

Abstract
Placental malaria, characterized by sequestration of Plasmodium falciparum in the maternal placental blood space and associated inflammatory damage, contributes to poor birth outcomes and ~200,000 infant deaths annually. Specific mechanisms that contribute to placental damage and dysfunction during malaria are not completely understood. To investigate a potential role for oxidative stress, antioxidant genes and markers for oxidative damage were assessed by quantitative PCR and immunohistochemistry in Plasmodium chabaudi AS-infected pregnant mice. Widespread evidence of lipid peroxidation was observed and was associated with higher antioxidant gene expression in conceptuses of infected mice. To assess the extent to which this oxidative damage might contribute to poor birth outcomes and be amenable to therapeutic intervention, infected pregnant mice were treated with N-acetylcysteine, a free radical scavenger, or tempol, an intracellular superoxide dismutase mimetic. The results show that mice treated with N-acetylcysteine experienced malaria induced–pregnancy loss at the same rate as control animals and failed to mitigate placental oxidative damage. In contrast, tempol-treated mice exhibited subtle improvement in embryo survival at gestation day 12. Although lipid peroxidation was not consistently reduced in the placentas of these mice, it was inversely related to embryo viability. Moreover, reduced IFN-γ and CCL2 plasma levels in treated mice were associated with midgestational embryo viability. Thus, although oxidative stress is remarkable in placental malaria and its mitigation by antioxidant therapy may improve pregnancy outcomes, the underlying mechanistic basis and potential therapeutic strategies require additional investigation.
INTRODUCTION
According to the World Health Organization, malaria remains a major public health problem in many countries of the world. Despite the progress in reducing malaria cases and deaths, it is estimated that 214 million cases of malaria occurred worldwide in 2015, leading to 438,000 deaths (1). In areas with moderate to high transmission of Plasmodium falciparum, a package of inter ventions for controlling malaria and its effects during pregnancy is recommended (2). Malaria in pregnant women is characterized by the accumulation of P. falciparum–infected RBCs, leukocyte infiltration, and excessive fibrin deposition in the maternal blood space (3) of the placenta, a syndrome known as placental malaria (PM). PM is associated with low birth weight and maternal anemia (4–7).
In addition to these pathogenic findings in PM, a recent study showed that poor pregnancy outcomes may also be attributed to malaria-induced oxidative stress in the placenta (8). In that work, the level of cell membrane lipid peroxidation, detected as malondialdehyde, was significantly increased in the placentas of malaria-infected and anemic women compared with noninfected and nonanemic counterparts (8). Moreover, oxidative stress was observed in a murine model of PM caused by P. berghei ANKA (9–11), which yields many features of human PM but is limited by parasite virulence and maternal mortality, restricting studies to late pregnancy.
Oxidative stress is defined as an imbalance between the production of reactive oxygen species (ROS), including superoxide, hydrogen peroxide, hydroxyl anion, and the ability of various antioxidant mechanisms to scavenge these radicals. By catalyzing the dismutation of superoxide radical into oxygen or hydrogen peroxide, superoxide dismutase (SOD) reacts with NO and ROS as an antioxidant and mitigates their negative physiological effects (12). Three isoforms of SOD have been identified. SOD1 has copper and zinc as cofactors and is localized to intracellular cytoplasmic compartments. SOD2, the second isoform, has manganese in its reactive center and has been localized to mitochondria of aerobic cells. Extracellular SOD (SOD3) is synthesized by only a few cell types including vascular smooth muscle cells, and is localized in the extracellular matrix of various tissues including the vascular wall and placenta (13). Catalase (Cat), another main antioxidant enzyme responsible for ROS control, further detoxifies a pro-oxidant response by degrading hydrogen peroxide into water and oxygen. To our knowledge, the extent to which these mediators of an antioxidant response are active in PM during early pregnancy malaria has not been explored. To address this issue, an alternative murine model (3, 14–18) in which P. chabaudi chabaudi AS (P. chabaudi AS) infection in early gestation induces placental damage and pregnancy loss in C57BL/6 and A/J mice (3, 17, 18) was used. Because apoptosis does not seem to play a universal role in this placental damage (17), we hypothesized that, consistent with the P. berghei model of PM (9, 11), oxidative stress participates in the pathogenesis of P. chabaudi AS infection during early pregnancy. To this end, oxidative stress was measured by quantitative PCR of markers of an antioxidant response and by immunohistochemistry of a reactive lipid mediator generated during oxidative stress, 4-hydroxynonenal (HNE). Additionally, the ability of treatment with antioxidants (N-acetylcysteine [NAC] and tempol [TPL]) to block malaria-associated oxidative stress and improve pregnancy outcome was assessed. Finally, because oxidative stress and elevated ROS are associated with inflammation in human and experimental malaria (19, 20), systemic cytokine levels were measured in antioxidant-treated and untreated animals.
MATERIALS AND METHODS
Animals, parasites, and infection
All animal procedures reported in this study followed the Guide for the Care and Use of Laboratory Animals. Animal protocols were approved by the University of Georgia Institutional Animal Care and Use Committee.
Male and female C57BL/6J (B6) mice were originally purchased from the Jackson Laboratory and were used to generate breeding stock and experimental animals in the University of Georgia Coverdell Vivarium (18). All mice were kept under specific pathogen-free conditions at the Coverdell Center Vivarium (University of Georgia, Athens, GA), and were devoid of Helicobacter infection, which has a clear negative impact on pregnancy in C57BL/6 mice (21). P. chabaudi chabaudi, strain AS (MR4-741), contributed by D. Walliker, was obtained from the BEI Resources Repository, National Institute of Allergy and Infectious Diseases, National Institutes of Health. The parasite was maintained by routine passage through female A/J mice and cryopreservation. Infection in experimental female mice, aged 8–10 wk, was initiated intravenously with 1000 infected RBCs per 20 g of body weight on gestation day 0; this is considered experiment day (ED) 0; thus, ED is directly comparable to gestation day. Parasitemia was detected beginning on ED6 in Giemsa-stained tail blood thin smears by examining at least 1000 RBCs in four high-power microscopic fields (3, 14–18). Thereafter, parasitemia was monitored daily by flow cytometry using Sytox-Orange staining (22). Percent of hematocrit was used as an indicator of anemia. Weight was also recorded every day (except from ED1 to ED5) in grams (18), and percent of weight loss or gain was calculated as previously described (3).
All mice were sacrificed on ED10 or ED12 as indicated (3, 14–18). Plasma and tissues were collected as described below. Mice were dissected for evaluation of embryo status and isolation of tissues as previously described (14, 15, 17, 18). Briefly, resorptions or nonviable embryos were identified by their necrotic and/or smaller size compared with viable normal embryos as they appear in uninfected pregnant (UP) mice. Mice undergoing active abortion, defined as evidence of bloody, mucoid vaginal discharge and/or evidence of embryos in the open cervix or vaginal canal were recorded(3,14, 16,18).Theuterus was separated by cutting directly below the oviduct and above the cervix, and the mesometrium was removed.
Antioxidant treatment and outcome monitoring
Groups of 6–16 female mice were paired 2:1 with a male overnight and checked the following morning for vaginal plugs. Observation of a plug prompted randomization, as numbers allowed, to infected pregnant (IP) and UP groups. Each of these randomization events (on a single day) is considered to be a discreet experiment. These groups were then further randomized on either ED2 or ED6 to receive control or vehicle (PBS or H2O) or antioxidant therapy. The antioxidants used in this study were NAC (pH 7.4) and TPL (4-hydroxy-2,2,6,6-tetramethylpiperidin-1-oxyl). Pairing was continued until the desired number of animals was achieved and is detailed for each treatment regimen below.
For NAC treatment, mice were initiated on 14 discreet experiment start dates, with two to five plugged mice obtained per day. In cases where fewer than four plugs were observed, mice were alternated to IP or UP groups only to allow later allocation to control (PBS) and treated groups within each start date–defined experiment. In initial experiments, NAC was administered i.p. from ED6 to ED12 at the dose of 50 mg/kg per day, with control mice receiving injections of PBS, and sacrifice was scheduled for ED12; for this goal, eight UP NAC, eight UP PBS, five IP NAC, and five IP PBS mice were generated. When these experiments revealed that pregnancy loss was occurring prior to ED12, an additional 21 mice from eight discreet experiment start dates (two to five mice per experiment; six UP NAC, six UP PBS, six IP NAC, and five IP PBS) were generated for sacrifice at ED10 (with treatment received from ED6 to ED10). ED10 sacrificed mice allowed collection of conceptus tissue for evaluation of lipid peroxidation (see below) and collection of plasma for assessment of systemic cytokine levels at the time of abortion.
TPL (at the dose of 40 mg/kg per day, dissolved in methanol, and diluted in PBS) was first administered i.p. starting at ED6; control mice received sterile PBS supplemented as appropriate with diluent (methanol). Overall, eight experiment start days were used with the number of mice varying from three to six per experiment, ultimately yielding 5 UP TPL, 7 UP PBS, 7 IP TPL, and 11 IP PBS mice that were sacrificed on ED12.
