

Abstract
Although histone modifications are associated with gene activities, studies of their causal relationships have been difficult. For this purpose, we developed an inducible system integrating dCas9-based targeting and chemically induced proximity technologies to allow small molecule induced recruitment of P300 acetyltransferase and the acetylation of H3K27 at precise gene loci in cells. Employing the new technique, we elucidated the temporal order of histone acetylation and gene activation, as well as the stability of the installed histone modification.
TOC image
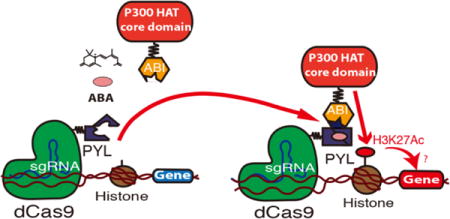
The histone tails are dynamically modified by histone modifying enzymes.1 These epigenetic modifications alter the compactness and/or surface of chromatin to modulate interactions with transcription regulators that control gene activity.2 It is proposed that combinations of histone posttranslational modifications (PTMs) form “histone codes” that guide gene expression.3 Although clear correlations exist between gene activity and histone PTMs,4 it is difficult to establish the causal relationships because tools are not available for precise temporal and gene locus specific editing of chosen histone PTMs. Because genetic or pharmacological interventions of histone modifying proteins leads to global changes in the epigenetic landscape, it is difficult to pinpoint whether a change in gene activity is a consequence of a local epigenetic modification or secondary effects arising from changes at remote genome loci.
The CRISPR/Cas9 system has become a powerful tool for genome editing.5 A catalytically inactive Cas9 (dCas9) enables targeting of dCas9-fused transcriptional regulators to defined genome loci by customized single guide RNAs (sgRNAs).6 Inducible Cas9/dCas9 systems have been developed to achieve temporal control of genome editing and gene activity.7 Although histone modifying proteins have also been fused to dCas9 to edit specific histone PTMs at chosen loci,8 these methods lack temporal control needed to investigate the dynamics of epigenetic regulation. Herein, we describe a new small molecule inducible system that integrates chemically induced proximity (CIP)9 and dCas9 methods to achieve temporally specific acetylation of lysine 27 on histone H3 (H3K27Ac) through gene locus specific recruitment of the catalytic acetyltransferase (HAT) domain of P300,10 which acetylates H3K27.11
To develop a new technique that temporally recruits P300 HAT to a chosen gene locus, we employed the abscisic acid (ABA)-based CIP system in which ABA induces rapid binding between PYL and ABI proteins.12 By linking PYL and ABI individually to dCas9 and the P300 HAT domain, P300 HAT can be recruited by ABA to specific genome loci targeted by customized sgRNAs (Fig. 1). Although chemical inducible systems for similar purposes have been reported,13 they rely on genome-targeting methods that are less easily customized. Consequently, this new system employing a readily customizable dCas9-based platform should facilitate epigenome editing. The reversible nature of the ABA system12 allows the rapid release of the recruited P300 HAT, which enables the studies on the stability and memory of the artificially established epigenetic environment.
Figure 1.
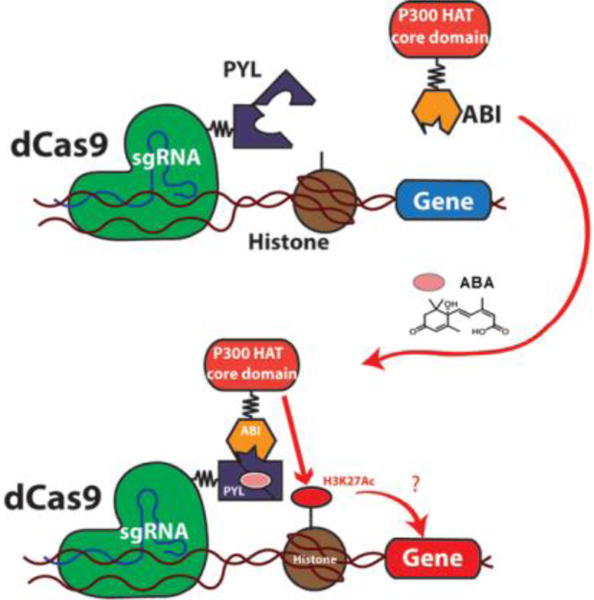
Small molecule inducible acetylation of H3K27.
