


Abstract
Human insulin-like growth factor II (IGF-II) mRNA can be cleaved at a specific site in its 4 kb long 3′-UTR. This yields a stable 3′ cleavage product of 1.8 kb consisting of a 3′-UTR and a poly(A) tail and an unstable 5′ cleavage product containing the IGF-II coding region. After cleavage, the 5′ cleavage product is targeted to rapid degradation and consequently is no longer involved in IGF-II protein synthesis. Cleavage is therefore thought to provide an additional way to control IGF-II gene expression. In this paper the kinetics and the efficiency of cleavage of IGF-II mRNAs are examined. The cleavage efficiency of IGF-II mRNAs carrying four different leaders (L1–L4) is enhanced in the highly structured leaders L1 and L3. Additionally, under standard cell culture conditions cleavage is a slow process that only plays a limited role in destabilisation and translation of the IGF-II mRNAs. However, in human Hep3B cells and CaCo2 cells which express IGF-II endogenously, cleavage is upregulated 3–5-fold at high cell densities. Regulated endonucleolytic cleavage of IGF-II mRNAs is restricted to cells in which IGF-II expression is related to specific cell processes.
INTRODUCTION
Human insulin-like growth factor II (IGF-II) is a mitogenic polypeptide of 67 amino acids with important functions in cell growth and development (1). Depending on the cell type in which it is expressed, IGF-II can exhibit remarkably different activities. IGF-II can promote either cell proliferation or differentiation; it strongly affects cellular survival and counteracts apoptosis in some cell systems, whereas in other cell lines an apoptosis-inducing effect by IGF-II is observed (2 and references therein). Thus, as IGF-II is involved in many physiological processes, a proper regulation of its expression is of crucial importance. Indeed, IGF-II gene expression is regulated at virtually all levels, ranging from developmental stage-dependent and tissue-specific activity of four different promoters to post-translational processing of the IGF-II precursor protein (reviewed in 3). The human IGF-II gene contains four promoters, P1–P4, and the mRNAs transcribed from these promoters range in size from 2.2 to 6.0 kb. Previously, we have observed that in addition to the different mRNAs that are transcribed from the four promoters, a smaller RNA species of 1.8 kb is formed (4). This RNA is the product of endonucleolytic cleavage at a specific site in the 3′-UTR of full-length IGF-II mRNA (5). In addition to this 1.8 kb RNA, which represents the 3′ cleavage product, a 5′ cleavage product that contains the coding region of the IGF-II mRNA is also formed. Similar observations were made for the rat IGF-II gene (6) and the mouse IGF-II gene (7). Because the 5′ cleavage product lacks a poly(A) tail, it is very unstable and not translated (6). Thus, as cleavage abolishes the protein-coding potential of a particular mRNA, it provides the cell with a putative additional mechanism to control IGF-II protein production, acting at the level of mRNA stability.
In recent years, it has become clear that gene regulation at the level of mRNA stability is very common (8–11). Initiation of mRNA degradation can occur through poly(A) shortening, arrest of translation at a premature stop codon [nonsense-mediated decay, (NMD)], or through endonucleolytic cleavage. To date, specific endonucleolytic cleavage has been observed for a number of mRNAs. Examples include the mRNAs encoding albumin (12,13), the cytokine groα (14), avian apo-very-low density lipoprotein II (15), the transferrin receptor (16) and the Xenopus homeodomain protein Xlhbox2 (17,18). The c-myc mRNA can be cleaved in the coding region, but can also be destabilised through AU-rich elements in the 3′-UTR (19,20). Recently, it has been reported that the human α-globin mRNA can also be cleaved at a specific site in the 3′-UTR (21).
Our previous studies were aimed at elucidating the determinants for recognition of the IGF-II mRNA cleavage site by an endoribonuclease. We have found that extensive RNA secondary structures formed by two widely separated sequence elements are required for cleavage (22). These RNA structures act co-operatively to maintain a sequence- and structure-specific cleavage site located in an RNA internal loop (23,24).
The aim of the present study was to obtain a better insight into the kinetics of specific endonucleolytic cleavage of the IGF-II mRNAs and the impact on the overall mRNA decay rate and protein production.
MATERIALS AND METHODS
Materials
Expression plasmids pTet-Splice and pTet-tTak were purchased from Gibco BRL Life Technologies. Plasmid pGL3 was from Promega. Tetracycline was purchased from Sigma (St Louis, MO). Restriction enzymes and T4 DNA ligase were purchased from Roche Molecular Biochemicals (Mannheim, Germany). Enzymes were used as specified by the manufacturers. dNTPs were obtained from Kabi-Pharmacia; BES [N,N-bis (2-hydroxyethyl)-2-aminoethanesulfonic acid] was from Sigma. A random primer labelling kit and a DNA sequencing kit were purchased from Amersham Pharmacia Biotech (Little Chalfont, UK). A poly(A)+ RNA isolation kit was purchased from Promega, RNAzol reagent was purchased from Biotecx Laboratories, Inc. (Houston, TX) and GeneScreen membranes were purchased from Du Pont de Nemours (Dreieich, Germany). [α-32P]dCTP (3000 Ci/mmol) was purchased from Amersham. Trichostatin A and dexamethasone were purchased from ICN Biomedicals, Inc. (Costa Mesa, CA) and Sigma, respectively. TCH‘, a serum-free defined media supplement was purchased from Celex Co. (Minnetonka, MN).
Construction of plasmids
Molecular cloning was performed according to established protocols (25). All positions within exon 9 of the IGF-II gene are indicated relative to the cleavage site; the nucleotide upstream of the cleavage site is –1; the nucleotide downstream is +1. When exon 9 sequences were deleted or additional restriction sites were introduced, the numbers indicated still refer to their original position in exon 9 relative to the cleavage site.
The leader plasmids were constructed as follows. The IGF-II expression plasmid EP7-9/Not (26) was digested with Alw44I to generate a fragment containing the IGF-II 3′-UTR sequence from position –17 relative to the cleavage site, to 210 nt downstream of the polyadenylation site; this fragment was inserted in the XbaI site of expression plasmid pSCT-GalX556, which contains the cytomegalovirus (CMV) promoter (27). The resulting plasmid, designated pCMV-1.8, was digested with BamHI and NotI (position +80). DNA containing PCR-generated leader sequences L1, -L2, -L3 and -L4 and the IGF-II open reading frame (28) was digested with BamHI and XhoI to generate fragments containing the 5′-UTR and the IGF-II coding region. Plasmid EP7-9/Not was digested with XhoI and NotI (positions –2291 and +80). The resulting fragment was first ligated with the leader-fragments and subsequently, the BamHI/NotI digested plasmid pCMV-1.8 was added to the ligation reaction mixtures followed by further ligation. This resulted in plasmids pCMV-L1, pCMV-L2, pCMV-L3 and pCMV-L4.