In subsequent experiments, TPL was delivered ad libitum in drinking water at 40 mg/kg per day (23, 24) with daily changes, and was initiated at either ED2 or ED6 through ED10 or ED12. This concentration was calculated to be comparable on a per-day basis to the concentration delivered by injection. TPL consumption was confirmed by tracking the volume of liquid consumed over the entire cage of mice and average per-mouse consumption calculated. For the ED6 TPL treatment regimen, nine experiment start dates were included, with daily inclusion ranging from three to six mice. Total group sizes were 4 UP TPL, 6 UP H2O, 15 IP TPL, and 6 IP H2O for a total of 31 mice. All of these mice were sacrificed on ED12. For the TPL drinking regimen started on ED2, 20 experiment start dates (numbers of mice ranging from three to six) were performed and yielded 45 mice(6UPTPL,5UPH2O,12IP TPL, 10 IP H2O) for sacrifice on ED10 and 40 mice (5 UP TPL, 12 UP H2O, 10 IP TPL, 6 IP H2O; nine experiment start dates) for sacrifice on ED12.
The mean volume of TPL solution drunk per mouse per day was respectively 4.93 ± 0.29 and 4.48 ± 0.09 in UP TPL and IP TPL, whereas the mean volume of water drunk was 5.26 ± 0.18 and 4.28±0.09 in UP H2O and IP H2O.Variation in water and TPL uptake was not significant between treatment groups.
Placental histology and immunohistochemistry
At sacrifice (ED10 and ED12), conceptuses and spleens were immediately harvested from TPL- and vehicle-treated mice and fixed in 10% neutral formalin for 12 h. The fixed tissues were embedded in paraffin and sectioned at 7 mm. H&E-stained organ sections were used by a pathologist to assess malaria-associated histopathology and the effects of TPL treatment. Unstained 7 mm tissue sections (3) were used for immunohistochemistry (IHC) according to previously reported methods (17) with a Polink-HRP kit for rabbit Abs following the manufacturer’s instructions (GBI Mukilteo, WA). Samples were stained the same day with the same reagents to avoid interassay variability. Briefly, unstained tissue sections were first deparaffinized and rehydrated through xylene and graded alcohol series. Ag retrieval was performed for 30 mn with a Pascal pressurized heating chamber (Code S2800; Dako, Carpinteria, CA). Detection of HNE adducts, markers for lipid peroxidation, was achieved with a polyclonal rabbit anti-HNE (ab46545; Abcam, Cambridge, MA) at 1/2000 for 1 h in a moist chamber at 37°C. HNE is one of the most quantitatively important products of lipid peroxidation (25), a process by which oxidants such as free radicals or nonradical species attack lipids, especially polyunsaturated fatty acids, yielding lipid peroxyl radicals, and hydroperoxides (26). Sections were counterstained with hematoxylin, dehydrated and mounted with Flo-Texx (Lerner Laboratories, Pittsburgh, PA). Stained slides were viewed and photographed on a Nikon Eclipse E400 microscope with a Nikon DS-L1-5M imaging camera at 3200 final magnification to monitor staining and assess results. Scoring for HNE in murine spleen and placental tissues was done by examining 10 power fields of the stained slide and assigning a score on a 0–4 scale. High-resolution images for data presentation were taken with an Olympus BX41 microscope coupled with an Olympus Digital camera DP 25, saved as TIFF files, and cropped and resized using Adobe Photoshop CS6.
Antioxidant gene expression by quantitative PCR
Total RNA from mouse conceptuses (ED9, 10, and 11) was extracted using Trizol Reagent (Ambion, Carlsbad CA), DNase-treated (Ambion), reverse-transcribed with High Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA). Genes of interest (Table I) were quantified with Power Sybr Green Master Mix (Applied Biosystems, Warrington, U.K.) with the C1000 Touch Thermal Cycler (CFX96 Real Time Systems; Bio-Rad, Hercules CA). Each sample was assayed in duplicate for all targets (antioxidant genes) and the housekeeping (18S RNA) gene. Average threshold cycle (Ct) values were normalized to the average of 18S RNA Ct values and relative expression was calculated for each target gene by the DDCt method (27). Details of mouse-specific primer sets (sequence forward and reverse) as well as the size of the amplification products and gene name are summarized in Table I.
TABLE I.
List of mouse-specific forward (PF) and reverse (PR) primers used in real time PCR for the amplification of pro-oxidants and antioxidant genes as well as housekeeping 18S rRNA and size of their amplification product in this study
| Gene Name | Forward and Reverse Primers | Size |
|---|---|---|
| PF: 5′-CCAGAGCGAAAGCATTTGCCAAGA-3′ | ||
| 18S RNA | PR: 5′-TCGGCATCGTTTATGGTCGGAACT-3′ | 106 |
| PF: 5′-CTACTCCCAGGTTGCCCACA-3′ | ||
| Nrf2 | PR: 5′-CGACTCATGGTCATCTACAAATGG-3′ | 103 |
| PF: 5′-GATGAAGAGAGGCATGTTGGA-3′ | ||
| Sod1 | PR: 5′-CCATTGTACGGCCAATGATGGAATG-3′ | 123 |
| PF: 5′-CACAAGCACAGCCTCCCAG-3′ | ||
| Sod2 | PR: 5′-CGCGTTAATGTGTGGCTCC-3′ | 63 |
| PF: 5′-TTCTTGTTCTACGGCTTGCTAC-3′ | ||
| Sod3 | PR: 5′-CTCCATCCAGATCTCCAGCACT-3′ | 174 |
| PF: 5′-AAATGCTTCAGGGCCGCCTT-3′ | ||
| Cat | PR: 5′-GTAGGGACAGTTCACAGGTA-3′ | 93 |
| PF: 5′-GGTGATGGCTTCCTTGTACC-3′ | ||
| Ho | PR: 5′-AGTGAGGCCCATACCAGAAG-3′ | 155 |
ELISA for TNF, IL-6, IFN, and CCL2
Plasma was collected from IP and UP mice by cardiac puncture at sacrifice on ED10. Cytokines and chemokine levels were assessed by ELISA using reference standard curves with known amounts of recombinant cytokines (TNF, IL-6, and IFN) and chemokine (CCL2) following the manufacturer’s instructions (BD and R&D as appropriate). For measurement of CCL2 in mouse conceptuses, total protein was isolated as previously described (17). Total proteins were quantified by CBQCA Protein Quantification Kit and diluted in assay diluent at the concentration of 1 mg/ml for use in ELISA.
Statistical analysis
Unless otherwise noted, the Graph Pad Prism Statistical Software (Version 5.03; GraphPad Software, La Jolla, CA) was used for quantitative data analysis. Sample sizes were in line with previous literature (cumulative 5–12 mice per group). Changes in percent starting weight, hematocrit, and parasitemia between groups were assessed by comparison of area under the curve (AUC). AUC was calculated in GraphPad Prism for weight using the ratio of starting weight (weight at day x divided by weight at day 0). AUC for hematocrit and parasitemia were calculated using the raw data for hematocrit and parasitemia, respectively. Nonparametric Mann–Whitney U test comparisons and Kruskal–Wallis test with Dunn multiple comparison test were applied when data were not normally distributed for pairwise and multiple group comparisons, respectively. Otherwise, Student t test and one-way ANOVA with Bonferroni multiple comparison test were used. A two-tailed Fisher exact test was used to compare embryo viability between antioxidant-treated versus control mice. A p value <0.05 was considered statistically significant.
RESULTS
Antioxidant gene expression is elevated in P. chabaudi AS–exposed conceptuses
Our mouse model of PM has consistently found that mice infected with P. chabaudi AS at ED0 abort their embryos at midgestation with no viable embryos remaining on ED12 (3, 16–18). To verify the extent to which embryos and their placentas are subjected to oxidative stress in this model, expression of genes involved in driving the transcriptional response to oxidative stress, NF erythroid derived 2 like-2 (Nrf2), and in detoxification of ROS, superoxide dismutase 1 (Sod1), Sod2, Sod3, and Cat as well as heme oxygenase (Ho), mRNA were assessed by quantitative PCR. On ED9, 1 d before abortion starts in IP B6 mice (3, 16–18), Nrf2, Sod1, Sod2, and Cat increased 4- to 5-fold relative to UP conceptuses, whereas Sod3 was upregulated with the fold increase reaching almost 20 (Fig. 1A). Sod2 remained elevated through ED10; Sod3 was strongly upregulated through ED11, peaking at a nearly a 30-fold increase on ED10 (Fig. 1A). Ho was modestly increased roughly 2-fold during the tested period (Fig. 1A).
FIGURE 1. Malaria-induced antioxidant response and lipid peroxidation in P. chabaudi AS–infected C57BL/6 (B6) conceptuses.