To achieve inducible epigenome editing, we fused PYL to dCas9, tagged with a nuclear localization sequence (NLS), and a HA tag to enable chromatin immunoprecipitation (ChIP) and western blotting analysis. ABI was linked to the P300 HAT domain with a FLAG tag and NLS (Fig 2a). Both dCas9-PYL-HA and FLAG-ABI-P300 fusion proteins were expressed robustly in human HEK293T cells (Fig. S1). Four sgRNAs were used simultaneously to target the promoter region (within 200 bp from the transcription start site, TSS) of the IL1RN gene (Fig 2b and Table S1).8b The IL1RN locus was selected for testing in HEK293T cells because of its low H3K27Ac level and the low expression level of IL1RN.8b To monitor localization of dCas9-PYL-HA, FLAG-ABI-P300 and the level of H3K27Ac by using ChIP assays, 9 pairs of primers were designed to cover a 3.2 kb-region at the IL1RN locus (Fig 2b and Table S2).
Figure 2.
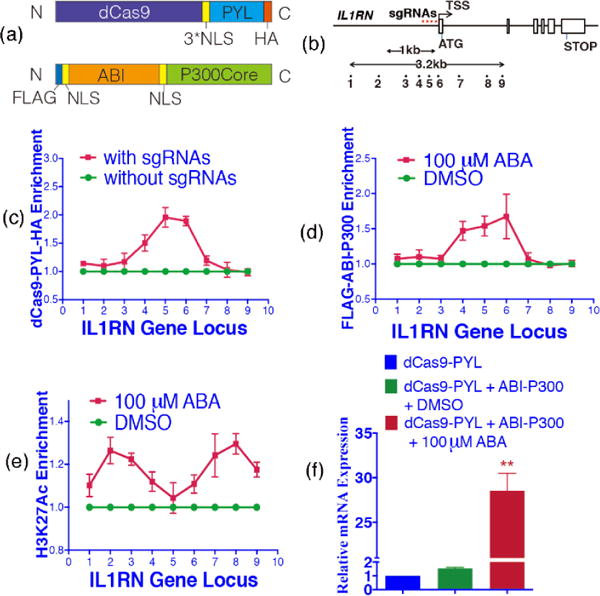
Small molecule induced acetylation of H3K27 at IL1RN locus. (a) Constructs of dCas9-PYL-HA and FLAG-ABI-P300. (b) The sites targeted by sgRNAs (red stars) and locations probed by qPCR amplicon (black solid circles) at the IL1RN locus. (c) Enrichment of dCas9-PYL-HA at the IL1RN locus with or without sgRNAs. Enrichment of FLAG-ABI-P300 (d) and H3K27Ac (e) at the IL1RN locus with or without ABA treatment. (f) Levels of IL1RN mRNA. ** represents P<0.01 compared with control group. Fold changes were calculated by comparing to cells treated without sgRNA (c), with DMSO (d, e) or transfected with dCas9-PYL (f). Error bars represent ± s.e.m. from biological replicates (n = 5).
To show that dCas9-PYL-HA is targeted to the IL1RN locus, HEK293T cells were transfected with plasmids encoding dCas9-PYL-HA with or without the co-transfection of IL1RN-specific sgRNAs and cultured for 24 h. Cells were then harvested and subjected to ChIP (using anti-HA antibody) and qPCR analysis in order to quantify the level of dCas9-PYL-HA at the IL1RN locus. The results show that, as expected, dCas9-PYL-HA is enriched over a narrow area that coincides with the sgRNA-binding locus (primer sets 4 to 6) (Fig. 2c).
To examine the recruitment of FLAG-ABI-P300, HEK293T cells were co-transfected with IL1RN sgRNAs, dCas9-PYL-HA and FLAG-ABI-P300 plasmids for 24 h. Cells were then either treated or not treated with 100 μ M ABA for another 24 h before being harvested. Portions of the cell lysates were analyzed by using ChIP (with anti-FLAG antibody) and qPCR assays to quantify the level of FLAG-ABI-P300. We observed that the addition of ABA leads to an increase in FLAG-ABI-P300 level at the IL1RN locus (Fig. 2d). To determine if the level of H3K27Ac changes in response to the FLAG-ABI-P300 localization, portions of the cell lysates produced in the above experiment were subjected to ChIP (using anti-H3K27Ac antibody) and qPCR assays to quantify the H3K27Ac enrichment. The results show that H3K27Ac levels increase at the IL1RN locus 24 h after ABA addition, and that this increase is not observed in lysates of cells that were not treated with ABA (Fig. 2e).