Plasmid pTet-L4-ΔWT was constructed as follows. Plasmid pTet-Splice was digested with XhoI and BamHI to generate a fragment containing the tetracycline-responsive minimal CMV promoter. This fragment was inserted in the AatII and BamHI sites of plasmid CMV-L4 that encompass the CMV promoter, resulting in plasmid pTet-L4. Plasmids ΔWT and ΔWTmut were digested with ClaI and XbaI (positions –1961 and +1051 in the 3′-UTR) and the resulting fragments were inserted in the ClaI and XbaI sites of plasmid pTet-L4. Construct ΔWTmut contains the following mutations: A(–144) and G(–143) have been deleted and U(–142), A(–141), G(–140) have been changed to C, U and C, respectively (23).
To generate plasmids pTet-L4-luc-ΔWT and pTet-L4-luc-ΔWTmut, the following procedure was followed. First, an XhoI–XbaI fragment of the IGF-II 3′-UTR (from position –2292 to +1047) was inserted in the XbaI site of the luciferase expression plasmid pGL3 (directly downstream of the luciferase coding region). The resulting plasmid was digested with HindIII and ClaI, yielding a fragment that contains the luciferase open reading frame and the IGF-II sequence from positions –2292 to –1961. This fragment was inserted in the HindIII/ClaI sites of plasmids pTet-L4-ΔWT and pTet-L4-ΔWTmut.
Culture and transient transfection of 293 cells
Human 293 cells were grown in Dulbecco’s Modified Eagle’s Medium (DMEM). The medium was supplemented with 10% fetal calf serum (FCS), 100 IU/ml penicillin, 100 µg/ml streptomycin and 300 µg/ml glutamine. The cells were transfected in the absence of tetracycline in 25 cm2 flasks at a confluence of 50% by the calcium phosphate coprecipitation method (29). Precipitates were prepared with BES-buffered saline (25) and contained 3.3 µg pTet-IGF-II constructs and 3.3 µg pTet-tTak (30) and were added to the cells. After 4 h, the medium was aspirated and replaced with fresh serum-containing medium. After 24 h, the tetracycline was removed to induce expression of the constructs. At various time points after transcription induction, the cells were washed with phosphate-buffered saline (PBS) and harvested in PBS with 0.025% trypsin, 0.02% EDTA. For the reverse experiment to determine the effect of cleavage on protein production, 3.3 µg pTet-L4-luc-ΔWT or pTet-L4-ΔWTmut was cotransfected with pTet-tTak in the absence of tetracycline. After 4 h, the medium was refreshed and 30 h later, cell extracts were prepared for luciferase assays (25,30).
Culture of Hep3B, CaCo2 and Ltk– cells
Human Hep3B cells were grown in α-Minimal Essential Medium (α-MEM) and 2% of the defined serum replacement TCH‘ or 10% FCS. Human CaCo2 cells were grown in high glucose (25mmol/l) DMEM with 10% FCS and murine Ltk– cells were grown in DMEM with 10% FCS. All cell culture media were supplemented with 100 IU/ml penicillin, 100 µg/ml streptomycin and 300 µg/ml glutamine.
To study the effect of serum/growth factors in Hep3B cells, cells growing on 10% FCS at a density of 50% confluence were deprived of serum by replacing the serum-containing medium with serum-free medium. At various time points ranging from 4 to 32 h, the cells were harvested for RNA isolation. The reverse experiment was also performed; cells growing in TCH‘ containing medium were supplied with 10% FCS and harvested at various time points ranging from 4 to 32 h thereafter.
The effect of the growth conditions in Hep3B cells and CaCo2 cells was examined as follows. Cells were seeded at a density of 1 × 104 cells/cm2 in 25 cm2 flasks and grown to a variety of different densities. Medium was refreshed daily. Cells were counted each day using a counting chamber after resuspending them in PBS with 0.025% trypsin, 0.02% EDTA. After harvesting the cells, RNA was isolated and subjected to northern blot analysis.
RNA isolation and analysis
Total RNA was isolated as described previously (24). The blots were probed with different radiolabelled DNA fragments. To detect the full-length IGF-II mRNAs as well as the 3′ cleavage product, a 1.0 kb SmaI fragment from the human IGF-II exon 9 (position +84 to +1096), hybridising to both the full-length IGF-II RNAs and the 3′ cleavage product, was used. To detect full-length IGF-II mRNAs as well as the 5′ cleavage product but not the 3′ product, a 970 bp IGF-II coding region cDNA fragment was used (31). To detect the GAPDH mRNA, a 1300 nt long GAPDH cDNA fragment was used. The DNA fragments were labelled by random priming with [α-32P]dCTP following the manufacturer’s protocols. After 2 h of pre-hybridisation, the probes were added to a final concentration of 106 c.p.m./ml. Blots were washed after overnight hybridisation, to a final stringency of 0.5× SSC, 1% SDS at 65°C (1× SSC is 0.15 M NaCl, 0.015 M sodium citrate) and exposed to Fuji RX X-ray film. The blots were also probed with a [γ-32P]ATP 5′-end-labelled oligonucleotide, 5′-AACGATCAGAGTAGTGGTATTTCACC-3′, that is complementary to 28S ribosomal RNA. Here, a slightly different procedure was used for hybridisation and washing of the blots. The blots were hybridised in the presence of 25% formamide and were washed to a final stringency of 1× SSC without SDS at 30°C. The amount of cleavage products was determined by phosphorimager scanning of the blots of at least three independent experiments. Cleavage efficiencies were determined by calculating the amount of 3′ cleavage product divided by the total amount of IGF-II RNA. Variation between the calculated cleavage efficiencies in separate experiments was <10%.
Poly(A)+ RNA was isolated from total RNA preparations according to the manufacturer’s instructions.
RESULTS
Effect of the various 5′ leaders on the efficiency of specific IGF-II mRNA endonucleolytic cleavage
The human IGF-II gene contains four promoters (P1–P4), which are differentially active during development and in different tissues. Transcription from these four promoters results in transcripts that differ in their 5′-UTRs (leaders), but have the IGF-II coding region and 3′-UTR in common (Fig. 1A). The length and composition of the leaders varies significantly; leader L1 consists of exons 1–3 and is 586 nt long and very GC rich; leader L2 (408 nt) consists of exon 4 sequences and is moderately rich in G and C residues; leader L3 (1171 nt) consists of exon 5 sequences, is rich in C residues and contains the longest 5′-UTR; leader L4 consists of exon 6 sequences and is the shortest 5′-UTR with only 109 nt. Computer-assisted RNA secondary structure prediction suggests that leaders L1 and L3 contain stable secondary structures while leader L2 is less structured and leader L4 does not contain any stable secondary structure (not shown). Previously, it was established that transcripts with leaders 1, 2 and 4 are efficiently translated, whereas leader 3 has a strong repressive effect on translation (32). Thus, it appears that the leaders significantly affect translation efficiency of the various IGF-II messengers.
