive Biosystems). The same samples were also subjected to amino acid sequence analysis, using a Procise pulsed liquid protein sequence (PE Applied Biosystems). Both of these procedures, carried out by the Macromolecular Structure, Sequencing and Synthesis Facility at Michigan State University, confirmed the identity and purity of the protein preparations. Bovine galectin-1 (15) used for the in vitro binding assay was kindly provided by Dr Ten Feizi (Imperial College School of Medicine, Northwick Park, Harrow, UK).
The cDNA of Gemin4(C50), identified through the yeast two-hybrid screen, was subcloned into the NcoI and EcoRI restriction sites of pGEX-2T (Pharmacia) to produce the fusion protein GST–Gemin4(C50). Glutathione–Sepharose beads (Pharmacia) were pretreated with bacterial cell lysate isolated from Escherichia coli used for recombinant fusion protein production. Approximately 1 µg of GST or GST–Gemin4(C50) was adsorbed to 20 µl of the pretreated beads. Then, 0.5 µg of bovine galectin-1 or recombinant murine galectin-3 was incubated with the GST or GST–Gemin4(C50) beads at 4°C for 2 h in 60% buffer D (20 mM HEPES, pH 7.9, 20% glycerol, 100 mM KCl and 0.2 mM EDTA) containing 0.05% Triton X-100. After extensive washing, the bound material was eluted with SDS–PAGE sample buffer and resolved by electrophoresis.
Immunoprecipitation of complexes from nuclear extracts of HeLa cells
HeLa S3 cells were grown in suspension culture by the National Cell Culture Center (Minneapolis, MN). Nuclear extract (NE) was prepared in buffer C (20 mM HEPES, pH 7.9, 25% glycerol, 0.42 M NaCl, 1.5 mM MgCl2, 0.2 mM EDTA, 0.5 mM phenylmethylsulfonyl fluoride, 0.5 mM dithiothreitol) or buffer D, as described by Dignam et al. (16). NEs were frozen as aliquots in a dry ice/ethanol bath and stored at –80°C. Protein concentrations were determined by the Bradford assay (17). In this study the protein concentration of NE was ∼4 mg/ml.
Polyclonal rabbit antiserum against recombinant rat galectin-1 (18) was a gift from Dr Sam Barondes (University of California, San Francisco, CA). We have previously described the adsorption and covalent coupling of this antibody to protein G–Sepharose beads (11). NE (∼80 µg protein) was incubated with beads containing anti-galectin-1 or the corresponding preimmune serum. The incubation was carried out at 4°C for 2 h in 60% buffer D containing 0.05% Triton X-100. After extensive washing with the same buffer, the bound material was eluted with SDS–PAGE sample buffer and resolved by electrophoresis.
SDS gel electrophoresis and immunoblotting
Proteins were resolved on 12 or 15% acrylamide gels electrophoresed in the presence of SDS (SDS–PAGE), as described by Laemmli (19). The polypeptides were revealed by staining with Coomassie blue, using the Gel Code Blue reagent of Pierce. Polypeptides were also revealed by immunoblotting of Hybond-C membranes (Amersham), following the general procedures previously described (10).
A rat monoclonal antibody was developed against the Mac-2 antigen (20), which has been shown to be galectin-3 (21) (αGal-3). The anti-galectin-1 antibodies used for immunoblotting were affinity purified from the serum of rabbits immunized with GST–human galectin-1 (αGal-1). Mouse monoclonal antibodies directed against SMN (αSMN) and Gemin2 (αGemin2) were purchased from Transduction Laboratories. The B, B′ and D polypeptides of the snRNPs were detected with human autoimmune serum (ENA anti-Sm) purchased from The Binding Site. HMG 14/17 polypeptides were detected with a rabbit polyclonal antiserum (a gift from Michael Bustin, NIH, Bethesda, MD). The polypeptides were revealed using horseradish peroxidase-conjugated secondary antibodies and the enhanced chemiluminescence system (Amersham Pharmacia).
In vitro splicing assay
The plasmid used to transcribe the MINX pre-mRNA substrate was a kind gift of Dr Susan Berget (Baylor College of Medicine, Houston, TX). The MINX pre-mRNA was labeled with [32P]GTP and the monomethyl cap was added during SP6 polymerase (Gibco BRL) transcription (22).
Samples of NE were dialyzed against 60% Dignam buffer D in the absence or presence of exogenously added proteins: GST, GST–Gemin4(C50), recombinant murine galectin-3, ND or CD. The dialysis was carried out for 75 min at 4°C in a microdialyzer with a 6–8 kDa cut-off dialysis membrane (11).
