


Abstract
CENP-C is an evolutionarily conserved centromere protein that is thought to be an important component in kinetochore assembly in vertebrate cells. However, the functional role of CENP-C in cell cycle progression remains unclear. To further understand CENP-C function, we developed a method incorporating the hyper-recombinogenic chicken B lymphocyte cell line DT40 to create several temperature-sensitive CENP-C mutants in DT40 cells. We found that, under restrictive conditions, one temperature-sensitive mutant, ts4-11, displayed metaphase delay and chromosome missegregation but proceeded through the cell cycle until arrest at G1 phase. Furthermore, ts4-11 cells were transfected with a human HeLa cell cDNA library maintained in a retroviral vector, and genes that suppressed the temperature-sensitive phenotype were identified. One of these suppressor genes encodes SUMO-1, which is a ubiquitin-like protein. This finding suggests that SUMO-1 may be involved in centromere function in vertebrate cells. The novel strategy reported here will be useful and applicable to a wide range of proteins that have general cell-autonomous function in vertebrate cells.
INTRODUCTION
The centromere plays a fundamental role in accurate chromosome segregation during mitosis and meiosis in eukaryotes. Its functions include sister chromatid adhesion and separation, microtubule attachment, chromosome movement and mitotic checkpoint control (1). Although errors in chromosome segregation are known to cause genetic diseases including some cancers (2), the mechanism by which functional centromeres are assembled and interact with microtubules of the spindle apparatus during cell division is not fully understood. Many mutants that show errors in chromosome segregation have been characterized to clarify the function of the centromere. This genetic approach has been applied extensively in yeasts and has led to the identification of genes that encode centromere components including CBF1, CBF3, NDC10, CEP3, CSE4 and MIF2 (3–7). Several genes involved in the evolutionarily conserved mitotic checkpoint pathway (8) have been identified through analysis of Saccharomyces cerevisiae mutants. Vertebrate homologs of these checkpoint proteins localize to the centromere prior to association of the centromere with the spindle (9–11).
It has been difficult to use genetic methods to analyze vertebrate centromeres because suitable assays for mutations that affect the accuracy of chromosome segregation are not currently available. Instead, vertebrate centromere proteins have been studied primarily by immunological methods. Antibodies against centromere proteins have been isolated from patients with various autoimmune diseases (12) and from immunized animals (13), and this has led to the identification of several proteins known as CENPs (14–18). Genetic approaches to analysis of vertebrate centromeres are desirable, however. Creation of temperature-sensitive mutants for genes of interest, for example, would provide a means by which we could know the function of these genes. Screening available libraries for genes that suppress the mutant phenotypes would help us identify interacting proteins. Thus, we were led to develop a genetic analysis method that utilizes the hyper-recombinogenic chicken B lymphocyte cell line DT40. We attempted to generate several temperature-sensitive CENP-C mutants in DT40 cells. Vertebrate CENP-C has been localized to the inner kinetochore plates adjacent to the centromeric DNA (19), and it is known to bind DNA directly (20). Disruption of the CENP-C gene results in cell death (21,22) due to chromosome missegregation and metaphase arrest (23). Metaphase arrest has also been observed after microinjection of anti-CENP-C antibodies into HeLa cells (24). Like CENP-A, CENP-C is found only at active centromeres (25,26) and is needed to induce formation of a functional centromere (23), suggesting that CENP-C is an important component of the kinetochore assembly. Here, we report the creation of several temperature-sensitive CENP-C mutants. We show that, at the restrictive temperature, mutant cells display metaphase delay and chromosome missegregation and that they proceed through the cell cycle but arrest at G1. In addition, by introducing a human HeLa cell cDNA library into mutant cells, we identified genes that suppress the temperature-sensitive phenotype.
MATERIALS AND METHODS
Plasmids
All plasmids were constructed by standard methods. The CENP-C targeting constructs that contain either the neomycin or puromycin resistance gene were described previously (21). To make the mutant CENP-C constructs, we used the QuickChange kit (Stratagene) according to the manufacturer’s specifications. CENP-C mutations were generated as follows (5′ to 3′ from the start site of the chicken CENP-C cDNA; mutant bases are underlined): E-671K, GAG→AAG; P-825L, CCA→CTA; R-655P, CGT→CCA; R-659W, AGG→TGG; P-664A, CCT→GCT; L-665V, CTG→GTG; Y-667F, TAT→TTT; W-668R, TGG→CGG; R-669G, CGA→GGA; and Y-675N, TAT→AAT. Mutant CENP-C fragments were inserted into the SmaI site of a CENP-C dicistronic construct containing the 5′ upstream region that is thought to include the CENP-C promoter (21). All gene constructs were verified by sequencing with the BigDye terminator sequencing kit (ABI) and a 377 ABI-DNA sequencer.
