

SUMMARY
Many studies have explored how neuromodulators affect synaptic function, yet little is known about how they modify computations at the microcircuit level. In the dorsal cochlear nucleus (DCN), a region that integrates auditory and multisensory inputs from two distinct pathways, serotonin (5-HT) enhances excitability of principal cells, predicting a generalized reduction in sensory thresholds. Surprisingly, we found that when looked at from the circuit level, 5-HT enhances signaling only from the multisensory input, while decreasing input from auditory fibers. This effect is only partially explained by an action on auditory nerve terminals. Rather, 5-HT biases processing for one input pathway by simultaneously enhancing excitability in the principal cell and in a pathway-specific feed-forward inhibitory interneuron. Thus, by acting on multiple targets, 5-HT orchestrates a fundamental shift in representation of convergent auditory and multisensory pathways, enhancing the potency of non-auditory signals in a classical auditory pathway.
In Brief
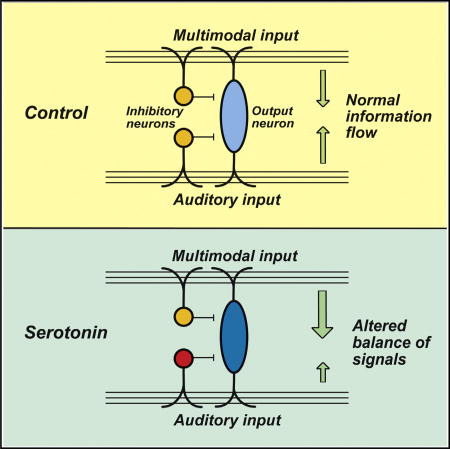
Neuromodulators may alter sensory processing upon changes in behavioral state. Tang and Trussell demonstrate that the neuromodulator serotonin shifts the representation of convergent auditory and multisensory pathways at a microcircuit level, enhancing the potency of non-auditory signals in a classical auditory brain region.
INTRODUCTION
Convergence of inputs from multiple sensory modalities, multisensory integration, is believed to enhance perceptual sensitivity and selectivity, thereby improving detection, localization, and orientation toward specific environmental cues (Stein and Stanford, 2008). During this selective process, context- and state-dependent patterns of neuromodulator release (e.g., acetylcholine, serotonin, dopamine, or norepinephrine) may reconfigure multisensory circuits, guiding behavior in adaptive ways (Froemke et al., 2013; Kayser et al., 2008; Mu et al., 2012; Parikh et al., 2007; Pinto et al., 2013). Serotonin (5-HT) in particular affects diverse physiological and behavioral functions (Lucki, 1998), and dysfunction of 5-HT signaling is implicated in a variety of psychiatric disorders (Huot et al., 2011; Meltzer et al., 1998). Despite evidence that 5-HT powerfully influences sensory processing (Brunert et al., 2016; Correia et al., 2017; Hurley and Pollak, 2005; Kapoor et al., 2016; Wood et al., 2013), it is unclear how 5-HT regulates the convergence of inputs in multisensory circuits.
The dorsal cochlear nucleus (DCN) is a well-established site of sensory integration, critical for sound source localization and orientation to sounds of interest (Imig et al., 2000; May, 2000; Oertel and Young, 2004; Sutherland et al., 1998). The layout of input pathways is clear and can be readily accessed using in vitro preparations (Figure 1A). Fusiform principal cells receive two distinct glutamatergic inputs: auditory input from auditory nerve fibers (ANFs) terminating on their basal dendrites (Brown and Ledwith, 1990; Ryugo and May, 1993; Smith and Rhode, 1985) and diverse multisensory input from parallel fibers (PFs) that terminate on their apical dendrites (Babalian, 2005; Davis et al., 1996; Itoh et al., 1987; Mugnaini et al., 1980; Shore et al., 2000; Weinberg and Rustioni, 1987; Zhou and Shore, 2004). The inhibitory microcircuits of the DCN are well characterized. ANF and PF inputs drive spiking in several subpopulations of inhibitory interneurons, notably vertical cells (also called tuberculoventral cells) (Rhode, 1999; Zhang and Oertel, 1993a) and cartwheel cells (Berrebi and Mugnaini, 1991; Golding and Oertel, 1997; Zhang and Oertel, 1993b), respectively, and these interneurons directly synapse on fusiform cells, thereby providing modality-specific feed-forward inhibition. The DCN receives a dense serotonergic input, originating predominantly from the dorsal and medial raphe nuclei (Klepper and Herbert, 1991; Steinbusch, 1981; Thompson and Thompson, 2001; Thompson et al., 1994, 1995; Willard et al., 1984). This anatomy therefore provides a foundation for understanding serotonergic modulation of multisensory integration. Importantly, aberrant 5-HT signaling is implicated in tinnitus, a condition associated with hyperactivity of the DCN (Simpson and Davies, 2000; Wu et al., 2016). Understanding the physiological basis of 5-HT modulation of the DCN may thus provide an insight into mechanisms underlying tinnitus.
