

ABSTRACT
Activation of the egg by the sperm is the first, vital stage of embryogenesis. The sperm protein PLCζ has been proposed as the physiological agent that triggers the Ca2+ oscillations that normally initiate embryogenesis. Consistent with this, recombinant PLCζ induces Ca2+ oscillations in eggs and debilitating mutations in the PLCZ1 gene are associated with infertility in men. However, there has been no evidence that knockout of the gene encoding PLCζ abolishes the ability of sperm to induce Ca2+ oscillations in eggs. Here, we show that sperm derived from Plcz1–/– male mice fail to trigger Ca2+ oscillations in eggs, cause polyspermy and thus demonstrate that PLCζ is the physiological trigger of these Ca2+ oscillations. Remarkably, some eggs fertilized by PLCζ-null sperm can develop, albeit at greatly reduced efficiency, and after a significant time-delay. In addition, Plcz1–/– males are subfertile but not sterile, suggesting that in the absence of PLCζ, spontaneous egg activation can eventually occur via an alternative route. This is the first demonstration that in vivo fertilization without the normal physiological trigger of egg activation can result in offspring. PLCζ-null sperm now make it possible to resolve long-standing questions in fertilization biology, and to test the efficacy and safety of procedures used to treat human infertility.
KEY WORDS: Sperm, Egg, Embryogenesis, Calcium signalling, PLCζ, Mouse
Highlighted article: In mice, PLCζ-null sperm fail to trigger the Ca2+ oscillations in eggs that normally initiate embryogenesis in mammals, resulting in polyspermy and subfertility but not sterility.
INTRODUCTION
Infertility is a world health problem that affects an estimated 1 in 7 couples and can be caused by problems in the development of viable gametes, but also by defects in fertilization or embryo development. Activation of the egg by the sperm at fertilization is a fundamental event in embryogenesis, and has been of scientific interest since Jacques Loeb's pioneering studies in the 1890s (Parrington et al., 2007). Defects in this process underlie some cases of human male infertility (Escoffier et al., 2016; Heytens et al., 2009; Kashir et al., 2012), and may have relevance for more general problems of embryo development in humans. In mammals, egg activation is triggered by sperm-induced Ca2+ oscillations in the egg (Igusa and Miyazaki, 1986; Kline and Kline, 1992). A key role of these Ca2+ oscillations is to downregulate, via a signalling pathway involving the Ca2+-dependent protein kinase gamma (CamKIIγ) and early mitotic inhibitor (Emi2), the maturation promotion factor (MPF) responsible for the metaphase II arrest of the egg (Sanders and Swann, 2016; Sobinoff et al., 2013). A later decline in mitogen-activated protein kinase (MAPK) activity correlates with the formation of pronuclei and entry into interphase of the first cell cycle. Another important event triggered following sperm-egg fusion is the block of polyspermy, which involves both changes in the egg membrane linked to the shedding of the egg protein Juno (Bianchi et al., 2014), and the cortical reaction: the Ca2+-dependent exocytosis of cortical granules and hardening of the zona pellucida of the egg (Li et al., 2013).
Despite advances in our understanding of the signalling pathways that lie downstream of the sperm-induced Ca2+ oscillations, exactly how the sperm triggers such Ca2+ oscillations in the egg has remained controversial. Previous studies have provided evidence that the physiological trigger of the Ca2+ oscillations is a sperm protein called phospholipase C zeta (PLCζ) (Ito et al., 2011; Swann and Lai, 2016). Thus, recombinant PLCζ RNA or protein trigger Ca2+ oscillations when injected into eggs, and depletion of PLCζ from sperm extracts eliminates their ability to trigger such Ca2+ oscillations (Kashir et al., 2011; Sanusi et al., 2015; Saunders et al., 2002). In addition, partial knockdown of Plcz1 expression in mouse sperm using RNA interference leads to sperm-induced Ca2+ oscillations that terminate prematurely, to lower rates of egg activation and to no transgenic offspring after fertilization in vivo (Knott et al., 2005). Further evidence for an important physiological role for PLCζ has come from the discovery of debilitating mutations in the PLCZ1 gene that are associated with infertility in men (Escoffier et al., 2016; Heytens et al., 2009; Kashir et al., 2012). However, despite these pieces of evidence implicating PLCζ as the physiological agent of egg activation, irrefutable evidence using genetic disruption methods to show that PLCζ is the sole initiator of Ca2+ oscillations in mammalian eggs has been lacking, and recently doubts have been raised about a role for PLCζ in the process of egg activation (Aarabi et al., 2012; Ferrer-Vaquer et al., 2016).
Another controversial issue is the importance of the Ca2+ oscillations induced by the sperm during mammalian fertilization for optimum embryo development. Although such oscillations constitute the physiological stimulus, previous studies have shown that egg activation and development to blastocyst in vitro can be induced by pharmacological or genetic treatments that reduce MPF levels in ways that do not require a Ca2+ stimulus (Knott et al., 2006; Miao et al., 2012; Phillips et al., 2002; Suzuki et al., 2010). In addition, currently infertility in men whose sperm fail to induce egg activation can be successfully treated by mechanical, electrical or chemical stimuli that induce a Ca2+ signal that is far from physiological (Sfontouris et al., 2015; Vanden Meerschaut et al., 2014). Such approaches have led to the successful births of hundreds of human infants worldwide. However, other studies of mouse parthenogenetic embryos stimulated by different patterns of artificially induced Ca2+ transients (Ducibella et al., 2002), or fertilized mouse eggs whose endogenous Ca2+ oscillations have been inhibited or hyper-stimulated (Ozil et al., 2005), show that altering the number and frequency of Ca2+ transients can subtly affect embryonic gene expression and developmental potential. Therefore, the long-term safety implications of the use of artificial egg activation stimuli in clinical practice remain far from clear.
To address these issues further experimentally, what has been lacking is a mouse in which PLCζ expression has been eliminated through gene knockout. This would make it possible to assess the physiological significance of PLCζ. In addition, if sperm lacking expression of functional PLCζ were found to be incapable of triggering Ca2+ oscillations in the egg, this would provide a null background in which to test the effects of artificial egg activation stimuli as inducers of egg activation and embryo development. Here, we describe the generation of such a PLCζ-null mouse through CRISPR/Cas9 gene editing (Doudna and Charpentier, 2014), and its phenotypic analysis.
RESULTS
Generation of PLCζ knockout mice using CRISPR/Cas9 gene editing
To create a PLCζ-null mouse, we used CRISPR/Cas9 gene editing to target the Plcz1 gene in fertilized mouse eggs. Two strategies were employed that targeted different exons of Plcz1 with independent single guide RNAs (sgRNAs) to control for potential off-target mutagenesis: one used Cas9WT endonuclease with one sgRNA; the other used Cas9D10A nickase with two paired sgRNAs (Fig. 1A). This second strategy was employed because it reduces off-target mutations (Ran et al., 2013; Shen et al., 2014). Two mutant Plcz1 mouse lines were produced, originating from two F0 females harbouring frameshift-producing deletions: Plcz1em1Jparr, generated using Cas9D10A nickase, has a 22-nucleotide deletion in exon 3 (Fig. 1B,D); Plcz1em2Jparr, generated using Cas9WT endonuclease, has a 17-nucleotide deletion in exon 5 (Fig. 1C,D).
Fig. 1.

Genomic characteristics of mouse Plcz1em1Jparr and Plcz1em2Jparr mutant alleles. (A) Gene structure of the mouse Plcz1 gene (NM_054066.4) and target sequences (red lines) for CRISPR/Cas9. Exons are represented by vertical bars (unfilled, untranslated regions; filled, coding regions). (B,C) Comparison of genomic sequences from wild-type Plcz1 (Plcz1WT) allele and two mutant Plcz1 alleles harbouring nucleotide deletions (deleted sequence shaded grey in Plcz1WT sequence). Capital letters correspond to exonic sequences and lower case letters to intronic sequences. (D) Typical result from a genotyping PCR reaction for both Plcz1 mutant lines: HET, Plcz1+/em1Jparr or Plcz1+/em2Jparr; MUT, homozygote Plcz1em1Jparr or Plcz1em2Jparr; WT, homozygote Plcz1WT.
Sequence analysis of RT-PCR products from testis mRNA obtained from these two lines confirmed that these corresponded to the predicted mutant cDNA sequences with no observable residual wild-type Plcz1 (Plcz1WT) sequence contamination (Fig. 2A,C), suggesting complete lack of Plcz1WT expression in these mutant lines. The truncated proteins that could be produced from the two mutant Plcz1 alleles (Fig. 2B,D) correspond to small N-terminal sequences lacking regions necessary for enzymatic activity (Nomikos et al., 2013). The absence of full-length PLCζ was further confirmed by immunoblotting analysis of sperm derived from Plcz1em1Jparr mice (Fig. 2E).
Fig. 2.

