


Abstract
Rpm2p is a protein subunit of yeast mitochondrial RNase P and is also required for the maturation of Rpm1r, the mitochondrially-encoded RNA subunit of the enzyme. Previous work demonstrated that an insertional disruption of RPM2, which produces the C-terminally truncated protein Rpm2-ΔCp, supports growth on glucose but cells lose some or all of their mitochondrial genome and become petite. These petites, even if they retain the RPM1 locus, lose their ability to process the 5′-ends of mitochondrial tRNA. We report here that if strains containing the truncated RPM2 allele are created and maintained on respiratory carbon sources they have wild-type mitochondrial genomes, and a significant portion of tRNA transcripts are processed. In contrast, precursor Rpm1r transcripts accumulate and mature Rpm1r is not made. These data show that one function of the deleted C-terminal region is in the maturation of Rpm1r, and that this region and mature Rpm1r are not absolutely required for RNase P activity. Finally, we demonstrate that full activity can be restored if the N-terminal and C-terminal domains of Rpm2p are supplied in trans.
INTRODUCTION
Mitochondrial transfer RNAs (tRNAs) are synthesized as precursors that undergo a number of essential processing steps before assuming a functional role in mitochondrial protein synthesis (1). Ribonuclease P (RNase P) is the endoribonuclease that generates the mature 5′-ends of tRNA by removal of the 5′-leaders from precursor-tRNAs in all organisms (reviewed in 2). All RNase P enzymes described to date, with the exception of plant chloroplast RNase P (3,4), are established ribonucleoproteins, composed of at least one RNA and one protein (5,6). In Eubacteria and a subset of Archea, the RNA component of RNase P is catalytically active in vitro in the absence of proteins (7,8). In contrast, the RNase P RNAs of the Eukarya are not catalytically active by themselves, and RNase P activity requires both RNA and protein subunits.
Yeast mitochondrial RNase P is essential for the production of mitochondrially-encoded tRNAs and, thus, is essential for growth on media containing non-fermentable carbon sources. These tRNAs translate a small number of proteins coded by mitochondrial DNA (mtDNA) that are necessary for electron transport and oxidative phosphorylation. Mitochondrial protein synthesis is also required for the maintenance of wild-type mtDNA in yeast (9), although the reason for this requirement is not known. Therefore, if cells cannot support mitochondrial tRNA processing, they should not be able to maintain the wild-type mitochondrial genome.
Subunits of mitochondrial RNase P are encoded in both mitochondrial and nuclear genomes. The RNA subunit of mitochondrial RNase P, Rpm1r, is encoded by mtDNA, whereas the protein subunit, Rpm2p, is encoded in the nuclear genome and imported into mitochondria (10,11). The gene coding for Rpm1r, RPM1, was originally identified by demonstrating that petite deletion mutants could only process tRNAs if they retained mtDNA located between the tRNAfMet and tRNAPro genes (10). Purification of yeast mitochondrial RNase P to near homogeneity revealed a 105 kDa protein named Rpm2p (12). The evidence that Rpm2p is a subunit of yeast mitochondrial RNase P is compelling. Rpm2p antibodies raised to a Trp–Rpm2p fusion protein immunoprecipitate all RNase P activity and all RNase P RNA from mitochondrial extracts (13). Moreover, an insertional disruption of the RPM2 gene causes the accumulation of precursor tRNAs in vivo (13). While it is clear that Rpm2p is a subunit of a mitochondrial RNase P enzyme, the role that it plays in enzyme activity is not yet known.
Further studies revealed that RPM2 is a complex, multi-functional gene. Experiments designed to delete the entire gene revealed, unexpectedly, that RPM2 is essential for growth on all carbon sources (14). Since mitochondrial protein synthesis is required for respiratory but not fermentative growth, Rpm2p must have at least one other function in addition to its role as a subunit of mitochondrial RNase P. We do not yet know why RPM2 is essential but cells containing the insertional disruption producing a protein with a C-terminal deletion of 468 amino acids sustain deletions in their mitochondrial DNA and do not process tRNA even if they retain the RPM1 gene (14). These results were consistent with a model that placed RNase P function in the C-terminal region of Rpm2p and the essential function in the N-terminal portion of the protein.
We now report that the C-terminal deletion allele can support RNase P activity if diploid cells with this allele are sporulated and maintained on non-fermentable carbon sources. Therefore, RNase P activity does not, as previously thought, require the C-terminal region of RPM2. We show here that the C-terminal deletion allele of RPM2 differentially affects processing of pre-tRNAs and Rpm1r. The C-terminal deletion mutant allele, rpm2-734::LEU2, produces Rpm2-ΔCp, a protein that does not support Rpm1r processing, but does support pre-tRNA processing, albeit not as efficiently as does Rpm2p. These data show that the two RNA maturation functions of Rpm2p can be separated. Finally, we demonstrate that all functions of Rpm2p are provided if the N-terminal and C-terminal domains, both with mitochondrial targeting signals, are expressed in trans.
