

Abstract
We speculated that ubiquitin specific peptidase 4 (USP4) may deubiquitinate interferon regulatory factor 4 (IRF4) and affect T helper type 2 (Th2) cell function. This study aimed to validate this hypothesis. Here, the interaction between USP4 and IRF4 were analyzed by co-immunoprecipitation assay. The deubiquitin effect of USP4 on IRF4 was analyzed by the Ni-NTA pull down assay. Luciferase reporter gene constructs were used to analyze the effects of USP4, IRF4 and nuclear factor of activated T cell-2 (NFATc2) on the activation of the interleukin-4 (IL-4) promoter. Then, the Th2 cells were infected with sh-USP4 to analyze the effects of USP4 on the expression levels of IRF4 and Th2-related cytokines. Western blotting and RT-qPCR were used to detect the protein and mRNA expression levels, respectively. To determine the levels of IL-4 and IRF4 in rheumatic heart disease (RHD) patients, peripheral blood mononuclear cells (PBMCs) were separated by density gradient centrifugation from RHD patients and healthy controls, and flow cytometric analysis was performed. Our results validated the interaction between USP4 and IRF4, and effects of USP4 on stabilization and deubiquitination of IRF4 were also found. Importantly, USP4 and IRF4 synergized with NFATc2 to specifically enhance NFAT-mediated activation of the IL-4 promoter. USP4 knockdown not only decreased the expression level of IRF4, but also affected the expression level of Th2-related cytokines. Finally, the increased level of IL-4 and IRF4 in PBMCs of RHD patients were observed. On the whole, our data indicate that USP4 interacts with and deubiquitinates IRF4, and also stabilizes IRF4 protein and promotes IRF4 function to facilitate IL-4 expression in Th2 cells, which may be related to the pathological process of RHD.
Keywords: rheumatic heart disease, T helper type 2 cells, ubiquitin specific peptidase 4, interferon regulatory factor 4
Introduction
Rheumatic heart disease (RHD) is an autoimmune disease that arises following infection of the throat by S. pyogenes in young individuals (3–19 years old) and children, who present genetic components that confer susceptibility to this disease (1). Both in developing countries and China, RHD remains to be a major public health concern that contributes to significant cardiac-related mortality and morbidity (2). RHD accounts for up to 250,000 premature deaths every year in the world and is considered as a physical manifestation of poverty and social inequality (3). A previous study indicated that molecular mimicry mediates the cross-reactions between streptococcal antigens and human proteins. Several autoantigens including vimentin, cardiac myosin epitopes, and other intracellular proteins have been identified (4).
It is well-accepted that both T and B lymphocytes can recognize pathogens and self-antigens through different types of molecular mimicry (4). Particularly, the role of T lymphocytes in the development of RHD was previously described in the 1980s (5). The epitope spreading, molecular mimicry and T cell receptor degeneracy were demonstrated as mechanisms that mediate rheumatic heart lesions through the identification of streptococcal and heart-tissue proteins cross-reactive intralesional T cell clones (4). Cytokines are important secondary signals following an infection since they cause deleterious responses in patients with autoimmune disease and trigger effective immune responses in most individuals (1). Antigen-activated CD4+ T cells polarize to the T helper type 1 (Th1), Th2 or Th17 subsets, three subsets of T helper cytokines, depending on the cytokine secreted. Th1 can produce interleukin-2 (IL-2), interferon-γ (IFN-γ), and tumor necrosis factor-α (TNF-α), and is involved with the cellular immune response. Th2 cells can produce IL-4, IL-5 and IL-13 and mediate humoral and allergic immune responses. Th17 is a type of proinflammatory response mediated by IL-17. In addition, the cytokines, transforming growth factor-β (TGF-β), IL-6 and IL-23, are the factors that induce the Th17 lineage. Among them, a significant role for IL-4 in Th2 development was recognized early on in the analysis of the CD4+ T cell subset and was also further confirmed by in vitro experimentation (6,7). Moreover, a previous study suggested that mimicry between streptococcal antigens and heart-tissue proteins, combined with high inflammatory cytokines and low IL-4 production, lead to the development of autoimmune reactions and cardiac tissue damage in RHD patients (8).
Interferon regulatory factor 4 (IRF4) is an immune system-restricted interferon regulatory factor that is required for lymphocyte activation. Researchers have demonstrated that IRF4 may be a component of such a Th2-specific protein complex serving to enhance the function of nuclear factor of activated T cell-2 (NFATc2) via its physical interaction with NFATc2 at the IL-4 locus, indicating the important role of IRF4 in Th2 development (9). It seems that IRF4 may play an important role in RHD progression by regulating IL-4 production and Th2 development. Deubiquitinating enzymes (DUBs) are proteases that process ubiquitin or ubiquitin-like gene products, reverse the modification of proteins by a single ubiquitin(-like) protein, and remodel polyubiquitin(-like) chains on target proteins (10). The ubiquitin-specific proteases (USP) have more than 50 members and represent the largest subclass of DUBs (11). Like other USPs, USP4 also mediates the removal and processing of ubiquitin. USP4 binds TGF-activated kinase 1 (TAK1), TNF receptor-associated factor 2 (TRAF2) and TRAF6 for deubiquitination and negatively regulates TNF-α-induced nuclear factor NF-κB (NF-κB) activation (12,13). USP4 directly interacts with and deubiquitinates TGF-β type I receptor (TβRI), regulating TGF-β signaling by controlling TβRI levels at the plasma membrane (14). USP4 also interacts directly with and deubiquitinates ARF-BP1, leading to the stabilization of ARF-BP1 and subsequent reduction of p53 levels (15).