Expression analysis of mouse Plcz1em1Jparr and Plcz1em2Jparr mutant alleles. (A,C) Comparison of cDNA sequences obtained from RT-PCR reactions using testis RNA from Plcz1WT and homozygote Plcz1em1Jparr (A) or Plcz1em2Jparr (C) animals. Codons are underlined, with amino acids indicated by their three-letter code. Numbers below amino acids correspond to the last amino acid position within the primary sequence not affected by the frameshift. The amino acid sequence after the frameshift is in grey. (B,D) Predicted protein-domain structures for wild-type mouse PLCζ protein (PLCζWT) and truncated proteins resulting from expression of mutant PLCζ em1Jparr (B) and PLCζ em2Jparr (D) alleles. (E) Immunoblotting analysis of PLCζ in sperm from Plcz1WT (two animals) and PLCζ em1Jparr (four animals) using a custom-made anti-PLCζ antibody (Kurokawa et al., 2005). The immunoreactive band corresponding to PLCζ is indicated by a red arrow. (F) Agarose gel image showing integrity of in vitro-produced cRNA. (G) Typical Ca2+ responses, monitored by the fluorescence ratio of Fura-2 in wild-type mouse eggs, elicited by injection of recombinant copy RNA (cRNA) in vitro transcribed from Plcz1WT, Plcz1em1Jparr or Plcz1em2Jparr cDNAs. Proportion of eggs showing Ca2+ oscillations: Plcz1WT ([cRNA] 1 ng/μl=10/10; [cRNA] 50 ng/μl=8/8), Plcz1em1Jparr ([cRNA] 50 ng/μl=5/5; [cRNA] 100 ng/μl=5/5) and Plcz1em1Jparr ([cRNA] 50 ng/μl=5/5; [cRNA] 100 ng/μl=5/5).
It has been previously suggested that the post-acrosomal WW domain-binding protein (Wbp2nl/PAWP), could function as the physiological agent of egg activation (Aarabi et al., 2014). We therefore investigated the status of Wbp2nl/PAWP expression in our Plcz1 mutant lines. RT-qPCR analysis for Wbp2nl mRNA revealed that the Plcz1 mutations present in our two Plcz1 mutant lines have no detrimental effects upon Wbp2nl mRNA expression in testis (Fig. S1A). Furthermore, immunoblotting analysis for PAWP revealed no apparent differences in the level of PAWP protein between Plcz1WT- and Plcz1em1Jparr-derived sperm (Fig. S1B).
We next carried out functional analysis of the mutant Plcz RNA predicted to be expressed in the two mutant mouse lines. Plcz1WT recombinant copy RNA (cRNA) (Fig. 2F) injected into mouse eggs triggered Ca2+ oscillations in a dose-dependent manner, consistent with production of functional PLCζ protein (Saunders et al., 2002) (Fig. 2G); however, injection of either mutant cRNA failed to induce Ca2+ oscillations (Fig. 2G). In combination, these results indicate that these mutant Plcz1 lines are null for functional PLCζ expression and will be referred to as Plcz1–/–.
PLCζ knockout does not affect spermatogenesis or sperm quality parameters
Histological analysis of Plcz1–/– male testes indicated no defects in spermatogenesis (Fig. 3A). Furthermore, epididymal sperm from Plcz1–/– males (Plcz1– sperm) had normal viability (Fig. 3B,C), motility (Fig. 3D) and hyperactivity, which reflect their ability to undergo capacitation (Fig. 3E-G). Additionally, the ability of Plcz1– sperm to undergo the acrosome reaction in response to either progesterone or ionomycin was undistinguishable from the sperm of wild-type animals (Plcz1+ sperm) (Fig. 3H). Collectively, these results indicate that loss of PLCζ has no apparent detrimental effects upon spermatogenesis or parameters associated with the ability of the sperm to bind and fuse with the egg.
Fig. 3.

Characterization of sperm from homozygote Plcz1em1Jparr and Plcz1em2Jparr mice. (A) Histological analysis of testes sections stained with Hematoxylin/Eosin. (B) Density plots of sperm stained with SYBR14 (live sperm) and propidium iodide (PI; dead sperm) and analysed by flow cytometry. A total of 10,000 events were collected for each group. (C) Quantification of data from B. Representative stained sperm were obtained using an Olympus BX40 microscope. (D-G) Analysis of sperm motility parameters using computer-assisted sperm analysis (CASA). (H) An acrosome reaction was induced either by treatment with progesterone or ionomycin, and reacted sperm were assessed by lack of reactivity towards fluorescently labelled lectin PNA. Representative images of reacted and unreacted sperm are shown. All data are presented as average±s.e.m. of four different animals. There are no statistically significant differences.
Loss of PLCζ abolishes the ability of the sperm to trigger Ca2+ oscillations in the egg
Using intracytoplasmic sperm injection (ICSI), we next investigated whether Plcz1– sperm could trigger Ca2+ oscillations in wild-type eggs. Injection of Plcz1+ sperm induced Ca2+ oscillations symptomatic of those seen at fertilization in nearly all eggs (Fig. 4A,B) (Wakai et al., 2011). In contrast, injected sperm from either Plcz1–/– line failed to induce Ca2+ oscillations (Fig. 4A,B), which were nevertheless triggered by subsequent injection of Plcz1WT cRNA (Fig. 4C). The inability of Plcz1– sperm to trigger Ca2+ oscillations was further confirmed following in vitro fertilization (IVF) using zona pellucida-free eggs (Fig. 4D,E); while 16/34 eggs showed Ca2+ oscillations following IVF with Plcz1+ sperm, none of the 40 eggs subjected to IVF with Plcz1em1Jparr-derived sperm showed such oscillations, with one egg showing a single Ca2+ transient (Fig. 4D,E). This is therefore the first direct evidence that PLCζ is the sole physiological trigger of the Ca2+ oscillations responsible for egg activation in mammals (Wakai et al., 2011).
Fig. 4.
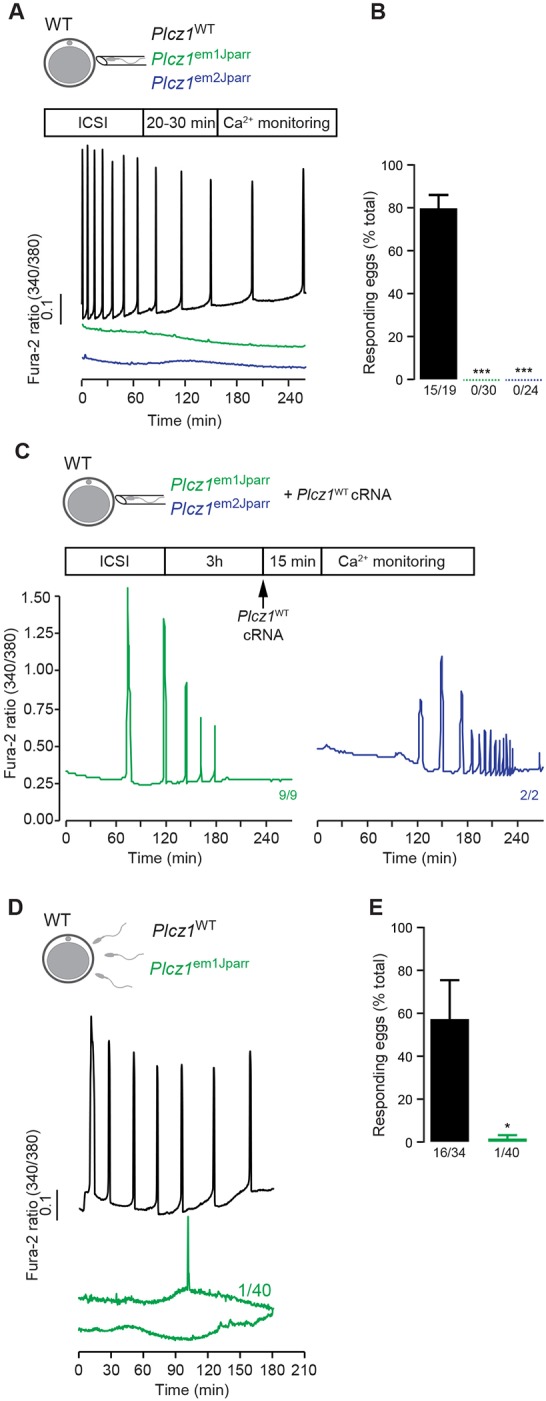
Ca2+ responses elicited during fertilization of mouse eggs. (A) Representative traces of Ca2+ responses, monitored by the fluorescence ratio (excitation 340/380 nm) of Fura-2, in wild-type mouse eggs, elicited by ICSI with sperm from homozygote Plcz1WT, Plcz1em1Jparr or Plcz1em2Jparr males. (B) Compilation of results from all eggs subjected to ICSI. Data are presented as average±s.e.m. followed by a one-way ANOVA analysis; numbers shown underneath each bar indicate proportion of responding eggs; the following number of animals were used for each group in a total of four or five experiments: Plcz1WT (3), Plcz1em1Jparr (2) and Plcz1em2Jparr (4). (C) Representative traces of Ca2+ responses, monitored by the fluorescence ratio (excitation 340/380 nm) of Fura-2, in wild-type mouse eggs, elicited by injection of Plcz1WT cRNA after ICSI with sperm from homozygote Plcz1em1Jparr and Plcz1em2Jparr males. Numbers shown next to traces indicate proportion of responding eggs subjected to ICSI and cRNA injection. (D) Representative traces of Ca2+ responses, in wild-type mouse eggs, elicited by IVF with sperm from Plcz1WT and Plcz1em1Jparr males; the Plcz1WT control sperm were from a transgenic line of the same strain background as the Plcz1em1Jparr sperm, in which the catalytic subunit of PKA was exclusively modified in the sperm (Morgan et al., 2008). (E) Compilation of results from all eggs subjected to IVF. Data are presented as average±s.e.m. followed by Student's t-test; numbers shown underneath each bar indicate proportion of responding eggs; the following number of animals were used for each group in a total of four or five experiments: Plcz1WT (5) and Plcz1em1Jparr (1).
