

Abstract
A sulfur-oxidizing chemolithoautotrophic bacterium, Sulfurovum lithotrophicum 42BKTT, isolated from hydrothermal sediments in Okinawa, Japan, has been used industrially for CO2 bio-mitigation owing to its ability to convert CO2 into C5H8NO4 − at a high rate of specific mitigation (0.42 g CO2/cell/h). The genome of S. lithotrophicum 42BKTT comprised of a single chromosome of 2217,891 bp with 2217 genes, including 2146 protein-coding genes and 54 RNA genes. Here, we present its complete genome-sequence information, including information about the genes encoding enzymes involved in CO2 fixation and sulfur oxidation.
Keywords: Complete genome, Sulfur-oxidizing bacterium, Chemolithoautotroph, CO2 bio-mitigation, Sulfurovum lithotrophicum
Introduction
10.1601/nm.3783 are well-known chemolithoautotrophic bacteria found in deep-sea hydrothermal fields that play significant roles in sulfur, nitrogen, and hydrogen flux [1, 2].
10.1601/nm.8874 42BKTT is a sulfur-oxidizing member of 10.1601/nm.3783 that was isolated from deep-sea hydrothermal sediments in Okinawa, Japan [3]. Strain 42BKTT is a Gram-negative, non-motile, and coccoid-to-short-rod-shaped bacterium that utilizes CO2 as a carbon source, S or S2O3 2− as electron donors, and O2 and NO3 − as electron acceptors [3, 4]. Recent studies have focused on its potential industrial applications for CO2 bio-mitigation, reporting that this strain could convert CO2 into C5H8NO4 − at a high specific mitigation rate of ~0.42 g CO2/cell/h [4].
The CO2-bio-mitigation ability of 10.1601/nm.8874 can be improved and optimized through genetic engineering; however, the present lack of genetic knowledge of 10.1601/nm.8874 renders the genetic engineering of this strain difficult. Here, we presented a preliminary description and the general features of 10.1601/nm.8874 42BKTT, along with its genome-sequence annotations and interactions with other 10.1601/nm.8873 species. This information would be helpful for improving the use of chemolithoautotrophic bacteria, including 10.1601/nm.8873 species, in industrial applications in CO2 bio-mitigation.
Organism information
Classification and features
A representative 16S rRNA gene of 10.1601/nm.8874 42BKTT was compared with that of other species using NCBI BLAST [5]. Figure 1 shows the phylogenetic tree with 10.1601/nm.8874 42BKTT, constructed based on the 16S rRNA sequence. This strain shared 99.1% (1393/1406 bp) and 95.1% (1312/1379) sequence identity with the 16S rRNA genes of 10.1601/nm.8873 sp. NBC37–1 [6] and 10.1601/nm.25804 Monchim33T, respectively.
Fig. 1.
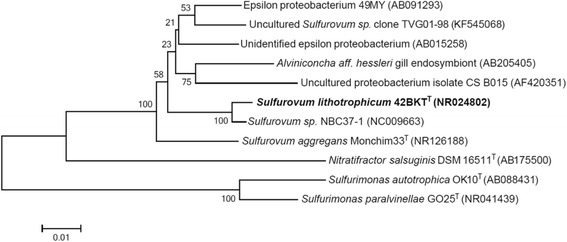
Phylogenetic tree showing the relative position of Sulfurovum lithotrophicum 42BKTT, based 16S rRNA gene sequence. All sites were informative and free of gaps. Evolutionary history was inferred using the neighbor-joining method [35]. The tree was built using the maximum composite-likelihood method [36]. The percentage of replicate trees with the associated taxa clustered together in the bootstrap test (1000 replicates) is shown next to the corresponding branches [37]. Evolutionary analyses were conducted in MEGA6 [38]. Corresponding GenBank accession numbers are shown in brackets next to the strain name
10.1601/nm.8874 42BKTT is a Gram-negative, non-motile, coccoid-to-short-rod-shaped bacterium that is 0.5–1.2 μm in length and 0.4–0.8 μm in width (Fig. 2). The 42BKTT strain is a mesophilic, facultative anaerobe that requires sea salt to grow and can use NH4Cl as a nitrogen source. Normal growth occurs at a temperature of 10–40 °C, pH of 5.0–9.0, and salinity of 5–60 g/l [3]. The basic details of its genome sequence are shown in Table 1.