Splicing reaction mixtures, in a total volume of 10 µl, contained dialyzed NE (6–8 µl), [32P]MINX pre-mRNA, 2.5 mM MgCl2, 1.5 mM ATP, 20 mM creatine phosphate, 0.5 mM dithiothreitol and 20 U RNasin (Promega). Splicing reactions were incubated at 30°C for 45–60 min. The assay was stopped by addition of proteinase K and SDS to final concentrations of 4 mg/ml and 0.1%, respectively. The samples were incubated at 37°C for 20 min, diluted to 100 µl with 125 mM Tris, pH 8, 1 mM EDTA, 0.3 M sodium acetate. RNA was extracted with 200 µl of phenol/chloroform (50:50 v/v), followed by 200 µl of chloroform. The RNAs were precipitated with 300 µl of ethanol at –80°C. The extracted RNAs were subjected to electrophoresis through 13% polyacrylamide (bisacrylamide:acrylamide 1.9:50 w/w)–8.3 M urea gels. The radioactive RNA bands were revealed by autoradiography.
Gel mobility shift assay for splicing complex formation
Ribonucleoprotein complexes representing intermediates in the splicing pathway were monitored by electrophoresis in non-denaturing gels (22,23). Splicing reaction mixtures (10 µl) were incubated at 30°C for 0–20 min. Heparin was then added to a final concentration of 1 mg/ml, along with 1 µl of glycerol containing 0.2% each of bromophenol blue, xylene cyanol and phenol red. The samples were loaded onto a pre-run 4% polyacrylamide gel (bisacrylamide:acrylamide 1:80 w/w). Electrophoresis was carried out in 50 mM Tris, 50 mM glycine, pH 8.8, at 4°C for ∼90 min at constant voltage (150 V). The positions of migration of the splicing complexes were revealed by autoradiography of the radioactive pre-mRNA. Quantitation of the amount of radioactivity was carried out on a STORM phosphorimager (Molecular Dynamics, Sunnyvale, CA).
RESULTS
Identification of Gemin4 as an interacting partner of galectin-1 and galectin-3
To identify proteins that interact with the galectins, we performed a yeast two-hybrid screen (24) using human galectin-1 as the bait protein and a HeLa cell cDNA library as prey. We screened 2 × 106 independent clones and isolated 12 His+/LacZ+ clones. Based on sequence analysis, four clones were identified as ribosomal proteins (RPL37, RPS26, RPS3 and 60S acidic ribosomal protein PO), four clones were identified as cytochrome oxidase subunit III and two clones were identified as ubiquitin-like proteins, human SMT3B/SUMO-3 (small ubiquitin-like modifier). Two clones contained novel sequences at the time of initial isolation. One of the latter, however, was identified as the C-terminal 50 amino acids of Gemin4 (accession no. AF173856) when the sequence was subjected to a BLAST search against the GenBank sequence database. There was 100% identity between the two sequences (Fig. 1). The sequence identified through the yeast two-hybrid screen is designated Gemin4(C50).
Figure 1.

Comparison of the nucleotide sequence of Gemin4 and the corresponding sequence of the clone identified as a ligand of galectin-1. DNA from the putative interacting clone was amplified by PCR and then sequenced. When the sequence of this clone (Query) was subjected to a BLAST search against the GenBank genome database, there was 100% identity with the C-terminal 50 amino acids of Gemin4 (accession no. AF173856).
The cDNA of Gemin4(C50) was subcloned into pGEX-2T and the fusion protein GST–Gemin4(C50) was purified from the bacterial expression system. Direct binding between galectin-1 and Gemin4(C50) was tested in GST pull-down assays. Galectin-1 bound to GST–Gemin4(C50) (Fig. 2A, lane 3) but not to GST alone (Fig. 2A, lane 2). We also tested the binding of galectin-3 to Gemin4(C50) using the same in vitro binding assay. Galectin-3 bound directly to GST–Gemin4(C50) (Fig. 2B, lane 6) but not to GST (Fig. 2B, lane 5).
Figure 2.

In vitro binding assay between GST–Gemin4(C50) and galectin-1 and galectin-3. Approximately 1 µg of GST (lanes 2, 5 and 7) or GST–Gemin4(C50) (lanes 3, 6 and 8) was bound to glutathione–Sepharose beads that had been pretreated with bacterial lysate from E.coli cells used for production of the recombinant fusion polypeptides. Bovine galectin-1 (0.5 µg) (A) or recombinant murine galectin-3 (0.5 µg) (B) was incubated with the GST or GST–Gemin4(C50) beads for 2 h at 4°C. After washing, the bound material was eluted and subjected to SDS–PAGE. Galectin-1 and galectin-3 were visualized by immunoblotting. In lanes 1 and 4, 10% of the input test protein is shown. After immunoblotting, one of the membranes was stained with Coomassie blue (C) to ascertain that equal quantities of GST and GST–Gemin4(C50) were bound to the glutathione beads.
In these experiments galectin-1 and galectin-3 were visualized by immunoblotting. The membranes used for immunoblotting were also stained with Coomassie blue to reveal GST (Mr ∼27 000) and GST–Gemin4(C50) (Mr ∼32 000). This was done as a control to ascertain that approximately the same amounts of GST and GST–Gemin4(C50) were adsorbed onto the glutathione beads (Fig. 2C, lanes 7 and 8). Finally, we also tested and found no binding of bovine serum albumin to the GST and GST–Gemin4(C50) beads (data not shown). Thus, it can be inferred that galectin-1, as well as galectin-3, interacts specifically with Gemin4.