Here, we speculated that USP4 may deubiquitinate IRF4, and affect Th2 cell function and may be associated with the pathogenesis of RHD. This study aimed to validate this hypothesis.
Materials and methods
Human samples
Twenty RHD patients (average age, 50 years) were enrolled from July, 2013 to January, 2014 at the First Affiliated Hospital of Anhui Medical University (Anhui, China). Preoperative echocardiography and postoperative pathological research were used to confirm RHD. Patients with dysfunction of liver and kidney, coronary heart disease, infectious diseases, other autoimmune diseases and malignant tumors were excluded. Ten healthy volunteers that were randomly selected from a population-based sample living in the same region with the same age distribution as the patient group, were considered as controls. Five milliliters of peripheral blood was drawn from every patient and control. This study was approved by the Ethics Committee of Anhui Medical University and the Institute Pasteur of Shanghai, the Chinese Academy of Sciences.
Plasmids and antibodies
The plasmids, Myc-IRF4 and Ha-USP4 and His-ubiquitin plasmids used in this study were provided by Dr Li Bin from the Institute Pasteur of Shanghai. Anti-flag and anti-USP4 (cat. no. U0635MSDS) antibodies were purchased from Sigma-Aldrich (St. Louis, MO, USA). The anti-Ha (cat. no. sc-7392 AC) and anti-ubiquitin (P4D1; cat. no. sc-8017) antibodies were purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA). The anti β-actin antibody (cat. no. LK9001T) was purchased from Tianjin Sungene Biotech Co., Ltd. (Tianjin, China). PerCP/Cy5.5 anti-human CD45RA (cat. no. 304121), FITC anti-human CD4 (cat. no. 300506), PE anti-human CD25 (cat. no. 302606), Pb anti-human/mouse IRF4 (cat. no. 48-9858-82) was purchased from eBioscience (San Diego, CA, USA). PerCP/Cy5.5 anti-human IL-4 (cat. no. 500822) was purchased from BioLegend, Inc. (San Diego, CA, USA).
Flow cytometry-based cell sorting
Peripheral blood mono-nuclear cells (PBMCs) were prepared by density gradient centrifugation according to standard Ficoll-Hypaque procedures (Pharmacia Biotech, Uppsala, Sweden). PBMCs were diluted with phosphate-buffered saline (PBS) and were stained with FITC anti-human CD4, PE-anti-human CD25, PerCP/Cy5.5 anti-human CD45RA (BD Biosciences, San Jose, CA, USA) or isotype-matched controls. The concentration of antibodies in each flow tube was ~20 µl/1–1.5×108 million cells, and the single-dyed and the non-dyed groups were set at the same time. The antibody concentration was ~1 µl/l×106 cells (dissolved in 50 µl PBS). After incubation on ice for 40 min, the cells were washed, resuspended in fluorescence-activated cell sorter (FACS) staining buffer (BD Biosciences). Naïve CD4+ T cells were gated on CD4, CD25 and CD45RA. After that, cells were centrifuged for 10 min, and resuspended in X-VIV015 medium (Lonza Co., Basel, Switzerland), and were cultured in an incubator (Thermo Fisher Scientific, Wilmington, DE, USA), with an atmosphere of 37°C and 5% CO2.
CD4+ T cell differentiation
Naïve CD4+ cells were seeded at a density of 2×105/well in 96-well plates, and were cultured in DMEM, containing 10% fetal bovine serum (FBS). For Th2 cell differentiation, cells were simulated with CD3/CD28 beads (Invitrogen, Carlsbad, USA) and the skewing conditions were as follows: IL-2 (50 U/ml), 20 ng/ml rIL-4 and anti-IFN-γ (10 µg/ml; BioLegend, Inc.). The cells were incubated for 3 days and further experimental analysis was performed after 7 days.
Cell culture
293T cells were purchased from the American Type Culture Collection (ATCC, Manassas, VA, USA), and were cultured in Dulbecco's modified Eagle's medium (DMEM) (HyClone, Logan, UT, USA) containing 10% FBS. The naïve CD4+ T cells and Th2 cells were obtained from peripheral blood of healthy samples, and were cultured in X-VIVO15 medium (Lonza Co.) containing 10% human AB serum (Weihong Biotechnology Co., Ltd., Shanghai, China), 1% non-essential amino-acid, 1% glutamine, 1% sodium pyruvate, 1% penicillin and streptomycin (all from Gibco, Grand Island, NY, USA). The cells were subsequently cultured in an incubator (Thermo Scientific), with an atmosphere of 37°C and 5% CO2.