Fig. 2.

Scanning electron micrograph of Sulfurovum lithotrophicum 42BKTT
Table 1.
Classification and general features of Sulfurovum lithotrophicum strain 42BKTT [11]
| MIGS ID | Property | Term | Evidence codea |
|---|---|---|---|
| Classification | Domain Bacteria | TAS [29] | |
| Phylum Proteobacteria | TAS [30] | ||
| Class Epsilonproteobacteria | TAS [31] | ||
| Order Campylobacterales | TAS [32] | ||
| Family Helicobacteraceae | TAS [33] | ||
| Genus Sulfurovum | TAS [3] | ||
| Species Sulfurovum lithotrophicum | TAS [3] | ||
| Type strain: 42BKTT (CP011308) | TAS [3] | ||
| Gram stain | Negative | TAS [3] | |
| Cell shape | Coccoid to short rods | TAS [3] | |
| Motility | None-motile | TAS [3] | |
| Sporulation | Not reported | NAS | |
| Temperature range | 10–40 °C | TAS [3] | |
| Optimum temperature | 28–30 °C | TAS [3] | |
| pH range; Optimum | 6.5–7.0 | TAS [3] | |
| Carbon source | Sodium bicarbonate | TAS [4] | |
| MIGS-6 | Habitat | Deep-sea hydrothermal vent | TAS [3] |
| MIGS-6.3 | Salinity | 0.5–6% NaCl (w/v) | TAS [3] |
| MIGS-22 | Oxygen requirement | Facultatively anaerobic | TAS [3] |
| MIGS-15 | Biotic relationship | Symbiont | TAS [3] |
| MIGS-14 | Pathogenicity | Not reported | NAS |
| MIGS-4 | Geographic location | Okinawa, Japan | TAS [3] |
| MIGS-5 | Sample collection | April 2002 | TAS [3] |
| MIGS-4.1 | Latitude | 27° 47·38′ N | TAS [3] |
| MIGS-4.2 | Longitude | 126° 53·87′ E | TAS [3] |
| MIGS-4.4 | Altitude | −1033 m | TAS [3] |
aEvidence codes - TAS Traceable Author Statement (i.e., a direct report exists in the literature); NAS Non-traceable Author Statement (i.e., not directly observed for the living, isolated sample, but based on a generally accepted property for the species or anecdotal evidence). These evidence codes are from the Gene Ontology project [34]
Chemotaxonomic data
The major cellular fatty acids that were present in strain 42BKTT included C16: 1 (53.7%), C16: 0 (31.3%), and C18: 0 (15.0%) [3]. It did not contain C14:0, C14:1, or C18:1, whereas 10.1601/nm.25804 Monchim33T contains 7.7, 5.9, and 9.4%, respectively, of these fatty acids [3, 7], and 10.1601/nm.3857 OK 10T, another chemolithoautotrophic bacteria, contains 8.4% of C14:0 and 9.4% of C18:1 [8]. 10.1601/nm.8874 42BKTT can fix CO2 via the reductive tricarboxylic acid (TCA) cycle, although the gene encoding phosphoenolpyruvate (PEP) carboxylase is not annotated in its genome. Sulfur or S2O3 2− are oxidized by bacteria of the genus 10.1601/nm.8873; 10.1601/nm.8874 42BKTT can oxidize S2− only using a sulfide-quinone reductase, whereas 10.1601/nm.8873 sp. NBC37–1 oxidizes S2− using a sulfide-quinone reductase or a sulfide dehydrogenase.
Genome sequencing information
Genome project history
10.1601/nm.8874 42BKTT was selected for sequencing based on its ability to convert CO2 into C5H8NO4 −, which can be industrially used for CO2 bio-mitigation. The draft sequencing and annotation were performed by ChunLab, Inc. (Seoul, Korea). The genome project was deposited in the Genomes OnLine Database [9] under the accession number Gp0118364. The complete genome sequence was also deposited in GenBank [10] under the accession number CP011308. Table 2 contains the details of the project and its association with MIGS version 2.0 compliance [11].
Table 2.