Cell culture and transfection
DT40 cells were cultured and transfected as described previously (21,23,27). Parental DT40 cells and all subclones were cultured in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% fetal calf serum, 1% chicken serum, penicillin and streptomycin at 37°C. G418 (Sigma) was added to a final concentration of 2 mg/ml, puromycin (Clontech) was used at a final concentration of 0.5 µg/ml, and histidinol (Sigma) was used at a final concentration of 1 mg/ml to select for stable transfectants. We used 96-well plates for drug selection. In each step, we picked a single colony from each well. We usually obtained 10–20 colonies per 96-well plate; therefore, we assumed that each single colony was derived from a single cell.
To generate a high-titer virus solution, ϕMP34 cells (28), a packaging cell line, were cultured in DMEM supplemented with 10% fetal calf serum, penicillin and streptomycin at 37°C. ϕMP34 cells were transfected with a human cDNA library with the CalPhos Mammalian Cell Transfection Kit (Clontech). This library was constructed in a pLiB vector (Clontech) with mRNA from HeLa cells, and the average cDNA insert size in this library is 2.4 kb. After cultures were incubated for 48–72 h, the culture medium was collected and centrifuged at 6000 g for 16 h at 4°C. After centrifugation, the supernatant was removed and the pellet was resuspended in an appropriate volume of cell culture medium. This high-titer viral suspension was passed gently through an 18G needle. To facilitate infection of DT40 cells, polybrene (Sigma) was added to the virus suspension to a final concentration of 4 µg/ml. To infect DT40 cells, virus supernatant containing polybrene was added to 1 × 107 DT40 cells in 50 ml culture medium. The virus was exposed to targeted cells for 6 h, and the culture medium was replaced with fresh medium. After incubation at 34°C for 24 h, the cells were incubated at 43°C. After 7 days, we selected colonies and performed PCR with the vector-specific primers for recovery of suppressor genes. Candidate suppressor genes were re-cloned into the EcoRI site of the pLib vector and were again introduced into temperature-sensitive mutant cells to confirm the suppressor activity of the gene.
Cell cycle analysis
Cells were pulse-labeled with 20 µM 5-bromodeoxyuridine (BrdU; Roche) for 20 min at the times indicated in Figure 5. Cells were harvested and fixed at 4°C overnight with 70% ethanol. After treatment with 4 M HCl in 0.5% Triton X-100 for 30 min, cells were incubated with anti-BrdU primary antibody (Becton Dickinson) for 1 h at room temperature and then with FITC-conjugated anti-mouse IgG (Jackson ImmunoResearch Laboratories) for 30 min at room temperature. Cells were stained with 5 µg/ml propidium iodide (Sigma) in phosphate-buffered saline (PBS). Between incubations, cells were washed with PBS containing 1% bovine serum albumin (BSA). Subsequent flow-cytometry was performed with an Epics Altra cytometer (Beckman-Coulter). Fluorescence data were displayed as dot plots produced by the Altra analysis software (Beckman-Coulter).
Figure 5.


Cell cycle progression of a temperature-sensitive CENP-C mutant. (A) Cell-cycle distribution of ts4-11 cells following culture at 43°C. Cells were stained with FITC-anti-BrdU (y-axis, log scale) to detect BrdU incorporation and with propidium iodide to detect total DNA (x-axis, linear scale). The lower-left box represents G1 phase cells, the upper box represents S phase cells, and the lower-right box represents G2/M phase cells. The numbers given in the boxes indicate the percentage of gated events. (B) Cell cycle progression of synchronized cultures of DT40 (left) or ts4-11 (right) cells. Both cell lines were cultured at 43°C. Cells were synchronized at mitosis with nocodazole. After release of the block, we measured DNA content by FACS at 2-h intervals.
For synchronization of cells, nocodazole (Sigma) was added to cultures at final concentration of 500 ng/ml. After incubation for 7 h at 43°C, cells were released from the nocodazole block by three washes in medium and re-cultured in fresh medium. At various time points after release, cells were harvested and analyzed with the Epics Altra cytometer (Beckman-Coulter).
Immunocytochemistry and FISH analysis
Immunofluorescent staining of whole cells was performed as described previously (21,23). Cells were collected onto slides by cytocentrifugation and fixed in 3% paraformaldehyde in PBS for 15 min at room temperature, permeabilized in 0.5% NP-40 in PBS for 15 min at room temperature, rinsed three times in 0.5% BSA and incubated for 1 h at 37°C with anti-α-tubulin antibody (1:50; Sigma), rabbit anti-CENP-A antibody (1:50; V.Regnier, unpublished data), rabbit anti-CENP-C antibody (1:1000; 23) or rabbit anti-CENP-H antibody (1:2000; 29). For double-staining experiments with CENP-A and CENP-C or CENP-C and CENP-H, we used the Fab fragment. After incubation with the primary antibody, we incubated cells with an excess of FITC-conjugated Fab antibody against rabbit IgG (Jackson ImmunoResearch Laboratories). We then labeled the second antigen as usual and detected the signals with Cy3-conjugated goat anti-rabbit IgG (Jackson ImmunoResearch Laboratories).