Cell transfection
293T cells were transfected with vectors using linear polyethyleneimine (PEI; Polysciences, Inc., Warrington, PA, USA). Cells (1×106) and plated in 6-well plates for 24 h before transfection. The plasmid was mixed with 100 µl OPTI-MEM (Gibco) low serum medium, and 12 µl of PEI was added and incubated for 10 min. The cell supernatant was then replaced with 900 µl fresh DMEM containing 10% FBS, and the transfection mixture was added, and then incubated in an incubator, with an atmosphere of 37°C and 5% CO2. After 4 h, the cell supernatant was replaced with 10% fresh FBS/fresh medium, and was cultured for 36–48 h for cell collection.
Transduction of shRNA lentiviruses
The lentiviral vector pLKO.1-puro was purchased from Addgene (Cambridge, MA, USA), and sh-USP41, sh-USP42 and sh-CK were obtained as described in a previous study (14). Helper plasmids were cotransfected with pLKO.1-based plasmids carrying shRNAs in 293T cells to package recombinant lentiviruses. HIV-1 virus titers were quantified by determining the TCID50 in 293T cells. Lentiviral vector supernatant was concentrated by centrifugation at 50,000 × g for 2.5 h directly after harvesting. Cell supernatants were harvested 48 h after transfection and were used to infect Th2 cells. Polybrene (8 µg/ml) was added and fresh culture solution was replaced on the next day. Cells were retrieved on day 7 for the detection of the transcriptional levels of USP4, IRF4, IL-4, IL-5, IL-10 and IL-13.
Co-immunoprecipitation
After 36–48 h of transfection, the cells was collected for co-immunoprecipitation analysis as previously described (16). Briefly, cells were washed, harvested, and centrifuged. The cell pellet was homogenized and cell lysis buffer was added. The homogenate was centrifuged, and the supernatant was incubated with anti-Ha/anti-Myc antibody and Protein A/G-Plus beads overnight. The resins were recovered and washed followed by centrifugation. The pellet was then washed twice with Tris-HCL (pH 7.5) containing 0.1% (w/v) SDS and 150 mM NaCl. After that, it was resuspended in 1X SDS loading buffer, heated at 100°C for 3 min and subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) followed by western blot analysis.
Ni-NTA pull down assay
Before being harvested for Ni-NTA pull down assay, the cells were treated with MG132 (10 nM) for 4 h to inhibit proteasomal degradation and prevent the ubiquitination of proteins. The cells were lysed with urea lysis buffer (8 M urea, 100 mM Na2HPO4, 10 mM Tris-HCl (pH 8.0), 10 mM imidazole 0.2% and Triton X-100) and fully disrupted by ultrasonication. After that, the cell lysate was incubated with Ni-NTA agarose beads for 3 h to obtain His-tag combined with Ni-NTA, followed by washing with wash buffer. Then, 2X SDS loading buffer was added to obtain samples for western blotting. The ubiquitination modification was analyzed using the indicated antibodies.
Construction of luciferase reporter gene constructs
The sequences of the promoter of IL-4 were cloned into pGL3 basic expression plasmid. pGL3-plasmid with promoter fragments were sequenced using automated DNA sequencing methods. 293T cells were transfected with pGL3 constructs along with 3 mg of pSV-β-galactosidase control vector. Lipofectamine 2000 (Invitrogen) was used according to the manufacturer's instructions. Luciferase and β-gal assays were conducted 24 h post-transfection using the Luciferase assay system and the β-galactosidase enzyme assay system (both from Promega, Madison, WI, USA), respectively.
Flow cytometric analysis
PBMCs were prepared by density gradient centrifugation from RHD patients and healthy controls according to standard Ficoll-Hypaque procedures (Pharmacia Biotech, Uppsala, Sweden). PBMCs were then treated with phorbol myristate acetate (PMA, 25 ng/ml) and ionomycin (1 µg/ml) for 4 h and PBMCs were stained and processed by flow cytometry.
Naïve CD4+ T cells were sorted using the cytometry-based analysis and were cultured for Th2 cell differentiation, and were treated with the inhibitor of USP4, vialinin A, for 12 h. Th2 cells were then treated with phorbol myristate acetate (PMA, 25 ng/ml) and ionomycin (1 µg/ml) for 4 h and processed by flow cytometry.
Western blot analysis
Following 36–48 h of cell transfection, cell lysates (30 mg/well) were subjected to sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to a polyvinylidine difluoride membrane (Millipore, Darmstadt, Germany). The blots were incubated with the primary antibodies overnight at 4°C. The membranes were washed and then incubated with horseradish peroxidase-conjugated secondary antibodies (Sigma-Aldrich). The immunoreactive proteins were visualized by the SuperSignal ECL (Pierce, Rockford, IL, USA) and the densities of the bands were analyzed by ImageJ software (National Institutes of Health, Bethesda, MD, USA).