MATERIALS AND METHODS
Strains, media and reagents
Standard yeast manipulations were used (15). Yeast cells were transformed with plasmid DNA using a lithium acetate method (16). Rich media included 1% Bacto-yeast extract, 2% Bacto-peptone and 2% glucose (YPD); in other cases 3% glycerol and 2% ethanol (YPGE), 2% galactose or 2% raffinose were used instead of glucose. Synthetic complete (SC) media lacking appropriate amino acids for plasmid retention contained 0.67% Bacto-nitrogen base and 2% glucose. Solid media for plates included 2% Bacto-agar. SC medium containing 1 g/l of 5-fluoroorotic acid (5-FOA) was used to select ura– yeast segregants. Culture media reagents were from Fisher Scientific (Pittsburgh, PA) or Difco (Detroit, MI). The haploid strain with chromosomal deletion of RPM2 was created by sporulation of the diploid strain W303 (ade2-1/ade2-1, his3-11,15/his3-11,15, leu2-3,112/leu2-3,112, ura3-1/ura3-1, trp1-1/trp1-1, can1-100/can1-100, RPM2/rpm2-144,1170::LEU2) containing a deletion of 3080 bp of the chromosomal RPM2 gene encoding amino acids 144–1170, replaced with the LEU2 gene. Wild-type RPM2 in this haploid strain is provided by the plasmid YEp352/RPM2. The C-terminally truncated RPM2, rpm2-734::LEU2, haploid strain was obtained by inserting the LEU2 gene into RPM2 at the unique HpaI site downstream of the codon for amino acid 734 as described by Morales et al. (12) into the diploid W303 strain. This strain was sporulated and dissected on glycerol/ethanol plates.
Plasmid construction
Standard procedures were used for the preparation and ligation of DNA fragments and recovery of plasmid DNA from Escherichia coli (17). Restriction and modification enzymes were used as recommended by the supplier (New England Biolabs, Beverly, MA). Plasmid DNA was isolated using Qiagen columns, and DNA fragments were isolated from agarose gels using a QIAEX II Extraction Kit (Qiagen, Chatsworth, CA). To construct plasmid pRS314/ rpm2-Δ735–1180 (ΔC), plasmid pRS314/RPM2 was cut with HpaI and PpuMI, filled in with Klenow fragment, gel-purified and religated. To construct pRS314/rpm2-Δ714–1097 (ΔC-20), plasmid pRS314/RPM2 was cut with PstI, gel-purified and religated. To construct pRS313/rpm2-Δ216–734 (ΔN), plasmid pRS314/RPM2 was cut with EcoRV and HpaI, gel-purified and religated.
RNA isolation
Total RNA was isolated by hot phenol extraction (18). Mitochondrial RNA was purified by guanidinium extraction (19) of isolated mitochondria (20).
Northern analysis
Total RNA or mitochondrial RNA (25–30 µg) was separated on a 6% polyacrylamide/8 M urea/Tris–borate/EDTA gel, transferred to a Zeta-Probe membrane, and processed as recommended by the manufacturer (Bio-Rad, Hercules, CA). A riboprobe complementary to the tRNAfMet was transcribed from PstI-digested pMM6 (20). A riboprobe complementary to Rpm1r was transcribed from HindIII-digested pMH777 (20). A riboprobe complementary to the tRNAPro was transcribed from HindIII-digested pT3/T7Pro. All plasmids were transcribed with T3 RNA polymerase in the presence of [α-32P]UTP (800 Ci/mmol; Dupont NEN, Boston, MA) using an Ambion T7/T3 Maxiscript kit as specified by the supplier. Oligonucleotide probes 5′-TTTATCGCTTTACCAC-3′ and 5′-CCATTAGACGATAAGGTC-3′ complementary to tRNAPhe and tRNAGlu, respectively, were radiolabeled with [γ-32P]ATP at the 5′-end as described (17). The signals were detected using a phosphorimager (PhosphorImager SF, Molecular Dynamics, Eugene, OR) or autoradiography.
RNA end mapping
Primer extension for the mapping of 5′-ends and RNA protection analysis for the 3′-end mapping of Rpm1r was performed as described by Stribinskis et al. (21).
Western analysis
Mitochondria were isolated as described by Fox et al. (22) and mitochondrial proteins were separated by 7% Tris–acetate polyacrylamide gel (Invitrogen, Carlsbad, CA), transferred to Immobilon-P membranes (Millipore, Bedford, MA) and treated with antibodies. The anti-Rpm2p antibodies were made against a peptide encoding amino acids 306–323 (QCB Inc., Hopkinton, MA) and were used at 1:200 dilution.