After cells were treated with colcemid at a final concentration of 100 ng/ml for 2 h, metaphase chromosome spreads were prepared by addition of a hypotonic solution and fixation in methanol/acetic acid (3:1). FISH was performed as described previously (30). Biotin-labeled painting probes were generated by degenerate-oligo-primed-PCR as described previously (30). Chromosomes and nuclei were counterstained with DAPI at 0.2 µg/ml in Vectashield antifade (Vector Labs). For chromosome stability assay, the numbers of chromosomes 1 and 2 (five in normal cells) were scored in at least 200 metaphase cells at the indicated time points.
All immunofluorescence and FISH images were obtained with a cooled CCD camera (Photometrics Image Point) mounted on a Zeiss Axioscope microscope with a 63× objective together with a filter wheel. Images were manipulated with IPLab software (Signal Analytics).
RESULTS
Generation of temperature-sensitive CENP-C mutants
We observed that DT40 cells were able to grow between 34 and 43°C. We then tried to generate temperature-sensitive mutants at the permissive temperature of 34°C and at the restrictive temperature of 43°C. The procedure for generating temperature-sensitive CENP-C mutants is shown in Figure 1A. We used the CENP-C-targeting constructs containing either the neomycin (neo) or puromycin (puro) resistance gene (21). First, we transfected the neo-targeting construct into DT40 cells and isolated neo-resistant clones heterozygous for the CENP-C+/– gene. Then, the CENP-C+/– clone was transfected with a mutant CENP-C cDNA under the control of the native CENP-C promoter and that integrated into the random sites in the genome of DT40 cells (CENP-C+/–/mutant CENP-C) at 34°C. To introduce mutations in CENP-C cDNA, we used two short regions homologous between CENP-Cs of several species (Fig. 1B). We used site-directed mutagenesis to generate 10 mutant CENP-C cDNA constructs with changes in conserved amino acids. Two of the mutations we generated were identical to the MIF2 mutations described in S.cerevisiae (31). The 1-19 mutation in CENP-C is identical to the MIF2-5 mutation (E→K), and the 2-15 mutation in CENP-C is identical to the MIF2-3 mutation (P→L). The other eight mutant cDNAs were random. We then transfected the puro-targeting construct into 10 different CENP-C+/–/mutant CENP-C clones to isolate CENP-C–/–/mutant CENP-C at 34°C. Targeted clones were moved to 43°C medium. At 43°C, temperature-sensitive mutants showed a severe phenotype that is described below. We isolated four temperature-sensitive mutants. The amino acid substitutions in CENP-C are shown in Figure 1B. The 3-17 and 4-11 mutations have not been reported in S.cerevisiae. We isolated mRNA from these temperature-sensitive mutants and confirmed the amino acid substitutions by RT–PCR and sequencing. Sequencing data for the 4-11 mutation, which causes the R659W substitution, are shown in Figure 1C. We characterized the 4-11 mutant (ts4-11) for further study. We should note that the four mutations obtained showed temperature-sensitivity even when the cells were heterozygous for the CENP-C mutation.
Figure 1.

Creation of temperature-sensitive CENP-C mutants. (A) Experimental strategy for isolation of temperature-sensitive mutant clones. (B) Positions of mutations in temperature-sensitive CENP-C mutants. Cells were transfected with 10 different constructs as described in the Materials and Methods. The four mutations shown here displayed temperature sensitivity. (C) Sequencing analysis of an RT–PCR product from the temperature-sensitive CENP-C mutant ts4-11.
The proliferative properties of ts4-11 cells were monitored with growth curves. Viable cells were assessed by trypan blue exclusion. The growth curve of ts4-11 cells was similar to that of wild-type DT40 cells at 34°C (Fig. 2). When ts4-11 cells were incubated at 43°C, cells ceased proliferating at ∼12 h and no cells survived to 60 h. Wild-type DT40 cells proliferated normally at 43°C (Fig. 2). These data indicate that the point mutation in the CENP-C gene results in growth arrest and subsequent cell death at 43°C.
Figure 2.

A temperature-sensitive CENP-C mutant displays growth arrest at 43°C. Representative growth curves for the indicated cell cultures. Cell viability was determined by trypan blue exclusion. Each experiment was performed twice, and each time point was examined in duplicate. When ts4-11 cells were incubated at 43°C, cells ceased proliferating by 12 h, and no viable cells were observed at 60 h. Wild-type DT40 cells proliferated normally at 43°C.
We examined localization of the mutant CENP-C protein in ts4-11 cells incubated at 43°C. Wild-type CENP-C is localized to the centromere throughout the cell cycle (19,23). Like CENP-C, CENP-A and CENP-H are constitutive centromere components present throughout the cell cycle (29,32). We then performed double-staining of ts4-11 cells with anti-CENP-C and anti-CENP-A or with anti-CENP-C and anti-CENP-H under restrictive conditions. Mutant CENP-C was co-localized with both CENP-A and CENP-H in ts4-11 cells after incubation at 43°C for 24 h (Fig. 3), indicating that the mutant CENP-C was not degraded and its localization to the centromere was not altered. The state (intact versus degraded) of mutant CENP-C was confirmed by western blot analysis (Fig. 3C). Western analysis confirmed that mutant CENP-C was not over-expressed (Fig. 3C).