Reverse transcription-quantitative PCR (RT-qPCR)
Total RNA was extracted from cells with TRIzol reagent (Invitrogen). The amount of each RNA sample with an A260/A280 ratio between 1.8 and 2.2 were considered for suitable use. cDNA was reverse-transcribed using PrimeScript® RT reagent kit (Takara, Tokyo, Japan). The primers used in this study were as follows: USP4 upstream, 5′-TCAGCCGCTATGTGAAACAG-3′ and downstream primer, 5′-GTGGTCTCACTGGGGTCATT-3′; actin upstream, 5′-CTCTTCCAGCCTTCCTTCCT-3′ and downstream primer, 5′-CAGGGCAGTGATCTCCTTCT-3′; IL-4 upstream, 5′-GCCACCATGAGAAGGACACT-3′ and downstream primer, 5′-ACTCTGGTTGGCTTCCTTCA-3′; IL-5 upstream, 5′-GGCACTGCTTTCTACTCATCGA-3′ and downstream primer, 5′-AGTTGGTGATTTTTATGTACAGGAACA-3′; IL-10 upstream, 5′-TGCAAAACCAAACCACAAGA-3′ and downstream primer, 5′-TCTC GGAGATCTCGAAGCAT-3′; IL-13 upstream, 5′-CTATGCATCCGCTCCTCAAT-3′ and downstream primer, 5′-GGTGATG TTGACCAGCTCCT-3′; IRF4 upstream, 5′-AGAACGAGGAGAAGAGCATC-3′ and downstream primer, 5′-CCTTTAAACAGTGCCCAAG-3′. Quantitative PCR was carried out with the SYBR® Premix Ex Taq™ kit (Takara). The 20-µl reaction mix consisted of 2 µl 30-fold diluted first-strand cDNA, 10 µl 2X SYBR® Premix Ex Taq™, 0.4 µl 10 µM forward and reverse primer, 0.4 µl 50X ROX Reference Dye and 6.8 µl DEPC-treated water. Reactions were performed in an ABI 7300 real-time quantitative instrument (Applied Biosystems, Foster City, CA, USA). Specificity of the PCR products was confirmed by melting curve analysis followed by the verification of the amplicon length on 1.5% agarose gels stained with ethidium bromide.
Statistical analysis
Data are expressed as the means ± standard error of the mean. Paired t-test was used for calculating statistically significant difference (p<0.05) by GraphPad Prism software (GraphPad Software Inc., San Diego, CA, USA).
Results
Interaction between USP4 and IRF4
Co-immunoprecipitation assays were performed to assess the functional interaction between USP4 and transcription factors GATA3, STAT6, IRF4 and PU.1. USP4 fused to the hemagglutinin (Ha) epitope (Ha-USP4) and Myc-Flag tagged -GATA3, -STAT6, -IRF4 or -PU.1 were ectopically coexpressed in 293T cells, and total protein lysates were prepared after stimulation with PMA and ionomycin. Immunoprecipitation was conducted with anti-Ha and immunoblotting was used to detect the expression level. Immunoprecipitation of Ha-USP4 using the anti-Ha antibody resulted in the co-immunoprecipitation of Myc-IRF4 as detected by immunoblotting with anti-Myc antibodies (Fig. 1A). These results demonstrated that USP4 and IRF4 interact directly with each other. Then, we conducted an immunoprecipitation assay using anti-Myc or anti-Ha separately. Compared with the cells that were transfected with Ha-labeled-USP4 or Myc-labeled-IRF4 alone, when Ha-USP4 and Myc-IRF4 were coexpressed, immunoprecipitation of Ha-USP4 using anti-Ha antibody resulted in the co-immunoprecipitation of Myc-IRF4 as detected by immunoblotting with anti-Myc antibodies. The association of USP4 and IRF4 was corroborated by reverse co-immunoprecipitation experiment using the anti-Myc antibody to immunoprecipitate IRF4 and anti-Ha to detect the associated Ha-IRF4 by western blot analysis (Fig. 1B). Th2 cell lysate was then immunoprecipitated separately by using normal IgG and anti-USP4. We found that immunoprecipitation of USP4 using the anti-USP4 antibody resulted in the co-immunoprecipitation of Myc-IRF4 as detected by immunoblotting with anti-Myc antibodies, indicating that USP4 interacts with IRF4 in Th2 cells (Fig. 1C).
Figure 1.

Interaction between ubiquitin specific peptidase 4 (USP4) and interferon regulatory factor 4 (IRF4). (A) IRF4 was co-immunoprecipitated by the anti-Ha antibody when Ha-USP4 and Myc-IRF4 were coexpressed. (B) IRF4 was co-immunoprecipitated by the anti-Ha antibody when Ha-USP4 and Myc-IRF4 were coexpressed, and USP4 was co-immunoprecipitated by the anti-Myc antibody when Ha-USP4 and Myc-IRF4 were coexpressed. (C) IRF4 was co-immunoprecipitated by the anti-USP4 antibody, indicating that USP4 interacts with IRF4 in Th2 cells. Flag-IRF4 and Ha-USP4 were co-transfected into 293T cells. After 48 h, the cells were retrieved, and 1 µg of anti-Ha or anti-Flag monoclonal antibody precipitate was added and immunoblotting was conducted in A and B. Th2 cells were amplified to 1×107, and co-immunoprecipitated with 1 µg of normal IgG or anti-USP4, then immunoblotting assay was conducted in C.
Effect of USP4 on stabilization of IRF4
We then aimed to test and verify the influence of USP4 on the stabilization of IRF4. Myc-IRF4 and different doses of Ha-USP4 were transfected into 293T cells, respectively, and immunoblotting was used. The results showed that the IRF4 protein expression level was influenced by USP4 in a dose-dependent manner (Fig. 2).
Figure 2.