Figure 1.

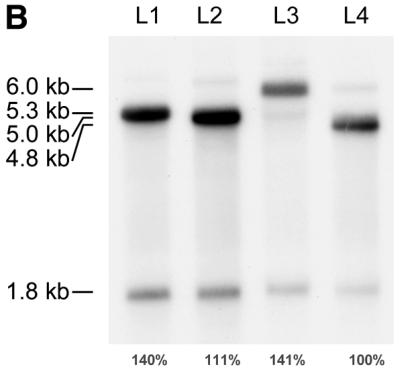
Effect of the various IGF-II 5′ leaders on mRNA cleavage efficiency. (A) Structure of the human IGF-II gene and the mRNAs generated by transcription from promoters P1–P4; the transcripts differ among each other in their 5′ leader sequences, but have their coding region and 3′-UTR in common. Promoters P1–P4 are indicated by bent arrows. Exons 1–9 are indicated by boxes; open boxes represent untranslated sequences, filled boxes represent the IGF-II coding region. The sizes of the transcripts with leaders L1–L4 are indicated in kb on the right. (B) Northern blot analysis of poly(A)+ RNA isolated from 293 cells transiently transfected with L1–L4 IGF-II expression plasmids shown in (A). The blot was hybridised with an IGF-II 3′-specific probe, detecting both the full-length transcripts and 3′ cleavage product. For each leader construct the amount of IGF-II RNA was quantified by phosphorimager scanning. The cleavage efficiency of the different leader transcripts was calculated and the mean value of three independent experiments is indicated below each lane. Variation between individual experiments was <10%.
In order to test the possibility that the nature of the individual leaders also affects cleavage, we set out to determine the cleavage efficiency of IGF-II mRNAs carrying the different leaders. For this purpose, constructs were generated in which the four different leaders (L1–L4) were linked to the IGF-II open reading frame and the 4 kb long 3′-UTR (Fig. 1A). The IGF-II leader constructs were transiently transfected to human 293 cells that are able to perform the cleavage reaction of these IGF-II minigene constructs but that do not express IGF-II endogenously. Transfection efficiency was monitored with a cotransfected RSV-luc construct. Twenty-four hours after transfection, poly(A)+ RNA was isolated from the cells and subjected to northern blot analysis using an IGF-II probe specific for detection of full-length IGF-II mRNAs and the 3′ cleavage product (Fig. 1B).
All four IGF-II transcripts were cleaved, as the 1.8 kb 3′ cleavage product was observed with all IGF-II mRNAs. The cleavage efficiency for each leader construct can be determined by the ratio of the cleavage product to the full-length product as previously described (23) using the IGF-II RNA signals quantified by phosphorimager scanning. For each leader construct the signal of the 3′ cleavage product was divided by the sum of the signals of the full-length RNA and the 3′ cleavage product (26), resulting in distinct differences in cleavage efficiency. In order to compare the cleavage efficiencies of the four different leader constructs directly, the cleavage efficiency of the least structured 5′-UTR, L4, was set at 100%. The L1 and L3 RNAs with predicted stable secondary structures were cleaved more efficiently than L4, with 140 and 141%, respectively, relative to L4 RNA; L2 was cleaved with lower efficiency than L1 and L3, but still more than L4, with 111%. Thus, although the 5′-UTR regions do not have all-or-none effects on cleavage, they do affect the efficiency of cleavage to a certain extent. These results suggest a positive correlation between complex structure formation in the 5′-UTR and cleavage, as the highly structured leaders L1 and L3 were cleaved more efficiently than the unstructured leader L4.
Effect of cleavage on the rate of mRNA decay and protein production
Subsequently, we investigated the consequence of cleavage on overall mRNA degradation. Initial studies were performed in human Hep3B cells which express IGF-II endogenously and in murine Ltk– cells which were stably transfected with IGF-II minigene expression plasmids. Actinomycin D was used to inhibit transcription and the IGF-II RNA levels were measured at various time points after administration of this drug. No significant changes in the IGF-II RNA levels were observed within the first 24 h after transcription arrest, whereas the unstable c-myc transcripts quickly vanished under the same conditions, indicating that transcription inhibition was complete (data not shown). These results suggested that the IGF-II transcripts are very stable, although it is generally known that actinomycin D can cause artificial stabilisation of some RNAs. Since actinomycin D non-specifically inhibits transcription of all genes, especially after longer time points, non-physiological conditions may result from extended actinomycin D incubations. Therefore, we chose to use a tetracycline-inducible system, in which the expression of the IGF-II gene was induced or repressed with minimal interference with the cellular physiology.
Two IGF-II expression plasmids were constructed with a promoter that can be repressed by tetracycline (Fig. 2A). In plasmid pTet-L4-ΔWT, the 5′-UTR of the 4.8 kb IGF-II mRNA (leader L4) is fused to the IGF-II open reading frame and the 3′-UTR of IGF-II mRNA. Leader L4 was selected because it is the shortest IGF-II leader of only 109 nt and it has no predicted secondary structure in the leader sequence. Therefore, it may be predicted that the influence of L4 on IGF-II mRNA cleavage in the 3′-UTR is minimal. The 3′-UTR lacks the region from position 876 to 2657 in the transcript that was designated ΔWT (23) because this region was previously shown to be dispensable for cleavage (26). To generate plasmid pTet-L4-ΔWTmut, point mutations were introduced around the cleavage site that completely abolish cleavage (Fig. 2A) (23). The plasmids were used for transient transfection assays in human 293 cells, which are a suitable model system to study in vivo cleavage, because these cells do not express IGF-II endogenously, but are able to perform the cleavage reaction.
Figure 2.



(A) Schematic representation of the transcripts from plasmids pTet-L4-ΔWT and pTet-L4-ΔWTmut. The nucleotide positions bordering the 5′-UTR (leader L4), the IGF-II open reading frame (ORF) and the 3′-UTR are indicated. The region in the 3′-UTR of ΔWT that is deleted and that was shown to be dispensable for cleavage (23) is indicated by an arrow. The cleavage site (CS) is also marked by an arrow. The asterisk indicates the positions of the point mutations in pTet-L4-ΔWTmut. (B) Northern blot analysis of total RNA isolated from human 293 cells transiently transfected with pTet-L4-ΔWT or pTet-L4-ΔWTmut in the absence of tetracycline. As a control for complete transcription arrest cells were grown in the presence of 500 ng/ml tetracycline. At 24 h after transfection, tetracycline was added to a concentration of 500 ng/ml to repress expression of the constructs and cells were harvested at the indicated time points. The blot was hybridised with an IGF-II 3′-UTR-specific probe detecting full-length RNAs as well as the 3′ cleavage product. A GAPDH coding region probe was used to correct for loading differences. (C) Graphic representation of IGF-II RNA decay of full-length L4-ΔWT RNA and ΔWTmut RNA levels (diamond) and of the formation of the 3′ cleavage product (triangle) after transcription arrest.