To study how IL1RN expression levels change, total RNAs from the cell lysates generated in the above experiment were purified and then subjected to qPCR analysis to quantify mRNA levels. The results show the IL1RN mRNA level from cells transfected with both ABI-P300 and dCas9-PYL is similar to the level of the one from cells transfected with dCas9-PYL only. This finding suggests that the overexpression of the P300 HAT without localizing to the IL1RN locus did not lead to IL1RN expression (Fig. 2f). On the other hand, the IL1RN mRNA level increases when transfected cells were treated with ABA (Fig. 2f), indicating that the increase in H3K27Ac activates IL1RN expression. Furthermore, when a P300 mutant (D1399Y) lacking the acetyltransferase activity8b was used, the IL1RN mRNA level did not increase upon ABA addition (Fig. S2), indicating that IL1RN expression directly depends on H3K27 acetylation. The observations described above demonstrate that the ABA-inducible system enables installation of H3K27Ac at the IL1RN locus and that the consequent increase in acetylation leads to an increased level of IL1RN expression.
To determine if the new method is applicable to modifying H3K27Ac at other gene loci, we designed sgRNAs that target the promoter region of other genes with low expression levels in HEK293T cells, including GRM2, HBA and MYOD1 (Table S1). The results of ChIP-qPCR assays employing primers designed to cover these gene loci (Fig. S3a, S4a, S5a and Table S2) show that in the presence of the corresponding sgRNAs, dCas9-PYL-HA is enrichment at each targeted gene locus (Fig. S3b, S4b and S5b) and ABA addition causes the enrichment of FLAG-ABI-P300 (Fig. S3c, S4c and S5c) and H3K27Ac (Fig. S3d, S4d and S5d). The level of mRNAs in each case also increases correspondingly (Fig. S3e, S4e and S5e). However, it is unclear at this moment the reasons that the observed H3K27Ac enrichment patterns were different at the 4 examined gene loci, which likely depend on the existing local epigenetic environments.
It is known that sgRNA/Cas9 can bind to unintended loci, which leads to off-target effects.14 To examine if aberrant binding interferes with the interpretation of results, we checked all reported sites in the genome that could potentially be targeted by the IL1RN sgRNAs used in our studies.15 We monitored changes in the mRNA levels of 6 genes having TSS close to the off-target sites (Table S3) after cells were co-transfected with IL1RN sgRNAs, dCas9-PYL-HA and FLAG-ABI-P300 plasmids followed by ABA treatment for 24 h. The observation that none of the 6 genes display increases in mRNA levels (Fig. S6) indicates that the off-target acetylation and gene activation are not significant in the studied cases. In addition, when sgRNAs designed for GRM2, HBA or MYOD1 were used in place of those for IL1RN, no increase in IL1RN mRNA was observed (Fig. S7), which demonstrates that the H3K27Ac enrichment at the IL1RN locus and gene expression are specific to IL1RN sgRNAs.
A significant advantage of this inducible system is that it possesses temporal control, which enables precise monitoring of the occurrence and progression of histone modification and gene expression events. To demonstrate this feature, we examined the relative timing of changes occurring in the levels of localized FLAG-ABI-P300, enriched H3K27Ac and expressed IL1RN mRNA after ABA induction. We confirmed that there is a significant level of sgRNA-dependent enrichment of dCas9-PYL-HA at the IL1RN locus at 24 h after transfection, which is the time point that we began adding ABA (Fig. S8). Within 1 h after ABA addition, FLAG-ABI-P300 is rapidly recruited to the IL1RN locus where it reaches a maximal level after 12 h (Fig. 3a). Correspondingly, the level of H3K27Ac begins to rise 1 h after ABA addition and continues to increase until 48 h (Fig. 3b). Finally, the mRNA level increases 4 h after ABA addition and reaches a maximum after 24 h (Fig. 3c).
Figure 3.