Effect of ubiquitin specific peptidase 4 (USP4) on stabilization of interferon regulatory factor 4 (IRF4). IRF4 protein expression level was influenced by USP4 in a dose-dependent manner. Equivalent dose of Myc-IRF4 and different doses of Ha-USP4 were cotransfected into 293T cells. After 48 h, the cells were retrieved and the expression levels of USP4 and IRF4 were detected by immunoblotting.
USP4 deubiquitinates IRF4
In order to validate whether USP4 exerts its effects on IRF4 through deubiquitination, the coding sequence of ubiquitin and its mutants pIPHis-K48 only and K63 only were cloned into the pIP-His vector. Myc-IRF4, Ha-USP4, Ha-USP4C311A, His-ubiquitin and its mutants were transiently expressed in 293T cells, respectively, and Ni-NTA was used for purified precipitation and then was detected by immunoblotting analysis. The results showed that USP4 simultaneously mediated the deubiquitination of IRF4 on K48 and K63. The experimental results (Fig. 3) proved that USP4 can mediate the deubiquitination of IRF4 on K48 and K63 sequences (amino acid residue sequence), revealing that USP4 exerts its effects on IRF4 through deubiquitination.
Figure 3.

Deubiquitination of interferon regulatory factor 4 (IRF4) by ubiquitin specific peptidase 4 (USP4). USP4 can mediate deubiquitination of IRF4 on K48 and K63 sequences (amino acid residue sequences). 293T cells were cotransfected with Myc-IRF4, Ha-USP4, His-ubiquitin or His-ubiquitin mutant K63 only and K48 only. Cells were collected after 48 h; MG132 (20 µM/ml) was used for processing and Ni-NTA was used for purified precipitation, and immunoblotting was conducted with monoclonal antibody of anti-Ha and anti-Flag.
USP4 and IRF4 synergize with NFATc2 to specifically enhance NFAT-mediated activation of the IL-4 promoter
To assess the ability of USP4 and IRF4 to transactivate the IL-4 promoter, either on their own or in conjunction with NFATc2, 293T cells were transfected with an IL-4 luciferase reporter construct. We found that cotransfection with USP4 and IRF4 had no significant effects on the luciferase activity of IL-4-luc (Fig. 4A), whereas cotransfection with USP4, IRF4 and NFATc2 markedly increased the luciferase activity of IL-4-luc compared with that of the controls (Fig. 4B and C). Naïve CD4+ T cells were sorted using flow cytometry-based analysis and were induced for Th2 cell differentiation, and were treated with the inhibitor of USP4, vialinin A, and then processed by flow cytometry. The results showed that the expression levels of IL-4 and IRF4 were decreased by the treatment of vialinin A (Fig. 4D).
Figure 4.
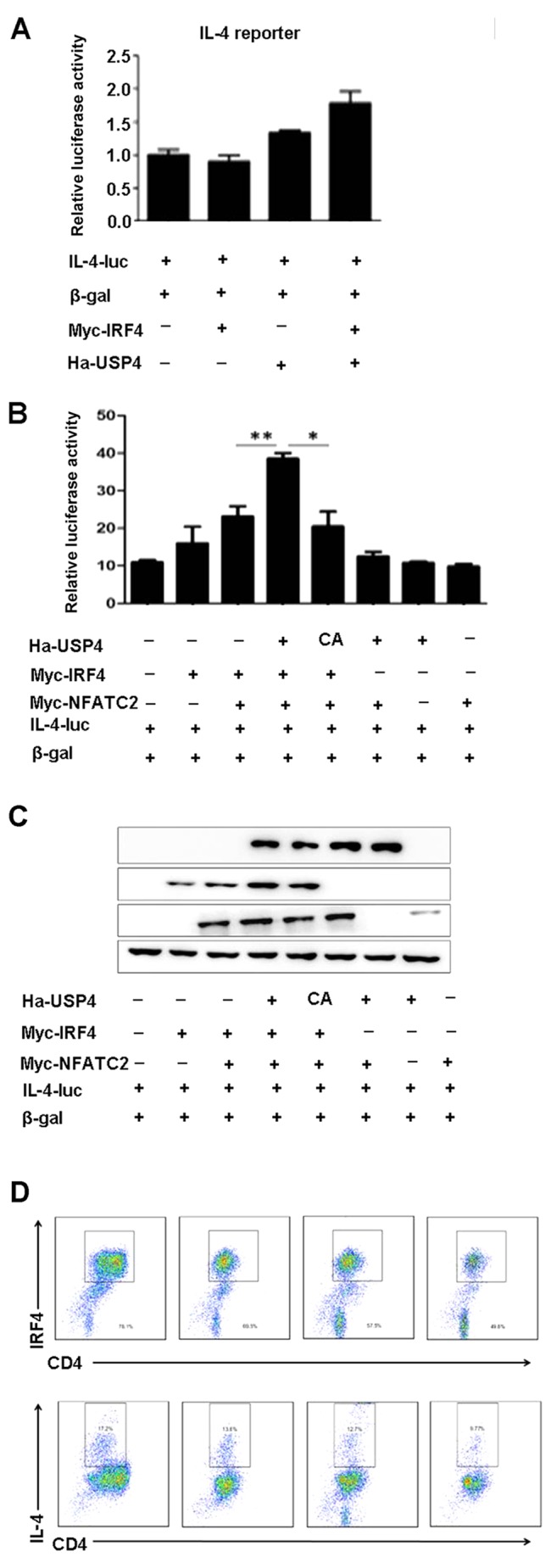
Ubiquitin specific peptidase 4 (USP4) and interferon regulatory factor 4 (IRF4) synergize with nuclear factor of activated T cell-2 (NFATc2) to specifically enhance NFAT-mediated activation of the interleukin-4 (IL-4) promoter. (A) Cotransfection with USP4 and IRF4 had no significant effects on the luciferase activity of IL-4-luc. (B and C) Cotransfection with USP4, IRF4 and NFATc2 markedly increased the luciferase activity of IL-4-luc compared with that of the controls. *p<0.05 and **p<0.01. (D) The expression levels of IL-4 and IRF4 were decreased by the treatment of vialinin A. Naïve CD4+ T cells were sorted using flow cytometry-based analysis and were cultured for Th2 cell differentiation, and were treated with the inhibitor of USP4, vialinin A, for 12 h. Th2 cells were then treated with phorbol myristate acetate (PMA, 25 ng/ml) and ionomycin (1 µg/ml) for 4 h and processed by flow cytometry.