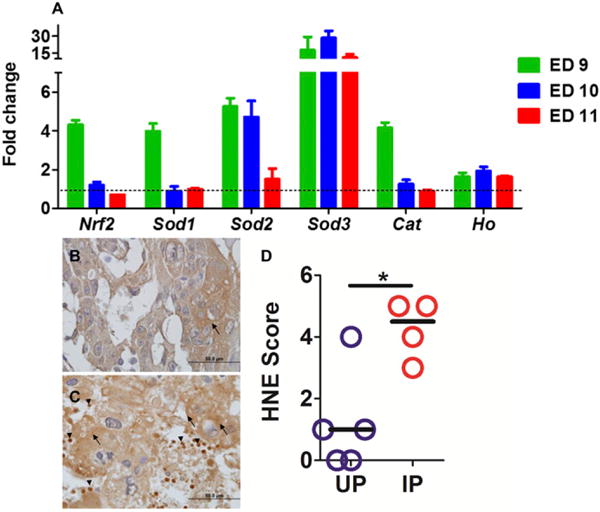
(A) Transcript levels of mouse Nrf2, Ho, Sod1, Sod2, Sod3, and Cat were quantified by quantitative PCR using forward and reverse primer sequences shown in Table I. Expression in IP mice is relative to the mean expression value in UP mice and normalized to mouse 18S RNA values. Columns and bars represent mean ± SEM of the relative quantities (fold-changes) for each gene from at least three conceptuses per mouse with a minimum of five mice per group. Representative images of UP (B) and IP (C) placenta stained for lipid adducts using an anti-HNE Ab. Arrows indicate stained trophoblast; arrowheads depict HNE staining in parasite-infected and uninfected RBCs in maternal blood space and within erythrophagocytosing trophoblast. (D) Summary scores for HNE staining. *p < 0.05.
Mouse PM is associated with lipid peroxidation
To verify that oxidative stress occurs in P. chabaudi AS–infected mouse placentas, in situ generation of HNE at ED10 was assessed. As shown in representative images (Fig. 1B, 1C), infected placentas showed significant (p = 0.0137; Fig. 1C, 1D) lipid peroxidation by IHC relative to uninfected placentas (Fig. 1B, 1D).
Antioxidant NAC does not rescue poor pregnancy outcomes in B6 mice
Based on the observations of increased antioxidant transcriptional response and lipid peroxidation in infected conceptuses (Fig. 1), we hypothesized that oxidative stress contributes to the pathogenesis of PM in this model and, in addition, that therapeutic interventions targeting oxygen radicals will improve pregnancy outcome. To this end, the efficacy of NAC was first tested. NAC interacts with ROS such as superoxide and hydrogen peroxide (28–33), and decreases the generation of free radicals by increasing glutathione synthesis (34, 35). Treatment was started on ED6 and ended on ED12, encompassing the time of detectable parasitemia to pregnancy loss in untreated P. chabaudi AS–infected pregnant B6 mice (3, 14, 16–18). Treatment efficacy was assessed by two outcome measures: embryo viability and uterus weight at ED12. Contrary to expectations, i.p.-delivered NAC (36) did not improve pregnancy outcome (Supplemental Fig. 1, Tables I, II). In fact, 50% of NAC-treated IP mice aborted earlier (ED8) than PBS-treated IP mice, indicating a potential toxic effect. IP uteri were significantly lighter that UP uteri (Supplemental Fig. 1G) and all embryos were nonviable at ED12 regardless of treatment (Table II). By AUC analysis, percent weight change (Supplemental Fig. 1B), percent hematocrit (Supplemental Fig. 1D), and percent parasitemia (Supplemental Fig. 1F) were not significantly different between NAC- and PBS-treated mice. These results indicate that, at the dose used in this study, NAC failed to improve pregnancy outcome in P. chabaudi AS–infected mice. Due to the observed toxicity of NAC and its failure to mitigate lipid peroxidation in spleen (Supplemental Fig. 2L) and placenta (Supplemental Fig. 2H, 2J), this therapeutic option was abandoned.
TABLE II.
Comparative percent of embryo viability in antioxidant-treated mice versus vehicle-treated mice
| Drug | Route | Started ED | Uninfected Pregnant (UP)
|
Infected Pregnant (IP)
|
||||
|---|---|---|---|---|---|---|---|---|
| Treated Viable/Total (%; n) | Vehicle Viable/Total (%; n) | p Value | Treated Viable/Total (%; n) | Vehicle Viable/total (%; n) | p Valuea | |||
| NAC | Injection | 6 | 41/43 (95; 5) | 44/46 (96; 5) | NS | 0/43 (0; 6) | 0/47 (0; 6) | NS |
| TPL | Injection | 6 | 39/43 (90; 5) | 42/44 (95; 5) | NS | 7/65 (10; 7) | 0/55 (0; 6) | 0.01 |
| TPL | Drinking | 2 | 40/42 (95; 5) | 80/85 (94; 10) | NS | 15/84 (18; 10) | 0/44 (0; 6) | 0.002 |
| TPL | Drinking | 6 | 38/41 (93; 4) | 62/67 (92; 6) | NS | 12/69 (17; 12) | 0/25 (0; 4) | 0.01 |
The pvalues were obtained with a two-tailed Fisher exact probability test when comparing UP-treated versus vehicle-treated and IP-treated versus IP-vehicle, respectively. Bold p values indicate significance between IP-treated and IP-vehicle.
%, proportion of viable embryos; ED, experiment day or gestation day, referring to the day when treatment was started; TPL, tempol; viable/total, the number of viable embryos divided by the total number of embryos on ED12.
TPL delivered i.p. incrementally improves pregnancy outcomes in P. chabaudi AS–infected mice
Given the failure of NAC to alleviate poor pregnancy outcomes in P. chabaudi AS–infected pregnant mice, we next turned to TPL, a nitroxide compound that has been studied extensively in animal models of increased ROS and oxidative stress (37). It metabolizes superoxide anions via a reducible nitroxide group (38) to form hydrogen peroxide, accounting for its superoxide dismutase mimetic action (37, 38). An advantage of using TPL is that it is cell permeable, specifically targeting superoxide anions that are generated intracellularly (39). Intraperitoneally delivered TPL did not impact the outcome of infection in terms of weight change (Fig. 2A, 2B), anemia (Fig. 2C, 2D), or course of parasitemia (Fig. 2E, 2F). Although uterus weight was reduced with infection regardless of treatment (Fig. 2G), the number of embryos per uterus (Fig. 2H) was equivalent in all groups (p > 0.05). Importantly, IP TPL-treated mice showed a significantly higher proportion of viable embryos relative to untreated IP mice (Table II). This result suggests that, unlike NAC, TPL has a modest protective effect for pregnancy outcomes in P. chabaudi AS–infected pregnant B6 mice.
FIGURE 2. Course of P. chabaudi AS infection and pregnancy outcome in mice receiving i.p.-delivered TPL from ED6 to ED12.
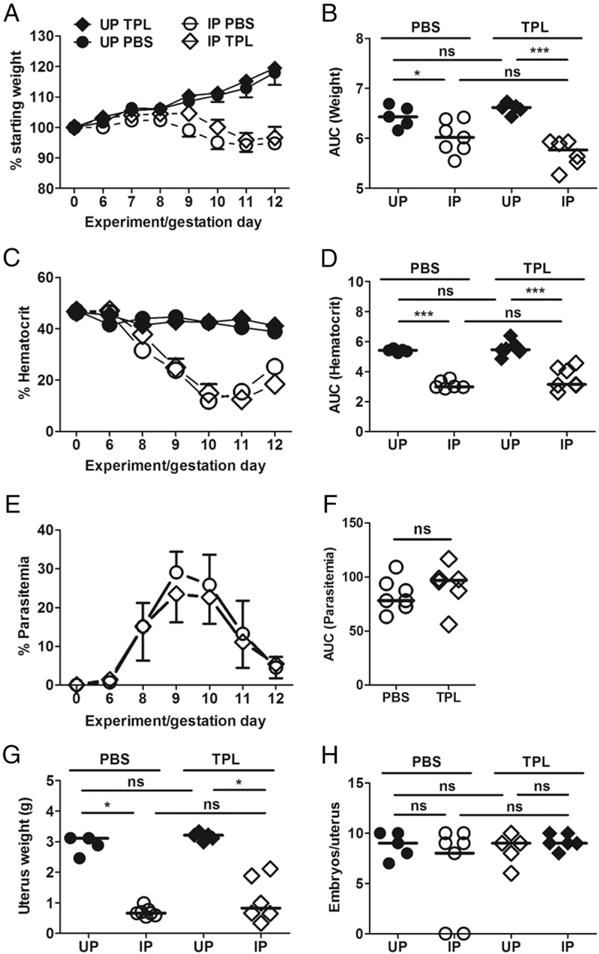
Unlike UP mice TPL-treated or not (UP PBS), percent weight change (A) and AUC for weight change (B) show evidence of IP mice losing weight. Percent hematocrit (C) and AUC for hematocrit (D) also depict anemia in IP mice whereas UP mice have stable hematocrit. Percent parasitemia (E) and AUC for parasitemia (F) are comparable in TPL-treated versus control mice. Pregnancy outcome as indicated by uterus weight (G) shows incremental improvement in pregnancy outcome in IP TPL-treated versus control mice. (H) Number of embryos per uterus. Data shown represent cumulative data for 30 mice from eight experiments. IP-PBS, open circle; IP-TPL, open diamond; UP-PBS, closed circle; UP-TPL, closed diamond. *p < 0.05, **p < 0.01, ***p < 0.001.