Increased polyspermy in eggs fertilized by PLCζ-null sperm after IVF or in vivo fertilization
The inability of Plcz1– sperm to trigger Ca2+ oscillations following IVF could be due to failure of Plcz1– sperm to bind and fuse with the egg. However, no significant differences were observed in the percentage of eggs containing sperm heads after 6 h post-IVF with Plcz1WT- or Plcz1em1Jparr-derived sperm (Plcz1WT, 143/193; Plcz1em1Jparr, 103/132) assessed by Hoechst DNA staining (Fig. 5A). Remarkably, we noticed a dramatic increase in the rate of polyspermy among eggs fertilized by IVF with Plcz1– sperm when compared with Plcz1+ sperm (Plcz1WT, 24/143; Plcz1em1Jparr, 74/103) (Fig. 5B).
Fig. 5.
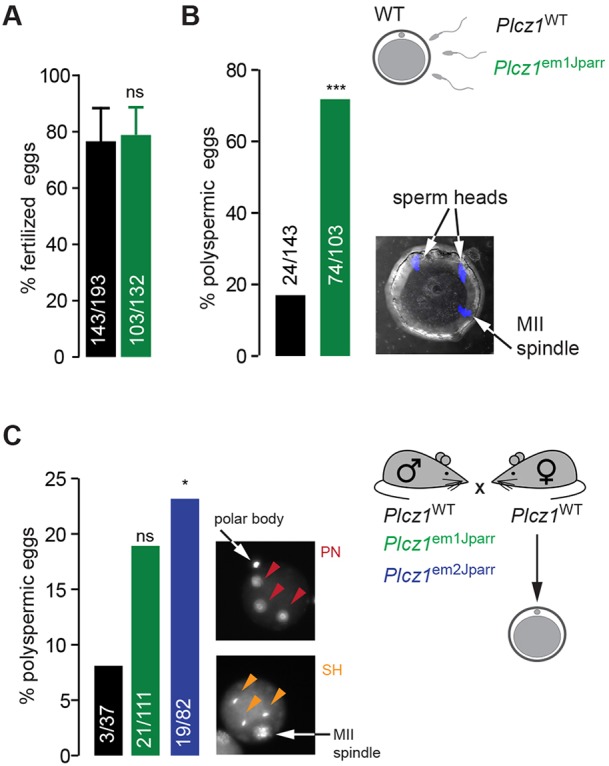
Increased polyspermy in eggs fertilized by Plcz1em1Jparr and Plcz1em2Jparr mutant sperm. (A,B) Wild-type eggs were stained with Hoechst 6 h post-IVF with Plcz1WT- and Plcz1em1Jparr-derived sperm (Plcz1WT: 193 eggs, 1 male; Plcz1em1Jparr: 132 eggs, 1 male). (A) Percentage of eggs showing sperm penetration as indicative of fertilization. (B) Percentage of eggs showing abnormal number (>1) of sperm heads. Data are presented as percentage of total eggs examined from three combined experiments. An example of a fertilized egg showing polyspermy is shown. (C) Number of polyspermic eggs derived from super-ovulated wild-type females fertilized in vivo by homozygote Plcz1WT, Plcz1em1Jparr or Plcz1em2Jparr males. Eggs were stained with Hoechst 18 h post-hCG treatment of females, for visualization of abnormal number of pronuclei (>2 PN) or sperm heads (>1 SH). Data are presented as percentage of total eggs examined from combined experiments (Plcz1WT, 37 eggs, 2 male; Plcz1em1Jparr, 111 eggs, 3 males; Plcz1em2Jparr, 82 eggs, 2 males) and statistical analysis performed using a 2×2 contingency table followed by a Fisher's exact test. Examples of fertilized eggs showing polyspermy are shown.
To discount possible artefacts associated with IVF, we investigated the levels of polyspermy in wild-type eggs from females fertilized in vivo by Plcz1+/+ and Plcz1–/– males. Hoechst DNA staining at 18 h post-hCG showed a clear increase in polyspermy among eggs fertilized by Plcz1–/– males (Plcz1em1Jparr, 21/111; Plcz1em2Jparr, 19/82) versus eggs fertilized by Plcz1+/+ males (3/37) (Fig. 5C), which confirms the findings observed following IVF.
These results indicate therefore that Plcz1– sperm are normal, regarding their ability to penetrate the egg, but suggest that, in the absence of Ca2+ oscillations, the mechanisms that block polyspermy (Fig. S2A,B) are not engaged in a timely fashion. Whether or not this failure is limited to cortical granule release is unknown; additional studies using Plcz1– sperm will be required to elucidate the status of other mechanisms that control the block to polyspermy and the role of Juno (Bianchi et al., 2014).
In vitro development of eggs fertilized by PLCζ-null sperm
As Ca2+ oscillations are considered to be the physiological trigger of mammalian egg activation (Fig. S2A,C) (Wakai et al., 2011), we assessed the ability of Plcz1– sperm to trigger this process. Following ICSI using Plcz1+ sperm, 35/40 of zygotes reached two-cell stage and 27/40 developed to the blastocyst stage (Fig. 6A). In contrast, Plcz1– sperm largely failed to trigger activation, with only a few eggs reaching the two-cell stage (Plcz1em1Jparr, 2/36; Plcz1em2Jparr, 1/47) and only one reaching the blastocyst stage (Plcz1em1Jparr, 1/36; Plcz1em2Jparr, 0/47) (Fig. 6A). This inability of Plcz1– sperm to trigger consistent egg activation was rescued by injection of Plcz1WT cRNA 3 h after ICSI with a significant proportion of eggs now developing to the two-cell stage (Plcz1em1Jparr, 16/20; Plcz1em2Jparr, 6/9) (Fig. 6A).
Fig. 6.
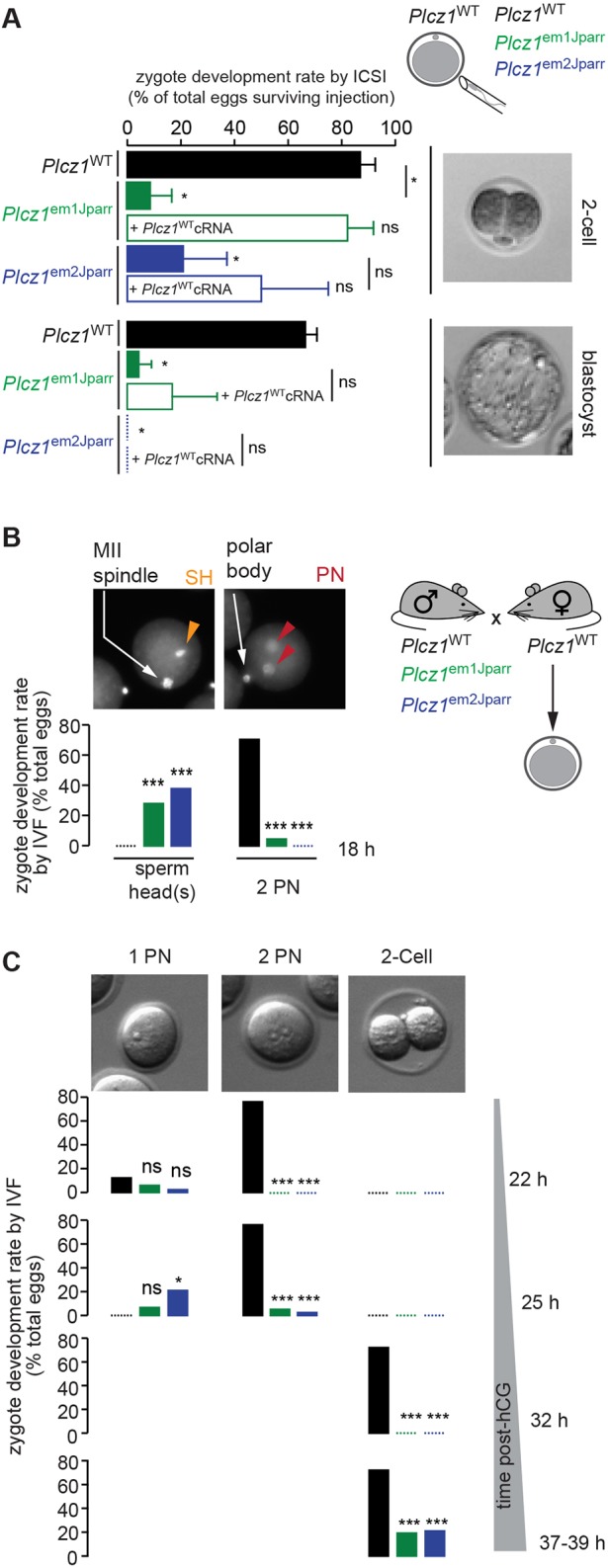
Embryo development in the absence of Ca2+ oscillations. (A) Proportion of two-cell and blastocyst embryos 2 days or 5 days post-ICSI, respectively (Plcz1WT, 40 eggs, 3 males; Plcz1em1Jparr, 36 eggs, 1 male; Plcz1em2Jparr, 47 eggs, 3 males). Plcz1WT cRNA was injected after ICSI, where indicated (Plcz1em1Jparr, 20 eggs, 1 male; Plcz1em2Jparr, 9 eggs, 2 males). Data are presented as average±s.e.m. of three to six experiments and example embryos are shown. (B,C) Eggs from super-ovulated females treated with hCG were fertilized in vivo, collected between 18 and 22 h post-hCG, and allowed to develop in vitro. Proportion of eggs showing sperm head(s) (SH), 1 pronucleus (1PN), 2 pronuclei (2PN) and 2-cell embryos at the indicated times. Examples of embryonic development stages are shown. (B) Data for 18 h were obtained from Hoechst-stained eggs (Plcz1WT, 37 eggs, 2 male; Plcz1em1Jparr, 111 eggs, 3 males; Plcz1em2Jparr, 82 eggs, 2 males). (C) Data from 22 h onwards were obtained from: Plcz1WT, 25 eggs, 2 males; Plcz1em1Jparr, 120 eggs, 3 males; Plcz1em2Jparr, 43 eggs, 1 male. For all panels, statistical significance was analysed using one-way ANOVA.