Figure 3.

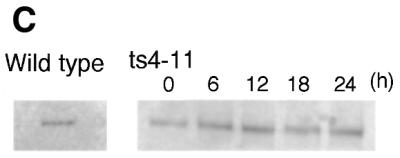
Localization of mutant CENP-C to the centromere is not changed under restrictive conditions. (A) Immunofluorescence analysis of ts4-11 cells stained with both anti-CENP-C and anti-CENP-H antibodies after 24 h at 43°C. The CENP-H signal is red, and the CENP-C signal is green. DNA was counterstained with DAPI (blue). (B) Immunofluorescence analysis of ts4-11 cells stained with both anti-CENP-A and anti-CENP-C antibodies after 24 h at 43°C. CENP-A signal is red, and CENP-C signal is green. DNA was counterstained with DAPI (blue). (C) Western blot analysis with anti-CENP-C antibody of ts4-11 cell extracts at the indicated times following incubation at 43°C. We also analyzed wild-type DT40 cells.
The phenotype of a temperature-sensitive CENP-C mutant
We examined whether the temperature-sensitive CENP-C mutation in ts4-11 cells causes chromosome missegregation. We performed FISH analysis of metaphase spreads with painting probes specific for chicken chromosomes 1 and 2. Because DT40 cells contain three copies of chromosome 2, we observed five fluorescent chromosomes in each wild-type cell (Fig. 4A, left panel). As shown in Figure 4A (right panel) and B, chromosome losses were observed in ts4-11 cells at the restrictive temperature. When cells were incubated at 43°C for 24 h, 3.4% of cells lost at least one copy of chromosome 1 or 2, a rate three times higher than the rate of loss observed in ts4-11 cells cultured at 34°C (Fig. 4B). This indicates that the temperature-sensitive CENP-C mutation in ts4-11 cells induces chromosome missegregation.
Figure 4.

A temperature-sensitive CENP-C mutant displays chromosome missegregation under restrictive conditions. (A) To examine chromosome loss, we used FISH analysis with chromosome-specific painting probes. We used probes specific for chicken chromosomes 1 and 2. Because DT40 has three copies of chromosome 2, we observed five painted chromosomes in ts4-11 cells at the permissive temperature (left panel). ts4-11 cells showing loss of chromosomes (right panel) were detected after incubation of cells at 43°C. (B) Distribution of the number of painted chromosomes per cell. ts4-11 cells were cultured at 43°C for 24 h. Wild-type DT40 cells were cultured at 43°C as a control. The numbers of painted chromosomes 1 and 2 were scored in approximately 200 metaphase cells. After colcemid treatment, we observed that ∼50% of cells were in metaphase in control cultures and ∼25% of cells were in metaphase after 24 h at 43°C.
Although we observed a significant increase in chromosome missegregation in ts4-11 cells at the restrictive temperature, it was unclear at what stage the mutant cells arrested during the cell cycle. To address this question, we measured both cellular DNA content and DNA synthesis by fluorescence-activated cell sorting (FACS) after pulse-BrdU-labeling (Fig. 5A). Asynchronous wild-type DT40 and ts4-11 cells (incubated at 34°C) showed similar cell cycle patterns (data not shown), with S phase accounting for two-thirds of the entire cell cycle time (Fig. 5A, 0 h). In contrast, after ts4-11 cells were cultured at 43°C, BrdU uptake decreased and was not detectable by 48 h. Thirty-nine percent of cells had accumulated in G2/M phase by 8 h (Fig. 5A). We also stained mutant cells with anti-tubulin and confirmed that 35% of the cells were arrested in metaphase. After metaphase arrest of the mutant cells at the restrictive temperature, cells proceeded further through mitosis and chromosome missegregation occurred. Cells finally arrested in G1. By 24 h, 52% of cells were arrested in G1 (Fig. 5A, 24 h). We also examined cell cycle progression of ts4-11 cells using synchronized cultures (Fig. 5B and C). After cells were synchronized with nocodazole for 7 h, the block was removed, and cell cycle progression was investigated at 2 h intervals under restrictive conditions. Comparison of the cell cycle patterns of wild-type and mutant cells at 2 h intervals revealed mitotic delay of ts4-11 cells (Fig. 5B and C, 2 h). After mitotic delay, the mutant cells exited mitosis, arrested at G1 and then died.