USP4 knockdown affects the expression level of Th2-related cytokines
Since IRF4 is an important transcription factor of Th2 cells, we ascertained whether deubiquitinating enzyme USP4 can affect the function of Th2 cells. Sh-USP4-1, Sh-USP4-2 and the corresponding control were cloned into the lentivirus vector pLKO.1, and transfected into 293T cells, and then Th2 cells were infected in vitro. Firstly, we found that the mRNA and protein expression levels of USP4 were significantly decreased after knockdown of USP4. The results also showed that Sh-USP4 significantly decreased the mRNA and protein expression levels of IRF4 in Th2 cells as compared to the control (Fig. 5A). In addition, the transcription levels of Th2-related cytokines IL-4, IL-10 and IL-13 were significantly decreased, whereas the transcription level of IL-5 was significantly increased as compared to the control (Fig. 5B).
Figure 5.

Ubiquitin specific peptidase 4 (USP4) knockdown affects the expression level of Th2-related cytokines. (A) sh-USP4 significantly decreased the mRNA and protein expression levels of interferon regulatory factor 4 (IRF4) in Th2 cells as compared to the control. (B) The transcription levels of Th2-related cytokines interleukin-4 (IL-4), IL-10 and IL-13 were significantly decreased as compared to the control. Quantitative real-time PCR was applied for detecting the transcriptional level of the relevant genes (USP4, IRF4, IL-4, IL-5, IL-10 and IL-13). *p<0.05 and **p<0.01 compared to the Sh-CK group.
Increased levels of IL-4 and IRF4 in serum of RHD patients
To determine the levels of IL-4 and IRF4 in serum of RHD patients, we collected peripheral blood of healthy controls and RHD patients. PBMCs were separated using density gradient centrifugation, and flow cytometric analysis was performed. As shown in Fig. 6, when compared to the healthy controls, the levels of IL-4 (Fig. 6A) and IRF4 (Fig. 6B) in the serum were all increased in the RHD patient group.
Figure 6.

Expression levels of interleukin-4 (IL-4) and interferon regulatory factor 4 (IRF4) were increased in the rheumatic heart disease (RHD) patients. (A) The expression level of IL-4 was increased in RHD patients. (B) The expression level of IRF4 was increased in RHD patients. We collected peripheral blood of healthy controls and RHD patients. Peripheral blood mononuclear cells (PBMCs) were separated using density gradient centrifugation, and flow cytometric analysis was performed.
Discussion
RHD is a complex autoimmune inflammatory disease. IL-4 is a Th2-type cytokine and plays a regulatory role in the inflammatory response mediated by Th1 cytokines. Infiltrating mononuclear cells secreting inflammatory cytokines IFN-γ and TNF-α are found in valvular and myocardium tissues. In mononuclear cells, IL-10 and IL-4 (regulatory cytokines) are also secreted in myocardium tissue. However, in valvular tissue, only a few cells secrete IL-4, suggesting that these low numbers of IL-4-producing cells may contribute to the progression of valvular RHD lesions (17). In this study, when compared to the healthy control group, the level of IL-4 in the serum was significantly increased in the RHD group.
We also found that the level of IRF4 in the serum was significantly increased in the RHD group when compared to the healthy control group. IRF4 is clearly critical for lymphocyte function since IRF4-deficient mice exhibit cell intrinsic defects in both B and T cells (18). A previous study indicated that naïve Th cells lacking IRF4 are unable to differentiate into IL-4-producing Th2 effector cells in vitro (9). They also identified IRF4 and NFATc2 as transcriptional partners that together regulate inducible IL-4 gene expression upon T cell activation (9). The IRF4 protein (full-length 1-450) possesses an NH2-terminal (1-134) DNA binding domain, a activation domain region that includes a proline-rich segment, and the COOH-terminal contains the regulatory domain (19,20). The previous study has proven that the amino acids 150–340 and the COOH-terminal region of the IRF4 protein are important for transcriptional synergy as well its interaction with NFATc2.