The unexpected, albeit greatly reduced, ability of some eggs injected with sperm from Plcz1–/– males to undergo egg activation and development after ICSI could be caused by the injection protocol. To bypass this possible artefact, and others associated with in vitro fertilization procedures, we directly examined fertilization and embryo development of wild-type eggs from females fertilized in vivo by Plcz1+/+ and Plcz1–/– males. Eggs/zygotes were collected at 18 h post-hCG and allowed to develop in vitro during which time they were routinely examined. At the early time point of 18 h, differences in activation events were already obvious, as Plcz1+ fertilized eggs displayed pronuclei formation, whereas Plcz1–-fertilized eggs showed mostly condensed sperm head(s) (Fig. 6B). Thus, Plcz1– sperm induced egg activation much less efficiently and protractedly, as further observed at subsequent time points (Fig. 6C): eggs fertilized by Plcz1+ sperm showed a high rate of two pronuclei formation (19/25) by 22 h and cleavage to two cells (18/25) by 32 h post-hCG treatment, whereas eggs fertilized by Plcz1– sperm displayed only two pronuclei formation (Plcz1em1Jparr, 6/120; Plcz1em2Jparr, 1/43) by 25 h and cleavage to the two-cell stage (Plcz1em1Jparr, 23/120; Plcz1em2Jparr, 9/43) after 38 h post-hCG treatment (Fig. 6C).
PLCζ knockout male mice are subfertile
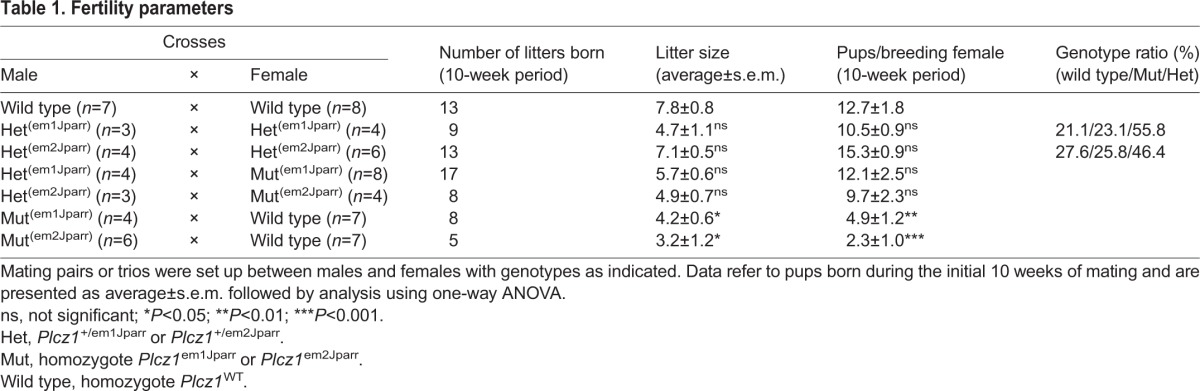
We next examined how the inability of Plcz1– sperm to trigger Ca2+ oscillations could affect the capacity of Plcz1–/– males to produce live offspring. Table 1 shows the compilation of results from various crosses. We found that, unlike crosses between Plcz1+/– males and females or between Plcz1–/– females and Plcz1+/– males, which resulted in offspring similar in number to Plcz1+/+ crosses and with genotypes close to Mendelian proportions, Plcz1–/– males could produce offspring, but at a much reduced efficiency compared with Plcz1+/+ males. Thus, although wild-type females crossed with Plcz1+/+ males produced average litter sizes of 7.8±0.8 and an average of 12.7±1.8 pups per female over the initial mating period of 10 weeks, wild-type females crossed with homozygote Plcz1em1Jparr or Plcz1em2Jparr males produced smaller litter sizes (4.2±0.6 or 3.2±1.2, respectively) with an average of 4.9±1.2 and 2.3±1.0 pups per female, respectively, over the same time period. This is the first demonstration in mammals that natural mating resulting in vivo fertilization without the normal physiological trigger of egg activation can result in offspring.
Table 1.
Fertility parameters
DISCUSSION
In summary, our findings demonstrate that absence of functional PLCζ does not compromise spermatogenesis and sperm quality parameters such as viability, motility, hyperactivity and ability to undergo the acrosome reaction. This is in contrast to a previous preliminary report that claimed that Plcz1–/– male mice were defective in the spermatogenic process (Ito et al., 2011). However, we show that Plcz1– sperm lack the ability to induce Ca2+ oscillations in the egg. Thus, we provide the first direct evidence that PLCζ is the physiological trigger of Ca2+ oscillations in mouse eggs, and probably in eggs of all mammals. Remarkably, a few eggs fertilized by Plcz1– sperm can develop to the blastocyst stage in vitro, and natural mating of Plcz1–/– males with wild-type females also results in low rates of egg activation that nevertheless can lead to development to term. Importantly, this is the first demonstration in mammals that natural mating and fertilization without the normal physiological trigger of egg activation can result in offspring. These findings are significant because PLCζ-null sperm and embryos offer a way to test the efficacy and safety of activation stimuli used in clinical IVF and will make it possible for the first time to ascertain the impact of Ca2+ oscillations on gene expression in the early embryo and on the health of offspring.
A key role of the sperm-induced Ca2+ oscillations is to downregulate, via a signalling pathway involving CamKIIγ and Emi2, MPF, which is responsible for the metaphase II arrest of the egg (Fig. S2A) (Sanders and Swann, 2016; Sobinoff et al., 2013). A later decline in MAPK activity correlates with the formation of pronuclei and entry into interphase of the first cell cycle. Importantly, MPF levels can be reduced artificially in various ways, even without a Ca2+ stimulus, triggering egg activation and development to blastocyst in vitro (Knott et al., 2006; Miao et al., 2012; Phillips et al., 2002; Suzuki et al., 2010).
This mode of egg activation, without Ca2+ release, may explain the spontaneous activation observed in vivo in ovulated hamster and mouse eggs left to reside in the oviduct for extended periods of time in the absence of fertilization (Whittingham and Siracusa, 1978; Xu et al., 1997; Yanagimachi and Chang, 1961). Moreover, eggs from the C57BL/6 strain, the strain used in our studies, have been shown to have a particularly high susceptibility to spontaneous activation during in vitro maturation (Cheng et al., 2012). Thus, it is possible that a related mechanism might be responsible for the activation we observe. In the situation described here, the self-activation that aging unfertilized eggs experience, which normally results in fragmentation or embryonic arrest, is rescued by the Plcz1– fertilizing sperm. It remains to be seen if these findings have relevance for other mammals, including humans, besides mice.
One other possibility is that Plcz1– sperm are inducing an atypical Ca2+ signal in the egg, which was not detected during the 4 h window during which Ca2+ monitoring was performed in our IVF studies. However, we failed to detect any form of Ca2+ response following ICSI or IVF with Plcz1– sperm. Furthermore, following IVF or in vivo fertilization, there was a marked increase in the rates of polyspermy, which suggest that the fertilizing sperm cannot induce Ca2+ release to engage the mechanisms that block polyspermy. Last, a final formal possibility is that sperm factors other than PLCζ might be involved in egg activation in this instance. The post-acrosomal sheath WW domain-binding protein (PAWP) has been suggested to play a role in egg activation (Aarabi et al., 2014). However, sperm from PAWP-null (Wbp2nl–/–) mice have no defect in egg activation capacity (Satouh et al., 2015), arguing against a role in this process. Future studies will be required to uncover which of these potential alternative mechanisms is allowing egg activation to occur in the absence of the normal physiological trigger.
There has been considerable debate about the importance of Ca2+ oscillations for optimal egg activation and subsequent embryo development. Although it is possible to bypass the Ca2+-dependent route in vitro in various ways (Knott et al., 2006; Miao et al., 2012; Phillips et al., 2002; Suzuki et al., 2010), altering the number and frequency of Ca2+ transients can subtly affect embryonic gene expression and developmental potential (Ducibella et al., 2002; Ozil et al., 2005). This is of important clinical significance given that currently infertility in men whose sperm fail to induce egg activation can be treated by inducing egg activation artificially with mechanical, electrical or chemical stimuli (Sfontouris et al., 2015; Vanden Meerschaut et al., 2014). However, the long-term effects of such treatments on human development remain far from clear, an issue of some concern given that the Ca2+ signals induced by these treatments are highly non-physiological (Sfontouris et al., 2015; Vanden Meerschaut et al., 2014).
Our findings are important in this respect in two main ways. First, our demonstration that Plcz1–/– mouse males can still conceive offspring in vivo, albeit at a reduced efficiency, seemingly in the absence of the normal sperm-induced Ca2+ oscillations, will allow us to study whether these offspring differ from those resulting from normal egg activation that is dependent on Ca2+ signals. Second, the availability of PLCζ-null sperm now makes it possible to assess, in a mouse model, how artificial egg activation stimuli, used in the clinic, might affect embryonic gene expression and offspring growth, metabolism and behaviour. Importantly, it will provide a way to test the efficacy and safety of recombinant PLCζ protein as an alternative therapeutic agent to treat infertility caused by egg activation deficiency.