Identification of a gene that suppresses the temperature-sensitive phenotype
We used our temperature-sensitive CENP-C mutant to identify gene products that interact genetically with CENP-C. For this purpose, we used a full-length human cDNA library constructed in a retroviral vector. The experimental procedure for isolation of genes that suppress the temperature-sensitive phenotype of ts4-11 cells is shown in Figure 6A. To obtain high-titer retroviruses, we used a modified packaging cell line, ϕMP34 (28). Packaging cells were transfected with library cDNAs, and the viral supernatants were harvested. Viral supernatants were enriched by centrifugation. ts4-11 cells were infected with the high-titer virus solution and cultured at 43°C. Cells that express a suppressor gene can grow at 43°C. Suppressor genes were then recovered from ts4-11 cells viable at 43°C by PCR with vector-specific primers. We also confirmed that each candidate suppressor was a legitimate suppressor gene with an additional transfection and selection experiment. DNA sequences of several suppressor genes were determined. Using this strategy, we identified several genes encoding suppressors of the temperature-sensitive CENP-C mutant (Table 1). One suppressor gene is SUMO-1 (Fig. 6B). SUMO-1, a ubiquitin-like protein, covalently modifies target proteins in a reaction mediated by the E2 ligase Ubc9; however, the biological significance of SUMO-1 modification (sumoylation) is unclear (33). The S.cerevisiae homolog of SUMO-1, SMT3Ap, can suppress the temperature-sensitive MIF2 mutant phenotype (36). MIF2p is the yeast functional homolog of CENP-C. This result suggests that the centromere assembly pathway is conserved. We attempted to assess whether CENP-C is modified directly by SUMO-1 but could not detect sumoylation of CENP-C (data not shown). We also generated Ubc9-deficient DT40 cells in which sumoylation does not occur (T.Hayashi, M.Seki, T.Fukagawa and T.Enomoto, unpublished data) and found abnormal progression of mitosis in Ubc9-deficient cells, suggesting that SUMO-1 may be related to mitotic function. The suppressor genes identified in the present study are summarized in Table 1. Further studies of these proteins will contribute to our understanding of centromere assembly and function. We also expressed the genes encoding CENP-A or CENP-H in ts4-11 cells by the retroviral vector system. Expression of these genes did not suppress the temperature-sensitive CENP-C phenotype (Fig. 6B).
Figure 6.


Identification of genes that suppress the temperature-sensitive CENP-C mutant phenotype. (A) Experimental strategy for isolation of genes that suppress the temperature-sensitive CENP-C mutant phenotype. (B) The temperature-sensitive phenotype was suppressed by introduction of the human SUMO-1 but not human CENP-A or chicken CENP-H gene.
Table 1. Suppressor genes for the CENP-C temperature-sensitive phenotype.
| No. of isolates |
Gene |
Accession no. |
| 5 | SUMO-1 | U67122 |
| 1 | Unkown protein (weak similarity to CENP-A) | AAH07864 |
| 1 | Kid (kinesin-like 4; essential for metaphase chromosome alignment) | NP_015556 |
| 2 | Ribosomal protein L23 | AAA03341 |
| 2 | T-complex protein, epsilon (molecular chaperone; M phase-specific interacting with 26S proteosome) | P48643 |
| 1 | Nudix (nucleoside diphosphate-linked moiety; MutT-like) | XP_0057960 |
| 1 | Ubiquitin-specific proteinase 16 | NP_006438 |
DISCUSSION
Creation of temperature-sensitive CENP-C mutants in DT40
To characterize the in vivo functions of a protein, it is helpful to identify temperature-sensitive mutations that can be studied in cultured cells. Significant efforts have been made to isolate mutations in genes from cultured vertebrate cells, but most attempts have been unsuccessful (34). In yeast, plasmid shuffling combined with in vitro mutagenesis has been useful for isolating conditional mutations in genes of interest (34). In the present study, we showed that temperature-sensitive CENP-C mutants could be created in the hyper-recombinogenic chicken cell line DT40. In DT40 cells, homologous recombination occurs very efficiently; the ratio of random to targeted integrations is 1:1 (27). To generate temperature-sensitive mutations, we focused on conserved regions of the CENP-C gene designated Blocks 2 and 3. In these regions, there are 24 amino acids conserved between the CENP-Cs of five species. We chose to change 10 of these amino acids. Of these 10 mutations, four yielded temperature-sensitive mutants. Although we generated temperature-sensitive mutants efficiently by site-directed mutagenesis, it may be possible to use random mutagenesis to generate additional temperature-sensitive mutations.
CENP-C function during the cell cycle
The phenotype of temperature-sensitive CENP-C mutants under restrictive conditions provided clues of the function of CENP-C during the cell cycle. At the restrictive temperature, ts4-11 cells were arrested at G1 phase after mitotic delay. There are two possible explanations for this arrest. One is that CENP-C functions mainly during mitosis. ts4-11 cells may have partial centromere function and thus be able to proceed through the cell cycle after mitotic delay. Chromosome loss appears to occur during progression, and cells may lose genes necessary for completion of G1 or entrance into S phase. We could not rule out this possibility because the mutant CENP-C protein was localized to centromeres. However, it is also possible that CENP-C has dual function during G1 and mitosis. Knehr et al. (35) examined cell cycle-dependent expression of human CENP-C. They reported that the highest expression of CENP-C occurs during G1 phase, and they postulated that, in addition to its role in mitosis, CENP-C might have a further role in G1 that is related to cell cycle control. Our observation of G1 arrest in ts4-11 cells is consistent with their data and hypothesis. We previously generated the Δ/CENP-C-ER DT40 cell line, which is a conditional loss-of-function mutant of CENP-C (21). In the presence of 4-hydroxytamoxifen (OHT), these cells proliferate similar to wild-type cells, but they lose CENP-C function when OHT is removed from the culture medium. Although we detected an accumulation of metaphase cells among the CENP-C-deficient cells, only 10% of cells were in metaphase in asynchronous cultures. One explanation for this observation was that CENP-C might serve further functions during the cell cycle in addition to its role in mitosis. The phenotypes of temperature-sensitive and CENP-C-deficient mutants suggest that CENP-C may have multiple functions during interphase and mitosis. Several proteins including CENP-A, -C and -H are thought to form a ‘prekinetochore structure’ prior to mitosis. During G1, the prekinetochore structure may be established, and abnormal structures may lead to cell cycle arrest. If CENP-C functions during G1 phase, it may be involved in prekinetochore assembly at interphase, and the prekinetochore structure may be related to the G1 checkpoint.