Gao et al found that USP22 interacts with and deubiquitinates NFATc2, and also stabilizes NFATc2 protein and promotes NFATc2 function to facilitate IL-2 expression in T cells (21). Chen et al found that the ubiquitin ligase Stub1 negatively modulates regulatory T cell suppressive activity by promoting degradation of the transcription factor Foxp3 (22). Hou et al found that the E3 deubiquitinase USP21 is a positive regulator of GATA3 in asthma (23). Other studies indicate that the deubiquitinase USP17 regulates the stability and nuclear function of IL-33, and is also a positive regulator of retinoic acid-related orphan nuclear receptor γt (RORγt) in Th17 cells (24,25). In this study, we not only found an interaction between USP4 and IRF4, but also proved the effect of USP4 on the stabilization and deubiquitination of IRF4. Our results also showed that sh-USP4 significantly decreased the mRNA and protein expression levels of IRF4 in Th2 cells as compared to the control. In addition, the transcription levels of Th2-related cytokines IL-4, IL-10 and IL-13 were significantly decreased as compared to the control.
There are seven lysines in ubiquitin and poly-ubiquitin chains can be assembled through linkage to any of the seven. The distinct chains are known as K6, K11, K27, K29, K33, K48 and K63 chains, depending upon the lysine through which the monomers are linked (26). The experimental results in our study proved that USP4 can mediate deubiquitination of IRF4 on K48 and K63 sequences (amino acid residue sequences), revealing that USP4 exerts its effects on IRF4 through deubiquitination.
To conclude, this study indicated that USP4 interacts with and deubiquitinates IRF4, and also stabilizes IRF4 protein and promotes IRF4 function to facilitate IL-4 expression in Th2 cells, which may be associated with the pathological process of RHD. Small-molecule inhibitors of USP4 may have a potential role in providing a new therapeutic strategy for the treatment of RHD.
Acknowledgments
This study was in part supported by grants from the Key Scientific Research Project of Anhui Provincial Science and Technology Department (no. 1301043025).
References
- 1.Guilherme L, Kalil J. Rheumatic heart disease: Molecules involved in valve tissue inflammation leading to the autoimmune process and anti-S. pyogenes vaccine. Front Immunol. 2013;4:352–352. doi: 10.3389/fimmu.2013.00352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Chopra P, Gulwani H. Pathology and pathogenesis of rheumatic heart disease. Indian J Pathol Microbiol. 2007;50:685–697. [PubMed] [Google Scholar]
- 3.Rothenbühler M, O'Sullivan CJ, Stortecky S, Stefanini GG, Spitzer E, Estill J, Shrestha NR, Keiser O, Jüni P, Pilgrim T. Active surveillance for rheumatic heart disease in endemic regions: A systematic review and meta-analysis of prevalence among children and adolescents. Lancet Glob Health. 2014;2:e717–e726. doi: 10.1016/S2214-109X(14)70310-9. [DOI] [PubMed] [Google Scholar]
- 4.Guilherme L, Köhler KF, Kalil J. Rheumatic heart disease: Mediation by complex immune events. Adv Clin Chem. 2011;53:31–50. doi: 10.1016/B978-0-12-385855-9.00002-3. [DOI] [PubMed] [Google Scholar]
- 5.Köhler G, Milstein C. Continuous cultures of fused cells secreting antibody of predefined specificity. 1975. Biotechnology. 1992;24:524–526. [PubMed] [Google Scholar]
- 6.Le Gros G, Ben-Sasson SZ, Seder R, Finkelman FD, Paul WE. Generation of interleukin-4 (IL-4)-producing cells in vivo and in vitro: IL-2 and IL-4 are required for in vitro generation of IL-4-producing cells. J Exp Med. 1990;172:921–929. doi: 10.1084/jem.172.3.921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Hsieh CS, Heimberger AB, Gold JS, O'Garra A, Murphy KM. Differential regulation of T helper phenotype development by interleukins 4 and 10 in an alpha beta T-cell-receptor transgenic system. Proc Natl Acad Sci USA. 1992;89:6065–6069. doi: 10.1073/pnas.89.13.6065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Faé KC, Oshiro SE, Toubert A, Charron D, Kalil J, Guilherme L. How an autoimmune reaction triggered by molecular mimicry between streptococcal M protein and cardiac tissue proteins leads to heart lesions in rheumatic heart disease. J Autoimmun. 2005;24:101–109. doi: 10.1016/j.jaut.2005.01.007. [DOI] [PubMed] [Google Scholar]