MATERIALS AND METHODS
Mice
The nomenclature of the Plcz1em1Jparr and Plcz1em2Jparr mice is in accordance with the MGI guidelines for nomenclature of mutant alleles. In Oxford, mice of the C57BL/6 strain were housed in individual ventilated cages at constant optimal temperature and humidity with 12 h of light per day in the Biomedical Science Building, or the Dunn School of Pathology, both part of the University of Oxford, and fed with standard dry pellets and water ad libitum. Animal use was approved by the University of Oxford's Local Ethical Review Committee and permitted by a licence from the UK Home Office in accordance with UK law (Animals [Scientific Procedures] Act 1986).
At the University of Massachusetts and the University of Ghent Hospital Medical School, mice were also used as a source of eggs for ICSI, IVF and microinjection experiments. Sperm used in these experiments were cryopreserved using straws at the University of Oxford and sent to the Massachusetts and Ghent labs where they were thawed before use in the experiments.
Mice used for egg collection in Massachusetts were 8- to 10-week-old CD1 females, which were handled following the US National Research Council's Animal Care and Welfare Guidelines. These procedures were approved by the Institutional Animal Care and Use Committee at the University of Massachusetts.
In Ghent, mice used for egg collection were 8- to 10-week-old B6D2F1 females (a cross between female C57BL/6 and male DBA/2 mice). All procedures involving live-animal handling and euthanasia were approved by the Ghent University Hospital Ethical Committee for Laboratory Animals (EC number 09/15).
sgRNA design and production
Single guide RNA (sgRNA) target sites were selected within the mouse Plcz1 sequence (NM_054066.4) using the CRISPR design tool (crispr.mit.edu). sgRNA sequences were cloned into either pX459 (Cas9WT) or pX461 (Cas9D10A) vectors (gifts from Prof. Feng Zhang, Massachusetts Institute of Technology, USA) and tested for efficacy by transfection into N2A mouse neuroblastoma cells using Fugene 6 followed by High Resolution Melting (HRM) analysis (Bassett and Liu, 2014). N2A mouse neuroblastoma cells were tested for mycoplasma, and were originally obtained from ATCC (https://www.lgcstandards-atcc.org/products/all/CCL-131.aspx?geo_country=gb). Based on HRM results, a pair of targeting sequences in exon 3 (GTATGAAGCACGaGAAGTTAAGG, AAGGTTCAGGATGATTTTAGAGG) and a single targeting sequencing in exon 5 (ACTCGGTCCCAATATTTGATCGG) were chosen for production of sgRNAs by in vitro transcription using a PCR template generated by Pfusion polymerase extension of a pair of overlapping primers, which add a T7 polymerase promoter (underlined) to the 5′ end and the common sgRNA sequence (bold) to the 3′ end: PLCZ1_exon3A(F), gaaattaatacgactcactataggAGGTTCAGGATGATTTTAGgttttagagctagaaatagc; PLCZ1_exon3B(F), gaaattaatacgactcactataggTATGAAGCACGaGAAGTTAgttttagagctagaaatagc; PLCZ1_exon5(F), gaaattaatacgactcactataggCTCGGTCCCAATATTTGATgttttagagctagaaatagc; sgRNA (R), aaaagcaccgactcggtgccactttttcaagttgataacggactagccttattttaacttgctatttctagctctaaaac.
Products were purified using a PCR purification kit (Qiagen), and in vitro transcribed using the T7 MEGAscript kit (Ambion) at 37°C for 4 h. sgRNAs were purified by phenol:chloroform extraction and precipitation using isopropanol followed by quantification by measuring absorbance at 260 nm. Total sgRNA (5 µg) was mixed with 10 µg capped and polyadenylated Cas9WT or Cas9D10A mRNA (Trilink) and 0.1 volumes 3 M sodium acetate (pH 5.2), and co-precipitated using three volumes ethanol. RNA was washed twice in 70% ethanol and resuspended in 15 µl water to a concentration of 1 mg/ml. Integrity of mRNA and sgRNA was confirmed by agarose gel electrophoresis.
Generation of Plcz1–/– mice
Three- to 4-week-old C57Bl/6J female mice (Charles River) were injected intraperitoneally with 5 IU of PMSG (Intervet) and 48 h later with 5 IU of hCG (Intervet), and were paired with C57BL/6J (Charles River) male mice. Zygotes were retrieved from oviductal ampullae at 20 h post-hCG. Cumulus-enclosed zygotes were denuded by exposure to 1 mg/ml hyaluronidase (Sigma) in modified human tubule fluid (HTF) (Zenith Biotech) for 3-6 min, washed through fresh modified HTF and cultured in HTF in the presence of 5% CO2 at 37°C. Two hours after release from the cumulus mass, zygotes with a normal morphology were microinjected into the cytoplasm in 30 µl drops of modified HTF media using a PMM-150FU Piezo impact drive (Primetech) with homemade glass capillaries with ∼5-10 pl RNA at a final concentration of 150 ng/µl (100 ng/µl Cas9 cRNA and 50 ng/µl sgRNA). Injected zygotes and non-injected controls were cultured overnight in KSOMaa (Zenith Biotech) in the presence of 5% CO2 at 37°C and checked for two-cell division the next day. B6CBF1 females (Charles River) were mated to a vasectomized male, and females showing the presence of a copulatory plug were surgically prepared to receive all zygotes with a normal two-cell morphology using a bilateral implantation. The resulting F0 offspring were biopsied, their genomic DNA was analysed by HRM, PCR products were cloned into PGEM-T vector and individual clones were sequenced. Founder F0 mice with mutant alleles were bred to C57BL/6J mice and offspring were tested for inheritance of mutant alleles by DNA sequencing. Heterozygote progeny (Plcz1+/–) were mated for production of homozygote mice (Plcz1–/–) and a colony for each Plcz1 mutant line was maintained by mating pairs using Plcz1–/– females and Plcz1+/– males.
Genotyping of Plcz1 mutant lines by PCR
Genomic DNA was extracted from tail or ear biopsies by digestion with proteinase K overnight at 65°C with agitation in 0.5 ml lysis buffer [100 mM Tris-HCl (pH 8), 5 mM EDTA, 200 mM NaCl, 0.1% SDS, 0.5 mg/ml proteinase K (Roche)]. Extracts were centrifuged at 17,000 g for 4 min, and supernatant was diluted 6.5× into water prior to inactivation of proteinase K at 95°C for 10 min. A 1 µl sample was used as a template for 10 µl PCR reactions using the following primers: Plcz1em1Jparr (F, ACAGGCAGGGAAAGGGTTT; R, TGTCTGAGCACAGGATAGATGA; wild-type product, 266 bp; mutant product, 244 bp); Plcz1em2Jparr (F, AGAAAGAAGGGAGGGGAATCTTC; R, TGATGGTATATCTGCATCCTTCTT; wild-type product, 342 bp; mutant product, 325 bp). Products were analysed by gel electrophoresis and by DNA sequencing.
Analysis of Plcz1 and Wbp2nl mRNA expression in testis
Total RNA was extracted from testes using single-step RNAzol RT (Sigma). For Plcz1 mRNA expression analysis, conventional RT-PCR was performed on the total RNA using OneTaq One-Step RT-PCR (New England Biolabs). The primers used to analyse Plcz1 mRNA expression, which are designed to anneal in different exons, were: Plcz1em1Jparr (F, AAGCCAACTTCATGAGCTCG; R, TCAATGGTGATTCTTCCTTGGT; wild-type product, 192 bp; mutant product, 170 bp); Plcz1em2Jparr (F, GAGAACTGTAAAACCGTGTACCA; R, CCCAGCAGTCAATTTCCAGA; wild-type product, 172 bp; mutant product, 155 bp).
Products were analysed by gel electrophoresis and by DNA sequencing. Wbp2nl mRNA expression levels were quantified in testes RNA by using OnestepPLUS qRT-PCR MasterMix SYBR Green (Primerdesign).
The primers used for Wbp2nl mRNA detection were: F, GGCAGCTCCAGACGGTGGCT; R, GCCCATCCAGAAGCTTGCAACTCG.
Wbp2nl mRNA expression levels were normalized to β-actin mRNA expression levels, and calculated using the relative quantification approach. Real-time qPCR was performed on a Roche LightCycler 480.
Immunoblot analysis of PLCζ and PAWP in sperm
Immunoblotting was performed as previously published (Kurokawa et al., 2005). Mouse sperm were diluted to appropriate concentrations, 2× sample buffer was added and samples were kept at −80°C until use. Thawed samples were boiled for 3 min, mixed well and loaded onto 7.5% SDS-PAGE gels; resolved polypeptides were transferred onto PVDF membranes (Millipore) using a Mini Trans-Blot Cell (Bio-Rad). The membranes were blocked in 6% nonfat dry milk in PBS-0.1% Tween and incubated overnight at 4°C with 1/1000 dilution of a custom-made anti-PLCζ rabbit sera [raised against a 19-mer sequence (GYRRVPLFSKSGANLEPSS) at the C-terminus of mouse PLCζ (mPLCζ)]; this was followed by a 1 h incubation with 1/2000 of a goat anti-rabbit horseradish peroxidase-labelled secondary antibody (Bio-Rad, 1662408EDU). Immunoreactivity was detected using chemiluminescence according to the manufacturer's instructions (PerkinElmer) using a Kodak Image Station 440CF. Immunoblot analysis of WPB2NL/PAWP was performed in a similar way, using overnight incubation at 4°C with 1/1000 dilution of an anti-WPB2NL/PAWP rabbit polyclonal antibody (Proteintech-22587-1-AP) (Kashir et al., 2017); this was followed by 1 h of incubation with 1/15,000 dilution of a goat anti-rabbit IRDye-800CW secondary antibody (Li-Cor, 926-32211) and fluorescence imaged using a Li-Cor Odyssey imaging system (Li-Cor). Immunoblotting procedures were repeated at least three times per sample.