RESULTS
Yeast strains with a truncation of Rpm2p grow slowly on a non-fermentable carbon source
Dang et al. (13) demonstrated that deletion of the C-terminal end of Rpm2p by inserting the LEU2 gene in RPM2 downstream of the codon for amino acid 734 induced loss of wild-type mtDNA on medium containing 2% glucose. To determine the effect of the C-terminal deletion in cells grown on non-fermentable media, RPM2/rpm2-734::LEU2 diploid cells (Fig. 1A) were sporulated, tetrads dissected and spores grown on plates containing glycerol and ethanol as carbon sources. We found that all four spores in each tetrad grew on glycerol/ethanol plates, but two spores formed smaller colonies (Fig. 1B). The slower growing cells carry the rpm2-734::LEU2 allele, whereas cells from larger-sized colonies contained RPM2. This result predicts that the mutant protein is properly expressed, localized to mitochondria and provides enough RNase P activity to support slow growth on glycerol. To test this prediction, mitochondrial extracts were prepared from wild-type and mutant cells grown in glycerol/ethanol medium and analyzed by western blotting. Figure 1C shows that both wild-type and mutant RPM2 alleles produce Rpm2 proteins that are localized to mitochondria. Thus, the deletion introduced into Rpm2p does not affect the stability and localization of the mutant protein.
Figure 1.

The rpm2-734::LEU2 allele supports respiratory growth if spores are grown directly on medium with glycerol/ethanol. (A) A schematic representation of the diploid RPM2/rpm2-734::LEU2. Numbers indicate amino acid residues in Rpm2p. (B) Diploid cells were sporulated and tetrads were dissected on YPGE plates. The small colony size segregated with the LEU2 allele. (C) Mitochondrial proteins were isolated from wild-type and rpm2-734::LEU2 cells, separated by Tris–acetate–polyacrylamide gel electrophoresis, and analyzed by immunoblotting using anti-Rpm2p polyclonal antibodies. The nuclear-encoded mitochondrial protein Mdh1p was used as a loading control. Lane 1, RPM2; lane 2, rpm2-734::LEU2. Protein molecular mass standards are indicated on the right.
One possible explanation for the decreased doubling time is that a percentage of the rpm2-734::LEU2 cells lose their wild-type mitochondrial genome (become petite) and cannot continue to divide on non-fermentable carbon sources. Such cells should grow if plated to glucose medium but not if plated to glycerol/ethanol medium. Alternatively, the entire population of cells might maintain its wild-type mitochondrial DNA but grow slowly. To differentiate between these two possibilities, the frequency of respiratory deficient cells generated during growth of wild-type and rpm2-734::LEU2 strains on glycerol/ethanol media was determined. In one approach, we removed aliquots from liquid cultures grown on glycerol/ethanol media and plated an equal number of cells to either glucose or glycerol/ethanol containing plates. In another, after 24 h of growth on glycerol/ethanol plates, colonies were dispersed by micromanipulation and the ability of single cells to multiply on glycerol/ethanol was monitored. Fewer than 3% of wild-type and mutant cells failed to divide under conditions that required respiration. Thus, we conclude that rpm2-734::LEU2 cells maintain their wild-type mitochondrial DNA and the slow growth phenotype observed on glycerol/ethanol medium cannot be attributed to higher than normal rates of petite formation.
C-terminal deletion of Rpm2p differentially affects processing of tRNA and Rpm1r
The tRNA processing capacity of the ΔC mutant grown in glycerol/ethanol liquid medium was assessed by northern analysis. Figure 2 demonstrates that while mature tRNAs are produced in the mutant, their steady-state levels are lower compared to wild-type and precursor tRNAs are detected. This occurs regardless of whether they have short (tRNAfMet and tRNAPhe, Fig. 2A) or long (tRNAGlu and tRNAPro, Fig. 2B and C, respectively) 5′-leader sequences.
Figure 2.

Processing of mitochondrial tRNAs in wild-type and rpm2-734::LEU2 cells. Mitochondrial RNA was isolated from cells grown in glycerol/ethanol medium (YPGE). A northern blot was performed with probes to (A) tRNAfMet and tRNAPhe (top and bottom, respectively), (B) tRNAGlu and (C) tRNAPro. Wild-type (lane 1) and rpm2-734::LEU2 (lane 2). Pre-tRNA and tRNA mark the positions of precursor and mature tRNA, respectively.