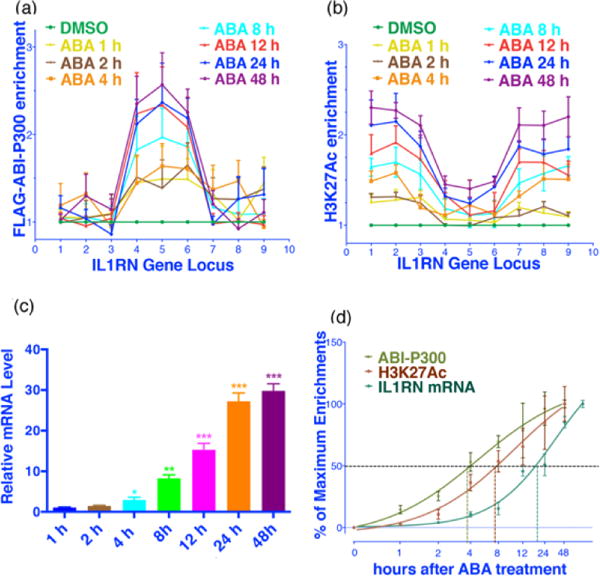
The time course of H3K27 acetylation and IL1RN expression. Enrichment of FLAG-ABI-P300 (a) and H3K27Ac (b) at the IL1RN locus at indicated time points after ABA treatment. (c) Levels of IL1RN mRNA at indicated time points after ABA treatment. ** represents P<0.01 compared with DMSO group; *** represents P<0.001 compared with DMSO group. Fold changes were calculated by comparing to cells treated with DMSO (a, b, c). (d) The time required to reach 50% maximum enrichments of FLAG-ABI-P300 (at primer 5 locus) and H3K27Ac (at primer 1 locus) at the IL1RN locus, and IL1RN mRNA expression. Error bars represent ± s.e.m. from biological replicates (n = 6; a, b, d or 8; c).
The time-dependent increases of IL1RN mRNA, and enrichments of FLAG-ABI-P300 and H3K27Ac at each of the probed loci were subjected to nonlinear regression analysis in order to elucidate the order of FLAG-ABI-P300 localization, H3K27Ac enrichment and IL1RN expression. A comparison of the time periods required to reach 50% maximum enrichments shows that FLAG-ABI-P300 localization takes place before H3K27Ac enrichment and that these events are followed by IL1RN mRNA expression (Fig. 3d and Table S4). The same analysis for the enrichments of H3K27Ac at each probed locus revealed that a longer time is required for the level of H3K27Ac to increase at loci farthest from the FLAG-ABI-P300 site (primer 1 and 9) than the closer sites (primer 3 and 7) (Fig. S9 and Table S4). This finding suggests a possibly spreading mechanism for H3K27Ac installation from the nucleation site at the IL1RN locus. Whether the same spreading phenomenon exists at the other loci remains to be studied.
Another major advantage of the new ABA inducible epigenome editing system is that it is readily reversible,12 which can be employed to remove an inducing factor rapidly so that the stability of artificially created histone PTMs can be monitored in a temporal specific manner. To determine the stability of H3K27Ac installed at the IL1RN locus after FLAG-ABI-P300 removal, HEK293T cells were co-transfected with IL1RN sgRNAs, dCas9-PYL-HA and FLAG-ABI-P300 plasmids for 24 h and then treated with ABA for another 12, 24 or 48 h. Cells were then washed with a non-ABA containing media (10 min for 3 times, or not washed as negative control) and then subjected to another 1 to 24 h culturing period. Cells were then harvested at different time points and assayed using ChIP and qPCR to monitor the levels of FLAG-ABI-P300, H3K27Ac and IL1RN mRNA. The results show that FLAG-ABI-P300 rapidly decreases to a near background level within 1 h following washing under all treatment conditions (Fig. 4a and S10). However, we observed that a longer ABA incubation time (48 h) increases the stability of H3K27Ac (Fig. 4b). A comprehensive time course analysis of H3K27Ac levels after ABA removal revealed that the longer cells were treated with ABA, the slower H3K27Ac was removed after washing (Fig. 4c and S10). The reasons why the stabilities of installed H3K27Ac are governed by ABA induction periods are not known at this point. Finally, an additional experiment demonstrated that changes of IL1RN mRNA levels after washing under conditions described above are well correlated with the changes in H3K27Ac levels (Fig. 4d), which further confirms the causal relationship between H3K27Ac and IL1RN activation. We also checked the reversibility of H3K27Ac at the GRM2 locus and observed that H3K27Ac enrichment was rapidly induced upon ABA addition and reversed after ABA removal (Fig. S11).
Figure 4.