Generation of recombinant PLCz1 RNA
A synthetic mouse Plcz1 cDNA (GeneArt Gene Synthesis, ThermoFisher Scientific) was subcloned into a pRNA vector for production of polyA+/capped recombinant mRNA using T3 RNA polymerase in an in vitro transcription reaction; capped transcripts were produced with the cap analogue m7G(5′)ppp(5′)G, following the manufacturer's instructions (mMessage mMachine, Ambion). Mutant Plcz1 cDNA versions were obtained by swapping internal fragments within wild-type Plcz1 cDNA, with corresponding synthetic sequences harbouring the deletions present in Plcz1em1Jparr or Plcz1em2Jparr. The quality of the recombinant mRNA was assessed by agarose gel electrophoresis.
Histology
Testes were fixed in 4% paraformaldehyde (PFA), dehydrated in increasing concentrations of ethanol, kept for 24 h in Histo-Clear II (National Diagnostics), then embedded in paraffin wax. Sections (5 μm) were cut and mounted on Superfrost Plus slides (Fisher Scientific). The sections were deparaffinized in Histo-Clear II, rehydrated in decreasing concentrations of ethanol, then stained with Harris Hematoxylin (Sigma-Aldrich) for 5 min. The sections were counterstained with eosin B solution (Sigma-Aldrich) and visualized under a Leica DM5000B microscope.
Sperm viability
Spermatozoa were collected from the cauda epididymis and vas deferens of 8-week-old mice and incubated in HEPES-buffered saline solution containing 4 mg/ml bovine serum albumin at 37°C for 30 min, and assayed using the LIVE/DEAD Sperm Viability Kit (Molecular Probes) following the manufacturer's instructions. In brief, the samples were incubated for 5 min with SYBR14 (Em 516 nm; labels live cells) at a final concentration of 100 nM, and then with PI (Em 617 nm; labels dead cells) at a final concentration of 12 µM for another 5 min. Stained samples were analysed using a BD FACS Canto II flow cytometer equipped with BD FACSDiva software version 6.1 collecting a minimum of 10,000 gated sperm cells for the log of the fluorescence. Samples within the SYBR14+/PI– quadrant were considered viable, and within the SYBR14–/PI+ quadrant were non-viable. Events within the SYBR14–/PI– quadrant were considered as debris. Data were analysed for the relative fluorescence of SYBR14/PI using FlowJo software v10.6. Sperm viability was calculated by the percentage of live cells relative to the total number. Images of fluorescently labelled sperm were obtained from samples loaded onto a microscope slide coated with 0.1% poly-D-lysine and imaged using an Olympus BX40 microscope.
Sperm motility and hyperactivation
Spermatozoa were collected as before and washed in a complete pre-warmed HTF medium at 37°C under 5% CO2. A 30 µl sample of the recovered sperm suspension (2×105 sperm/ml) after 1 h incubation with HTF was loaded into a pre-warmed Leja slide chamber by capillary action on an Olympus CX41 microscope. Sperm motility and hyperactivation parameters were analysed using computer-assisted sperm analysis (CASA) with software HTM-CEROS (version 12.3, Hamilton Thorne), recording at least 800 mouse sperm/slide for each group. The image-acquiring settings were: 60 frames per second, minimum contrast (50), minimum cell size (10 pixels), VAP cutoff (7.5 µm/s), VSL cutoff (6 µm/s), cell intensity (80). Different progressive sperm motility grading scores (rapid, medium, slow, static) were recorded according to the Average Path Velocity (VAP) and Straight Line Velocity (VSL) parameters, while sperm hyperactivation (vigour) values were evaluated by Curvilinear Velocity (VCL), Amplitude of Lateral Head displacement (ALH) and Beat Cross Frequency (BCF).
Acrosome reaction
Spermatozoa were collected as before, washed, then capacitated in complete HTF medium for 30 min at 37°C and 5% CO2. The samples were divided into three groups at a sperm density of 3×106 sperm/ml: the first group was treated with ionomycin at a final concentration of 10 µM in DMSO, the second group was treated with progesterone at a final concentration of 20 µM in DMSO and the third group was only treated with DMSO as a control. At different time points, spermatozoa were collected and pipetted onto a microscope slide coated with 0.1% poly-d-lysine, then fixed with acetone/methanol (1:1) on ice for 10 min. The slides were washed twice with phosphate-buffered saline before incubating with lectin PNA-Alexa Fluor 594 conjugate (Life Technologies) for 30 min in the dark. Slides were washed twice with H2O before mounting with ProLong Gold antifade reagent with DAPI for sperm DNA staining. The slides were assessed for acrosomal status (lectin PNA+, unreacted; lectin PNA–, reacted) using an Olympus BX40 microscope. The percentage of acrosome-reacted spermatozoa was calculated as percentage of total numbers.
Intracytoplasmic sperm injection (ICSI)
ICSI was carried out both at the University of Ghent Hospital Medical School and at the University of Massachusetts. Briefly, hybrid female mice were stimulated using pregnant mare serum gonadotrophin (PMSG). MII eggs were obtained from the oviducts 12 to 14 h after injection of hCG. After treatment with hyaluronidase to remove cumulus cells, MII eggs were washed and transferred into KSOM (potassium simplex optimized medium) under culture conditions (37°C, 6% CO2, 5% O2). Sperm microinjection was performed by piezo-drilling following protocols previously described (Yoshida and Perry, 2007; Yoon and Fissore, 2007). All manipulations were carried out using HEPES-buffered medium at 37°C, under light mineral oil at room temperature. Sperm heads were separated from the tail and microinjected by applying Piezo pulses of different intensity to penetrate the zona pellucida and plasma membrane of the egg. The sperm head was then released into the ooplasm and the pipette carefully withdrawn from the egg.
In vitro fertilization and zona pellucida-free eggs
Mouse eggs were collected as above and as previously described following standard superovulation procedures (Navarrete et al., 2015). Up to four cumulus-oocyte complexes (COCs) were placed into a 90 μl drop of media (TYH standard medium) covered with mineral oil and previously equilibrated in an incubator with 5% CO2 at 37°C. Fertilization drops containing 20-30 eggs were inseminated with sperm from each genotype incubated as described above in medium supporting capacitation (final concentration of 1×106 cells/ml). Fertilization and polyspermy were evaluated using Hoechst staining at different hours post-insemination. In the case where [Ca2+]i monitoring was performed, following egg collection the zonae pellucidae were removed using acidic Tyrode's solution (pH 1.6) (Bernhardt et al., 2015). Zona pellucida-free eggs were adhered to Cell-Tak (MatTek)-treated glass-bottom dishes in 90 μl BSA-free TYH, and sperm were added to a final concentration of 105 sperm/ml, along with 4 μl TYH containing 30 mg/ml BSA for a final concentration of ∼1.5 mg/ml BSA. Imaging began immediately after sperm addition and was performed essentially as described below. Because the sperm from both Plcz1–/– males was cryopreserved, the control sperm was also cryopreserved and it was collected from a transgenic mouse line originally generated by Dr G. S. McKnight (University of Washington, WA, USA) (Morgan et al., 2008). Sperm penetration was confirmed by the presence of Ca2+ responses, as the large number of frozen sperm needed to achieve fertilization made it impossible to assess sperm entry.
Sperm penetration
Sperm-egg penetration analysis was performed 6 h post-in vitro fertilization. Briefly, C57BL/6J female mice at 3-4 weeks old (Charles River) were stimulated using 5 IU pregnant mare serum gonadotrophin (PMSG, Folligon, Intervets). Ovulation was induced 46-48 h later using 5 IU hCG (Chorulon, Intervet). Oocyte-granulosa cell complexes (MII eggs) were collected 13-14 h following hCG administration in a complete pre-warmed HTF medium (Zenith Biotech), and then inseminated with pre-capacitated sperm from each animal group under mineral oil (Zenith Biotech) at 37°C and 5% CO2. Six hours later, eggs were washed and treated for 60 s with acid Tyrode's solution to remove the extra sperm attached to the zona pellucida. The eggs were then fixed in 3.7% paraformaldehyde for 30 min, and washed and permeabilized before DNA staining with Hoechst 33324. Sperm DNA was detected using a 358⁄461 (ex/em.) laser line on a LSX8 Leica confocal scanning laser microscope. High-resolution image series on z plane were generated in order to build 3-dimensional (3D) images of the embryos. Counting of sperm heads inside the eggs confirmed by the DNA staining method, was carried out on 3D projections. Processing and analysis of the digital images were performed using Image-J/Fiji.
Microinjection of PLCz1 cRNA
Microinjections were performed as described previously (Lee et al., 2016). cRNA was prepared as previously described and 1-2 µl used to prepare micro drops from which glass micropipettes were loaded by aspiration. cRNAs were delivered into eggs and/or zygotes by pneumatic pressure (PLI-100 picoinjector, Harvard Apparatus). Each egg received 5-10 pl, ∼1-3% of the total volume of the egg. Injection of cRNA into zygotes was performed 3 h after completion of ICSI.