Rpm2p is also required for maturation of the RNA subunit of mitochondrial RNase P, Rpm1r (21). To analyze the effect of the C-terminal deletion mutation on the synthesis of Rpm1r, steady-state levels of mature Rpm1r and pre-Rpm1r were assessed by northern analysis. The majority of RPM1 transcripts were mature in wild-type cells with a small amount of an intermediate in the processing pathway detected (Fig. 3B, lane 1). In contrast, only a small amount of an intermediate and no mature Rpm1r was detected in the mutant and instead, high molecular weight primary transcripts accumulated (Fig. 3B, lane 2). Hybridization of the same filter with a probe complementary to tRNAPro (Fig. 3C) identified the same major high molecular weight primary transcripts as those detected with the Rpm1r probe (Fig. 3B and C, compare lanes 2). There was no comparable transcript detected in wild-type RNA at this exposure (Fig. 3C, lane 1). These data suggest that Rpm1r processing is disrupted at both 5′- and 3′-ends in cells producing Rpm2-ΔCp.
Figure 3.

rpm2-734::LEU2 cells do not make mature Rpm1r. (A) Rpm1r is transcribed either with tRNAfMet and tRNAPro from the FP promoter, or with only tRNAPro from the SP promoter (top). Vertical arrows correspond to processing events and putative processing intermediates are represented below. The dotted line corresponds to 5′-ends, which can be generated either by cleavage at the 3′-end of tRNAfMet or by transcription initiation at SP promoter. Mitochondrial RNA was isolated from cells grown in YPGE. A northern blot was performed with probes to Rpm1r (B) and tRNAPro (C). Lane 1, wild-type; lane 2, rpm2-734::LEU2.
Mature Rpm1r 5′- and 3′-ends are not made in rpm2-734::LEU2 cells
The 5′-ends of transcripts from RPM1 were mapped by primer extension. A labeled oligonucleotide complementary to Rpm1r was annealed to wild-type and mutant mitochondrial RNA and extended with reverse transcriptase. The extension products were separated on a 6% denaturing polyacrylamide gel and compared to a DNA sequence generated with the same oligonucleotide (Fig. 4A). The most abundant termination product in wild-type RNA results from the mature Rpm1r (Fig. 4A, lanes 6 and 7). No corresponding extension product was present in the mutant (Fig. 4A, lane 8). Transcripts corresponding to Rpm1r precursors have the same 5′-ends, previously mapped at the nucleotide level to the SP promoter or to the 3′-end of the tRNAfMet gene (21), in RNA isolated from wild-type and mutant cells.
Figure 4.

Mapping the 5′- and 3′-ends of RPM1 transcripts. (A) 5′-ends were determined by extending a radiolabeled primer complementary to a sequence in Rpm1r. A dideoxy sequencing ladder (lanes 1–4) and extension products from wild-type (lanes 6 and 7, mitochondrial RNA isolated from YPD and YPGE grown cells, respectively), and rpm2-734::LEU2 (lane 8, mitochondrial RNA isolated from YPGE grown cells) RNAs were separated on a 6% denaturing gel. 5′ end represents the mature 5′-end of Rpm1r. SP and FP represent 5′-ends of primary transcripts, which originate at consensus promoter sequences downstream and upstream of the tRNAfMet gene, respectively. 3′ Metf represents the 5′-end of a product that is likely formed by an endonuclease cleavage at the 3′-end of tRNA fMet. (B) A single-stranded DNA probe was hybridized with mitochondrial RNA isolated from YPGE grown cells. The samples were digested with S1 nuclease, and protected fragments were separated on a 6% polyacrylamide denaturing gel. Probe alone (lanes 1 and 2, in the absence or presence of S1 nuclease, respectively); probe plus S1 nuclease in the presence of wild-type RNA (lane 3) and increasing amounts of rpm2-734::LEU RNA (10, 20 and 30 µg in lanes 4, 5 and 6, respectively). 3′ end represents the mature 3′end of Rpm1r. Intermediate marks a putative processing intermediate discussed in the text.
To determine if 3′-end maturation occurred, a DNA fragment spanning the mature 3′-end of Rpm1r was end-labeled, strand-separated, hybridized to mitochondrial RNA from wild-type and mutant cells, and digested with S1 nuclease as described (21). The major fragment protected by wild-type RNA corresponds to the mature 3′-end of Rpm1r and is labeled 3′-end (Fig. 4B, lane 3). Its identity was determined by sizing against a chemical sequencing ladder of the same fragment used in the protection experiment (data not shown). No corresponding reaction product was obtained from RNA isolated from the mutant (Fig. 4B, lanes 4–6). This result demonstrates that the 3′-end of mutant Rpm1r is extended relative to the wild-type Rpm1r 3′-end. There is a product formed in RNA isolated from both wild-type and mutant cells (Fig. 4B, lanes 3–6, Intermediate), that could have arisen from an RNA extended 80 nt downstream of the mature 3′-end of Rpm1r (21).