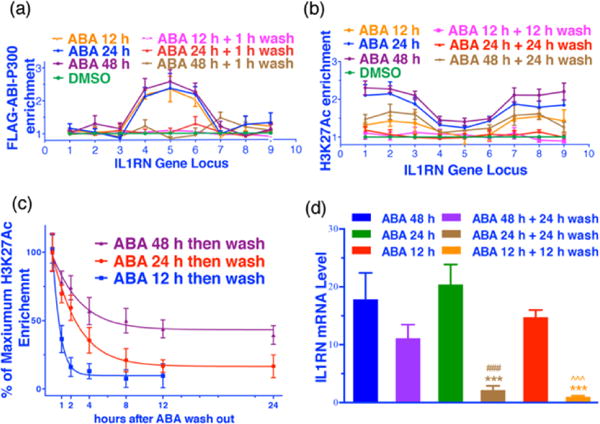
The reversibility of H3K27 acetylation and IL1RN expression. Enrichments of FLAG-ABI-P300 at primer 5 locus (a) and H3K27Ac at primer 1 locus (b) at the IL1RN locus at indicated time points after ABA treatment with or without washing. (c) Reversing rate of H3K27Ac at the IL1RN locus after ABA removal. (d) Levels of IL1RN mRNA at indicated time points after ABA treatment, or after washing. ### represents P<0.001 compared with 48 h; *** represents P<0.001 compared with 24 h; ˆˆˆ represents P<0.001 compared with 12 h. Fold enrichments were calculated by comparing to DMSO controls. Error bars represent ± s.e.m. from biological replicates (n = 6; a, b or n = 8; c).
The last phase of this effort was aimed at demonstrating precise temporal control in histone PTM editing by this method. For this purpose, HEK293T cells were co-transfected with IL1RN sgRNAs, dCas9-PYL-HA and FLAG-ABI-P300 plasmids for 24 h before adding ABA. Cells were then incubated for 12 h with ABA, washed, and incubated for another 12 h period in the absence of ABA. A second dose of ABA was then added to cells, which were incubated for another 12 h before they were harvested and their H3K27Ac and mRNA levels analyzed. The results show that the H3K27Ac level can be tightly controlled temporally at the IL1RN locus and that the IL1RN mRNA levels change accordingly (Fig. S12).
In summary, we have demonstrated a new strategy, which combines the ABA-based CIP method with the dCas9 system, for temporally-controlled (both induction and reversion) and gene locus-specific editing of H3K27Ac. The observations made in this effort show that H3K27Ac leads to an increased activity of the IL1RN gene. We anticipate that the new strategy will be applicable to studies of other histone modifying or chromatin remodeling proteins at other target genes that are aimed at elucidating the functions and mechanisms of diverse epigenetic regulation events.
Supplementary Material
Acknowledgments
This work was supported by the National Institutes of Health R21 HG008776.
Footnotes
Notes
The authors declare no competing financial interest.
References
- 1.Berger SL. Nature. 2007;447:407. doi: 10.1038/nature05915. [DOI] [PubMed] [Google Scholar]
- 2.Kouzarides T. Cell. 2007;128:693. doi: 10.1016/j.cell.2007.02.005. [DOI] [PubMed] [Google Scholar]
- 3.Strahl BD, Allis CD. Nature. 2000;403:41. doi: 10.1038/47412. [DOI] [PubMed] [Google Scholar]
- 4.Zhou VW, Goren A, Bernstein BE. Nat Rev Genet. 2011;12:7. doi: 10.1038/nrg2905. [DOI] [PubMed] [Google Scholar]
- 5.(a) Jinek M, Chylinski K, Fonfara I, Hauer M, Doudna JA, Charpentier E. Science. 2012;337 doi: 10.1126/science.1225829. [DOI] [PMC free article] [PubMed] [Google Scholar]; (b) Cong L, Ran FA, Cox D, Lin S, Barretto R, Habib N, Hsu PD, Wu X, Jiang W, Marraffini LA, Zhang F. Science. 2013;339:819. doi: 10.1126/science.1231143. [DOI] [PMC free article] [PubMed] [Google Scholar]; (c) Mali P, Yang L, Esvelt KM, Aach J, Guell M, DiCarlo JE, Norville JE, Church GM. Science. 2013;339:823. doi: 10.1126/science.1232033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.(a) Qi LS, Larson MH, Gilbert LA, Doudna JA, Weissman JS, Arkin AP, Lim WA. Cell. 2013;152:1173. doi: 10.1016/j.cell.2013.02.022. [DOI] [PMC free article] [PubMed] [Google Scholar]; (b) Sander JD, Joung JK. Nat Biotechnol. 