Identification of genes that suppress the temperature-sensitive CENP-C mutant phenotype
In yeast, multi-copy genes encoding suppressors of temperature-sensitive phenotypes were identified to investigate genetic interactions with gene products of interest. Because it is difficult to use plasmid vectors in vertebrate cells, we developed a genetic method using a retroviral vector system to further our understanding of CENP-C function. Retroviral vectors are advantageous because there is no damage to cells as a result of gene transfer, and the transferred gene integrates into the host genome and shows steady, elevated expression. Recently, Yoshimatsu et al. (28) developed a new method to obtain high-titer recombinant retroviruses that is based on a modified packaging cell line and virus enrichment by centrifugation. We applied their method to our temperature-sensitive CENP-C mutants and successfully isolated genes that suppressed the temperature-sensitive phenotype.
One suppressor of the CENP-C temperature-sensitive phenotype is SUMO-1. The S.cerevisiae homolog of SUMO-1, SMT3Ap, suppresses the phenotype of a temperature-sensitive MIF2 mutation (36). Though this connection is interesting, neither CENP-C nor MIF2p is modified directly by SUMO-1. The present data and data from recent reports (33,37,38) suggest SUMO-1 to be involved indirectly in centromere function. Ishov et al. (33) performed immunostaining with anti-SUMO-1 antibody and showed that SUMO-1 is localized to ND10 bodies. These bodies are punctate foci in interphase nuclei that contain several proteins including PML, Sp100 and Daxx. The function of ND10 has not yet been determined, but Everett et al. (38) reported a connection between centromeres and ND10 protein. In addition, human CENP-C interacts with Daxx, a component of ND10, as determined by yeast two-hybrid assays (37). These findings suggest that there is a cell-cycle regulated connection between CENP-C and ND10 proteins modified by SUMO-1. Our identification of SUMO-1 as a suppressor of the CENP-C mutant phenotype suggests a genetic interaction between CENP-C and SUMO-1. We recently created a Ubc9-deficient DT40 cell line that showed abnormal mitosis, indicating that modification of proteins by SUMO-1 may be associated with mitosis (T.Hayashi, M.Seki, T.Fukagawa and T.Enomoto, unpublished data). Further analysis of ND10-related proteins may reveal the functional connection between the centromere, sumoylation and ND10.
We also identified several additional suppressor, which encode kid, ribosomal protein L23, T-complex protein ɛ, nudix and ubiquitin proteinase. Although we do not know of any direct associations of these gene products with CENP-C, several are known to contribute to mitotic function. Chromokinesin, kid, is essential for chromosome alignment and anaphase movement (39,40), and T-complex protein ɛ may be required for metaphase/anaphase transition (41). With this suppressor screening, we also identified a novel gene that is functionally unknown but shares weak similarity with CENP-A. Functional analyses of these suppressor genes is expected to contribute to our understanding of CENP-C function.
Acknowledgments
ACKNOWLEDGEMENTS
The authors are very grateful to Ms Y. Miyauchi, Ms K. Suzuki and Ms K. Kita for technical assistance and Dr K. Ikenaka and Dr T. Yoshimatsu for the gift of ϕMP34 cell line. We also thank Dr W. Brown and Dr P. Nurse for advice about the temperature-sensitive mutants in DT40. This work was supported by a Grant-in-Aid for Scientific Research on Priority Areas (A) ‘Cell Cycle’ and (C) ‘Genome Biology’ and ‘Cancer Biology’ from the Ministry of Education, Science, Sports and Culture of Japan.
References
- 1.Choo K.H.A. (1997) The Centromere. Oxford University Press, Oxford, New York, Tokyo.