These results, together with those identifying the 5′-ends (Fig. 4A), predict discrete RNAs which were detected, but not separated, in the northern analysis shown in Figure 3B (lanes 1 and 2). The results reported above indicate that removal of the C-terminus of Rpm2p blocks processing of Rpm1r. Transfer RNA processing is only partially affected. This suggests that the C-terminal domain of Rpm2p plays a more significant role in the processing of Rpm1r than it plays in tRNA processing.
N- and C-terminal truncated Rpm2 proteins function in trans
To determine if the C-terminal domain can act in trans to restore full activity of Rpm2-ΔCp, we used three deletion mutants (Fig. 5A). The RPM2 alleles rpm2Δ735–1180 (ΔC) and rpm2Δ216–734 (ΔN) together cover the entire RPM2 sequence, while the rpm2Δ715–1097 (ΔC-20) and rpm2Δ216–734 (ΔN) pair is missing amino acids 715–734. All three ΔC, ΔC-20 and ΔN deletion mutants retain DNA coding for the putative mitochondrial targeting information at the N-terminus, so that the protein products they produce should be localized to mitochondria. We introduced the ΔC + ΔN, or ΔC-20 + ΔN genes, in combination on different low-copy plasmids, into Δrpm2 cells containing RPM2 on a URA3 plasmid. After selection for transformants, we then measured the ability of cells to grow on 5-FOA plates. Only cells that have lost the URA3 plasmid and have forms of Rpm2p that provide the essential function can grow. RPM2 and ΔC + ΔN containing cells appeared on 5-FOA plates at the same rate (Fig. 5B). However, the combination of ΔC(–20) and ΔN plasmids does not support growth on glucose medium. This indicates that amino acids 715–734 are essential for growth. Figure 5C demonstrates that cells expressing ΔC + ΔN fragments in trans grow on the non-fermentable carbon sources glycerol/ethanol at a rate similar, but slightly slower, compared to cells expressing an intact, plasmid-encoded Rpm2p. To determine if ΔC and ΔN are maintained in trans during these experiments, mitochondrial extracts were prepared and western analysis with anti-Rpm2p antibodies was performed. Our anti-peptide antibodies do not recognize Rpm2-ΔN protein because the deletion includes the epitope-coding region but it is clear from the phenotype that Rpm2-ΔNp is being produced. Figure 5D shows that Rpm2-ΔC protein from the in trans strain and Rpm2-ΔCp from the original disruption strain migrate identically. There was no full-length Rpm2p, which indicates that Rpm2-ΔC protein is maintained as a separate protein.
Figure 5.


N- and C-terminal truncated Rpm2p proteins function in trans. (A) A schematic representation of Rpm2 proteins. Numbers indicate amino acid residues. The filled box represents a putative mitochondrial targeting sequence. (B) RPM2, ΔC + ΔN or ΔC(–20) + ΔN alleles on different low-copy plasmids were introduced into a Δrpm2 haploid strain containing RPM2 on a URA3 plasmid and serial dilutions of transformants plated onto 5-FOA plates to assess growth in the absence of wild-type RPM2. (C) Strains were grown overnight in liquid YPGE medium and serial dilutions plated on YPD and YPGE. Cells were incubated 2 and 4 days for YPD and YPGE plates, respectively. (D) Mitochondrial proteins were isolated from the glycerol/ethanol grown cells shown in (C), separated on Tris–acetate–polyacrylamide gel electrophoresis and analyzed by immunoblotting using anti-Rpm2p polyclonal antibodies. The nuclear-encoded mitochondrial protein Mdh1p was used as a loading control. Lane 1, RPM2; lane 2, ΔC + ΔN; lane 3, ΔC (rpm2-734::LEU2). Protein molecular mass standards are indicated on the right.
Cells containing both ΔC and ΔN also maintain the ability to be transferred from glucose to glycerol/ethanol media, indicating that these cells keep their wild-type mtDNA independent of growth on a particular carbon source. Indeed, Figure 6 shows that the ΔC and ΔN genes together support Rpm1r processing and consequently RNase P activity (lanes 2 and 4 in upper and lower panel, respectively) in both fermentative and respiratory conditions. The amount of mature Rpm1r, however, does appear lower than that found in wild-type cells. Together, the results indicate that the C-terminus harbors a domain that is required for Rpm1r processing and is capable of functioning to a significant degree in trans.
Figure 6.