Co-immunoprecipitation of galectin-1 and galectin-3 in complexes containing SMN
Gemin4 is one of approximately 15 polypeptides of a macromolecular complex containing SMN, Gemin2, Gemin3 and the core polypeptides of snRNPs bearing the Sm epitope (Sm B/B′, Sm D1–D3 and Sm E–G) (25,26). Thus, our demonstration of a direct interaction between galectin-1 and Gemin4 prompted us to test whether galectin-1 is also a component of such nuclear complexes containing SMN. Polyclonal rabbit anti-galectin-1 was used to immunoprecipitate complexes from NE derived from HeLa cells. Proteins immunoprecipitated with anti-galectin-1 were separated by SDS–PAGE and subjected to immunoblotting with various antibodies (Fig. 3). First, anti-galectin-1 immunoprecipitated galectin-1 from NE (Fig. 3, top). In addition, the following polypeptides, for which we had antibody reagents, were also found in the same anti-galectin-1 immunoprecipitate: (i) SMN (∼38 kDa); (ii) Gemin2 (∼36 kDa); (iii) galectin-3 (∼30 kDa); and (iv) the Sm B, B′ and D polypeptides of the snRNPs.
Figure 3.
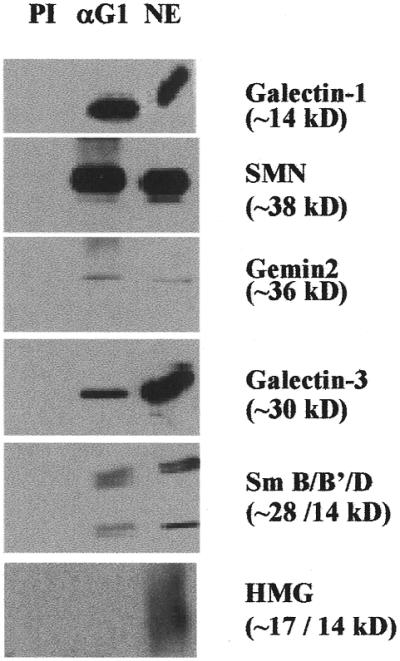
Analysis of the polypeptides immunoprecipitated from NE by anti-galectin-1. NE (∼80 µg protein) was incubated for 2 h at 4°C with protein G–Sepharose beads covalently coupled with polyclonal rabbit anti-rat galectin-1. After washing, the bound material was eluted and subjected to SDS–PAGE. Polypeptides (indicated to the right of each panel) in the immunoprecipitates were identified with the following antibodies: affinity purified rabbit polyclonal antibodies against human galectin-1; mouse monoclonal antibody against SMN; mouse monoclonal antibody against Gemin2; rat monoclonal antibody against galectin-3; human autoimmune serum reactive against the Sm epitopes on the core polypeptides of snRNPs; rabbit polyclonal antibodies to HMGs 14/17. In each panel, PI represents material precipitated by preimmune serum; αG1 represents material precipitated by rabbit anti-rat galectin-1; NE represents 50% of the nuclear extract subjected to immunoprecipitation, except the α galectin-1 immunoblot, in which the NE is ∼80% of that used for immunoprecipitation.
Control experiments using preimmune serum did not precipitate any of the proteins (Fig. 3, labeled PI). Moreover, we also tested and found that irrelevant nuclear proteins, such as high mobility group (HMG) 14/17, were not detectable in the anti-galectin-1 immunoprecipitate (Fig. 3, bottom). These results suggest that the polypeptides identified above represented bona fide components of macromolecular complexes co-precipitated by anti-galectin-1.
Effect of addition of the N-terminal domain of galectin-3 on the in vitro splicing reaction
It is interesting to note that galectin-3 is also found in the immunoprecipitate of anti-galectin-1 (Fig. 3). This is consistent with the observation that galectin-3 can interact directly with Gemin4 (Fig. 2B), which may, in turn, incorporate it into the macromolecular complex. The polypeptide of galectin-3 contains two domains: (i) a proline- and glycine-rich ND; and (ii) the carbohydrate-binding CD. We have isolated preparations of ND and CD and have carefully characterized them in terms of purity by SDS–PAGE, immunoblotting, mass spectrometry and amino acid sequence analysis. The availability of these reagents, representing fragments of the intact galectin-3 polypeptide, suggested the opportunity to test their effects in a cell-free splicing assay.
NE derived from HeLa cells exhibited strong splicing activity in our in vitro assay, converting >50% of the pre-mRNA substrate into the mature mRNA product (Fig. 4, lane 1). When the splicing reaction was carried out in the presence of exogenously added ND, there was a dose-dependent inhibition of splicing (Fig. 4, lanes 5–8). Inhibition of splicing was noticeable at a concentration as low as 1.2 µM (Fig. 4, lane 6). Complete inhibition was observed at 10 µM ND: there were barely detectable levels of products and splicing intermediates and substantially higher levels of the starting substrate (Fig. 4, lane 8).