- 2.Lengauer C., Kinzler,K.W. and Vogelstein,B. (1998) Genetic instabilities in human cancers. Nature, 396, 643–649. [DOI] [PubMed] [Google Scholar]
- 3.Doheny K.F., Soger,P.K., Hyman,A.A., Tugendreich,S., Spencer,F. and Hieter,P. (1993) Identification of essential components of the S. cerevisiae kinetochore. Cell, 73, 761–774. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Pidoux A.L. and Allshire,R.C. (2000) Centromeres: getting a grip of chromosomes. Curr. Opin. Cell Biol., 12, 308–319. [DOI] [PubMed] [Google Scholar]
- 5.Stoler S., Keith,K.C., Curnick,K.E. and Fitzgerald-Hayes,M. (1995) A mutation in CSE4, an essential gene encoding a novel chromatin associated protein in yeast, cause chromosomes nondisjunction and cell cycle arrest at mitosis. Genes Dev., 9, 573–586. [DOI] [PubMed] [Google Scholar]
- 6.Meluh P.B., Yang,P., Glowczewski,L., Koshland,D. and Smith,M. (1998) CSE4p is a component of the core centromere of Saccharomyces cerevisiae. Cell, 94, 607–613. [DOI] [PubMed] [Google Scholar]
- 7.Brown M.T., Goetsch,L., and Hartwell,L.H. (1993) MIF2 is required for mitotic integrity during anaphase spindle elongation in Sacchromyces cerevisiae. J. Cell Biol., 123, 387–403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Amon A. (1999) The spindle checkpoint. Curr. Opin. Genet. Dev., 9, 69–75. [DOI] [PubMed] [Google Scholar]
- 9.Chen R.-H., Water,J.W., Salmon,E.D. and Murray,A.W. (1996) Association of spindle assembly checkpoint component XMAD2 with unattached kinetochores. Science, 274, 242–246. [DOI] [PubMed] [Google Scholar]
- 10.Taylor S.S. and McKeon,F. (1997) Kinetochore localization of murine Bub1 is required for normal mitotic timing and checkpoint response to spindle damage. Cell, 89, 727–735. [DOI] [PubMed] [Google Scholar]
- 11.Taylor S.S., Ha,E. and McKeon,F. (1998) The human homologue of Bub3 is required for kinetochore localization of Bub1 and a Mad3/Bub1 related protein kinase. J. Cell Biol., 142, 1–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Moroi Y., Peebles,C., Fritzler,M.J., Steigerwald,J. and Tan,E.M. (1980) Autoantibody to centromere (kinetochore) in scleroderma sera. Proc. Natl Acad. Sci. USA, 77, 1627–1631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Compton D.A., Yen,T.J. and Cleveland,D.W. (1991) Identification of novel centromere/kinetochore-associated proteins using monoclonal antibodies generated against human mitotic chromosome scaffolds. J. Cell Biol., 112, 1083–1097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Pluta A.F., Mackay,A.M., Ainsztein,A.M., Goldberg,I.G. and Earnshaw,W.C. (1995) The centromere: Hub of chromosomal activities. Science, 270, 1591–1594. [DOI] [PubMed] [Google Scholar]
- 15.Craig J.M., Earnshaw,W.C. and Vagnarelli,P. (1999) Mammalian centromeres: DNA sequence, protein composition, and role in cell cycle progression. Exp. Cell Res., 246, 249–262. [DOI] [PubMed] [Google Scholar]
- 16.Dobie K.W., Hari,K.L., Maggert,K.A. and Karpen,G.H. (1999) Centromere proteins and chromosome in heritance: a complex affair. Curr. Opin. Genet. Dev., 9, 206–217. [DOI] [PubMed] [Google Scholar]
- 17.Choo K.H.A. (2000) Centromerization. Trends Cell Biol., 10, 182–188. [DOI] [PubMed] [Google Scholar]
- 18.Tyler-Smith C. and Floridia,G. (2000) Many paths to the top of the mountain: diverse evolutionary solutions to centromere structure. Cell, 102, 5–8. [DOI] [PubMed] [Google Scholar]
- 19.Saitoh H., Tomkiel,J., Cooke,C.A., Ratrie,H., Maurer,M., Rothfield,N.F. and Earnshaw,W.C. (1992) CENP-C, an autoantigen in scleroderma, is a component of the human inner kinetochore plate. Cell, 70, 115–125. [DOI] [PubMed] [Google Scholar]
- 20.Yang C.H., Tomkiel,J., Saitoh,H., Johnson,D.H. and Earnshaw,W.C. (1996) Identification of overlapping DNA-binding and centromere-targeting domains in the human kinetochore protein CENP-C. Mol. Cell. Biol., 16, 3576–3586. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Fukagawa T. and Brown,W.R.A. (1997) Efficient conditional mutation of the vertebrate CENP-C gene. Hum. Mol. Genet., 6, 2301–2308. [DOI] [PubMed] [Google Scholar]