Project information
| MIGS ID | Property | Term |
|---|---|---|
| MIGS 31 | Finishing quality | Completely finished |
| MIGS 28 | Libraries used | Illumina 300-bp paired-end library, PacBio 20 K library |
| MIGS 29 | Sequencing platforms | Miseq PE 300, PacBio 10 K |
| MIGS 31.2 | Fold coverage | 852.21× |
| MIGS 30 | Assemblers | CLC Genomics Workbench v.7.5.1, SMRT Analysis v.2.3 |
| MIGS 32 | Gene-calling method | Prodigal 2.6.2 |
| Locus Tag | YH65 | |
| Genbank ID | CP011308.1 | |
| Genbank Date of Release | 08/20/2015 | |
| GOLD ID | Gp0118364 | |
| BIOPROJECT | PRJNA279430 | |
| MIGS 13 | Source-material identifier | 42BKTT/ ATCC BAA-797T |
| Project relevance | CO2 fixation |
Growth conditions and genomic DNA preparation
10.1601/nm.8874 42BKTT was grown in a 125-mL serum bottle (Wheaton Industries, Millville, NJ, USA) with 20 mL of MJ basal medium and filled with a CO2/N2 gas mixture. The bottle was incubated at 29 °C while shaking at 120 rpm (Green Shaker, Vision Scientific Co., Daejeon, Korea) [4]. Genomic DNA was isolated using a QIAmp DNA mini kit (Qiagen, Hilden, Germany), according to the manufacturer’s instructions.
Genome sequencing and assembly
The genomic library was sequenced using an Illumina MiSeq PE 300 and PacBio 10 K with the Illumina 300-bp paired-end library (Illumina, San Diego, CA, USA) and the PacBio 20 K library (Pacific Biosciences, Menlo Park, CA, USA), respectively. The generated paired-end sequencing reads (total read length: 2217,891 bp) were assembled using the CLC Genomics Workbench version 7.5.1 (CLC Bio, Aarhus, Denmark) and PacBio SMRT Analysis version 2.3 (Pacific Biosciences), resulting in one contig with an average genome coverage of 852.21 × .
Genome annotation
The genome was annotated using the NCBI Prokaryotic Genome Annotation Pipeline [12], which was designed to annotate bacterial genomes. Genome annotation was performed by predicting protein-coding, rRNA, tRNA, ncRNA, and pseudo genes. Phobius [13] was used to predict signal-peptide genes, and TMHMM Server version 2.0 [14] was used to predict transmembrane helix genes [15, 16]. Protein families [17] were investigated using Pfam 29.0 [18], and GeneMarkS+ [19], which uses alignment data for gene prediction, was used as an annotation tool [20].
Genome properties
The genome of 10.1601/nm.8874 42BKTT comprised a single circular chromosome of 2217,891 bp with a GC content of 44.26%. Among the 2217 genes predicted, 2146 (96.80%) were protein-coding DNA sequences, 17 of which were pseudogenes. Among the CDSs, 89.66% were grouped into cluster of orthologous group functional categories. The genome contained a CRISPR array and 54 RNA genes, including 44 tRNAs, 9 rRNAs, and one ncRNA. The properties and statistics of the genome are summarized in Fig. 3 and Tables 3 and 4, 5.
Fig. 3.
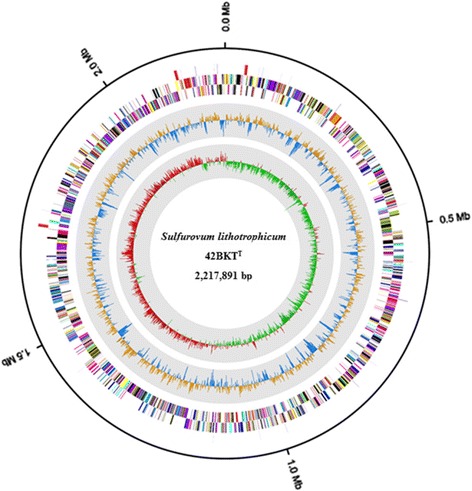
Genome map of Sulfurovum lithotrophicum 42BKTT. From the outer to the inner circle: RNA regions (rRNA, red; tRNA, lavender), CDS on the reverse strand (colored based on COG categories), CDS on the forward strand (colored based on COG categories), G + C skew (blue/goldenrod), and GC ratio (green/red)
Table 3.