TPL delivered via drinking water partially improves pregnancy outcomes in P. chabaudi AS–infected mice
Given the promise of TPL treatment by injection, we next tested whether the outcome could improve with constant administration via drinking water at the same dose (40 mg/kg per day) as previously described in models for fetal growth restriction and preeclampsia (23, 24). One regimen started on ED6 (Supplemental Fig. 2) and the second on ED2 (to allow longer exposure to the drug; Fig. 3). In both cases, ED12 was initially chosen as the endpoint. Weight gain was suppressed in control IP mice relative to UP mice as expected (Fig. 3A, 3B) (3, 14, 16–18). With treatment starting on ED2, weight change among the dams was not different between the TPL-treated groups, apparently due to reduced weight gain in UP mice (Fig. 3A, 3B). As observed with injected TPL, treatment did not improve anemia (Fig. 3C, 3D), nor alter the development of parasitemia (Fig. 3E, 3F). Likewise, uterus weight was reduced with infection regardless of treatment (Fig. 3G). Because uterus weight in control UP mice was comparable to that in treated UP mice (Fig. 3G), reduced weight gain in the latter (Fig. 3A) was not due to embryo toxicity; in fact, the proportion of viable embryos in TPL-treated (93%) relative to UP controls (94%; Table II) was equivalent. Notably, uterus weight in treated IP mice was not different from that of treated UP mice (Fig. 3G), likely owing to improved outcome (increased uterus weight) in 2 out of 10 IP mice. Improved outcome is also reflected in the number of embryos per uterus (Fig. 3H), which differed across the groups (Kruskal–Wallis test, p < 0.0218; pairwise post hoc tests did not yield significant results). Indeed, the proportion of viable embryos in IP TPL-treated mice was significantly higher than in the untreated IP cohort (Table II). Consistent with this, the number of viable embryos and uterus weight were positively correlated in the treated IP mice (Spearman r = 0.86; p = 0.0045; Supplemental Fig. 3).
FIGURE 3. Course of P. chabaudi AS infection and pregnancy outcome in ad libitum orally delivered TPL treatment from ED2 to ED12.
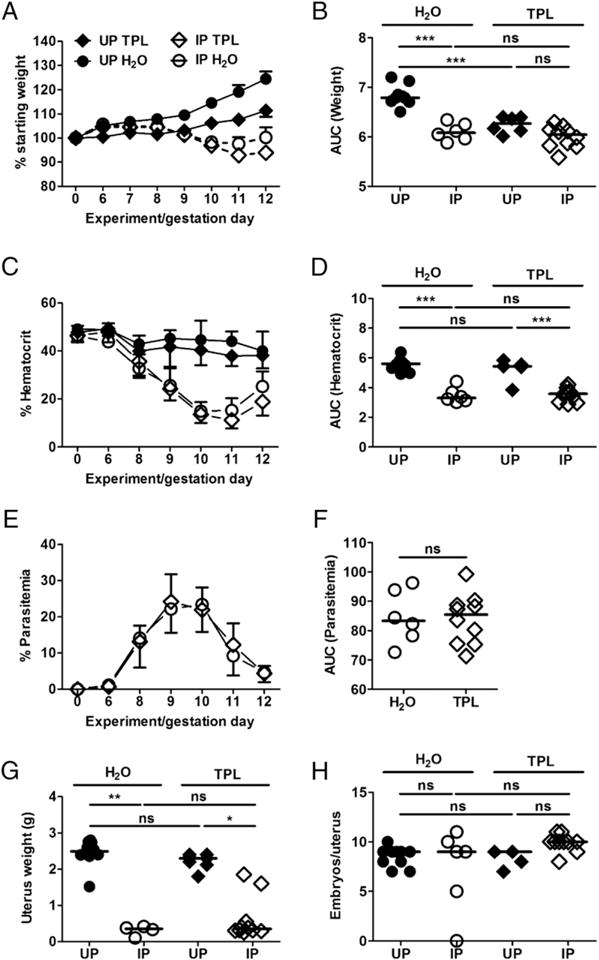
Percent weight change (A) with AUC (B) are indicated. Percent hematocrit (C) and AUC (D) depict anemia in IP mice, whereas UP mice have stable hematocrit. Percent parasitemia (E) and AUC (F) are comparable in mice orally delivered TPL versus mice under drinking water. Uterus weight (G) shows relative improvement of pregnancy outcome in mice receiving treatment versus those under drinking water, whereas number of embryos per uterus is the same regardless of infection or treatment status (H). The data from this experiment cumulatively represent 31 mice (5 UP-TPL, 10 UP-H2O, 10 IP-TPL, and 6 IP-H2O) generated with 10 experiment start days. IP-H2O, open circle; IP-TPL, open diamond; UP-H2O, closed circle; UP-TPL, closed diamond. Number of embryos per uterus varies across the groups (Kruskal–Wallis test, p = 0.0218). Pairwise post hoc tests are not significant. *p < 0.05, **p < 0.01, *** p < 0.001. ns, not statistically significant.
Finally, similar improved pregnancy outcomes were observed with oral TPL treatment initiated at ED6 (Supplemental Fig. 4, Table II). Collectively, these results indicate that TPL delivered orally ad libitum has a limited but significant positive effect on embryo survival in IP mice.
Chronic TPL exposure does not significantly mitigate placental lipid peroxidation
To assess the extent to which TPL treatment improves embryo viability by reducing placental oxidative stress, placental lipid peroxidation at ED10 in mice receiving oral TPL from ED2 was compared with control mice.
Lipid peroxidation was evident in placentas from some TPL-treated (Fig. 4D, 4H), and all control mice (Fig. 4B, 4F). Interestingly, there was significant variability in HNE staining among IP placentas, with some specimens having staining scores comparable to those from UP dams (Fig. 4N). This suggests variable efficacy for superoxide scavenging by TPL. Importantly, embryo viability was negatively correlated with the IHC score (Spearman r = −0.82; p = 0.0341; Fig. 4O), indicating that in cases in which TPL treatment did mediate reduction in placental lipid peroxidation, a protective effect (i.e., enhanced embryo viability) was achieved. In contrast to the lipid peroxidation patterns observed in placentas, splenic HNE was substantially and universally reduced in TPL-treated animals (Fig. 4K, 4L). Thus, whereas orally administered TPL effectively scavenges ROS and protects the spleen against lipid peroxidation in IP mice (Fig. 4M), this treatment is inconsistently effective at the placental level (Fig. 4N).
FIGURE 4. Immunostaining for 4-hydroxynonenal, marker of lipid peroxidation, in ED10 placenta and spleen from mice orally administered TPL beginning at ED2 versus mice under drinking water (H2O).
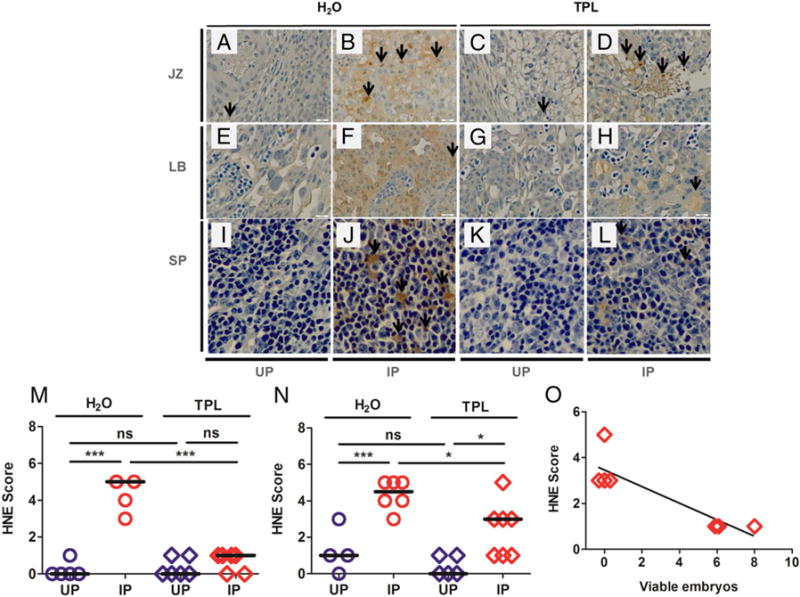
The two upper panels show immunostaining in the placental junctional zone [JZ; (A–D)] and labyrinth [LB; (E–H)] and the lower panels show immunostaining in the spleen [SP; (I–L)] of UP and P. chabaudi AS–IP mice. (M) shows HNE staining scores in the spleen and panel (N) shows staining in the placenta, whereas (O) depicts the correlation between viable embryos and HNE immunostaining scores in the placenta of TPL-treated IP mice. The data from this experiment were generated once with at least five mice per group, representing 45 mice [6 UP TPL, 5 UP H2O, 12 IP TPL, 10 IP H2O; 10 experimental start days (three to six mice per experiment)]. *p < 0.05, **p < 0.01. ns, not statistically significant.