Figure 1. 5-HT Causes Enhanced Multimodal Transmission Relative to Auditory Input.

(A) Simplified diagram for ANF (auditory nerve fiber) and PF (parallel fiber) input-driven microcircuits.
(B and C) Left: representative traces of (B) ANF or (C) PF stimulation (10 pulses at 50 Hz) evoked spike activity recorded in fusiform cells before and during 10 µM 5-HT application. Right: an example of spike raster plots for the activity of spike during ANF or PF stimulation presentation in the absence and presence of 5-HT (trials 11–30 highlighted by red dashed lines indicate 5-HT application). Black trace, control; red trace, during 5-HT application.
(D and E) Summary of 5-HT’s effects on the firing probability of spikes triggered by (D) ANF (n = 10) or (E) PF stimulation (n = 8). Open symbols represent the mean firing probability of individual neurons, and filled symbols represent the averaged firing probability of all neurons.
(F) 5-HT shifted the balance of convergent ANF and PF inputs toward multisensory enhancement. Change index: mean firing probability during 5-HT application relative to control. Error bars represent SEM. *p < 0.05, **p < 0.01, ***p < 0.001. CWC, cartwheel cell; FC, fusiform cell; VC, vertical cell.
A mouse brain slice preparation was developed that preserved both ANF- and PF-driven microcircuits, to determine whether and how 5-HT regulates the convergence of auditory and multisensory inputs in the DCN. We found that 5-HT differentially targets auditory and multisensory components of the microcircuit, biasing the response of fusiform cells toward multisensory transmission. This is accomplished predominantly by a facilitating action on hyperpolarization-activated cyclic nucleotide-gated (HCN) channels in both the fusiform cell and a pathway-specific interneuron, the vertical cell. In this way, the principal cell becomes consistently more responsive to multisensory input while its response to auditory signals is stymied by increased feed-forward inhibition.
RESULTS
Pathway-Specific Effects of 5-HT on Inputs to Fusiform Cells
We previously showed that 5-HT increases excitability of fusiform cells by activating HCN channels (Tang and Trussell, 2015). In the current study, we predicted that because the enhanced excitability is postsynaptic, all excitatory synaptic inputs should be more effective in driving the fusiform cell. We tested this idea in vitro by recording postsynaptic potentials and spikes in fusiform cells following 50-Hz trains of stimuli delivered to either ANFs or PFs (see Supplemental Experimental Procedures) in the absence and presence of 5-HT (Figure 1A). As expected (Tang and Trussell, 2015), 5-HT enhanced spontaneous firing of fusiform cells recorded in current-clamp (data not shown). What was unexpected was that 10 µM 5-HT decreased the firing probability of postsynaptic spikes triggered by stimulation of ANFs (40.0% ± 9.5% decrease; p < 0.01, paired t test; n = 10; Figures 1B and 1D), yet increased the firing probability of PF-triggered spikes (increased by 67.3% ± 22.1%; p < 0.05, paired t test; n = 8; Figures 1C and 1E). The result of these opposing actions was a shift in the normal balance of convergent ANF and PF inputs toward the multisensory pathway (change index: mean firing probability during 5-HT relative to control, ANF: 0.60 ± 0.10, PF: 1.67 ± 0.22; p < 0.001, unpaired t test; Figure 1F).
It seemed reasonable that the increase in PF-induced excitation of fusiform cells was due to the increased postsynaptic excitability (Tang and Trussell, 2015), and indeed 5-HT increased spike probability of fusiform cells to PF-evoked excitatory postsynaptic potentials (EPSPs) across a wide range of fiber stimulus intensities (Figure S1). However, this postsynaptic change cannot explain the decreased effectiveness of ANF input to fusiform cells. To determine whether 5-HT exerted a selective inhibitory effect specifically on auditory synapses, we recorded pairs of excitatory postsynaptic currents (EPSCs) (termed EPSC1 and EPSC2) in voltage-clamped fusiform cells by independently stimulating ANFs and PFs in the presence of inhibitory synaptic blockers (Figure 2A). Bath application of 10 µM 5-HT indeed produced a small reduction in the amplitude of ANF EPSC1 (27.0% ± 2.8% reduction; p < 0.001, paired t test; n = 19; Figures 2B and 2C) and increased the paired-pulse ratio (PPR; EPSC2/EPSC1) by 10.7% ± 2.9% from 0.82 to 0.90 (paired t test, p < 0.001; n = 19), suggesting that the reduction in ANF EPSCs could be caused by a decrease in release probability. The increase in the normalized PPR was correlated with the degree of 5-HT-induced inhibition of ANF EPSCs (r2 = 0.21; p < 0.05; Figure 2E), consistent with a presynaptic action of 5-HT. By contrast, 5-HT had no effects on PF EPSCs (EPSC1: 102.0% ± 3.4% of control; p > 0.05, n = 18; Figures 2B and 2D) or PPR (98.0% ± 1.7% of control; p > 0.05; n = 18), and no correlation was observed between the change in normalized PPR and the effect of 5-HT on PF EPSC amplitude (r2 = 0.00; p = 0.98; Figure 2E). 