Genome statistics
| Attribute | Value | % of total |
|---|---|---|
| Genome size (bp) | 2217,891 | 100.00 |
| DNA coding (bp) | 2,028,222 | 91.44 |
| DNA G + C (bp) | 981,638 | 44.26 |
| DNA scaffolds | 1 | |
| Total genes | 2217 | 100.00 |
| Protein-coding genes | 2146 | 96.80 |
| RNA genes | 54 | 2.44 |
| Pseudo genes | 17 | 0.77 |
| Genes in internal clusters | NA | NA |
| Genes with function prediction | 1559 | 70.32 |
| Genes assigned to COGs | 1979 | 89.26 |
| Genes with Pfam domains | 1770 | 79.84 |
| Genes with signal peptides | 412 | 18.58 |
| Genes with transmembrane helices | 513 | 23.14 |
| CRISPR repeats | 1 |
Table 4.
Number of genes associated with the general COG functional categories
| Code | Value | % agea | Description |
|---|---|---|---|
| J | 138 | 6.43 | Translation, ribosomal structure, and biogenesis |
| A | 0 | 0.00 | RNA processing and modification |
| K | 47 | 2.19 | Transcription |
| L | 94 | 4.38 | Replication, recombination, and repair |
| B | 1 | 0.05 | Chromatin structure and dynamics |
| D | 14 | 0.65 | Cell cycle control, cell division, chromosome partitioning |
| V | 18 | 0.84 | Defense mechanisms |
| T | 88 | 4.10 | Signal-transduction mechanisms |
| M | 144 | 6.71 | Cell wall/membrane/envelope biogenesis |
| N | 6 | 0.28 | Cell motility |
| U | 39 | 1.82 | Intracellular trafficking and secretion |
| O | 95 | 4.43 | Post-translational modification, protein turnover, chaperones |
| C | 138 | 6.43 | Energy production and conversion |
| G | 53 | 2.47 | Carbohydrate transport and metabolism |
| E | 119 | 5.55 | Amino acid transport and metabolism |
| F | 60 | 2.80 | Nucleotide transport and metabolism |
| H | 85 | 3.96 | Coenzyme transport and metabolism |
| I | 43 | 2.00 | Lipid transport and metabolism |
| P | 106 | 4.94 | Inorganic ion transport and metabolism |
| Q | 22 | 1.03 | Secondary metabolites biosynthesis, transport and catabolism |
| R | 143 | 6.66 | General function prediction only |
| S | 526 | 24.51 | Function unknown |
| - | 238 | 11.09 | Not in COGs |
aPercentage of the total number of protein-coding genes in the genome
Table 5.
Species in the genus Sulfurovum
| Species (isolation source) | Genome size (Mb) | Accession no. | CDS | GC (%) | Reference |
|---|---|---|---|---|---|
|
Sulfurovum lithotrophicum 42BKTT
(Deep-sea hydrothermal sediment) |
2.21 | CP011308 | 2092 | 44.3 | This report |
|
Sulfurovum sp. NBC37–1 (Deep-sea hydrothermal vent) |
2.56 | AP009179 | 2466 | 43.8 | [6] |
|
Candidatus Sulfurovum sediminum AR (Marine sediment) |
2.12 | AJLE01000000 | 2114 | 39.2 | [26] |
Insights from the genome sequence
10.1601/nm.8874 42BKTT is a sulfur-oxidizing bacterium that can fix CO2 through the reductive TCA cycle. Here, we focused on investigating its abilities for CO2 fixation and sulfur oxidation (sox), based on its genome sequence.