Figure 4.

Comparison of the effect of addition of recombinant galectin-3 or its N- and C-terminal domains on the splicing activity of nuclear extract. Lane 1, nuclear extract (NE); lanes 2–4, recombinant galectin-3 (rG3) at 3, 6 and 12 µM added to NE; lanes 5–8, ND at 0.6, 1.2, 5 and 10 µM added to NE; lanes 9–12, CD at 3, 10, 28 and 57 µM added to NE. All reactions contained 32P-labeled MINX pre-mRNA substrate (5000 c.p.m.) and products of the splicing reactions were analyzed by electrophoresis through a 13% polyacrylamide–urea gel, followed by autoradiography. The positions of migration of pre-mRNA substrate, splicing intermediates (exon 1 and lariat–exon 2) and RNA products (lariat and ligated exon 1–exon 2) are highlighted on the right.
The splicing reaction was also carried out in the presence of exogenously added full-length recombinant galectin-3. Over a concentration range of 3–12 µM, the intact galectin-3 polypeptide had no effect on the in vitro splicing reaction (Fig. 4, lanes 2–4). Similarly, no inhibition was observed when the splicing reaction was carried out in the presence of exogenously added CD (Fig. 4, lanes 9–12). The concentration of CD tested ranged from 3 to 57 µM. Thus, the inhibitory effect of ND on the cell-free splicing assay was specific, inasmuch as parallel addition of either full-length galectin-3 or CD failed to yield the same effect.
Several possibilities were considered to account for the observed inhibition of splicing by ND. For example, the ND preparation could be contaminated by a protease that degraded essential components of the splicing machinery. On the basis of our previous experience showing that depletion of NE with antibodies directed against the Sm antigens of snRNPs resulted in the loss of splicing activity in this assay (11), we specifically tested for any effects of ND on the Sm antigens. Incubation of NE with ND, CD or full-length recombinant galectin-3 under the conditions of the splicing assay did not reduce the levels of the B, B′ and D polypeptides of the Sm antigens (data not shown).
We also tested for possible RNase activity in our protein preparations. The 32P-labeled pre-mRNA substrate was incubated with NE in the presence or absence of exogenous ND, CD and recombinant galectin-3. This incubation was carried out in the absence of ATP. (In the presence of ATP such incubation would simply correspond to a splicing assay and the pre-mRNA would be converted to the mature RNA product.) Surprisingly, the inclusion of any of our protein preparations (ND, CD or full-length recombinant galectin-3) resulted in a higher recovery of 32P-labeled pre-mRNA than incubation with NE alone. The basis for this better recovery is not known, but these results are consistent with the results of the splicing assay, in which substantially higher levels of the starting pre-mRNA substrate were found in reactions inhibited by ND (Fig. 4, lanes 7–8). In any case, the ND preparation did not degrade the pre-mRNA substrate.
Effect of addition of galectin-3 ND on spliceosome formation
The MINX pre-mRNA substrate forms several complexes, distinguished by the composition of the RNP components, during the course of the splicing reaction (22). Initially, the pre-mRNA is complexed with hnRNP proteins to form the H-complex. In the presence of ATP, specific snRNPs become associated with the H-complex to form the A and B active spliceosomal complexes. We have used the gel mobility shift assay to detect complexes formed by the splicing extract in the presence of exogenously added ND, CD or full-length recombinant galectin-3.
Under our electrophoretic conditions (10.7 cm gel, 150 V, 90 min), 32P-labeled pre-mRNA, in the absence of any proteins (no NE added), migrated off the bottom of the gel. When NE is added in the absence of ATP, only a radiolabeled smear corresponding to the H-complex (Fig. 5, lane 1) can be found, even after 5 min incubation. In the presence of ATP, the H-complex is also observed at 0 min (Fig. 5, lane 2); after 5 min incubation, bands corresponding to the higher order active complexes (A and B) become prominent (Fig. 5, lane 3). The summation of complexes A and B accounted for ∼70% of the radioactivity in lane 3 (Fig. 5). These higher order complexes are also observed when full-length recombinant galectin-3 (Fig. 5, lane 4) or CD (Fig. 5, lane 6) is added to the NE, pre-mRNA substrate and ATP for 5 min. In contrast, the addition of ND arrested the predominant portion of the radioactive pre-mRNA at the H-complex (Fig. 5, lane 5). The H-complex accounted for 83% of the radioactivity in lane 5 (Fig. 5). Although a small amount of radioactivity (∼16%) could be found at a position corresponding to the A-complex, no B-complex formation was evident. Finally, there was essentially no difference in the total amount of radioactivity between the samples shown in Figure 5.
Figure 5.