First, we determined the amount of tetracycline required for complete repression of transcription in 293 cells. We found that 50 ng/ml of tetracycline was sufficient to reduce the level of IGF-II mRNA to undetectable levels and a concentration of tetracycline up to 500 ng/ml did not affect growth rate or cell survival (data not shown). To test the effect of cleavage on the rate of IGF-II mRNA turnover directly 293 cells were transiently transfected with plasmid pTet-L4-ΔWT or pTet-L4-ΔWTmut in the absence of tetracycline to allow expression of the constructs. At 24 h after transfection, tetracycline was added to a concentration of 500 ng/ml to ensure complete and rapid inhibition of transcription. Subsequently, RNA was isolated at various time points and subjected to northern blot analysis using the IGF-II 3′-UTR-specific probe and the GAPDH probe (Fig. 2B). Control transfections carried out in the presence of 500 ng/ml tetracycline did not yield any expression of the constructs, showing that the inhibition of transcription by tetracycline was complete. For both constructs, no significant decrease in the full-length IGF-II RNA levels was observed up to 6 h after transcription inhibition, indicating that the RNAs are rather stable. After 8 and 10 h, however, the RNA levels were decreased to ∼85% of the initial levels at time point 0 and after 28 h a reduction to 33% was found. Similar decay kinetics were observed for both the L4-ΔWT and the L4-ΔWTmut RNAs, irrespective of the presence or absence of cleavage, indicating that cleavage did not accelerate the decay rate of the L4-ΔWT RNA (Fig. 2C).
While the L4-ΔWT full-length RNA levels declined, the abundance of the 3′ cleavage product did not decrease throughout the course of the experiment (Fig. 2B and C). Although the formation of the 3′ cleavage product continued due to ongoing cleavage during the experiment, this occurred at a diminished rate because the substrate full-length RNA levels declined. This result confirms previous observations that the 3′ cleavage product is a very stable RNA species. Notably, the level of the 3′ cleavage product at time point 0 was about 4-fold lower than that at 6 h after transcription inhibition. This suggests that at time point 0 (24 h after transfection), the levels of the 3′ cleavage product had not yet reached a steady-state level, reflecting the slow rate of cleavage. Thus, both the formation and the degradation of the 1.8 kb 3′ cleavage product appear to be very slow.
In order to examine whether protein synthesis is affected by cleavage of the IGF-II mRNAs, reporter plasmids pTet-L4-luc-ΔWT and pTet-L4-luc-ΔWTmut were constructed. In these plasmids, the IGF-II open reading frame was replaced by the luciferase open reading frame. This was necessary, because it is impossible to measure IGF-II production by the cells due to the large amounts of IGF-II present in FCS in the medium. The 293 cells were transiently transfected with these plasmids; suboptimal amounts of DNA were used for transfection to ensure that the expression levels of the constructs were in the linear range. After transfection, RNA was isolated from the cells and analysed on northern blots. The transcript derived from pTet-L4-luc-ΔWT was cleaved, indicating that cleavage can still occur when the IGF-II open reading frame is replaced by the luciferase open reading frame (Fig. 3A). This is in agreement with our previous results showing that a region from the IGF-II 3′-UTR containing sequence elements I and II can confer cleavage of a heterologous mRNA (26). In contrast, the transcript from pTet-L4-luc-ΔWTmut was not cleaved at all, confirming our previous observation that the introduced mutations completely abolish cleavage (Fig. 3A) (23). At 30 h after transfection the cells were tested for their luciferase activity. No difference was observed in luciferase activity of the cells transfected with either pTet-L4-ΔWT or pTet-L4-ΔWTmut, indicating that cleavage did not affect luciferase expression (Fig. 3B).
Figure 3.
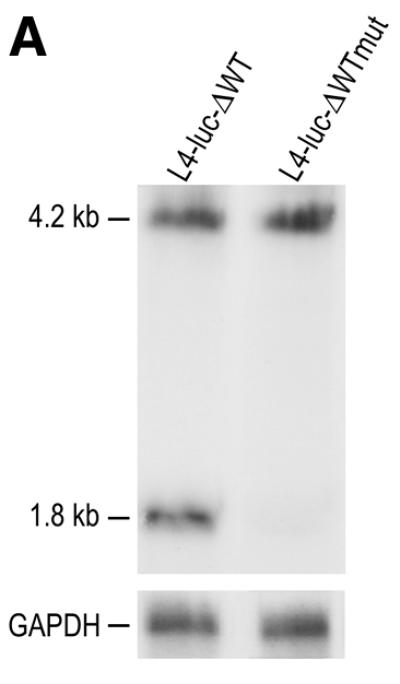

Effect of cleavage on luciferase expression. (A) Northern blot analysis of RNA isolated from 293 cells which were transiently transfected with pTet-L4-luc-ΔWT or pTet-L4-luc-ΔWTmut. The blot was hybridised with an IGF-II 3′-UTR-specific probe detecting full-length RNAs as well as the 3′ cleavage product. The sizes of the full-length RNAs and the 3′ cleavage product are indicated in kb on the left. As a control for loading differences, the blot was also probed with a GAPDH DNA fragment. (B) The luciferase activity of cells transiently transfected with pTet-L4-luc-ΔWTmut relative to the activity of cells transfected with pTet-L4-luc-ΔWT, which was set at 100%. The result shown represents the mean value of five independent experiments and the error bar shows the calculated standard deviation.
In conclusion, the results indicate that under the standard cell culture conditions used in our transient transfection assays, IGF-II mRNA cleavage is a slow process that has little effect on the mRNA decay rate and protein production.
Endonucleolytic cleavage of IGF-II mRNAs is up-regulated at high cell density
It has been described that IGF-II gene expression in several systems is affected by cell density and cell differentiation. For this reason, we used a time course to examine the growth status of two human cell lines known to express IGF-II endogenously: (i) the human hepatocarcinoma cell line Hep3B that is capable of growing to high cell densities forming multiple cell layers; and (ii) the human colonic carcinoma cell line CaCo2 that spontaneously differentiates at high cell densities (33,34).