Calcium imaging
This was carried out in two different ways. Using one approach, eggs were pre-incubated for 30 min at 37°C in KSOM containing the Ca2+-sensitive fluorescent dye Fura-2 acetoxymethyl ester (7.5 µM) (Fura-2 AM, Invitrogen). After ICSI, eggs were transferred to a 10 µl KSOM droplet in a glass dish (MatTek) for Ca2+ analysis. Imaging was performed on an inverted epifluorescence microscope (Nikon Eclipse TE 300, Analis) using a 20× objective and filter switch (Lambda DG-4 filter switch, Sutter Instrument), for excitation at 340 and 380 nm corresponding to Fura-2 AM spectrum. Images were acquired every 5 s for a duration of 2 h and intracellular Ca2+ changes were registered as the ratio of fluorescence (340/380 nm). In another approach, [Ca2+]i monitoring was performed as previously described (Lee et al., 2016), also using Fura 2-AM.
In vitro development of in vivo fertilized eggs
Three- to 4-week-old C57Bl/6J female mice (Charles River) were injected intraperitoneally with 5 IU of PMSG (Intervet) and 48 h later with 5 IU of hCG (Intervet), and paired with male mice. Zygotes were retrieved from oviductal ampullae at 18 h post-hCG. Cumulus-enclosed zygotes were denuded by exposure to 1 mg/ml hyaluronidase (Sigma) in modified HTF (Zenith Biotech) for 3-6 min, washed through fresh modified HTF and cultured in KSOMaa in the presence of 5% CO2 at 37°C. For assessment of polyspermy, denuded and washed zygotes were cultured in KSOMaa in the presence of 5% CO2 at 37°C for 30 min. The zygotes were then placed in fresh modified HTF containing 1 µg/ml Hoechst 33342 and incubated for 20-30 min. They were washed in fresh modified HTF and imaged using a fluorescence microscope.
Statistical analyses
Where indicated, data are presented as average±s.e.m. and analysed by a one-way ANOVA with Bunnett's or Tukey's multiple comparison tests, or Student's t-test, as appropriate. In some figures, data are presented as the proportion of total eggs examined showing a specific characteristic from combined experiments; contingency tables with a Fisher's exact test were used in these cases. Graphs are annotated with the following conventions: ns, P>0.05; *P<0.05; **P<0.01; ***P<0.001.
Acknowledgements
We thank the technical staff of the University of Oxford, University of Massachusetts, and the University of Ghent Hospital Medical School, for their help in breeding and maintaining the mice used in this study.
Footnotes
Competing interests
The authors declare no competing or financial interests.
Author contributions
Conceptualization: A.H., J.G., M.R., H.C.L., M.F.B., G.A., A.B., S.F., F.N., B.H., R.F., J.P.; Methodology: A.H., J.G., M.R., H.C.L., M.F.B., G.A., A.B., S.F., F.N., B.H., R.F., J.P.; Validation: A.H., J.G., M.R., H.C.L., M.F.B., G.A., A.B., S.F., F.N.; Formal analysis: J.G., M.R., H.C.L., M.F.B., G.A., A.B., S.F., F.N.; Investigation: A.H., J.G., M.R., H.C.L., M.F.B., G.A., A.B., S.F., F.N.; Data curation: A.H., J.G., M.R., H.C.L., M.F.B., G.A., A.B., S.F., F.N.; Writing - original draft: J.P.; Writing - review & editing: M.R., B.H., R.F., J.P.; Supervision: P.d.S., B.H., R.F., J.P.; Project administration: J.P.; Funding acquisition: P.d.S., B.H., R.F., J.P.
Funding
Work in J.P.’s lab was funded by a DPhil grant to A.H. from the Iraqi Higher Committee for Education Development and by an Academic Scholarship Award grant from the Society for Reproduction and Fertility. Work in B.H.’s and P.d.S.’s labs was funded by a project funding grant from the Fonds Wetenschappelijk Onderzoek (FWO) (G060615N) and by a fundamental clinical research mandate from FWO to P.d.S. Work in R.F.’s lab was funded by a National Institutes of Health grant (HD051872). Deposited in PMC for release after 12 months.
Supplementary information
Supplementary information available online at http://dev.biologists.org/lookup/doi/10.1242/dev.150227.supplemental
References
- Aarabi M., Yu Y., Xu W., Tse M. Y., Pang S. C., Yi Y.-J., Sutovsky P. and Oko R. (2012). The testicular and epididymal expression profile of PLCzeta in mouse and human does not support its role as a sperm-borne oocyte activating factor. PLoS ONE 7, e33496 10.1371/journal.pone.0033496 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aarabi M., Balakier H., Bashar S., Moskovtsev S. I., Sutovsky P., Librach C. L. and Oko R. (2014). Sperm-derived WW domain-binding protein, PAWP, elicits calcium oscillations and oocyte activation in humans and mice. FASEB J. 28, 4434-4440. 10.1096/fj.14-256495 [DOI] [PubMed] [Google Scholar]
- Bassett A. and Liu J.-L. (2014). CRISPR/Cas9 mediated genome engineering in Drosophila. Methods 69, 128-136. 10.1016/j.ymeth.2014.02.019 [DOI] [PubMed] [Google Scholar]
- Bernhardt M. L., Lowther K. M., Padilla-Banks E., McDonough C. E., Lee K. N., Evsikov A. V., Uliasz T. F., Chidiac P., Williams C. J. and Mehlmann L. M. (2015). Regulator of G-protein signaling 2 (RGS2) suppresses premature calcium release in mouse eggs. Development 142, 2633-2640. 10.1242/dev.121707 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bianchi E., Doe B., Goulding D. and Wright G. J. (2014). Juno is the egg Izumo receptor and is essential for mammalian fertilization. Nature 508, 483-487. 10.1038/nature13203 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheng Y., Zhong Z. and Latham K. E. (2012). Strain-specific spontaneous activation during mouse oocyte maturation. Fertil. Steril. 98, 200-206. 10.1016/j.fertnstert.2012.03.060 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doudna J. A. and Charpentier E. (2014). Genome editing. The new frontier of genome engineering with CRISPR-Cas9. Science 346, 1258096 10.1126/science.1258096 [DOI] [PubMed] [Google Scholar]
- Ducibella T., Huneau D., Angelichio E., Xu Z., Schultz R. M., Kopf G. S., Fissore R., Madoux S. and Ozil J.-P. (2002). Egg-to-embryo transition is driven by differential responses to Ca(2+) oscillation number. Dev. Biol. 250, 280-291. 10.1006/dbio.2002.0788 [DOI] [PubMed] [Google Scholar]
- Escoffier J., Lee H. C., Yassine S., Zouari R., Martinez G., Karaouzène T., Coutton C., Kherraf Z.-E., Halouani L., Triki C. et al. (2016). Homozygous mutation of PLCZ1 leads to defective human oocyte activation and infertility that is not rescued by the WW-binding protein PAWP. Hum. Mol. Genet. 25, 878-891. 10.1093/hmg/ddv617 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferrer-Vaquer A., Barragan M., Freour T., Vernaeve V. and Vassena R. (2016). PLCzeta sequence, protein levels, and distribution in human sperm do not correlate with semen characteristics and fertilization rates after ICSI. J. Assist. Reprod. Genet. 33, 747-756. 10.1007/s10815-016-0718-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heytens E., Parrington J., Coward K., Young C., Lambrecht S., Yoon S.-Y., Fissore R. A., Hamer R., Deane C. M., Ruas M. et al. (2009). Reduced amounts and abnormal forms of phospholipase C zeta (PLCzeta) in spermatozoa from infertile men. Hum. Reprod. 24, 2417-2428. 10.1093/humrep/dep207 [DOI] [PubMed] [Google Scholar]
- Igusa Y. and Miyazaki S. (1986). Periodic increase of cytoplasmic free calcium in fertilized hamster eggs measured with calcium-sensitive electrodes. J. Physiol. 377, 193-205. 10.1113/jphysiol.1986.sp016181 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ito J., Parrington J. and Fissore R. A. (2011). PLCzeta and its role as a trigger of development in vertebrates. Mol. Reprod. Dev. 78, 846-853. 10.1002/mrd.21359 [DOI] [PubMed] [Google Scholar]
- Kashir J., Jones C., Lee H. C., Rietdorf K., Nikiforaki D., Durrans C., Ruas M., Tee S. T., Heindryckx B., Galione A. et al. (2011). Loss of activity mutations in phospholipase C zeta (PLCzeta) abolishes calcium oscillatory ability of human recombinant protein in mouse oocytes. Hum. Reprod. 26, 3372-3387. 10.1093/humrep/der336 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kashir J., Konstantinidis M., Jones C., Lemmon B., Lee H. C., Hamer R., Heindryckx B., Deane C. M., De Sutter P., Fissore R. A. et al. (2012). A maternally inherited autosomal point mutation in human phospholipase C zeta (PLCzeta) leads to male infertility. Hum. Reprod. 27, 222-231. 10.1093/humrep/der384 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kashir J., Buntwal L., Nomikos M., Calver B. L., Stamatiadis P., Ashley P., Vassilakopoulou V., Sanders D., Knaggs P., Livaniou E. et al. (2017). Antigen unmasking enhances visualization efficacy of the oocyte activation factor, phospholipase C zeta, in mammalian sperm. Mol. Hum. Reprod. 23, 54-67. 10.1093/molehr/gaw073 [DOI] [PubMed] [Google Scholar]