Impact of chronic TPL treatment on local and systemic inflammatory markers
To assess the impact of TPL treatment on the inflammatory response in pregnant mice receiving treatment from ED2, the levels of cytokines (TNF, IL-6, and IFN-γ) and the chemokine CCL2 were assessed at ED10 in plasma and protein extracts from conceptus tissue by ELISA (Fig. 5). Irrespective of treatment, the levels of TNF, IL-10, and CCL2 were universally higher in IP compared with UP mice (Fig. 5B, 5D, 5E); CCL2 was also elevated in conceptus lysates from IP mice (Fig. 5F). IFN-γ (Fig. 5A) and IL-6 (Fig. 5C) also varied across the groups (Kruskal–Wallis test, p < 0.05), but pairwise post hoc tests did not yield significant results. When the levels of these factors were assessed as a function of pregnancy outcome (aborting or not) among IP mice, both IFN-γ (Fig. 6A) and CCL2 (Fig. 6E) were found to clearly segregate to low levels in nonaborting mice and high levels in aborting mice in TPL-treated mice, but not in the control group. Interestingly, TPL treatment appeared to suppress TNF among aborting mice (Fig. 6B). Tissue CCL2 (Fig. 6F) differed across the groups (Kruskal–Wallis test, p, 0.05), but pairwise post hoc tests did not yield significant results. Neither IL-6 (Fig. 6C) nor IL-10 (Fig. 6D) levels varied significantly as a function of treatment or outcome.
FIGURE 5. Levels of cytokines and chemokines in TPL-treated (from ED2) and control (H2O) mice at midgestation (ED10).
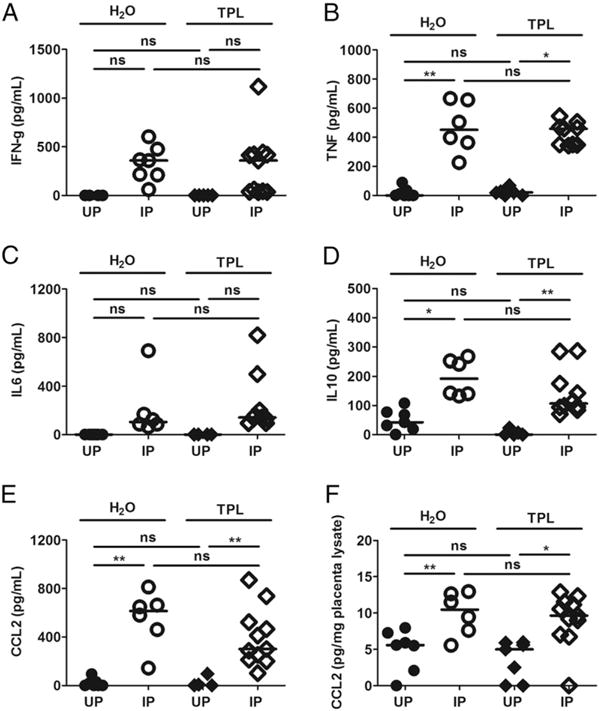
Levels of plasma IFN (A), TNF (B), IL-6 (C), IL-10 (D), and CCL2 (E), as well as placental tissue lysate CCL2 (F) are indicated. The data shown are derived from 10 experimental start days (three to six mice per experiment), which yielded a total of 45 mice. The Kruskal–Wallis test showed significance across the groups for IFN-γ (p = 0.0120) and IL-6 (p = 0.0219), but pairwise post hoc tests did not. *p < 0.05, **p < 0.01. ns, not statistically significant.
FIGURE 6. Levels of cytokines and chemokines in TPL-treated and control (H2O) infected pregnant mice undergoing abortion (A) or not (NA) at midgestation (ED10).
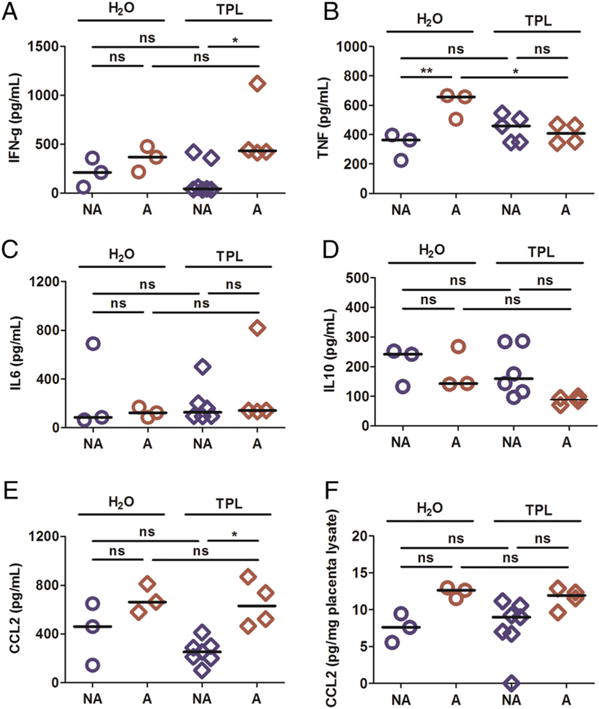
Levels of plasma IFN (A), TNF (B), IL-6 (C), IL-10 (D), and CCL2 (E) as well as placental tissue lysate CCL2 (F) are indicated. The data shown are derived from 10 experimental start days, which yielded a total of 45 mice. The Kruskal–Wallis test showed significance but the post hoc tests did not. *p < 0.05. ns, not statistically significant.
DISCUSSION
In this study, a potential role for placental oxidative stress in compromise of early pregnancy in P. chabaudi AS infection was explored. Placental antioxidant gene expression and lipid peroxidation in mouse conceptuses were examined and the impact of antioxidant treatment on midgestational pregnancy outcomes in malaria-infected mice was assessed. We present evidence that supports the hypothesis that oxidative stress contributes to poor pregnancy outcomes during PM.
Previous studies in murine P. berghei NK65 infection in late pregnancy malaria and in human PM have suggested that malaria impacts the antioxidant response in the placenta and promotes oxidative stress (8, 9, 11). Our results corroborate those studies.
In P. chabaudi AS infection in B6 mice, we observed increased expression of Nrf2 in conceptuses. This increase in Nrf2 mRNA was coincident with peak parasitemia, and corresponded to the day before abortion begins in P. chabaudi AS–infected mice. This upregulation was observed together with increased expression of antioxidant genes Sod1, Sod2, Sod3, and Cat. Expression levels for most of these genes returned nearly to baseline on ED10. A different pattern for placental catalase activity was observed in BALB/c mice infected at midgestation with P. berghei NK65: relative to uninfected mice during ascending parasitemia, catalase activity was reduced (9). Although the basis for this discrepancy is not clear, there are multiple obvious differences in study design between that and the present work. It is noteworthy that P. berghei ANKA infection in B6 mice was associated with a significant decrease in Cat and Sod1 expression in the brain (40). Thus, different parasite species may mediate variable oxidative responses in mice.
The most remarkable antioxidant response in the P. chabaudi AS–infected placenta was Sod3, the expression of which was highly elevated at all tested time points. Although the source of these transcripts in the placenta was not identified, it is noteworthy that this enzyme plays an important role in scavenging ROS extracellularly (41). In PM, where sequestered parasites (42) and responding immune cells (19, 20, 43) are likely to represent a potent source of ROS, extracellular SOD may be critical to counterbalance high levels of oxidative stress. Further studies will be required to determine the extent to which this particular enzyme may be a critical player in the response to PM, as well as cellular sources.
Regardless of the observed significant upregulation of antioxidant gene expression, intense staining for HNE (lipid peroxidation) in the placentas of infected mice suggests that the local antioxidant response was inadequate to protect the placenta from infection-induced oxidative stress. Given that excessive or uncontrolled production of HNE interferes with cellular signaling, and a role in the pathogenesis of several diseases, including severe malaria, has been suggested (44), placental lipid peroxidation may be central to PM-associated compromise of pregnancy, Indeed, at very high levels, HNE reportedly induces apoptosis or necrosis (45). Although we previously demonstrated that the former is unlikely to mediate embryo loss in infected B6 mice (17), necrotic placental damage is observed (D. Sarr, J. Ecker, J.M. Moore, unpublished observations). The extent to which necroptosis is operational in PM has not been reported. At moderate levels, HNE promotes organelle and protein damage leading to induction of autophagy (45). Inhibition of mTOR signaling has recently been shown to provide a link between human PM and low birth weight (46), but has not been explored in the context of malaria-induced oxidative stress.