So far, six pathways have been associated with CO2 fixation: the Calvin-Benson-Bassham or reductive pentose pathway, the reductive TCA cycle or reverse citric acid cycle, the reductive acetyl CoA or Wood-Ljungdahl pathway, the 3-hydroxypropionate pathway or malyl CoA pathway, the 3-hydroxypropionate/4-hydroxy-butyrate cycle, and the dicarboxylate/4-hydroxybutyrate cycle [21, 22]. Similar to the majority of 10.1601/nm.3783, 10.1601/nm.8874 42BKTT can also grow chemoautotrophically through its adenosine triphosphate citrate lyase, 2-oxoglutarate:ferredoxin oxidoreductase, and pyruvate:ferredoxin oxidoreductase via the reductive TCA cycle [23–25]. We annotated these three key enzymes, as well as other relevant enzymes such as malate dehydrogenase, fumarate hydratase, fumarate reductase, isocitrate dehydrogenase, aconitate hydratase, PEP synthase, and PEP carboxylase, in the genome sequence of 42BKTT. Notably, 10.1601/nm.8873 sp. NBC37–1 and Candidatus Sulfurovum sediminum AR could also assimilate CO2 via the reductive TCA cycle [6, 26].
10.1601/nm.8874 42BKTT is known to oxidize or S2S O3 2− via a sox system using SoxB, SoxXA, SoxYZ, and Sox(CD)2 periplasmic proteins [27]. These enzymes catalyze the oxidation of S or S2O3 2− using horse cytochrome c as the final electron acceptor [28]. Here, we confirmed the presence of SoxA, SoxB, SoxZ, SoxY, and SoxX genes in the 42BKTT genome.
Conclusions
To the best of our knowledge, this is the first report describing the genome sequence of 10.1601/nm.8874 42BKTT, which comprised a circular chromosome of 2217,891 bp (44.26% GC content) with 2217 genes, among which 2146 were CDSs, 17 were pseudogenes, and 54 were RNA genes. 10.1601/nm.8874 42BKTT assimilates CO2 via the reductive TCA cycle and oxidizes S or S2O3 2− via the sox system. The details of the genome sequence of this strain could provide potential strategies to enhance the industrial application of such bacteria for CO2 bio-mitigation.
Acknowledgments
Funding
This study was supported by a grant from the KRIBB Research Initiative Program, and Industrial Strategic Technology Development Program (10067772, Development of bio-glutaric acid based plasticizers) funded by the Ministry of Trade, Industry & Energy (MOTIE, Korea).
Abbreviations
- CDS
Coding DNA sequence
- COG
Cluster of orthologous group
- PEP
Phosphoenolpyruvate
- TCA
Tricarboxylic acid
Authors’ contributions
WJ and GP performed the microbial cultivation and genomic DNA isolation. LP and HL performed the phylogenetic analysis. WJ, LP, and NL performed sequencing and data analysis. WJ, LP, and JA drafted the manuscript. DL, HK, IA, CL, HL, and JA edited the manuscript. All the authors have read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Nakagawa S, Takai K, Inagaki F, Hirayama H, Nunoura T, Horikoshi K, Sako Y. Distribution, phylogenetic diversity and physiological characteristics of epsilon-Proteobacteria in a deep-sea hydrothermal field. Environ Microbiol. 2005;7:1619–1632. doi: 10.1111/j.1462-2920.2005.00856.x. [DOI] [PubMed] [Google Scholar]
- 2.Huber JA, Butterfield DA, Baross JA. Bacterial diversity in a subseafloor habitat following a deep-sea volcanic eruption. FEMS Microbiol Ecol. 2003;43:393–409. doi: 10.1111/j.1574-6941.2003.tb01080.x. [DOI] [PubMed] [Google Scholar]
- 3.Inagaki F, Takai K, Nealson KH, Horikoshi K. Sulfurovum lithotrophicum gen. nov., sp. nov., a novel sulfur-oxidizing chemolithoautotroph within the epsilon-Proteobacteria isolated from Okinawa trough hydrothermal sediments. Int J Syst Evol Microbiol. 2004;54:1477–1482. doi: 10.1099/ijs.0.03042-0. [DOI] [PubMed] [Google Scholar]
- 4.Kwon HS, Lee JH, Kim T, Kim JJ, Jeon P, Lee CH, Ahn IS. Biofixation of a high-concentration of carbon dioxide using a deep-sea bacterium: Sulfurovum lithotrophicum 42BKTT. RSC Adv. 2015;5:7151–7159. doi: 10.1039/C4RA07830F. [DOI] [Google Scholar]
- 5.NCBI BLAST. https://blast.ncbi.nlm.nih.gov/Blast.cgi. Accessed 17 Jan 2017.