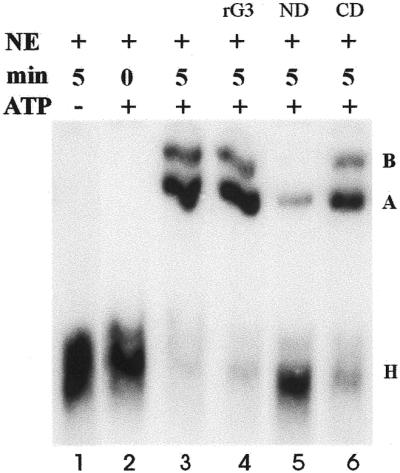
Comparison of the effect of addition of recombinant galectin-3 or its N- and C-terminal domains on formation of splicing complexes. All samples contained nuclear extract (NE) and 32P-labeled MINX pre-mRNA substrate (5000 c.p.m.). The sample in lane 1 was incubated for 5 min in the absence of ATP; the samples in lanes 2–6 were incubated in the presence of ATP. Lane 2, no additions to NE, 0 min; lane 3, no additions to NE, 5 min; lane 4, 12 µM recombinant galectin-3 (rG3), 5 min; lane 5, 10 µM ND, 5 min; lane 6, 10 µM CD, 5 min. Splicing complexes were analyzed by non-denaturing gel electrophoresis and autoradiography. The positions of the H-, A- and B-complexes (22) are highlighted on the right.
It should perhaps be noted that using the electrophoretic conditions described for native gels, the position in the gel indicated as the H-complex region actually consists of two complexes, the H- and E-complexes (27). Both are formed in the absence of ATP. H-complexes contain the splicing substrate and hnRNP proteins while E-complexes additionally incorporate U1 snRNP at the 5′ splice site (2). Therefore, our present use of the term H-complex is meant to indicate a region of the gel rather than a distinction between H- and E-complexes.
Effect of addition of GST–Gemin4(C50) on the splicing assay
Since the ND fragment of galectin-3 exerts a dominant negative effect when added to a splicing reaction, we tested whether a fragment of Gemin4, the other component of the galectin-3–Gemin4 interacting partners, might yield a similar effect. Two lines of evidence indicate that the C-terminal 50 amino acids of Gemin4 possessed sufficient structural information for proper folding and binding to other proteins: (i) galectin-1 interacted with this 50 amino acid fragment to activate the transcriptional reporter in the yeast two-hybrid screen (Fig. 1); and (ii) in vitro GST pull-down assays showed that Gemin4(C50) can bind both galectin-1 and galectin-3 (Fig. 2). Thus, Gemin4(C50) represents a fragment with retention of at least some of the binding properties of the full-length Gemin4 polypeptide.
The NE used in this experiment converted ∼30% of the pre-mRNA substrate into the mature mRNA product (Fig. 6, lane 1). When the splicing reaction was carried out in the presence of GST–Gemin4(C50), <2% conversion into the final product was observed (Fig. 6, lanes 4 and 5). No such inhibition was observed with corresponding concentrations of GST (mature mRNA product accounted for 30–40% of the radioactive RNA species; Fig. 6, lanes 2 and 3). The purities of the GST and GST–Gemin4(C50) protein preparations were monitored during our GST pull-down studies (see for example Fig. 2C). Neither preparation appeared to contain non-specific proteases inasmuch as little or no degradation of galectin-1 or galectin-3 was observed in those binding assays. Finally, neither the GST nor the GST–Gemin4(C50) protein preparations appeared to degrade the pre-mRNA substrate.
Figure 6.
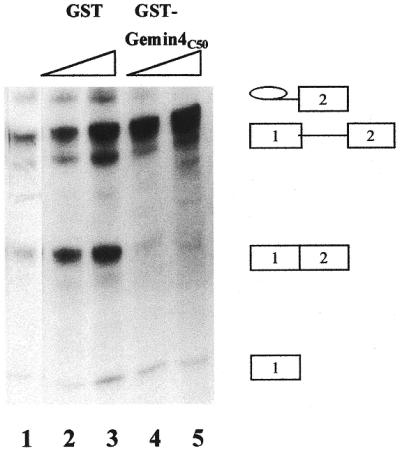
The effect of Gemin4(C50) addition on the splicing activity of nuclear extract. Lane 1, nuclear extract (NE); lanes 2 and 3, GST at 10 and 30 µM added to NE; lanes 4 and 5, GST–Gemin4(C50) at 10 and 30 µM added to NE. All reactions contained 32P-labeled MINX substrate (5000 c.p.m.) and products of the splicing reactions were analyzed by electrophoresis through a 13% polyacrylamide–urea gel, followed by autoradiography. The positions of migration of pre-mRNA substrate, splicing intermediates and RNA product are highlighted on the right.
DISCUSSION
The key findings of the present study include: (i) Gemin4 is an interacting partner of galectin-1 as identified through a yeast two-hybrid screen and it binds galectin-1 and galectin-3 directly in GST pull-down assays; (ii) galectin-1 and galectin-3 are associated with macromolecular complexes that also contain polypeptides previously identified with one or more steps of the pre-mRNA splicing process; (iii) fragments of either one of the interacting partners, Gemin4 or galectin-3, exert a dominant negative effect when added exogenously to the cell-free splicing assay; and (iv) inhibition of splicing by the galectin-3 ND appears to be due to a block at the H-complex in assembly of the active spliceosome. The key points of these findings are summarized in the schematic diagram of Figure 7.