Considering the coincidence of the antioxidant response and embryo loss in P. chabaudi AS–infected mice, it is likely that oxidative stress plays an important role in PM pathogenesis. Compromised fetal development, resulting in intrauterine growth retardation, is a hallmark of human PM (47, 48). Notably, in noninfected pregnancies, intrauterine growth retardation is associated with increased free radicals and markers of lipid peroxidation in maternal peripheral blood (49). The association of pregnancy complications like pre-eclampsia, recurrent spontaneous abortion, and antiphospholipid syndrome with oxidative stress has motivated a number of intervention studies with antioxidant therapies (50–53). This precedence led to our hypothesis that antioxidant drug delivery, by mitigating oxidative stress, may improve pregnancy outcomes in our mouse model. In keeping with the observed elevated expression of Sod3, which was suggestive of extracellular oxidative stress, mice were first treated with NAC, an extracellular reactive oxygen scavenger. Although accumulation of maternal inflammatory monocytes and macrophages, which might contribute ROS, is not evident in placentas of P. chabaudi AS–infected pregnant B6 mice (17), an NAC treatment approach was rationalized by work in a rat model of LPS-induced preterm labor and fetal demise, which responded positively to NAC pretreatment (54) at the same concentrations used in this study. Contrary to expectations, however, NAC mediated no protective effect in malaria-infected mice, and in fact, appeared to be embryotoxic. This may be related to the ability of NAC, at low concentrations, to actually enhance superoxide anion generation (37), negating any potential antioxidant effect and resulting in no net change in HNE generation in placenta and spleen. In the absence of an improved outcome with the concentration and timing of NAC used in this study, we were not motivated to test alternative delivery modes or regimens. Rather, we next tested TPL, an SOD mimetic that has been shown to cross biological membranes (39) and effectively scavenge superoxide (55–58). A recent study in experimental cerebral malaria showed TPL slightly increased survival in treated mice (59). Similar to that model, i.p. delivery of TPL to malaria-infected mice increased embryo survival by a modest 10%. Encouraged by that outcome, however, we tested TPL efficacy via oral administration. This mode, with a 6- or 10-d treatment period, yielded outcomes that were comparable to delivery via injection; that is, modest improvement in embryo viability in TPL-treated IP mice. Unexpectedly, UP mice receiving oral TPL treatment had significantly lower weight gain than UP controls. A previous study found that oral administration of TPL to obese/Zucker rats fed a high-fat diet reduced abdominal fat and weight gain, without changing food intake (60). Additionally, mice fed TPL from birth had a reduced weight gain but appeared fully healthy (61, 62). In the current study, reduced pregnancy-associated weight gain in TPL-treated uninfected mice was not reflected in uterus weight, which was comparable in treated and control mice, nor did it negatively impact number of embryos per dam. Thus, we conclude that there is no embryotoxicity with TPL. Indeed, studies designed to assess the efficacy of TPL in improving pregnancy outcome in mouse models of preeclampsia (24) and fetal growth restriction (23) found that oral administration of this drug enhances fetal growth (23, 24). These studies used TPL at the concentration of 1 mmol/l in drinking water starting at 2 d before mating (24) and gestation day 12.5 (23), respectively. The present work used a comparable drug concentration, but outcomes were relatively modest; this could be due to a more intense impact of malaria infection on embryo viability. Interestingly, whereas TPL treatment did not significantly impact placental ROS levels in the fetal growth restriction model (23), we observed a reduction of placental lipid peroxidation in some animals, with staining being negatively correlated with embryo viability. Thus, TPL is variably efficacious at the placental level in the context of malaria infection. The basis for this variation is not clear, but is unlikely to be attributable to variation in drug consumption or parasite dynamics, given that lipid peroxidation in the spleen was effectively and consistently controlled in all treated animals. Importantly, other studies have shown an organ-specific effect of TPL (37). In the absence of confirmation of the drug in plasma or urine, we tentatively conclude that bioavailability for TPL in the placenta is poor, but in cases in which it can reach effective local concentrations, oral TPL can positively impact embryo outcome and suppress placental lipid peroxidation.
Our readout for both oxidative damage and TPL efficacy, aside from infection parameters and embryo outcome, was the detection of placental HNE. It is important to acknowledge that HNE can be contributed exogenously, such as by components of the malaria parasite, rather than being generated in situ by endogenous ROS. Only the latter would be targeted by TPL (37, 38), because this drug will not have a direct effect on de novo generation of HNE by the parasite, namely via hemozoin. Notably, HNE generated by hemozoin in an infected erythrocyte can be transferred to uninfected erythrocytes (63); it is therefore plausible that parasite-derived HNE is transferred to trophoblast in the infected mouse by accumulating parasites (16, 18), and may overcome the effects of NAC and TPL. Hemozoin, which accumulates in trophoblast in mouse (15) and human (64) PM, could also directly contribute to HNE production in situ. Indeed, HNE staining in maternal erythrocytes is evident in P. chabaudi AS–infected placentas (Fig. 1C). Future studies in our model and others will be necessary to confirm the sources and contribution of HNE to malaria pathogenesis in pregnancy.
Finally, previous studies have demonstrated the anti-inflammatory effects of TPL (65, 66), including in experimental cerebral malaria (59). Because inflammatory responses are critical in PM pathogenesis (3, 14–18), the extent to which these responses are altered was a logical consideration in TPL-treated mice. Interestingly, just as TPL efficacy in altering HNE production was variable in malaria-infected pregnant mice, so was its ability to alter systemic inflammatory responses. Cytokine levels were not different in IP mice as a function of treatment status; however, plasma levels of IFN-γ and CCL2 were clearly segregated in TPL-treated animals, with lower levels being observed in nonaborting relative to aborting mice. Thus, an indicator of TPL efficacy might be its ability to suppress inflammatory cytokine and chemokine production. Future studies of antioxidant therapy in IP mice will need to resolve the relative contributions of TPL-mediated antioxidant and anti-inflammatory effects.
In summary, this study provides evidence that in early gestational P. chabaudi AS infection, significant lipid peroxidation and transcriptional antioxidant response, both indicators of oxidative stress, occur in the infected placenta. Whereas NAC was unsuccessful in improving pregnancy outcomes in the mouse, antioxidant TPL therapy yielded a modest and variable, but promising, improvement in pregnancy outcome that may be dually related to reduced oxidative damage and inflammatory responses. These results suggest that antioxidant therapies may be useful as adjunctive therapies with antimalarial drugs in PM, but inconsistent drug efficacy in the placenta is a significant concern.
Supplementary Material
Acknowledgments
We thank MR4 for providing malaria parasites contributed by David Walliker. We thank Dr. Julie Nelson and Dr. Muthugapatti K. Kandasamy, respectively, at the Flow Cytometry Core Facility of the Center for Tropical and Emerging Global Diseases and at the Biomedical Microscopy Core at University of Georgia for technical assistance. We also thank the staff members of the histology laboratory, Department of Pathology, College of Veterinary Medicine at University of Georgia.
This work was supported by the National Institutes of Health Grants HD46860 and AI50240 to J.M.M.
Abbreviations used in this article
- AUC
area under the curve
- Cat
catalase
- Ct
threshold cycle
- ED
experiment day
- HNE
4-hydroxynonenal
- Ho
heme oxygenase
- IHC
immunohistochemistry
- IP
infected pregnant
- NAC
N-acetylcysteine
- Nrf2
NF erythroid derived 2 like-2
- PM
placental malaria
- ROS
reactive oxygen species
- Sod
superoxide dismutase
- TPL
tempol
- UP
uninfected pregnant
Footnotes
The content is solely the responsibility of the authors and does not necessarily represent the official views of the Eunice Kennedy Shriver National Institute of Child Health and Human Development, the National Institute of Allergy and Infectious Diseases, or the National Institutes of Health.
The online version of this article contains supplemental material.
DISCLOSURES
The authors have no financial conflicts of interest.
References
- 1.WHO. World malaria report. WHO Press; Geneva, Switzerland: 2015. [Google Scholar]
- 2.WHO. Intermittent preventive treatment in pregnancy (IPTp) 2016 Available at: http://www.who.int/malaria/areas/preventive_therapies/pregnancy/en/. Accessed: June 1, 2017.