First, we examined the endogenous IGF-II expression pattern of both cell lines at two cell densities using specific probes for detection of full-length IGF-II mRNAs and 5′ or 3′ cleavage product. In the human neuroblastoma cell line SHSY-5Y, it was originally observed that cleavage of IGF-II mRNA results in the formation of an abundant 1.8 kb 3′ cleavage product and much lower levels of the 4.2 kb 5′ cleavage product, indicating that the 5′ cleavage product is less stable than the 3′ cleavage product (7). RNA isolated from exponentially growing Hep3B cells on serum replacement medium and from CaCo2 cells growing in 10% FCS-containing medium, was subjected to northern blot analysis. An IGF-II 3′-UTR-specific probe was used to detect full-length IGF-II mRNAs and the 3′ cleavage product and an IGF-II coding region-specific probe was used to detect full-length IGF-II mRNAs and the 5′ cleavage product (Fig. 4). RNA from Hep3B cells grown to 50% cell density showed the full-length 6.0 and 4.8 kb mRNAs with both probes, the 1.8 kb 3′ cleavage product with the 3′-specific probe and very little of the 4.2 kb 5′ cleavage product with the coding region probe. This indicates that in Hep3B cells the 5′ cleavage product is much less stable than the 3′ cleavage product. At 100% cell density, the full-length IGF-II mRNA levels were decreased, while the levels of both the 5′ and the 3′ cleavage products increased significantly (Fig. 4A). This indicates that the cleavage efficiency was increased at high cell density in Hep3B cells. A similar experiment was done with the CaCo2 cells (Fig. 4B). Hybridisation with the IGF-II coding region-specific probe revealed that both the 6.0 and the 4.8 kb full-length IGF-II mRNAs were expressed in these cells. Again the amount of 1.8 kb 3′ cleavage product was increased at 100% cell density due to increased endonucleolytic cleavage of the full-length IGF-II mRNAs. In these cells no 5′ cleavage product could be detected with the coding region probe, not even at high cell density, suggesting that the 5′ cleavage product is even less stable in CaCo2 cells than in Hep3B cells.
Figure 4.

Northern blot analyses of RNAs isolated from Hep3B cells (A) and CaCo2 cells (B) at 50 and 100% confluence. The blots were first probed with an IGF-II 3′-UTR-specific probe to detect full-length IGF-II mRNAs and the 3′ cleavage product (3′-UTR). After stripping the blots were reprobed with an IGF-II coding region-specific probe to detect full-length IGF-II mRNAs and the 5′ cleavage product (CR). The sizes of the full-length IGF-II mRNAs, 6.0 and 4.8 kb, as well as the size of the 5′ cleavage product (4.2 kb) or the 3′ cleavage product (1.8 kb) are indicated.
To further examine the cell density-dependent regulation of IGF-II expression and cleavage, a time course experiment was performed. Cells were seeded at a density of 1 × 104 cells/cm2 (day 0) and were grown in serum replacement medium (Hep3B) or in serum-containing medium (CaCo2). Medium was replaced daily and cells were counted to determine the cell density at the time they were harvested for RNA isolation and subjected to northern blot analysis using the IGF-II 3′-UTR-specific probe. From day 1 to day 7–8, the cells proliferated rapidly until 100% confluence was reached at a density of 15 × 104 cells/cm2 for the Hep3B cells and 26 × 104 cells/cm2 for the CaCo2 cells (Fig. 5A). A slight decrease in Hep3B cell number was observed on the subsequent day after growth arrest due to cell death.
Figure 5.


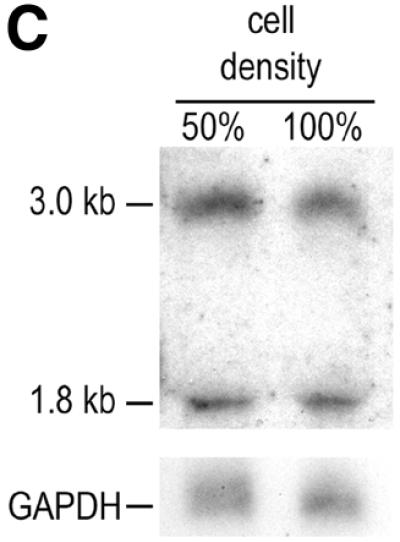
IGF-II gene expression and mRNA cleavage efficiency as a function of Hep3B and CaCo2 cell density. (A) Growth curves of Hep3B cells growing in the TCH‘ serum replacement medium and CaCo2 cells growing in 10% FCS. Cells were seeded at 1 × 104 cells/cm2 (day 0). Medium was refreshed daily. (B) Northern blot analyses of RNAs isolated from the Hep3B and CaCo2 cells harvested at days 3–9 after seeding and grown to the densities indicated in (A). The blots were first hybridised with the IGF-II 3′-UTR-specific probe and subsequently re-probed with the GAPDH probe and a 5′ end-labelled oligonucleotide complementary to the 28S rRNA to correct for loading differences. The RNA signals were quantified by phosphorimager scanning. (C) Northern blot analysis of RNA isolated from murine Ltk– cells stably transfected with the human IGF-II minigene ΔWT (23). The cells express a full-length IGF-II mRNA of 3.0 kb, which after cleavage yield the 1.8 kb 3′-UTR cleavage product. The blot was probed with the IGF-II 3′-UTR-specific probe detecting both the full-length IGF-II mRNA and the 3′ cleavage product. The cells were grown to 50 and 100% confluence as indicated above the lanes.
The result shows that for the Hep3B cells, a slight increase in the full-length IGF-II mRNA levels of 1.7-fold was seen during exponential growth of the cells. When cells reached growth arrest (day 9), the full-length IGF-II mRNA levels decreased. The levels of the 3′ cleavage product increased along with the full-length mRNAs during exponential growth of the cells. Interestingly, at higher cell densities the increase of the abundance of the 3′ cleavage product was stronger than the increase of the full-length mRNA levels; a 3-fold increase was observed at day 5, after which the ratio 1.8/6.0 kb remained high. As the Hep3B cells reached confluence, the amount of 6.0 kb mRNA decreased whereas more 1.8 kb mRNA could be detected, indicating that the cleavage efficiency increased upon reaching confluence (Fig. 5B).
A similar, but not identical, pattern was observed with the CaCo2 cells. The full-length IGF-II mRNA levels increased 3-fold during exponential growth and reached a peak level at day 7 after seeding (Fig. 4B). At day 9, the full-length IGF-II mRNA levels decreased to the initial levels again, while the level of the 3′ cleavage product was further increased to 5-fold of the level seen on day 3. These results indicate that in Hep3B cells and to a greater extent in CaCo2 cells, the cleavage efficiency significantly increases at high cell density. Thus, in addition to the previously observed general decrease in transcriptional activity (35), endonucleolytic cleavage of the full-length IGF-II mRNAs significantly contributes to the observed down-regulation of the full-length IGF-II mRNA levels.