- 22.Kalitsis P., Fowler,K.J., Earle,E., Hill,J., and Choo,K.H.A. (1998) Targeted disruption of mouse centromere protein C gene leads to mitotic disarray and early embryo death. Proc. Natl Acad. Sci. USA, 95, 576–582. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Fukagawa T., Pendon,C., Morris,J., and Brown,W. (1999) CENP-C is necessary but not sufficient to induce formation of functional centromere. EMBO J., 18, 4196–4209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Tomkiel J., Cooke,C.A., Saitoh,H., Bernat,R.L., and Earnshaw,W.C. (1994) CENP-C is required for maintaining proper kinetochore size and for a timely transition to anaphase. J. Cell Biol., 125, 531–545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Earnshaw W.C., Ratrie,H., and Stetten,A.C.G. (1989) Visualization of centromere proteins CENP-B and CENP-C on a stable dicentric chromosome in cytological spreads. Chromosoma, 98, 1–12. [DOI] [PubMed] [Google Scholar]
- 26.Sullivan B.A. and Schwartz,S. (1995) Identification of centromeric antigens in dicentric Robertsonian translocations: CENP-C and CENP-E are necessary components of functional centromeres. Hum. Mol. Genet., 4, 2189–2197. [DOI] [PubMed] [Google Scholar]
- 27.Buerstedde J.M. and Takeda,S. (1991) Increased ratio of targeted to random integration after transfection of chicken B cell lines. Cell, 67, 179–188. [DOI] [PubMed] [Google Scholar]
- 28.Yoshimatsu T., Tamura,M., Kuriyama,S. and Ikenaka,K. (1998) Improvement of retroviral packaging cell lines by introducing the polyomavirus early region. Hum. Gene Ther., 9, 161–172. [DOI] [PubMed] [Google Scholar]
- 29.Fukagawa T., Mikami,Y., Nishihashi,A., Regnier,V., Haraguchi,T., Hiraoka,Y., Sugata,N., Todokoro,K., Brown,W. and Ikemura,T. (2001) CENP-H, a constitutive centromere component, is required for centromere targeting of CENP-C in vertebrate cells. EMBO J., 20, 4603–4617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Fukagawa T., Hayward,N., Yang,J., Azzalin,C., Griffin,D., Stwart,A.F. and Brown,W. (1999) The chicken HPRT gene: a counter selectable marker for the DT40 cell line. Nucleic Acids Res., 27, 1966–1969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Brown M.T. (1995) Sequence similarities between the yeast chromosome segregation protein MIF2 and the mammalian centromere protein CENP-C. Gene, 160, 111–116. [DOI] [PubMed] [Google Scholar]
- 32.Sullivan K.F., Hechenberger,M. and Masri,K. (1994) Human CENP-A contains a histone H3 related histone fold domain that is required for targeting to the centromere. J. Cell Biol. 127, 581–592. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Ishov A.M., Sotnikov,A.G., Negorev,D., Vladimirova,O.V., Neff,N., Kamitani,T., Yeh,E.T., Strauss,J.F.,III and Maul,G.G. (1999) PML is critical for ND10 formation and recruits the PML-interating protein Daxx to this nuclear structure when modified by SUMO-1. J. Cell Biol., 147, 221–233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Adachi N., Ikeda,H. and Kikuchi,A. (1994) Mutant isolation of mouse DNA topoisomerase IIα in yeast. Nucleic Acids Res., 22, 4229–4233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Knehr M., Poppe,M., Schroeter,D., Eickelbaum,W., Finze,E.M., Kiesewetter,U.L., Enulescu,M., Arand,M. and Paweletz,N. (1996) Cellular expression of human centromere protein C demonstrates a cyclic behavior with highest abundance in the G1 phase. Proc. Natl Acad. Sci. USA, 93, 10234–10239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Meluh P.B. and Koshland,D. (1995) Evidence that the MIF2 gene of Saccharomyces cerevisiae encodes a centromere protein with homology to the mammalian centromere protein CENP-C. Mol. Biol. Cell, 6, 793–807. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Pluta A.F., Earnshaw,W.C. and Goldberg,I.G. (1998). Interphase-specific association of intrinsic centromere protein CENP-C with HDaxx, a death domain-binding protein implicated in Fas-mediated cell death. J. Cell Sci., 111, 2029–2041. [DOI] [PubMed] [Google Scholar]
- 38.Everett R.D., Earnshaw,W.C., Pluta,A.F., Sternsdorf,T., Ainsztein,A.M., Carmena,M., Ruchaud,S., Hsu,W.L. and Orr,A. (1999) A dynamic connection between centromeres and ND10 proteins. J. Cell Sci., 112, 3443–3454. [DOI] [PubMed] [Google Scholar]
- 39.Funabiki H. and Murray,A.W. (2000) The Xenopus chromokinesin Xkid is essential for metaphase chromosome alignment and must be degraded to allow anaphase chromosome movement. Cell, 102, 411–424. [DOI] [PubMed] [Google Scholar]
- 40.Antonio C., Ferby,I., Wilhelm,H., Jones,M., Karsenti,E., Nebreda,A.R. and Vernos,I. (2000) Xkid, a chromokinesin required for chromosome alignment on the metaphase plate. Cell, 102, 425–435. [DOI] [PubMed] [Google Scholar]
- 41.Tokumoto M., Horiguchi,R., Nagahama,Y., Ishikawa,K. and Tokumoto,T. (2000) Two protein, a goldfish 20S proteasome subunit and the protein interacting with 26S proteasome, change in the meiotic cell cycle. Eur. J. Biochem. 267, 97–103. [DOI] [PubMed] [Google Scholar]