- Kline D. and Kline J. T. (1992). Repetitive calcium transients and the role of calcium in exocytosis and cell cycle activation in the mouse egg. Dev. Biol. 149, 80-89. 10.1016/0012-1606(92)90265-I [DOI] [PubMed] [Google Scholar]
- Knott J. G., Kurokawa M., Fissore R. A., Schultz R. M. and Williams C. J. (2005). Transgenic RNA interference reveals role for mouse sperm phospholipase Czeta in triggering Ca2+ oscillations during fertilization. Biol. Reprod. 72, 992-996. 10.1095/biolreprod.104.036244 [DOI] [PubMed] [Google Scholar]
- Knott J. G., Gardner A. J., Madgwick S., Jones K. T., Williams C. J. and Schultz R. M. (2006). Calmodulin-dependent protein kinase II triggers mouse egg activation and embryo development in the absence of Ca2+ oscillations. Dev. Biol. 296, 388-395. 10.1016/j.ydbio.2006.06.004 [DOI] [PubMed] [Google Scholar]
- Kurokawa M., Sato K., Wu H., He C., Malcuit C., Black S. J., Fukami K. and Fissore R. A. (2005). Functional, biochemical, and chromatographic characterization of the complete [Ca2+]i oscillation-inducing activity of porcine sperm. Dev. Biol. 285, 376-392. 10.1016/j.ydbio.2005.06.029 [DOI] [PubMed] [Google Scholar]
- Lee H. C., Yoon S.-Y., Lykke-Hartmann K., Fissore R. A. and Carvacho I. (2016). TRPV3 channels mediate Ca(2)(+) influx induced by 2-APB in mouse eggs. Cell Calcium 59, 21-31. 10.1016/j.ceca.2015.12.001 [DOI] [PubMed] [Google Scholar]
- Li L., Lu X. and Dean J. (2013). The maternal to zygotic transition in mammals. Mol. Aspects Med. 34, 919-938. 10.1016/j.mam.2013.01.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miao Y.-L., Stein P., Jefferson W. N., Padilla-Banks E. and Williams C. J. (2012). Calcium influx-mediated signaling is required for complete mouse egg activation. Proc. Natl. Acad. Sci. USA 109, 4169-4174. 10.1073/pnas.1112333109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morgan D. J., Weisenhaus M., Shum S., Su T., Zheng R., Zhang C., Shokat K. M., Hille B., Babcock D. F. and McKnight G. S., (2008). Tissue-specific PKA inhibition using a chemical genetic approach and its application to studies on sperm capacitation. Proc. Natl. Acad. Sci. USA 105, 20740-20745. 10.1073/pnas.0810971105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Navarrete F. A., García-Vázquez F. A., Alvau A., Escoffier J., Krapf D., Sánchez-Cárdenas C., Salicioni A. M., Darszon A. and Visconti P. E. (2015). Biphasic role of calcium in mouse sperm capacitation signaling pathways. J. Cell. Physiol. 230, 1758-1769. 10.1002/jcp.24873 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nomikos M., Kashir J., Swann K. and Lai F. A. (2013). Sperm PLCzeta: from structure to Ca2+ oscillations, egg activation and therapeutic potential. FEBS Lett. 587, 3609-3616. 10.1016/j.febslet.2013.10.008 [DOI] [PubMed] [Google Scholar]
- Ozil J.-P., Markoulaki S., Toth S., Matson S., Banrezes B., Knott J. G., Schultz R. M., Huneau D. and Ducibella T. (2005). Egg activation events are regulated by the duration of a sustained [Ca2+]cyt signal in the mouse. Dev. Biol. 282, 39-54. 10.1016/j.ydbio.2005.02.035 [DOI] [PubMed] [Google Scholar]
- Parrington J., Davis L. C., Galione A. and Wessel G. (2007). Flipping the switch: how a sperm activates the egg at fertilization. Dev. Dyn. 236, 2027-2038. 10.1002/dvdy.21255 [DOI] [PubMed] [Google Scholar]
- Phillips K. P., Petrunewich M. A. F., Collins J. L., Booth R. A., Liu X. J. and Baltz J. M. (2002). Inhibition of MEK or cdc2 kinase parthenogenetically activates mouse eggs and yields the same phenotypes as Mos(−/−) parthenogenotes. Dev. Biol. 247, 210-223. 10.1006/dbio.2002.0680 [DOI] [PubMed] [Google Scholar]
- Ran F. A., Hsu P. D., Lin C.-Y., Gootenberg J. S., Konermann S., Trevino A. E., Scott D. A., Inoue A., Matoba S., Zhang Y. et al. (2013). Double nicking by RNA-guided CRISPR Cas9 for enhanced genome editing specificity. Cell 154, 1380-1389. 10.1016/j.cell.2013.08.021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanders J. R. and Swann K. (2016). Molecular triggers of egg activation at fertilization in mammals. Reproduction 152, R41-R50. 10.1530/REP-16-0123 [DOI] [PubMed] [Google Scholar]
- Sanusi R., Yu Y., Nomikos M., Lai F. A. and Swann K. (2015). Rescue of failed oocyte activation after ICSI in a mouse model of male factor infertility by recombinant phospholipase Czeta. Mol. Hum. Reprod. 21, 783-791. 10.1093/molehr/gav042 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Satouh Y., Nozawa K. and Ikawa M. (2015). Sperm postacrosomal WW domain-binding protein is not required for mouse egg activation. Biol. Reprod. 93, 94 10.1095/biolreprod.115.131441 [DOI] [PubMed] [Google Scholar]
- Saunders C. M., Larman M. G., Parrington J., Cox L. J., Royse J., Blayney L. M., Swann K. and Lai F. A. (2002). PLC zeta: a sperm-specific trigger of Ca(2+) oscillations in eggs and embryo development. Development 129, 3533-3544. [DOI] [PubMed] [Google Scholar]
- Sfontouris I. A., Nastri C. O., Lima M. L. S., Tahmasbpourmarzouni E., Raine-Fenning N. and Martins W. P. (2015). Artificial oocyte activation to improve reproductive outcomes in women with previous fertilization failure: a systematic review and meta-analysis of RCTs. Hum. Reprod. 30, 1831-1841. 10.1093/humrep/dev136 [DOI] [PubMed] [Google Scholar]
- Shen B., Zhang W., Zhang J., Zhou J., Wang J., Chen L., Wang L., Hodgkins A., Iyer V., Huang X. et al. (2014). Efficient genome modification by CRISPR-Cas9 nickase with minimal off-target effects. Nat. Methods 11, 399-402. 10.1038/nmeth.2857 [DOI] [PubMed] [Google Scholar]
- Sobinoff A. P., Sutherland J. M. and McLaughlin E. A. (2013). Intracellular signalling during female gametogenesis. Mol. Hum. Reprod. 19, 265-278. 10.1093/molehr/gas065 [DOI] [PubMed] [Google Scholar]
- Suzuki T., Yoshida N., Suzuki E., Okuda E. and Perry A. C. F. (2010). Full-term mouse development by abolishing Zn2+-dependent metaphase II arrest without Ca2+ release. Development 137, 2659-2669. 10.1242/dev.049791 [DOI] [PubMed] [Google Scholar]
- Swann K. and Lai F. A. (2016). The sperm phospholipase C-zeta and Ca2+ signalling at fertilization in mammals. Biochem. Soc. Trans. 44, 267-272. 10.1042/BST20150221 [DOI] [PubMed] [Google Scholar]
- Vanden Meerschaut F., Nikiforaki D., Heindryckx B. and De Sutter P., (2014). Assisted oocyte activation following ICSI fertilization failure. Reprod. Biomed. Online 28, 560-571. 10.1016/j.rbmo.2014.01.008 [DOI] [PubMed] [Google Scholar]
- Wakai T., Vanderheyden V. and Fissore R. A. (2011). Ca2+ signaling during mammalian fertilization: requirements, players, and adaptations. Cold Spring Harb. Perspect. Biol. 3, a006767 10.1101/cshperspect.a006767 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whittingham D. G. and Siracusa G. (1978). The involvement of calcium in the activation of mammalian oocytes. Exp. Cell Res. 113, 311-317. 10.1016/0014-4827(78)90371-3 [DOI] [PubMed] [Google Scholar]
- Xu Z., Abbott A., Kopf G. S., Schultz R. M. and Ducibella T. (1997). Spontaneous activation of ovulated mouse eggs: time-dependent effects on M-phase exit, cortical granule exocytosis, maternal messenger ribonucleic acid recruitment, and inositol 1,4,5-trisphosphate sensitivity. Biol. Reprod. 57, 743-750. 10.1095/biolreprod57.4.743 [DOI] [PubMed] [Google Scholar]
- Yanagimachi R. and Chang M. C. (1961). Fertilizable life of golden hamster ova and their morphological changes at the time of losing fertilizability. J. Exp. Zool. 148, 185-203. 10.1002/jez.1401480303 [DOI] [PubMed] [Google Scholar]
- Yoon S.-Y. and Fissore R. A. (2007). Release of phospholipase C zeta and [Ca2+]i oscillation-inducing activity during mammalian fertilization. Reproduction 134, 695-704. 10.1530/REP-07-0259 [DOI] [PubMed] [Google Scholar]
- Yoshida N. and Perry A. C. F. (2007). Piezo-actuated mouse intracytoplasmic sperm injection (ICSI). Nat. Protoc. 2, 296-304. 10.1038/nprot.2007.7 [DOI] [PubMed] [Google Scholar]