Figure 7.

Schematic diagram summarizing the components of SMN-containing complexes and the intermediates in spliceosome assembly. The known and newly identified polypeptides of SMN complexes are delineated on the left. The steps in active spliceosome formation are shown on the right. The dotted rectangles represent exons and the horizontal line represents the intron on the pre-mRNA. The ATP-independent H- and E-complexes migrate to a position designated as the H-complex in our native gel electrophoretic system. The SMN-containing complexes are illustrated here to supply snRNPs to the H-complex, leading to the formation of ATP-dependent active spliceosomal A-, B- and C-complexes. This delivery may also involve galectin-1 and galectin-3 inasmuch as galectin depletion results in arrest of spliceosome assembly at the H-complex.
Three different lines of experiments now converge at the H-complex to implicate it as a site of action of galectins-1 and -3 in the splicing of pre-mRNA. First, we have identified galectin-1 and galectin-3 as novel components of macromolecular complexes containing SMN, Gemin2 and the core snRNP polypeptides bearing the Sm epitope (Fig. 7, left). Both galectins interact directly with at least one component of these complexes, Gemin4. The SMN-containing complexes are found in the cytoplasm and in discrete nuclear bodies called gems (gemini of coiled bodies) (28,29). In the former the SMN complex is associated with snRNP core proteins and plays a critical role in the biogenesis of snRNPs (30,31). In the nucleus SMN is required for pre-mRNA splicing by supplying snRNPs to the H-complex in the assembly of spliceosomes (26,32; Fig. 7, right). This H-complex juncture is indeed where galectins appear to be required, as demonstrated by the effect of galectin depletion and by the effect of galectin-3 ND addition.
Secondly, in previous studies (10) we had documented that NEs depleted of the galectins by affinity adsorption on lactose–agarose beads failed to form active spliceosomal complexes and gel mobility shift assays of 32P-labeled pre-mRNA revealed only bands migrating in the H-complex region. Moreover, the activities of the galectin-depleted extract, in forming splicing complexes and in performing the in vitro splicing reaction, were reconstituted by addition of recombinant galectin-3 with similar dose–response curves. On the basis of these results, we hypothesized that the galectins may be required at an early stage in the splicing pathway, such as organization of the H-complex for addition of other splicing factors (10,33). Interpreted in the context of our current findings, the effect of galectin depletion is most likely to be exerted at either the assembly of SMN-containing complexes (Fig. 7, left) or docking of such complexes at the H-complex (Fig. 7, right).
Finally, when the splicing assay was carried out in the presence of exogenously added ND of galectin-3, we also observed a dose-dependent inhibition of product formation. Again, this inhibition was associated with the arrest of spliceosome assembly at the H-complex. In both the in vitro splicing reaction and in assembly of active spliceosomes parallel additions of either the full-length galectin-3 polypeptide or the CD failed to yield the same effect.
Recently, the ND of galectin-3 has been expressed as a polypeptide fused to GST. Addition of this GST–ND fusion protein to our splicing assay also resulted in inhibition of splicing, as did addition of ND. In contrast, GST itself had no effect when added to the same splicing assay (see also Fig. 6). Thus, two preparations of the galectin-3 ND, expressed in distinct systems and purified by different procedures, both inhibited the splicing reaction.
Physico-chemical studies have been carried out on the mouse (14) and hamster (34) homologs of galectin-3, as well as isolated ND and CD preparations derived from the full-length polypeptide. These studies have indicated that the two domains of galectin-3 are structurally as well as functionally distinct and independent. For example, differential scanning calorimetry of mouse galectin-3 yielded distinct transition temperatures for ND (∼40°C) and CD (∼55°C), both in the full-length polypeptide and as isolated preparations of individual domains (14). Similarly, the circular dichroic spectrum of intact hamster galectin-3 is reproduced by an appropriately weighted summation of the spectra of the ND and CD fragments, suggesting that the isolated domains retain the structure that they had in the intact molecule (34).
Both of these studies, as well as studies on the human homolog (35,36), clearly show that the CD bears the carbohydrate-binding activity of the protein. In addition, the CD of galectin-3 has been shown to interact with a cytoplasmic cysteine- and histidine-rich protein (37). In previous studies (11) we had found that polypeptides corresponding to the CD (the CD of galectin-3 or galectin-1, which contains a single domain homologous to it) were sufficient to reconstitute splicing activity in a galectin-depleted NE. However, the minimum concentrations required for activity were four to eight times higher than that of the full-length galectin-3 polypeptide, which contains the proline- and glycine-rich ND.
This differential concentration effect on splicing reconstitution by intact galectin-3 versus its CD (or galectin-1) implies that both domains of galectin-3 interact with the splicing machinery. The nature of these interactions is unclear. Regardless of this, the observed splicing inhibition by excess ND likely occurs through perturbation of one set of these interactions and supports this suggestion.