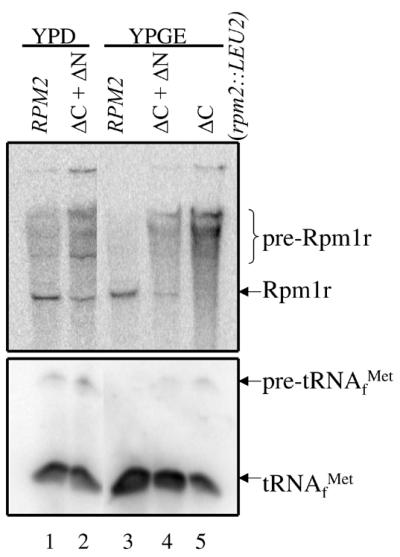
The C-terminal domain together with the N-terminal domain restores full activity of Rpm2p. Northern analysis of wild-type (lanes 1 and 3), ΔC + ΔN (lanes 2 and 4) and rpm2-734::LEU2 (lane 5) total RNAs. Upper panel shows the membrane probed with a riboprobe to Rpm1r; lower panel shows the membrane probed with a riboprobe to tRNAfMet.
DISCUSSION
Petite deletion mutants result when rpm2-734::LEU2 cells are grown on glucose (13) and even though Rpm1r and tRNA genes are retained, the tRNA gene transcripts are not processed. Providing RPM2 by transformation restores the tRNA processing ability to these petites. We report here that the same rpm2-734::LEU2 allele supports mitochondrial tRNA processing and maintenance of wild-type mtDNA if cells are grown on respiratory carbon sources. Therefore, under conditions that require respiration for growth, a truncated version of Rpm2p can support RNase P activity and provide enough tRNA for mitochondrial protein synthesis.
The observation that rpm2-734::LEU2 cells grow on respiratory carbon sources clearly demonstrates that the entire RPM2 open reading frame is not essential for mitochondrial RNase P activity. Nevertheless, the ratio of precursor to mature mitochondrial tRNA is greater in these cells relative to wild-type, indicating that removal of the C-terminus of Rpm2p has a negative effect on RNase P activity. This effect may not, however, be a direct effect of the truncation on RNase P activity but instead may be related to the role of Rpm2p in the maturation of Rpm1r, the RNA subunit of mitochondrial RNase P.
In Saccharomyces cerevisiae, Rpm1r is made as a part of a polycistronic transcript, tRNAfMet–Rpm1r–tRNAPro, that undergoes a series of processing steps to release mature Rpm1r (23). Previous results have shown that neither 5′- nor 3′-ends of Rpm1r are matured when rpm2-734::LEU2 cells are grown on glucose (21). Interestingly, the results shown here demonstrate that even though rpm2-734::LEU2 cells grown on non-fermentable carbon sources are capable of processing mitochondrial tRNAs, Rpm1r processing is blocked. Thus, the C-terminal region of Rpm2p plays a more significant role in Rpm1r processing than tRNA processing. Moreover, the role of the C-terminal region in Rpm1r processing can be provided in trans suggesting that it may be a relatively autonomous domain. Together, these results indicate that the primary defect in rpm2-734::LEU2 cells is in the processing of Rpm1r, and defects in tRNA processing may be linked to the absence of mature Rpm1r.
Rpm1r is necessary for mitochondrial RNase P activity both in vivo and in vitro (11,20). Therefore, in the absence of detectable mature Rpm1r, it is surprising to find mature tRNAs in rpm2-734::LEU2 mitochondria. This observation suggests that pre-Rpm1r may be capable of functioning as a component of RNase P. If this is the case, pre-Rpm1r is not as efficient as mature Rpm1r in supporting tRNA processing since tRNA precursors accumulate. In this context, it has been shown that bacterial RNase P RNA precursors function in tRNA processing in vitro (24). Alternatively, levels of mature Rpm1r below our detection limit may be supporting RNase P activity in rpm2-734::LEU2 cells grown on non-fermentable carbon sources.
The mechanism by which Rpm2p influences Rpm1r processing is unknown. There are several scenarios that could lead to the accumulation of Rpm1r precursors with both 5′ and 3′ extensions. First of all, the RNase P activity per se could be required for maturation of one, or both, ends of Rpm1r. In this context, addition of the Bacillus subtilis RNase P protein to a precursor form of the P RNA in vitro results in autolytic processing of the 5′- and 3′-ends of the precursor (25). Another possibility is that one or more additional nuclease activities necessary for Rpm1r maturation are encoded by RPM2 and function independently of RNase P. Finally, Rpm2p could play a chaperone role, facilitating Rpm1r folding into a conformation recognized by 5′- and 3′-end processing enzymes.
It is clear that growth conditions have a profound effect on the ability of the C-terminally truncated form of Rpm2p to provide RNase P activity in mitochondria. One possibility is that there is more enzyme produced in respiring cells and an increase in amount could compensate for reduced activity caused by the truncation. Indeed, increased expression of nuclear-encoded mitochondrial proteins, as well as expression of genes encoded in mtDNA, does accompany growth on glycerol/ethanol (26). Similarly, expression of genes involved in mitochondrial biogenesis are repressed by glucose (catabolite repression) and this decrease coupled with the truncation could cause RNase P activity to fall below the threshold required for mitochondrial protein synthesis necessary to maintain a wild-type mitochondrial genome. Cells with deletions of ABF2 and MMF1, two other yeast genes encoding mitochondrial proteins, also maintain their wild-type mitochondrial genome if grown by respiration but lose it when grown by fermentation (27,28).