- 3.Avery JW, Smith GM, Owino SO, Sarr D, Nagy T, Mwalimu S, Matthias J, Kelly LF, Poovassery JS, Middii JD, et al. Maternal malaria induces a procoagulant and antifibrinolytic state that is embryotoxic but responsive to anticoagulant therapy. PLoS One. 2012;7:e31090. doi: 10.1371/journal.pone.0031090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Desai M, ter Kuile FO, Nosten F, McGready R, Asamoa K, Brabin B, Newman RD. Epidemiology and burden of malaria in pregnancy. Lancet Infect Dis. 2007;7:93–104. doi: 10.1016/S1473-3099(07)70021-X. [DOI] [PubMed] [Google Scholar]
- 5.Menendez C, Ordi J, Ismail MR, Ventura PJ, Aponte JJ, Kahigwa E, Font F, Alonso PL. The impact of placental malaria on gestational age and birth weight. J Infect Dis. 2000;181:1740–1745. doi: 10.1086/315449. [DOI] [PubMed] [Google Scholar]
- 6.Fried M, Muga RO, Misore AO, Duffy PE. Malaria elicits type 1 cytokines in the human placenta: IFN-gamma and TNF-alpha associated with pregnancy outcomes. J Immunol. 1998;160:2523–2530. [PubMed] [Google Scholar]
- 7.Rogerson SJ, Brown HC, Pollina E, Abrams ET, Tadesse E, Lema VM, Molyneux ME. Placental tumor necrosis factor alpha but not gamma interferon is associated with placental malaria and low birth weight in Malawian women. Infect Immun. 2003;71:267–270. doi: 10.1128/IAI.71.1.267-270.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Megnekou R, Djontu JC, Bigoga JD, Medou FM, Tenou S, Lissom A. Impact of placental Plasmodium falciparum malaria on the profile of some oxidative stress biomarkers in women living in Yaoundé, Cameroon. PLoS One. 2015;10:e0134633. doi: 10.1371/journal.pone.0134633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Sharma L, Kaur J, Shukla G. Role of oxidative stress and apoptosis in the placental pathology of Plasmodium berghei infected mice. PLoS One. 2012;7:e32694. doi: 10.1371/journal.pone.0032694. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Sharma L, Shukla G. Treatment of pregnant BALB/c mice with sulphadoxine pyrimethamine or chloroquine abrogates Plasmodium berghei induced placental pathology. Parasitol Int. 2014;63:49–56. doi: 10.1016/j.parint.2013.08.016. [DOI] [PubMed] [Google Scholar]
- 11.Sharma L, Kaur J, Rishi P, Shukla G. Plasmodium berghei: influence of infection on the oxidant and antioxidants levels in pregnant BALB/c mice. Exp Parasitol. 2012;131:215–222. doi: 10.1016/j.exppara.2012.04.005. [DOI] [PubMed] [Google Scholar]
- 12.Matsubara K, Higaki T, Matsubara Y, Nawa A. Nitric oxide and reactive oxygen species in the pathogenesis of pre-eclampsia. Int J Mol Sci. 2015;16:4600–4614. doi: 10.3390/ijms16034600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Rosta K, Molvarec A, Enzsöly A, Nagy B, Rónai Z, Fekete A, Sasvári-Székely M, Rigó J, Jr, Vér A. Association of extracellular superoxide dismutase (SOD3) Ala40Thr gene polymorphism with pre-eclampsia complicated by severe fetal growth restriction. Eur J Obstet Gynecol Reprod Biol. 2009;142:134–138. doi: 10.1016/j.ejogrb.2008.10.014. [DOI] [PubMed] [Google Scholar]
- 14.Poovassery JS, Sarr D, Smith G, Nagy T, Moore JM. Malaria-induced murine pregnancy failure: distinct roles for IFN-gamma and TNF. J Immunol. 2009;183:5342–5349. doi: 10.4049/jimmunol.0901669. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Poovassery J, Moore JM. Association of malaria-induced murine pregnancy failure with robust peripheral and placental cytokine responses. Infect Immun. 2009;77:4998–5006. doi: 10.1128/IAI.00617-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Sarr D, Smith GM, Poovassery JS, Nagy T, Moore JM. Plasmodium chabaudi AS induces pregnancy loss in association with systemic pro-inflammatory immune responses in A/J and C57BL/6 mice. Parasite Immunol. 2012;34:224–235. doi: 10.1111/j.1365-3024.2012.01355.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Sarr D, Bracken TC, Owino SO, Cooper CA, Smith GM, Nagy T, Moore JM. Differential roles of inflammation and apoptosis in initiation of mid-gestational abortion in malaria-infected C57BL/6 and A/J mice. Placenta. 2015;36:738–749. doi: 10.1016/j.placenta.2015.04.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Poovassery J, Moore JM. Murine malaria infection induces fetal loss associated with accumulation of Plasmodium chabaudi AS-infected erythrocytes in the placenta. Infect Immun. 2006;74:2839–2848. doi: 10.1128/IAI.74.5.2839-2848.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Becker K, Tilley L, Vennerstrom JL, Roberts D, Rogerson S, Ginsburg H. Oxidative stress in malaria parasite-infected erythrocytes: host-parasite interactions. Int J Parasitol. 2004;34:163–189. doi: 10.1016/j.ijpara.2003.09.011. [DOI] [PubMed] [Google Scholar]
- 20.Percário S, Moreira DR, Gomes BA, Ferreira ME, Gonçalves AC, Laurindo PS, Vilhena TC, Dolabela MF, Green MD. Oxidative stress in malaria. Int J Mol Sci. 2012;13:16346–16372. doi: 10.3390/ijms131216346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Bracken TC, Cooper AC, Ali Z, Truong H, Moore MJ. Helicobacter infection significantly alters pregnancy success in laboratory mice. J Am Assoc Lab Anim Sci. 2017;56:322–329. [PMC free article] [PubMed] [Google Scholar]
- 22.Jimenez-Diaz MB, Mulet T, Gomez V, Viera S, Alvarez A, Garuti H, Vazquez Y, Fernandez A, Ibanez J, Jimenez M, et al. Quantitative measurement of Plasmodium-infected erythrocytes in murine models of malaria by flow cytometry using bidimensional assessment of SYTO-16 fluorescence. Cytometry A. 2008;75:225–235. doi: 10.1002/cyto.a.20647. [DOI] [PubMed] [Google Scholar]
- 23.Stanley JL, Andersson IJ, Hirt CJ, Moore L, Dilworth MR, Chade AR, Sibley CP, Davidge ST, Baker PN. Effect of the anti-oxidant tempol on fetal growth in a mouse model of fetal growth restriction. Biol Reprod. 2012;87:25, 1–8. doi: 10.1095/biolreprod.111.096198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Hoffmann DS, Weydert CJ, Lazartigues E, Kutschke WJ, Kienzle MF, Leach JE, Sharma JA, Sharma RV, Davisson RL. Chronic tempol prevents hypertension, proteinuria, and poor feto-placental outcomes in BPH/5 mouse model of preeclampsia. Hypertension. 2008;51:1058–1065. doi: 10.1161/HYPERTENSIONAHA.107.107219. [DOI] [PubMed] [Google Scholar]
- 25.Castro JP, Jung T, Grune T, Siems W. 4-Hydroxy-nonenal (HNE) modified proteins in metabolic diseases. Free Radic Biol Med. 2016 doi: 10.1016/j.freeradbiomed.2016.10.497. [DOI] [PubMed] [Google Scholar]
- 26.Yin H, Xu L, Porter NA. Free radical lipid peroxidation: mechanisms and analysis. Chem Rev. 2011;111:5944–5972. doi: 10.1021/cr200084z. [DOI] [PubMed] [Google Scholar]
- 27.Schmittgen TD, Livak KJ. Analyzing real-time PCR data by the comparative C(T) method. Nat Protoc. 2008;3:1101–1108. doi: 10.1038/nprot.2008.73. [DOI] [PubMed] [Google Scholar]
- 28.Sadowska AM, Verbraecken J, Darquennes K, De Backer WA. Role of N-acetylcysteine in the management of COPD. Int J Chron Obstruct Pulmon Dis. 2006;1:425–434. doi: 10.2147/copd.2006.1.4.425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Sadowska AM, van Overveld FJ, Górecka D, Zdral A, Filewska M, Demkow UA, Luyten C, Saenen E, Zielinski J, De Backer WA. The interrelationship between markers of inflammation and oxidative stress in chronic obstructive pulmonary disease: modulation by inhaled steroids and antioxidant. Respir Med. 2005;99:241–249. doi: 10.1016/j.rmed.2004.07.005. [DOI] [PubMed] [Google Scholar]
- 30.Sadowska AM, Manuel-Y-Keenoy B, De Backer WA. Antioxidant and anti-inflammatory efficacy of NAC in the treatment of COPD: discordant in vitro and in vivo dose-effects: a review. Pulm Pharmacol Ther. 2007;20:9–22. doi: 10.1016/j.pupt.2005.12.007. [DOI] [PubMed] [Google Scholar]
- 31.Sadowska AM, Luyten C, Vints AM, Verbraecken J, Van Ranst D, De Backer WA. Systemic antioxidant defences during acute exacerbation of chronic obstructive pulmonary disease. Respirology. 2006;11:741–747. doi: 10.1111/j.1440-1843.2006.00943.x. [DOI] [PubMed] [Google Scholar]
- 32.Aruoma OI, Halliwell B, Hoey BM, Butler J. The antioxidant action of N-acetylcysteine: its reaction with hydrogen peroxide, hydroxyl radical, superoxide, and hypochlorous acid. Free Radic Biol Med. 1989;6:593–597. doi: 10.1016/0891-5849(89)90066-x. [DOI] [PubMed] [Google Scholar]
- 33.Benrahmoune M, Thérond P, Abedinzadeh Z. The reaction of superoxide radical with N-acetylcysteine. Free Radic Biol Med. 2000;29:775–782. doi: 10.1016/s0891-5849(00)00380-4. [DOI] [PubMed] [Google Scholar]
- 34.Neuschwander-Tetri BA, Rozin T. Diurnal variability of cysteine and glutathione content in the pancreas and liver of the mouse. Comp Biochem Physiol B Biochem Mol Biol. 1996;114:91–95. doi: 10.1016/0305-0491(96)83706-0. [DOI] [PubMed] [Google Scholar]