Having established that Hep3B cells and CaCo2 cells show increased cleavage efficiency at high cell density, we determined whether this phenomenon was specific for cells endogenously expressing IGF-II or if regulated cleavage could occur in other cell types as well. Murine fibroblast Ltk– cells, which do not produce endogenous IGF-II, were stably transfected with the human IGF-II minigene construct ΔWT and it was shown that these cells express and cleave the transcript from the introduced IGF-II minigene (23,26). The stably transfected Ltk– cells were grown to 50 and 100% confluence and subjected to northern blot analysis of total RNA. Using the IGF-II 3′-UTR-specific probe, it was shown that the IGF-II RNA levels were not changed in these cells at higher densities; both the levels of the full-length IGF-II mRNA as well as the levels of the 3′ cleavage product remained constant (Fig. 5C). This indicates that the increase in cleavage efficiency under high cell density conditions is not a general phenomenon that occurs in every cell type that is able to perform IGF-II mRNA cleavage. Similar results were obtained in transient assays with human 293 cells, also not endogenously expressing IGF-II (data not shown).
In conclusion, our data indicate that regulated endonucleolytic cleavage is specific for a limited set of cell types such as Hep3B and CaCo2 cells which express IGF-II endogenously. This phenomenon could be restricted to cells for which altered IGF-II expression is related to specific cell processes such as growth arrest or cell differentiation.
DISCUSSION
In this study we have investigated the effects on cleavage efficiency of the different 5′ leader sequences present in the various IGF-II transcripts in transient transfection assays. Subsequently, the kinetics of IGF-II mRNA cleavage and its effect on mRNA decay and protein production was studied. In addition, we have examined the regulation of IGF-II mRNA cleavage in human Hep3B cells and CaCo2 cells, which endogenously express IGF-II.
The human IGF-II gene contains four promoters, which are differentially active during development and in different tissues. The 5′ leaders L2 and L4 do not show significant secondary structure, whereas L1 and L3 are rather long and highly structured. It was established previously that the leaders have significant effects on translation efficiency; while IGF-II mRNAs carrying L1, L2 and L4 are efficiently translated, L3 strongly represses translation (32). In this report we demonstrate that the leaders also differentially influence cleavage efficiency of the IGF-II mRNAs. mRNAs containing leaders L1 and L3 that have predicted stable secondary structures were cleaved more efficiently than RNAs with the non-structured leaders L2 and L4, suggesting a positive correlation between secondary structure formation in the leader and cleavage efficiency of the corresponding mRNA. Thus, the leaders appear to affect both translation and cleavage, but in a different manner. Only the translational activity of the L3-transcript is strongly inhibited, while the cleavage efficiency of both L1 and L3 are enhanced. This suggests that there is no direct link between translation and cleavage, in contrast to several other systems; cleavage of the 9E3, the c-myc and the groα transcripts have been described to be dependent on translation (14,36,37). Similar to IGF-II mRNA cleavage, however, endonucleolytic cleavage of the Xenopus homeobox mRNA Xlhbox2 was also found to be independent of translation (17). A separate regulation for endonucleolytic cleavage of IGF-II mRNAs allows for a response to exogenous stimuli independently of translation regulation.
Another important issue to be addressed is the effect of cleavage on IGF-II mRNA turnover. Using a tetracycline-inducible system the decay of IGF-II mRNAs was examined. We have made the following observations. (i) Point mutations that inactivate cleavage do not lead to a slower degradation rate of the RNAs after transcription inhibition; both RNAs that are cleaved and RNAs that are not cleaved were reduced to 33% of the initial levels after 28 h. This indicates that endonucleolytic cleavage did not contribute significantly to the general decay of the IGF-II mRNAs in the 293 cells used. In contrast to the full-length RNAs, no decline in the levels of the 3′ cleavage product was seen throughout the experiment indicating that the 1.8 kb RNA species is very stable. (ii) No significant difference in luciferase expression was observed between cleaved and non-cleaved RNA, indicating that cleavage had little effect on the protein production.
From these data, we conclude that under the standard cell culture conditions used in our transient transfection assays, cleavage is a slow process that does not play a significant role in overall IGF-II mRNA degradation or IGF-II protein production. Cleavage leads to the production of a very stable 1.8 kb 3′ cleavage product, which remains present in the cells for a long time after its formation.
Several studies have shown that cell density affects IGF-II expression in different cell types. Both in the rat IEC-6 and the rat BRL-3A cell lines an up-regulation of IGF-II gene expression has been observed at increasing cell densities, leading to the hypothesis that IGF-II may act as a survival factor at high cell densities in these cells (38,39). These reports prompted us to investigate whether IGF-II gene expression and IGF-II mRNA cleavage are also regulated in a cell density-dependent manner. Hep3B cells and CaCo2 cells, which express IGF-II endogenously, were grown to different densities and analysed for IGF-II expression and cleavage. During exponential growth of the cells the abundance of the IGF-II full-length mRNAs and the 3′ cleavage product are enhanced due to increased transcription (Fig. 5). However, when cells reach confluence a consistent increase of 3–5-fold in the abundance of only the 3′ cleavage product was observed, indicating that cleavage of full-length IGF-II mRNAs is upregulated (Fig. 5). Interestingly, this regulated response in cleavage occurs at high cell density only, when Hep3B cells stop growing and go into cell-arrest and when CaCo2 cells start to differentiate. This suggests that under these altered growth conditions the IGF-II synthesis is inhibited not only by down-regulation of transcription, but more importantly by enhanced cleavage of the full-length IGF-II mRNAs. These two concerted actions will lead to a rapid decrease of IGF-II synthesis in the cells. We proposed that this regulated cleavage occurs only in cells that endogenously express IGF-II. This was confirmed when non-IGF-II expressing murine Ltk– cells were stably transfected with the human IGF-II minigene ΔWT. Upon expression of IGF-II from this minigene no cell density-dependent increase in cleavage efficiency was observed (Fig. 5C). This suggests that the regulated cleavage is a cell type-specific phenomenon, rather than a general effect of growth inhibition.
What might be the physiological relevance of the regulated cleavage efficiency? The increased levels of the 3′ cleavage product were accompanied by a reduction in the full-length IGF-II mRNA levels, indicating that the increase in cleavage efficiency may contribute to a down-regulation of IGF-II expression at high cell density. Thus, although cleavage is probably not exclusively responsible for the reduced IGF-II mRNA levels at high cell densities, it accelerates the disappearance of the very stable IGF-II mRNAs. An additional interesting possibility is that the 3′ cleavage product may have an intrinsic cellular function under altered growth conditions. Similarly, it was demonstrated that 3′-UTR sequences of the mRNAs from murine muscle structural genes intrinsically inhibit cell division and promote cell differentiation (40). The unusual stability of the 1.8 kb IGF-II 3′ cleavage product caused by a protective 5′ G-quadruplex structure provides support for this hypothesis, especially because stable non-coding RNAs have been shown to carry out diverse functions in both prokaryotes and eukaryotes (reviewed in 41–43). In addition, computer-assisted secondary structure prediction suggests that the 1.8 kb 3′ cleavage product is highly base-paired and, interestingly, this RNA appears to be rich in structural motifs that are common to functional non-coding RNAs as determined with a specialised computer algorithm available on the World Wide Web at http://rnagene.lbl.gov/.