While rpm2-734::LEU2 cells are capable of growth on non-fermentable carbon sources, their growth is slow. This reduction in growth rate could be a consequence of inefficient tRNA processing linked to truncated Rpm2p and/or inefficient Rpm1r processing. However, Cliften et al. (29) showed that there is normal respiratory growth if only 10–20% of wild-type mitochondrial transcripts are present. Clearly, at least 20% of tRNAs are processed in rpm2-734::LEU2 mitochondria suggesting that tRNA maturation may not be limiting for growth on respiratory carbon sources. Therefore, it is possible that the slow respiratory growth of rpm2-734::LEU2 might be a reflection of some other mitochondrial dysfunction. In this regard, we have recently demonstrated that Rpm2p, apart from its role in RNase P, has an additional function in the synthesis of mitochondrially-encoded subunits of cytochrome c oxidase (30). This function is also required for respiratory growth and may also be affected by the Rpm2p truncation.
An approach to understand why cells expressing the same Rpm2-ΔC protein display differences in their ability to maintain tRNA processing on different carbon sources would be to search for suppressors that allow maintenance of a wild-type mtDNA under fermentative growth. Preliminary results with this approach show promise and will be reported elsewhere.
Acknowledgments
ACKNOWLEDGEMENTS
We thank Ms Marlene Steffen and Mr Paul Weis for technical assistance. National Institutes of Health grant GM27597 to N.C.M. supported this work.
References
- 1.Hopper A.K. and Martin,N.C. (1992) Processing of yeast cytoplasmic and mitochondrial precursor tRNAs. In Jones,E.W., Pringle,J.R. and Broach,J.R. (eds), The Molecular and Cellular Biology of the Yeast Saccharomyces: Gene Expression. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, pp. 99–141.
- 2.Frank D.N. and Pace,N.R. (1998) Ribonuclease P: unity and diversity in a tRNA processing enzyme. Annu. Rev. Biochem., 67, 153–180. [DOI] [PubMed] [Google Scholar]
- 3.Wang M.J., Davis,N.W. and Gegenheimer,P. (1988) Novel mechanisms for maturation of chloroplast transfer RNA precursors. EMBO J., 7, 1567–1574. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Thomas B.C., Li,X. and Gegenheimer,P. (2000) Chloroplast ribonuclease P does not utilize the ribozyme-type pre-tRNA cleavage mechanism. RNA, 6, 545–553. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Stark B.C., Kole,R., Bowman,E.J. and Altman,S. (1978) Ribonuclease P: an enzyme with an essential RNA component. Proc. Natl Acad. Sci. USA, 75, 3717–3721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Pace N.R. and Brown,J.W. (1995) Evolutionary perspective on the structure and function of ribonuclease P, a ribozyme. J. Bacteriol., 177, 1919–1928. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Guerrier-Takada C., Gardiner,K., Marsh,T., Pace,N. and Altman,S. (1983) The RNA moiety of ribonuclease P is the catalytic subunit of the enzyme. Cell, 35, 849–857. [DOI] [PubMed] [Google Scholar]
- 8.Pannucci J.A., Haas,E.S., Hall,T.A., Harris,J.K. and Brown,J.W. (1999) RNase P RNAs from some archea are catalytically active. Proc. Natl Acad. Sci. USA, 96, 7803–7808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Myers A.M., Pape,L.K. and Tzagoloff,A. (1985) Mitochondrial protein synthesis is required for maintenance of intact mitochondrial genomes in Saccharomyces cerevisiae. EMBO J., 4, 2087–2092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Underbrink-Lyon K., Miller,D.L., Ross,N.A., Fukuhara,H. and Martin,N.C. (1983) Characterization of a yeast mitochondrial locus necessary for tRNA biosynthesis. Mol. Gen. Genet., 191, 512–518. [DOI] [PubMed] [Google Scholar]
- 11.Hollingsworth M.J. and Martin,N.C. (1986) RNase P activity in the mitochondria of Saccharomyces cerevisiae depends on both mitochondrial and nucleus-encoded components. Mol. Cell. Biol., 6, 1058–1064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Morales M.J., Dang,Y.L., Sulo,P. and Martin,N.C. (1992) A 105 kDa protein is required for yeast mitochondrial RNase P activity. Proc. Natl Acad. Sci. USA, 89, 9875–9879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Dang Y.L. and Martin,N.C. (1993) Yeast mitochondrial RNase P: sequence of the RPM2 gene and demonstration that its product is a protein subunit of the enzyme. J. Biol. Chem., 268, 19791–19796. [PubMed] [Google Scholar]