- 9.Rengarajan J, Mowen KA, McBride KD, Smith ED, Singh H, Glimcher LH. Interferon regulatory factor 4 (IRF4) interacts with NFATc2 to modulate interleukin 4 gene expression. J Exp Med. 2002;195:1003–1012. doi: 10.1084/jem.20011128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Reyes-Turcu FE, Ventii KH, Wilkinson KD. Regulation and cellular roles of ubiquitin-specific deubiquitinating enzymes. Annu Rev Biochem. 2009;78:363–397. doi: 10.1146/annurev.biochem.78.082307.091526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Nijman SM, Luna-Vargas MP, Velds A, Brummelkamp TR, Dirac AM, Sixma TK, Bernards R. A genomic and functional inventory of deubiquitinating enzymes. Cell. 2005;123:773–786. doi: 10.1016/j.cell.2005.11.007. [DOI] [PubMed] [Google Scholar]
- 12.Xiao N, Li H, Luo J, Wang R, Chen H, Chen J, Wang P. Ubiquitin-specific protease 4 (USP4) targets TRAF2 and TRAF6 for deubiquitination and inhibits TNFα-induced cancer cell migration. Biochem J. 2012;441:979–986. doi: 10.1042/BJ20111358. [DOI] [PubMed] [Google Scholar]
- 13.Fan YH, Yu Y, Mao RF, Tan XJ, Xu GF, Zhang H, Lu XB, Fu SB, Yang J. USP4 targets TAK1 to downregulate TNFα-induced NF-κB activation. Cell Death Differ. 2011;18:1547–1560. doi: 10.1038/cdd.2011.11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Zhang L, Zhou F, Drabsch Y, Gao R, Snaar-Jagalska BE, Mickanin C, Huang H, Sheppard KA, Porter JA, Lu CX, et al. USP4 is regulated by AKT phosphorylation and directly deubiquitylates TGF-β type I receptor. Nat Cell Biol. 2012;14:717–726. doi: 10.1038/ncb2522. [DOI] [PubMed] [Google Scholar]
- 15.Zhang X, Berger FG, Yang J, Lu X. USP4 inhibits p53 through deubiquitinating and stabilizing ARF-BP1. EMBO J. 2011;30:2177–2189. doi: 10.1038/emboj.2011.125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Zhu C, Zheng F, Sun T, Duan Y, Cao J, Feng H, Shang L, Zhu Y, Liu H. Interaction of avian influenza virus NS1 protein and nucleolar and coiled-body phosphoprotein 1. Virus Genes. 2013;46:287–292. doi: 10.1007/s11262-012-0849-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Guilherme L, Cury P, Demarchi LM, Coelho V, Abel L, Lopez AP, Oshiro SE, Aliotti S, Cunha-Neto E, Pomerantzeff PM, et al. Rheumatic heart disease: Proinflammatory cytokines play a role in the progression and maintenance of valvular lesions. Am J Pathol. 2004;165:1583–1591. doi: 10.1016/S0002-9440(10)63415-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Mittrücker HW, Matsuyama T, Grossman A, Kündig TM, Potter J, Shahinian A, Wakeham A, Patterson B, Ohashi PS, Mak TW. Requirement for the transcription factor LSIRF/IRF4 for mature B and T lymphocyte function. Science. 1997;275:540–543. doi: 10.1126/science.275.5299.540. [DOI] [PubMed] [Google Scholar]
- 19.Eisenbeis CF, Singh H, Storb U. Pip, a novel IRF family member, is a lymphoid-specific, PU.1-dependent transcriptional activator. Genes Dev. 1995;9:1377–1387. doi: 10.1101/gad.9.11.1377. [DOI] [PubMed] [Google Scholar]
- 20.Brass AL, Kehrli E, Eisenbeis CF, Storb U, Singh H. Pip, a lymphoid-restricted IRF, contains a regulatory domain that is important for autoinhibition and ternary complex formation with the Ets factor PU.1. Genes Dev. 1996;10:2335–2347. doi: 10.1101/gad.10.18.2335. [DOI] [PubMed] [Google Scholar]
- 21.Gao Y, Lin F, Xu P, Nie J, Chen Z, Su J, Tang J, Wu Q, Li Y, Guo Z, et al. USP22 is a positive regulator of NFATc2 on promoting IL-2 expression. FEBS Lett. 2014;588:878–883. doi: 10.1016/j.febslet.2014.02.016. [DOI] [PubMed] [Google Scholar]
- 22.Chen Z, Barbi J, Bu S, Yang HY, Li Z, Gao Y, Jinasena D, Fu J, Lin F, Chen C, et al. The ubiquitin ligase Stub1 negatively modulates regulatory T cell suppressive activity by promoting degradation of the transcription factor Foxp3. Immunity. 2013;39:272–285. doi: 10.1016/j.immuni.2013.08.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Hou X, Zhang J, Chen C, Li B, Shi G. Identification of the E3 deubiquitinase USP21 as a positive regulator Of GATA3 in Asthma. Am J Respir Crit Care Med. 2012;185:A6478. [Google Scholar]
- 24.Ni Y, Tao L, Chen C, Song H, Li Z, Gao Y, Nie J, Piccioni M, Shi G, Li B. The Deubiquitinase USP17 Regulates the Stability and Nuclear Function of IL-33. Int J Mol Sci. 2015;16:27956–27966. doi: 10.3390/ijms161126063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Han L, Yang J, Wang X, Wu Q, Yin S, Li Z, Zhang J, Xing Y, Chen Z, Tsun A, et al. The E3 deubiquitinase USP17 is a positive regulator of retinoic acid-related orphan nuclear receptor γt (RORγt) in Th17 cells. J Biol Chem. 2014;289:25546–25555. doi: 10.1074/jbc.M114.565291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Dikic I, Robertson M. Ubiquitin ligases and beyond. BMC Biol. 2012;10:22. doi: 10.1186/1741-7007-10-22. [DOI] [PMC free article] [PubMed] [Google Scholar]