- 35.McWalter GK, Higgins LG, McLellan LI, Henderson CJ, Song L, Thornalley PJ, Itoh K, Yamamoto M, Hayes JD. Transcription factor Nrf2 is essential for induction of NAD(P)H: quinone oxidoreductase 1, glutathione S-transferases, and glutamate cysteine ligase by broccoli seeds and isothiocyanates. J Nutr. 2004;134(Suppl):3499S–3506S. doi: 10.1093/jn/134.12.3499S. [DOI] [PubMed] [Google Scholar]
- 36.Xu DX, Chen YH, Wang H, Zhao L, Wang JP, Wei W. Effect of N-acetylcysteine on lipopolysaccharide-induced intrauterine fetal death and intra-uterine growth retardation in mice. Toxicol Sci. 2005;88:525–533. doi: 10.1093/toxsci/kfi300. [DOI] [PubMed] [Google Scholar]
- 37.Wilcox CS. Effects of tempol and redox-cycling nitroxides in models of oxidative stress. Pharmacol Ther. 2010;126:119–145. doi: 10.1016/j.pharmthera.2010.01.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Wilcox CS, Pearlman A. Chemistry and antihypertensive effects of tempol and other nitroxides. Pharmacol Rev. 2008;60:418–469. doi: 10.1124/pr.108.000240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Laight DW, Andrews TJ, Haj-Yehia AI, Carrier MJ, Anggård EE. Microassay of superoxide anion scavenging activity in vitro. Environ Toxicol Pharmacol. 1997;3:65–68. doi: 10.1016/s1382-6689(96)00143-3. [DOI] [PubMed] [Google Scholar]
- 40.Serghides L, McDonald CR, Lu Z, Friedel M, Cui C, Ho KT, Mount HT, Sled JG, Kain KC. PPARγ agonists improve survival and neurocognitive outcomes in experimental cerebral malaria and induce neuroprotective pathways in human malaria. PLoS Pathog. 2014;10:e1003980. doi: 10.1371/journal.ppat.1003980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Nozik-Grayck E, Suliman HB, Piantadosi CA. Extracellular superoxide dismutase. Int J Biochem Cell Biol. 2005;37:2466–2471. doi: 10.1016/j.biocel.2005.06.012. [DOI] [PubMed] [Google Scholar]
- 42.Debierre-Grockiego F, Schwarz RT. Immunological reactions in response to apicomplexan glycosylphosphatidylinositols. Glycobiology. 2010;20:801–811. doi: 10.1093/glycob/cwq038. [DOI] [PubMed] [Google Scholar]
- 43.Niessen F, Schaffner F, Furlan-Freguia C, Pawlinski R, Bhattacharjee G, Chun J, Derian CK, Andrade-Gordon P, Rosen H, Ruf W. Dendritic cell PAR1-S1P3 signalling couples coagulation and inflammation. Nature. 2008;452:654–658. doi: 10.1038/nature06663. [DOI] [PubMed] [Google Scholar]
- 44.Schwarzer E, Arese P, Skorokhod OA. Role of the lip-operoxidation product 4-hydroxynonenal in the pathogenesis of severe malaria anemia and malaria immunodepression. Oxid Med Cell Longev. 2015;2015:638416. doi: 10.1155/2015/638416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Barrera G. Oxidative stress and lipid peroxidation products in cancer progression and therapy. ISRN Oncol. 2012;2012:137289. doi: 10.5402/2012/137289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Dimasuay KG, Aitken EH, Rosario F, Njie M, Glazier J, Rogerson SJ, Fowkes FJ, Beeson JG, Powell T, Jansson T, Boeuf P. Inhibition of placental mTOR signaling provides a link between placental malaria and reduced birthweight. BMC Med. 2017;15:1. doi: 10.1186/s12916-016-0759-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Steketee RW, Nahlen BL, Parise ME, Menendez C. The burden of malaria in pregnancy in malaria-endemic areas. Am J Trop Med Hyg. 2001;64(Suppl):28–35. doi: 10.4269/ajtmh.2001.64.28. [DOI] [PubMed] [Google Scholar]
- 48.Umbers AJ, Aitken EH, Rogerson SJ. Malaria in pregnancy: small babies, big problem. Trends Parasitol. 2011;27:168–175. doi: 10.1016/j.pt.2011.01.007. [DOI] [PubMed] [Google Scholar]
- 49.Karowicz-Bilińska A. Lipid peroxides concentration in women with intrauterine growth restriction. Ginekol Pol. 2004;75:6–9. [PubMed] [Google Scholar]
- 50.Jauniaux E, Burton GJ. The role of oxidative stress in placental-related diseases of pregnancy. J Gynecol Obstet Biol Reprod. 2016;45:775–785. doi: 10.1016/j.jgyn.2016.02.012. [DOI] [PubMed] [Google Scholar]
- 51.Cotechini T, Graham CH. Aberrant maternal inflammation as a cause of pregnancy complications: a potential therapeutic target? Placenta. 2015;36:960–966. doi: 10.1016/j.placenta.2015.05.016. [DOI] [PubMed] [Google Scholar]
- 52.Agarwal A, Aponte-Mellado A, Premkumar BJ, Shaman A, Gupta S. The effects of oxidative stress on female reproduction: a review. Reprod Biol Endocrinol. 2012;10:49. doi: 10.1186/1477-7827-10-49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Lisitsyna TA, Reshetnik TM, Durnev AD, Nasonov EL. Oxidative stress in the pathogenesis of anti-phospholipid syndrome. Vestn Akad Med Nauk SSSR. 2004;7:19–24. [PubMed] [Google Scholar]
- 54.Paintlia MK, Paintlia AS, Singh AK, Singh I. Attenuation of lipopolysaccharide-induced inflammatory response and phospholipids metabolism at the feto-maternal interface by N-acetyl cysteine. Pediatr Res. 2008;64:334–339. doi: 10.1203/PDR.0b013e318181e07c. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Cuzzocrea S, McDonald MC, Mota-Filipe H, Mazzon E, Costantino G, Britti D, Mazzullo G, Caputi AP, Thiemermann C. Beneficial effects of tempol, a membrane-permeable radical scavenger, in a rodent model of collagen-induced arthritis. Arthritis Rheum. 2000;43:320–328. doi: 10.1002/1529-0131(200002)43:2<320::AID-ANR11>3.0.CO;2-9. [DOI] [PubMed] [Google Scholar]
- 56.Cuzzocrea S, McDonald MC, Mazzon E, Siriwardena D, Costantino G, Fulia F, Cucinotta G, Gitto E, Cordaro S, Barberi I, et al. Effects of tempol, a membrane-permeable radical scavenger, in a gerbil model of brain injury. Brain Res. 2000;875:96–106. doi: 10.1016/s0006-8993(00)02582-8. [DOI] [PubMed] [Google Scholar]
- 57.Cuzzocrea S, McDonald MC, Mazzon E, Dugo L, Lepore V, Fonti MT, Ciccolo A, Terranova ML, Caputi AP, Thiemermann C. Tempol, a membrane-permeable radical scavenger, reduces dinitrobenzene sulfonic acid-induced colitis. Eur J Pharmacol. 2000;406:127–137. doi: 10.1016/s0014-2999(00)00623-3. [DOI] [PubMed] [Google Scholar]
- 58.Cuzzocrea S, McDonald MC, Filipe HM, Costantino G, Mazzon E, Santagati S, Caputi AP, Thiemermann C. Effects of tempol, a membrane-permeable radical scavenger, in a rodent model of carrageenan-induced pleurisy. Eur J Pharmacol. 2000;390:209–222. doi: 10.1016/s0014-2999(99)00910-3. [DOI] [PubMed] [Google Scholar]
- 59.Francischetti IM, Gordon E, Bizzarro B, Gera N, Andrade BB, Oliveira F, Ma D, Assumpção TC, Ribeiro JM, Pena M, et al. Tempol, an intracellular antioxidant, inhibits tissue factor expression, attenuates dendritic cell function, and is partially protective in a murine model of cerebral malaria. PLoS One. 2014;9:e87140. doi: 10.1371/journal.pone.0087140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Ebenezer PJ, Mariappan N, Elks CM, Haque M, Francis J. Diet-induced renal changes in Zucker rats are ameliorated by the superoxide dismutase mimetic TEMPOL. Obesity (Silver Spring) 2009;17:1994–2002. doi: 10.1038/oby.2009.137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Mitchell JB, Xavier S, DeLuca AM, Sowers AL, Cook JA, Krishna MC, Hahn SM, Russo A. A low molecular weight antioxidant decreases weight and lowers tumor incidence. Free Radic Biol Med. 2003;34:93–102. doi: 10.1016/s0891-5849(02)01193-0. [DOI] [PubMed] [Google Scholar]
- 62.Schubert R, Erker L, Barlow C, Yakushiji H, Larson D, Russo A, Mitchell JB, Wynshaw-Boris A. Cancer chemoprevention by the antioxidant tempol in Atm-deficient mice. Hum Mol Genet. 2004;13:1793–1802. doi: 10.1093/hmg/ddh189. [DOI] [PubMed] [Google Scholar]
- 63.Uyoga S, Skorokhod OA, Opiyo M, Orori EN, Williams TN, Arese P, Schwarzer E. Transfer of 4-hydroxynonenal from parasitized to non-parasitized erythrocytes in rosettes. Proposed role in severe malaria anemia. Br J Haematol. 2012;157:116–124. doi: 10.1111/j.1365-2141.2011.09015.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Lucchi NW, Sarr D, Owino SO, Mwalimu SM, Peterson DS, Moore JM. Natural hemozoin stimulates syncytiotrophoblast to secrete chemokines and recruit peripheral blood mononuclear cells. Placenta. 2011;32:579–585. doi: 10.1016/j.placenta.2011.05.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Volk T, Hensel M, Schuster H, Kox WJ. Secretion of MCP-1 and IL-6 by cytokine stimulated production of reactive oxygen species in endothelial cells. Mol Cell Biochem. 2000;206:105–112. doi: 10.1023/a:1007059616914. [DOI] [PubMed] [Google Scholar]
- 66.Di Paola R, Mazzon E, Zito D, Maiere D, Britti D, Genovese T, Cuzzocrea S. Effects of Tempol, a membrane-permeable radical scavenger, in a rodent model periodontitis. J Clin Periodontol. 2005;32:1062–1068. doi: 10.1111/j.1600-051X.2005.00818.x. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.