How does cleavage of the IGF-II mRNAs relate to other systems? Endonucleolytic cleavage has been described for a number of mRNAs. Cleavage of the IGF-II mRNAs is distinct from these systems in a number of respects. For example, the determinants for recognition of the cleavage site are very complex in the case of IGF-II mRNA cleavage, whereas for cleavage of the apoII, the albumin and the Xenopus Xlhbox2B mRNAs, short sequences in a single-stranded environment appear to be sufficient (13,15,18). IGF-II mRNAs cleavage requires extensive RNA structures that act in a co-operative manner to stabilise a highly specific cleavage site (23,24). To our knowledge, the transferrin receptor mRNA is the only other system that has been described to require large regions of RNA structure for specific cleavage in its 3′-UTR. Interestingly, an upstream sequence element was found to be required for cleavage of this mRNA as well (16), similar to the requirement for the upstream element I in the IGF-II mRNAs. Cleavage of the IGF-II mRNAs occurs independently of deadenylation, similar to most, but not all systems; it was shown that cleavage of groα mRNA is associated with poly(A) tail shortening (14) and also cleavage of the yeast PGK1 mRNA and the human α-globin mRNA have been reported to be coupled with poly(A) tail shortening (21,44). In addition to its complex structural requirements and its independence of deadenylation, also its independence of translation distinguishes IGF-II mRNA cleavage from a number of other systems, as mentioned above. This makes the regulated IGF-II mRNA endonucleolytic cleavage a novel example of functional RNA processing, involving both structural RNA determinants as well as specific physiological conditions of the cells.
In summary, we have established that cleavage is regulated in a cell density-dependent manner and that this regulation is cell type-specific. The unusually stable 1.8 kb 3′ cleavage product shows interesting structural features that are common to functional RNAs. This makes it tempting to speculate that the 1.8 kb 3′ cleavage product of IGF-II mRNA might have an intrinsic physiological function.
Acknowledgments
ACKNOWLEDGEMENTS
The authors would like to thank Mrs A. M. C. B. Koonen-Reemst for excellent technical assistance. This work was supported by a grant from the Netherlands Organization for the Advancement of Pure Research (NWO).
References
- 1.Jones J.I. and Clemmons,D.R. (1995) Insulin-like growth factors and their binding proteins: biological actions. Endocr. Rev., 16, 3–34. [DOI] [PubMed] [Google Scholar]
- 2.Engström W., Shokrai,A., Otte,K., Granerus,M., Gessbo,A., Bierke,P., Madej,A., Sjolund,M. and Ward,A. (1998) Transcriptional regulation and biological significance of the insulin like growth factor II gene. Cell Prolif., 31, 173–189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Holthuizen P.E., Steenbergh,P.H. and Sussenbach,J.S. (1999) Regulation of IGF gene expression. In Rosenfeld,R.G. and Roberts,C.T. (eds), The IGF System. Molecular Biology, Physiology and Clinical Applications. Humana Press, Totowa, NJ, pp. 37–61.
- 4.De Pagter-Holthuizen P., Jansen,M., van der Kammen,R.A., van Schaik,F.M.A. and Sussenbach,J.S. (1988) Differential expression of the human insulin-like growth factor II gene. Characterization of the IGF-II mRNAs and an mRNA encoding a putative IGF-II-associated protein. Biochim. Biophys. Acta, 950, 282–295. [DOI] [PubMed] [Google Scholar]
- 5.Meinsma D., Holthuizen,P., van den Brande,J.L. and Sussenbach,J.S. (1991) Specific endonucleolytic cleavage of IGF-II mRNAs. Biochem. Biophys. Res. Commun., 179, 1509–1516. [DOI] [PubMed] [Google Scholar]
- 6.Nielsen F.C. and Christiansen,J. (1992) Endonucleolysis in the turnover of insulin-like growth factor II mRNA. J. Biol. Chem., 267, 19404–19411. [PubMed] [Google Scholar]
- 7.Holthuizen P.E., Cleutjens,C.B.J.M., Veenstra,G.J.C., van der Lee,F.M., Koonen-Reemst,A.M.C.B. and Sussenbach,J.S. (1993) Differential expression of the human, mouse and rat IGF- II genes. Regul. Pept., 48, 77–89. [DOI] [PubMed] [Google Scholar]
- 8.Beelman C.A. and Parker,R. (1995) Degradation of mRNA in eukaryotes. Cell, 81, 179–183. [DOI] [PubMed] [Google Scholar]
- 9.Ross J. (1995) mRNA stability in mammalian cells. Microbiol. Rev., 59, 423–450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Jacobson A. and Peltz,S.W. (1996) Interrelationships of the pathways of mRNA decay and translation in eukaryotic cells. Annu. Rev. Biochem., 65, 693–739. [DOI] [PubMed] [Google Scholar]
- 11.Wickens M., Anderson,P. and Jackson,R.J. (1997) Life and death in the cytoplasm: messages from the 3′ end. Curr. Opin. Genet. Dev., 7, 220–232. [DOI] [PubMed] [Google Scholar]
- 12.Dompenciel R.E., Garnepudi,V.R. and Schoenberg,D.R. (1995) Purification and characterization of an estrogen-regulated Xenopus liver polysomal nuclease involved in the selective destabilization of albumin mRNA. J. Biol. Chem., 270, 6108–6118. [DOI] [PubMed] [Google Scholar]
- 13.Chernokalskaya E., Dompenciel,R. and Schoenberg,D.R. (1997) Cleavage properties of an estrogen-regulated polysomal ribonuclease involved in the destabilization of albumin mRNA. Nucleic Acids Res., 25, 735–742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Stoeckle M.Y. (1992) Removal of a 3′ non-coding sequence is an initial step in degradation of groαmRNA and is regulated by interleukin-1. Nucleic Acids Res., 20, 1123–1127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Binder R., Hwang,S.P., Ratnasabapathy,R. and Williams,D.L. (1989) Degradation of apolipoprotein II mRNA occurs via endonucleolytic cleavage at 5′-AAU-3′/5′-UAA-3′ elements in single-stranded loop domains of the 3′-noncoding region. J. Biol. Chem., 264, 16910–16918. [PubMed] [Google Scholar]
- 16.Binder R., Horowitz,J.A., Basilionm,J.P., Koeller,D.M., Klausner,R.D. and Harford,J.B. (1994) Evidence that the pathway of transferrin receptor mRNA degradation involves an endonucleolytic cleavage within the 3′ UTR and does not involve poly(A) tail shortening. EMBO J., 13, 1969–1980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Brown B.D. and Harland,R.M. (1990) Endonucleolytic cleavage of a maternal homeo box mRNA in Xenopus oocytes. Genes Dev., 4, 1925–1935. [DOI] [PubMed] [Google Scholar]