The identification of Gemin4 as an interacting partner of galectin-1 and galectin-3 represents an important advance in our investigation of the role of galectins in the cell nucleus. First, it provides evidence that galectins-1 and -3 interact with splicing components in vivo, as had been inferred previously on the basis of depletion–reconstitution and co-localization experiments. The dominant negative effect on splicing exerted by Gemin4(C50) strengthens the notion that direct binding between the galectins and Gemin4 are functionally relevant in the splicing pathway. Secondly, it supports our previous notion that, mechanistically, the locus of galectin action is at the H-complex during assembly of spliceosomes. Finally, it offers new insights on the analysis of the composition and identities of polypeptides in the macromolecular complexes required for the splicing machinery and how galectin-1 or galectin-3 may facilitate this process.
Acknowledgments
ACKNOWLEDGEMENTS
We thank Kyle Openo for preparation of recombinant galectin-3 and its domains and Joe Leykam of the Macromolecular Structure, Sequencing and Synthesis Facility at Michigan State University for his help in characterizing the protein samples by mass spectrometry and amino acid sequence analysis. We also thank Drs S. Barondes, T. Feizi, J. Hirabayashi and K. Kasai for various galectin and antibody reagents detailed in the text. This work was supported by grants GM-38740 from the National Institutes of Health and MCB 97-23615 from the National Science Foundation.
References
- 1.Sharp P.A. (1994) Split genes and RNA splicing. Cell, 77, 805–815. [DOI] [PubMed] [Google Scholar]
- 2.Kramer A. (1996) The structure and function of proteins involved in mammalian pre-mRNA splicing. Annu. Rev. Biochem., 65, 367–409. [DOI] [PubMed] [Google Scholar]
- 3.Fu X.-D. (1995) The superfamily of arginine/serine-rich splicing factors. RNA, 1, 663–680. [PMC free article] [PubMed] [Google Scholar]
- 4.Raker V.A., Hartmuth,K., Kastner,B. and Luhrmann,R. (1999) Spliceosomal U snRNP core assembly: Sm proteins assemble onto an Sm site RNA nonanucleotide in a specific and thermodynamically stable manner. Mol. Cell. Biol., 19, 6554–6565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Reed R. (2000) Mechanisms of fidelity in pre-mRNA splicing. Curr. Opin. Cell Biol., 12, 340–345. [DOI] [PubMed] [Google Scholar]
- 6.Arenas J.E. and Abelson,J.N. (1997) Prp43: an RNA helicase-like factor involved in spliceosome disassembly. Proc. Natl Acad. Sci. USA, 94, 11798–11802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Leffler H. (1997) Introduction to galectins. Trends Glycosci. Glycotechnol., 9, 9–19. [Google Scholar]
- 8.Moutsatsos I.K., Davis,J.M., and Wang,J.L. (1986) Endogenous lectins from cultured cells: subcellular localization of carbohydrate-binding protein 35 in 3T3 fibroblasts. J. Cell Biol., 102, 477–483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Laing J.G. and Wang,J.L. (1988) Identification of carbohydrate binding protein 35 in heterogeneous nuclear ribonucleoprotein complex. Biochemistry, 27, 5329–5334. [DOI] [PubMed] [Google Scholar]
- 10.Dagher S.F., Wang,J.L. and Patterson,R.J. (1995) Identification of galectin-3 as a factor in pre-mRNA splicing. Proc. Natl Acad. Sci. USA, 92, 1213–1217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Vyakarnam A., Dagher,S.F., Wang,J.L. and Patterson,R.J. (1997) Evidence for a role for galectin-1 in pre-mRNA splicing. Mol. Cell. Biol., 17, 4730–4737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Vyakarnam A., Lenneman,A.J., Lakkides,K.M., Patterson,R.J. and Wang,J.L. (1998) A comparative nuclear localization study of galectin-1 with other splicing components. Exp. Cell Res., 242, 419–428. [DOI] [PubMed] [Google Scholar]
- 13.Hirabayashi J. and Kasai,K. (1991) Effect of amino acid substitution by site-directed mutagenesis on the carbohydrate recognition and stability of human 14 kDa β-galactoside-binding lectin. J. Biol. Chem., 266, 23648–23653. [PubMed] [Google Scholar]
- 14.Agrwal N., Sun,Q., Wang,S.-Y. and Wang,J.L. (1993) Carbohydrate binding protein 35. I. Properties of the recombinant polypeptide and the individuality of the domains. J. Biol. Chem., 268, 14931–14939. [PubMed] [Google Scholar]