- 14.Kassenbrock C.K., Gao,G.-J., Groom,K.R., Sulo,P., Douglas,M.G. and Martin,N.C. (1995) RPM2, independent of its mitochondrial RNase P function, suppresses an ISP42 mutant defective in mitochondrial import and is essential for normal growth. Mol. Cell. Biol., 15, 4763–4770. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kaiser C., Michaelis,S. and Mitchell,A. (1994) Methods in Yeast Genetics. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
- 16.Chen D.C., Yang,B.C. and Kuo,T.T. (1992) One-step transformation of yeast in stationary phase. Curr. Genet., 21, 83–84. [DOI] [PubMed] [Google Scholar]
- 17.Sambrook J., Fritsch,E.F. and Maniatis,T. (1989) Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
- 18.Köhrer K. and Domdey,H. (1991) Preparation of high molecular weight RNA. Methods Enzymol., 194, 398–405. [DOI] [PubMed] [Google Scholar]
- 19.Chomczynski P. and Sacchi,N. (1987) Single-step method of RNA isolation by acid guanidinium thiocyanite-phenol-chlorophorm extraction. Anal. Biochem., 162, 156–159. [DOI] [PubMed] [Google Scholar]
- 20.Morales M.J., Wise,C.A., Hollingsworth,M.J. and Martin,N.C. (1989) Characterization of yeast mitochondrial RNase P: an intact RNA subunit is not essential for activity in vitro. Nucleic Acids Res., 17, 6865–6881. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Stribinskis V., Gao,G.-J., Sulo,P., Dang,Y.-L. and Martin,N.C. (1996) Yeast mitochondrial RNase P synthesis is altered in an RNase P protein subunit mutant: insights into the biogenesis of a mitochondrial RNA-processing enzyme. Mol. Cell. Biol., 16, 3429–3436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Fox T.D., Foley,L.S., Mulero,J.J., McMullin,T.W., Thorsness,P.E., Heidin,L.O. and Costanzo,M.C. (1991) Analysis and manipulation of yeast mitochondrial genes. Methods Enzymol., 194, 149–165. [DOI] [PubMed] [Google Scholar]
- 23.Martin N.C., Hollingsworth,M.J., Shu,H.-H. and Najarian,D.R. (1985) Expression and function of the tRNA synthesis locus in Saccharomyces cerevisiae. In Quagliariello,E. et al. (eds), Achievements and Perspectives of Mitochondrial Research. Elsevier Science Publishers, Amsterdam, Vol. 2, pp. 193–202.
- 24.Reich C., Gardiner,K.J., Olsen,G.J., Pace,B., Marsh,T. and Pace,N.R. (1986) The RNA component of Bacillus subtilis RNase P: sequence, activity, and partial secondary structure. J. Biol. Chem., 261, 7888–7893. [PubMed] [Google Scholar]
- 25.Loria A. and Pan,T. (2000) The 3′ substrate determinants for the catalytic efficiency of the Bacillus subtilis RNase P holoenzyme suggest autolytic processing of the RNase P RNA in vivo. RNA, 6, 1413–1422. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Ulery T.L., Jang,S.-H. and Jaehning,J.A. (1994) Glucose repression of yeast mitochondrial transcription: kinetics of derepression and role of nuclear genes. Mol. Cell. Biol., 14, 1160–1170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Diffley J.F. and Stillman,B. (1991) A close relative of the nuclear, chromosomal high-mobility group protein HMG1 in yeast mitochondria. Proc. Natl Acad. Sci. USA, 88, 7864–7868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Oxelmark E., Marchini,A., Malanchi,I., Magherini,F., Jaquet,L., Hajibagheri,M.A., Blight,K.J., Jauniaux,J.C. and Tommasino,M. (2000) Mmf1p, a novel mitochondrial protein conserved through evolution and involved in maintenance of the mitochondrial genome. Mol. Cell. Biol., 20, 7784–7797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Cliften P.F., Park,J.-Y., Davis,B.P., Jang,S.-H. and Jaehning,J.A. (1997) Identification of three regions essential for interaction between a σ-like factor and core RNA polymerase. Genes Dev., 11, 2897–2990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Stribinskis V., Gao,G.-J., Ellis,S.R. and Martin,N.C. (2001) Rpm2p, the protein subunit of mitochondrial RNase P in Saccharomyces cerevisiae also has a role in the translation of mitochondrially encoded subunits of cytochrome c oxidase. Genetics, 158, 573–585. [DOI] [PMC free article] [PubMed] [Google Scholar]