- 18.Brown B.D., Zipkin,I.D. and Harland,R.M. (1993) Sequence-specific endonucleolytic cleavage and protection of mRNA in Xenopus and Drosophila. Genes Dev., 7, 1620–1631. [DOI] [PubMed] [Google Scholar]
- 19.Swartwout S.G. and Kinniburgh,A.J. (1989) c-myc RNA degradation in growing and differentiating cells: possible alternate pathways. Mol. Cell. Biol., 9, 288–295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Ioannidis P., Havredaki,M., Courtis,N. and Trangas,T. (1996) In vivo generation of 3′ and 5′ truncated species in the process of c-myc mRNA decay. Nucleic Acids Res., 24, 4969–4977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Wang Z. and Kiledjian,M. (2000) Identification of an erythroid-enriched endoribonuclease activity involved in specific mRNA cleavage. EMBO J., 19, 295–305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Scheper W., Meinsma,D., Holthuizen,P.E. and Sussenbach,J.S. (1995) Long-range RNA interaction of two sequence elements required for endonucleolytic cleavage of human insulin-like growth factor II mRNAs. Mol. Cell. Biol., 15, 235–245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.van Dijk E.L., Sussenbach,J.S. and Holthuizen,P.E. (1998) Identification of RNA sequences and structures involved in site-specific cleavage of IGF-II mRNAs. RNA, 4, 1623–1635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.van Dijk E.L., Sussenbach,J.S. and Holthuizen,P.E. (2000) Distinct RNA structural domains cooperate to maintain a specific cleavage site in the 3′-UTR of IGF-II mRNAs. J. Mol. Biol., 300, 449–467. [DOI] [PubMed] [Google Scholar]
- 25.Sambrook J., Fritsch,E.F. and Maniatis,T. (1989) Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
- 26.Meinsma D., Scheper,W., Holthuizen,P., van den Brande,J.L. and Sussenbach,J.S. (1992) Site-specific cleavage of IGF-II mRNAs requires sequence elements from two distinct regions of the IGF-II gene. Nucleic Acids Res., 20, 5003–5009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Rusconi S., Severne,Y., Georgiev,O., Galli,I. and Wieland,S. (1990) A novel expression assay to study transcriptional activators. Gene, 89, 211–221. [DOI] [PubMed] [Google Scholar]
- 28.de Moor C.H. (1994) Translation of Insulin-like Growth Factor 2 mRNAs. Utrecht University, Utrecht, The Netherlands.
- 29.Graham F.L. and van der Eb,A.J. (1973) A new technique for the assay of infectivity of human adenovirus 5 DNA. Virology, 52, 456–467. [DOI] [PubMed] [Google Scholar]
- 30.de Wet J.R., Wood,K.V., DeLuca,M., Helinski,D.R. and Subramani,S. (1987) Firefly luciferase gene: structure and expression in mammalian cells. Mol. Cell. Biol. 7, 725–737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Jansen M., van Schaik,F.M.A., van Tol,H., van den Brande,J.L. and Sussenbach,J.S. (1985) Nt sequence analysis of cDNAs encoding precursors of human insulin-like growth factor II (IGF-II) and an IGF-II variant. FEBS Lett., 179, 243–246. [DOI] [PubMed] [Google Scholar]
- 32.de Moor C.H., Jansen,M., Sussenbach,J.S. and van den Brande,J.L. (1994) Differential polysomal localization of human insulin-like growth factor 2 mRNAs, both in cell lines and fetal liver. Eur. J. Biochem., 222, 1017–1022. [DOI] [PubMed] [Google Scholar]
- 33.Peterson M.D. and Mooseker,M.S. (1992) Characterization of the enterocyte-like brush border cytoskeleton of the C2BBe clones of the human intestinal cell line, Caco-2. J. Cell Sci., 102, 581–600. [DOI] [PubMed] [Google Scholar]
- 34.Singh P., Dai,B., Yallampalli,U., Lu,X. and Schroy,P.C. (1996) Proliferation and differentiation of a human colon cancer cell line (Caco2) is associated with significant changes in the expression and secretion of insulin-like growth factor (IGF) IGF-II and IGF binding protein-4: role of IGF-II. Endocrinology, 137, 1764–1774. [DOI] [PubMed] [Google Scholar]
- 35.Singh P., Dai,B., Given,R.L., Lu,X. and Holthuizen,P.E. (1998) Differential activation of IGF-II promoters P3 and P4 in Caco-2 cells during growth and differentiation. Gastroenterology, 114, 1221–1229. [DOI] [PubMed] [Google Scholar]
- 36.Stoeckle M.Y. and Hanafusa,H. (1989) Processing of 9E3 mRNA and regulation of its stability in normal and Rous sarcoma virus-transformed cells. Mol. Cell. Biol., 9, 4738–4745. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Wisdom R. and Lee,W. (1990) Translation of c-myc mRNA is required for its post-transcriptional regulation during myogenesis. J. Biol. Chem., 265, 19015–19021. [PubMed] [Google Scholar]
- 38.Corkins M.R., Park,J.H., Davis,D.V., Slentz,D.H. and MacDonald,RG. (1999) Regulation of the insulin-like growth factor axis by increasing cell number in intestinal epithelial (IEC-6) cells. Growth Horm. IGF Res., 9, 414–424. [DOI] [PubMed] [Google Scholar]
- 39.Kutoh E., Schwander,J. and Margot,J.B. (1995) Cell-density-dependent modulation of the rat insulin-like-growth-factor-binding protein 2 and its gene. Eur. J. Biochem., 234, 557–562. [DOI] [PubMed] [Google Scholar]
- 40.Rastinejad F. and Blau,H.M. (1993) Genetic complementation reveals a novel regulatory role for 3′ untranslated regions in growth and differentiation. Cell, 72, 903–917. [DOI] [PubMed] [Google Scholar]
- 41.Wagner E.G. and Simons,R.W. (1994) Antisense RNA control in bacteria, phages and plasmids. Annu. Rev. Microbiol. 48, 713–742. [DOI] [PubMed] [Google Scholar]
- 42.Delihas N. (1995) Regulation of gene expression by trans-encoded antisense RNAs. Mol. Microbiol., 15, 411–414. [DOI] [PubMed] [Google Scholar]
- 43.Kelley R.L. and Kuroda,M.I. (2000) Noncoding RNA genes in dosage compensation and imprinting. Cell, 103, 9–12. [DOI] [PubMed] [Google Scholar]
- 44.Vreken P. and Raue,H.A. (1992) The rate-limiting step in yeast PGK1 mRNA degradation is an endonucleolytic cleavage in the 3′-terminal part of the coding region. Mol. Cell. Biol., 12, 2986–2996. [DOI] [PMC free article] [PubMed] [Google Scholar]