- 15.Childs R.A. and Feizi,T. (1979) β-Galactoside-binding muscle lectins of man and monkey show antigenic cross-reactions with those of bovine origin. Biochem. J., 183, 755–758. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Dignam J.D., Lebovitz,R.M. and Roeder,R.G. (1983) Accurate transcription initiation by RNA polymerase II in a soluble extract from isolated mammalian nuclei. Nucleic Acids Res., 11, 1475–1489. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Bradford M.M. (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72, 248–254. [DOI] [PubMed] [Google Scholar]
- 18.Cooper D.N.W., Massa,S.M. and Barondes,S.H. (1991) Endogenous muscle lectin inhibits myoblast adhesion to laminin. J. Cell Biol., 115, 1437–1448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Laemmli U.K. (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227, 680–685. [DOI] [PubMed] [Google Scholar]
- 20.Ho M.K. and Springer,T.A. (1982) Mac-2, a novel 32,000 Mr mouse macrophage subpopulation-specific antigen defined by monoclonal antibodies. J. Immunol ., 128, 1221–1228. [PubMed] [Google Scholar]
- 21.Cherayil B.J., Weiner,S.J. and Pillai,S. (1989) The Mac-2 antigen is a galactoside specific lectin that binds IgE. J. Exp. Med., 170, 1959–1972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Zillman M., Zapp,M.L. and Berget,S.M. (1988) Gel electrophoretic isolation of splicing complexes containing U1 small nuclear ribonucleoprotein particles. Mol. Cell. Biol., 8, 814–821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Konarska M.M. and Sharp,P.A. (1986) Electrophoretic separation of complexes involved in the splicing of precursors to mRNAs. Cell, 46, 845–855. [DOI] [PubMed] [Google Scholar]
- 24.Fields S. and Song,O. (1989) A novel genetic system to detect protein-protein interactions. Nature, 340, 245–247. [DOI] [PubMed] [Google Scholar]
- 25.Charroux B., Pellizzoni,L., Perkinson,R.A., Yong,J., Shevchenko,A., Mann,M. and Dreyfuss,G. (2000) Gemin4: a novel component of the SMN complex that is found in both gems and nucleoli. J. Cell Biol., 148, 1177–1186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Meister G., Buhler,D., Laggerbauer,B., Zobawa,M., Lottspeich,F. and Fisher,U. (2000) Characterization of a nuclear 20S complex containing the survival of motor neurons (SMN) protein and a specific subset of spliceosomal Sm proteins. Hum. Mol. Genet., 9, 1977–1986. [DOI] [PubMed] [Google Scholar]
- 27.Michaud S. and Reed,R. (1993) A functional association between the 5′ and 3′ splice site is established in the earliest prespliceosome complex (E) in mammals. Genes Dev., 7, 1008–1020. [DOI] [PubMed] [Google Scholar]
- 28.Dietz H. (1998) Polishing the cutting edge of gems. Nature Genet., 20, 321–322. [DOI] [PubMed] [Google Scholar]
- 29.Matera A.G. and Frey,M.R. (1998) Nuclear structure 98. Coiled bodies and gems: janus or gemini? Am. J. Hum. Genet., 63, 317–321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Fisher U., Liu,Q. and Dreyfuss,G. (1997) The SMN-SIP1 complex has an essential role in spliceosomal snRNP biogenesis. Cell, 90, 1023–1029. [DOI] [PubMed] [Google Scholar]
- 31.Mattaj I.W. (1998) Ribonucleoprotein assembly: clues from spinal muscular atrophy. Curr. Biol., 8, 93–95. [DOI] [PubMed] [Google Scholar]
- 32.Pellizzoni L., Kataoka,N., Charroux,B. and Dreyfuss,G. (1998) A novel function for SMN, the spinal muscular atrophy disease gene product, in pre-mRNA splicing. Cell, 95, 615–624. [DOI] [PubMed] [Google Scholar]
- 33.Patterson R.J., Dagher,S.F., Vyakarnam,A. and Wang,J.L. (1997) Nuclear galectins: functionally redundant components in processing of pre-mRNA. Trends Glycosci. Glycotechnol., 9, 77–85. [Google Scholar]
- 34.Mehul B., Bawumia,S., Martin,S.R. and Hughes,R.C. (1994) Structure of baby hamster kidney carbohydrate-binding protein CBP30, an S-type animal lectin. J. Biol. Chem., 269, 18250–18258. [PubMed] [Google Scholar]
- 35.Hsu D.K., Zuberi,Z.I. and Liu,F.-T. (1992) Biochemical and biophysical chacracterization of human recombinant IgE-binding protein, an S-type lectin. J. Biol. Chem., 267, 14167–14174. [PubMed] [Google Scholar]
- 36.Ochieng J., Platt,D., Tait,L., Hogan,V., Raz,T., Carmi,P. and Raz,A. (1993) Structure-function relationship of a recombinant human galactoside-binding protein. Biochemistry, 32, 4455–4460. [DOI] [PubMed] [Google Scholar]
- 37.Menon R.P., Strom,M. and Hughes,R.C. (2000) Interaction of a novel cysteine and histidine-rich cytoplasmic protein with galectin-3 in a carbohydrate-independent manner. FEBS Lett., 470, 227–231. [DOI] [PubMed] [Google Scholar]