- 6.Nakagawa S, Takaki Y, Shimamura S, Reysenbach AL, Takai K, Horikoshi K. Deep-sea vent epsilon-proteobacterial genomes provide insights into emergence of pathogens. Proc Natl Acad Sci U S A. 2007;104:12146–12150. doi: 10.1073/pnas.0700687104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Mino S, Kudo H, Arai T, Sawabe T, Takai K, Nakagawa S. Sulfurovum aggregans sp. nov.,a hydrogen-oxidizing, thiosulfate-reducing chemolithoautotroph within the Epsilonproteobacteria isolated from a deep-sea hydrothermal vent chimney, and an emended description of the genus Sulfurovum. Int J Syst Evol Microbiol. 2014;64:3195–3201. doi: 10.1099/ijs.0.065094-0. [DOI] [PubMed] [Google Scholar]
- 8.Inagaki F, Takai K, Kobayashi H, Nealson KH, Horikoshi K. Sulfurimonas autotrophica gen. nov., sp. nov., a novel sulfur-oxidizing epsilon-proteobacterium isolated from hydrothermal sediments in the Mid-Okinawa Trough. Int J Syst Evol Microbiol. 2003;53:1801–1805. doi: 10.1099/ijs.0.02682-0. [DOI] [PubMed] [Google Scholar]
- 9.Genomes OnLine Database. https://gold.jgi.doe.gov/. Accessed 17 Jan 2017.
- 10.GenBank. https://www.ncbi.nlm.nih.gov/genbank/. Accessed 17 Jan 2017.
- 11.Field D, Garrity G, Gray T, Morrison N, Selengut J, Sterk P, Tatusova T, Thomson N, Allen MJ. Angiuoli SV and others. The minimum information about a genome sequence (MIGS) specification. Nat Biotechnol. 2008;26:541–547. doi: 10.1038/nbt1360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.NCBI Prokaryotic Genome Annotation Pipeline. https://www.ncbi.nlm.nih.gov/genome/annotation_prok/. Accessed 17 Jan 2017.
- 13.Phobius. http://phobius.sbc.su.se/. Accessed 17 Jan 2017.
- 14.TMHMM Server version 2.0. http://www.cbs.dtu.dk/services/TMHMM/. Accessed 17 Jan 2017.
- 15.Kall L, Krogh A, Sonnhammer EL. Advantages of combined transmembrane topology and signal peptide prediction-the Phobius web server. Nucleic Acids Res. 2007;35:W429–W432. doi: 10.1093/nar/gkm256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Krogh A, Larsson B, von Heijne G, Sonnhammer EL. Predicting transmembrane protein topology with a hidden Markov model: application to complete genomes. J Mol Biol. 2001;305:567–580. doi: 10.1006/jmbi.2000.4315. [DOI] [PubMed] [Google Scholar]
- 17.Bateman A, Birney E, Durbin R, Eddy SR, Howe KL, Sonnhammer ELL. The Pfam protein families database. Nucleic Acids Res. 2000;28:263–266. doi: 10.1093/nar/28.1.263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Pfam 29.0. http://pfam.xfam.org/. Accessed 17 Jan 2017.
- 19.GeneMarkS+. http://exon.gatech.edu/Genemark/genemarks.cgi. Accessed 17 Jan 2017.