2014;32:347. doi: 10.1038/nbt.2842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.(a) Zetsche B, Volz SE, Zhang F. Nat Biotechnol. 2015;33:139. doi: 10.1038/nbt.3149. [DOI] [PMC free article] [PubMed] [Google Scholar]; (b) Davis KM, Pattanayak V, Thompson DB, Zuris JA, Liu DR. Nat Chem Biol. 2015;11:316. doi: 10.1038/nchembio.1793. [DOI] [PMC free article] [PubMed] [Google Scholar]; (c) Liu KI, Ramli MN, Woo CW, Wang Y, Zhao T, Zhang X, Yim GR, Chong BY, Gowher A, Chua MZ, Jung J, Lee JH, Tan MH. Nat Chem Biol. 2016;12:980. doi: 10.1038/nchembio.2179. [DOI] [PubMed] [Google Scholar]; (d) Maji B, Moore CL, Zetsche B, Volz SE, Zhang F, Shoulders MD, Choudhary A. Nat Chem Biol. 2017;13:9. doi: 10.1038/nchembio.2224. [DOI] [PMC free article] [PubMed] [Google Scholar]; (e) Gao Y, Xiong X, Wong S, Charles EJ, Lim WA, Qi LS. Nat Methods. 2016;13:1043. doi: 10.1038/nmeth.4042. [DOI] [PMC free article] [PubMed] [Google Scholar]; (f) Hemphill J, Borchardt EK, Brown K, Asokan A, Deiters A. J Am Chem Soc. 2015;137:5642. doi: 10.1021/ja512664v. [DOI] [PMC free article] [PubMed] [Google Scholar]; (g) Zhou W, Deiters A. Angew Chem Int Ed Engl. 2016;55:5394. doi: 10.1002/anie.201511441. [DOI] [PubMed] [Google Scholar]
- 8.(a) Kearns NA, Pham H, Tabak B, Genga RM, Silverstein NJ, Garber M, Maehr R. Nat Methods. 2015;12:401. doi: 10.1038/nmeth.3325. [DOI] [PMC free article] [PubMed] [Google Scholar]; (b) Hilton IB, D’Ippolito AM, Vockley CM, Thakore PI, Crawford GE, Reddy TE, Gersbach CA. Nat Biotechnol. 2015;33:510. doi: 10.1038/nbt.3199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Fegan A, White B, Carlson JC, Wagner CR. Chem Rev. 2010;110:3315. doi: 10.1021/cr8002888. [DOI] [PubMed] [Google Scholar]
- 10.Delvecchio M, Gaucher J, Aguilar-Gurrieri C, Ortega E, Panne D. Nat Struct Mol Biol. 2013;20:1040. doi: 10.1038/nsmb.2642. [DOI] [PubMed] [Google Scholar]
- 11.Dancy BM, Cole PA. Chem Rev. 2015;115:2419. doi: 10.1021/cr500452k. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.(a) Liang FS, Ho WQ, Crabtree GR. Sci Signal. 2011;4:rs2. doi: 10.1126/scisignal.2001449. [DOI] [PMC free article] [PubMed] [Google Scholar]; (b) Wright CW, Guo ZF, Liang FS. Chembiochem. 2015;16:254. doi: 10.1002/cbic.201402576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.(a) Konermann S, Brigham MD, Trevino AE, Hsu PD, Heidenreich M, Cong L, Platt RJ, Scott DA, Church GM, Zhang F. Nature. 2013;500:472. doi: 10.1038/nature12466. [DOI] [PMC free article] [PubMed] [Google Scholar]; (b) Hathaway NA, Bell O, Hodges C, Miller EL, Neel DS, Crabtree GR. Cell. 2012;149:1447. doi: 10.1016/j.cell.2012.03.052. [DOI] [PMC free article] [PubMed] [Google Scholar]; (c) Kadoch C, Williams RT, Calarco JP, Miller EL, Weber CM, Braun SM, Pulice JL, Chory EJ, Crabtree GR. Nat Genet. 2017;49:213. doi: 10.1038/ng.3734. [DOI] [PMC free article] [PubMed] [Google Scholar]; (d) Stanton BZ, Hodges C, Calarco JP, Braun SM, Ku WL, Kadoch C, Zhao K, Crabtree GR. Nat Genet. 2017;49:282. doi: 10.1038/ng.3735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Cradick TJ, Fine EJ, Antico CJ, Bao G. Nucleic Acids Res. 2013;41:9584. doi: 10.1093/nar/gkt714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Polstein LR, Perez-Pinera P, Kocak DD, Vockley CM, Bledsoe P, Song L, Safi A, Crawford GE, Reddy TE, Gersbach CA. Genome Res. 2015;25:1158. doi: 10.1101/gr.179044.114. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.