- 20.Besemer J, Lomsadze A, Borodovsky M. GeneMarkS: a self-training method for prediction of gene starts in microbial genomes. Implications for finding sequence motifs in regulatory regions. Nucleic Acids Res. 2001;29:2607–2618. doi: 10.1093/nar/29.12.2607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Saini R, Kapoor R, Kumar R, Siddiqi TO, Kumar A. CO(2) utilizing microbes--a comprehensive review. Biotechnol Adv. 2011;29:949–960. doi: 10.1016/j.biotechadv.2011.08.009. [DOI] [PubMed] [Google Scholar]
- 22.Kanao T, Fukui T, Atomi H, Imanaka T. ATP-citrate lyase from the green sulfur bacterium Chlorobium limicola is a heteromeric enzyme composed of two distinct gene products. Eur J Biochem. 2001;268:1670–1678. doi: 10.1046/j.1432-1327.2001.02034.x. [DOI] [PubMed] [Google Scholar]
- 23.Hügler M, Gärtner A, Imhoff JF. Functional genes as markers for sulfur cycling and CO2 fixation in microbial communities of hydrothermal vents of the Logatchev field. FEMS Microbiol Ecol. 2010;73:526–537. doi: 10.1111/j.1574-6941.2010.00919.x. [DOI] [PubMed] [Google Scholar]
- 24.Hugler M, Wirsen CO, Fuchs G, Taylor CD, Sievert SM. Evidence for autotrophic CO2 fixation via the reductive tricarboxylic acid cycle by members of the epsilon subdivision of proteobacteria. J Bacteriol. 2005;187:3020–3027. doi: 10.1128/JB.187.9.3020-3027.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Takai K, Campbell BJ, Cary SC, Suzuki M, Oida H, Nunoura T, Hirayama H, Nakagawa S, Suzuki Y, Inagaki F, et al. Enzymatic and genetic characterization of carbon and energy metabolisms by deep-sea hydrothermal chemolithoautotrophic isolates of Epsilonproteobacteria. Appl Environ Microbiol. 2005;71:7310–7320. doi: 10.1128/AEM.71.11.7310-7320.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Park SJ, Ghai R, Martin-Cuadrado AB, Rodriguez-Valera F, Jung MY, Kim JG, Rhee SK. Draft genome sequence of the sulfur-oxidizing bacterium “Candidatus Sulfurovum sediminum” AR, which belongs to the Epsilonproteobacteria. J Bacteriol. 2012;194:4128–4129. doi: 10.1128/JB.00741-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Friedrich CG, Bardischewsky F, Rother D, Quentmeier A, Fischer J. Prokaryotic sulfur oxidation. Curr Opin Microbiol. 2005;8:253–259. doi: 10.1016/j.mib.2005.04.005. [DOI] [PubMed] [Google Scholar]
- 28.Bardischewsky F, Quentmeier A, Rother D, Hellwig P, Kostka S, Friedrich CG. Sulfur dehydrogenase of Paracoccus pantotrophus: the heme-2 domain of the molybdoprotein cytochrome c complex is dispensable for catalytic activity. Biochemistry. 2005;44:7024–7034. doi: 10.1021/bi047334b. [DOI] [PubMed] [Google Scholar]
- 29.Woese CR, Kandler O, Wheelis ML. Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya. Proc Natl Acad Sci U S A. 1980;87:4576–4579. doi: 10.1073/pnas.87.12.4576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Garrity GM, Bell JA, Phylum LT. Bergey’s manual of systematic bacteriology. 2005. XIV. Proteobacteria phyl. nov; p. 1. [Google Scholar]
- 31.Garrity GM, Bell JA, Lilburn T. Bergey’s manual of systematic bacteriology. 2005. Class V. Epsilonproteobacteria class. nov; p. 1145. [Google Scholar]
- 32.Garrity GM, Bell JA, Lilburn T. Bergey’s manual of systematic bacteriology. 2005. Order I. Campylobacterales ord. nov; p. 1145. [Google Scholar]
- 33.Garrity GM, Bell JA, Lilburn T. Bergey’s manual of systematic bacteriology. 2005. Family II. Helicobacteraceae fam. nov; p. 1168. [Google Scholar]
- 34.Ashburner M, Ball CA, Blake JA, Botstein D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT, et al. Gene ontology: tool for the unification of biology. The gene ontology consortium. Nat Genet. 2000;25:25–29. doi: 10.1038/75556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Saitou N, Nei M. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol. 1987;4:406–425. doi: 10.1093/oxfordjournals.molbev.a040454. [DOI] [PubMed] [Google Scholar]
- 36.Tamura K, Nei M, Kumar S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc Natl Acad Sci U S A. 2004;101:11030–11035. doi: 10.1073/pnas.0404206101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Felsenstein J. Confidence limits on phylogenies: an approach using the bootstrap. Evolution. 1985;39:783–791. doi: 10.1111/j.1558-5646.1985.tb00420.x. [DOI] [PubMed] [Google Scholar]
- 38.Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. MEGA6: molecular evolutionary genetics analysis version 6.0. Mol Biol Evol. 2013;30:2725–2729. doi: 10.1093/molbev/mst197. [DOI] [PMC free article] [PubMed] [Google Scholar]