5-HT also reduced trains of ANF EPSCs, without affecting trains of PF EPSCs (Figures S2A–S2D), and did not affect inward currents evoked by direct microiontophoresis of glutamate on fusiform cells near their basal dendrites (Figures S2E and S2F). Previous evidence demonstrated the presence of 5-HT1A receptors (5-HT1ARs) in the DCN (Oh et al., 1999; Thompson and Wiechmann, 2002; Wright et al., 1995). Indeed, we found that the presynaptic effect of 5-HT on ANF synapses was mediated by 5-HT1ARs, because the selective agonist 8-OH-DPAT (10 µM) reversibly decreased the EPSC1 amplitude by 32.8% ± 2.1% (Figures 2F and 2G) and increased the PPR by 15.1% ± 5.3% (Figure 2G). Moreover, 5-HT did not affect ANF EPSCs in the presence of the 5-HT1AR antagonist 3 µM WAY100635 (Figure 2G). Finally, we made recordings from auditory granule cells, while stimulating their mossy fiber inputs (Balakrishnan and Trussell, 2008), and did not observe significant effects of 10 µM 5-HT on the amplitude of the mossy fiber EPSCs (n = 9; p > 0.05) or on the granule cells’ holding current (n = 14; p > 0.05; data not shown), further supporting that the PF pathway is not affected by 5-HT.
Figure 2. Suppression of Excitatory Synaptic Transmission to Fusiform Cells by 5-HT Is Input-Pathway Specific.

(A) Experimental configuration of separate stimulation of ANF and PF.
(B) Representative average traces from fusiform cells show that 10 µM 5-HT selectively reduced ANF EPSCs (top), but does not affect PF EPSCs (bottom). Black dashed lines denote the peak of EPSC1 before application of 5-HT.
(C and D) Pooled data show that 5-HT reduced the ANF EPSC1 amplitude and increased the PPR (paired-pulse ratio of EPSC2/EPSC1) (n = 19) (C), yet 5-HT did not affect PF EPSCs and short-term plasticity (n = 18) (D). Each line represents an individual neuron, and filled circles represent the averaged EPSC amplitude of all neurons.
(E) Correlation of 5-HT-induced inhibition of ANF EPSCs versus the normalized PPR (PPR5-HT/PPRcontrol: the PPR during application of 5-HT was normalized to that of the control) suggests that 5-HT inhibited ANF EPSCs via a presynaptic mechanism. The red lines are regression lines. There is no correlation between 5-HT-induced inhibition of PF EPSCs and the normalized PPR. (F) Representative averaged traces of ANF EPSCs before and during 8-OH-DPAT (10 µM).
(G) Pooled data show that 8-OH-DPAT reduced the amplitude of ANF EPSCs and increases the PPR (n = 5), and 5-HT did not change the amplitude of ANF EPSC1 and the PPR in the presence of 3 µM WAY100635 (n = 4). Open symbols represent the EPSC amplitude of individual neurons, and filled symbols represent the averaged EPSC amplitude of all neurons.
EPSCs were recorded in the presence of 1 µM strychnine and 10 µM SR95531. Stimulus artifacts were removed for clarity. Error bars represent SEM. *p < 0.05; **p < 0.01; ***p < 0.001.
Pathway-Specific Enhancement of Feed-Forward Inhibition by 5-HT
The presynaptic inhibitory effect of 5-HT on ANF EPSCs seems to oppose the postsynaptic excitatory actions of 5-HT described earlier, and thus cannot fully account for the inhibition of postsynaptic spikes driven by ANF (Figure 1). Thus, we asked whether other mechanisms might additionally reduce the impact of ANF activity following 5-HT exposure. ANF and PF inputs not only excite fusiform cells, but also initiate feed-forward inhibition by way of vertical cells and cartwheel cells, respectively (Figure 1A). To address whether 5-HT could modulate synaptic inhibition, we determined the effects of 5-HT on feed-forward inhibitory postsynaptic currents (IPSCs) in fusiform cells evoked by ANF or PF stimuli. Feed-forward inhibition was revealed by eliciting ANF- and PF-mediated synaptic responses in voltage-clamped fusiform cells held at −60 mV, a voltage between the reversal potentials for EPSCs and IPSCs (EGlu = 0 mV; ECl = −85 mV). Typically, ANF- and PF-evoked disynaptic currents exhibited a brief EPSC immediately followed by an IPSC; both phases of current were completely abolished by NBQX (10 µM) (Figure 3A), an antagonist for AMPA/kainate receptors, confirming that the inhibitory component reflects disynaptic activation of local interneurons. Further confirmation of the disynaptic nature of the IPSCs was seen through analysis of their latencies. EPSCs and IPSCs were isolated by voltage clamping at their respective reversal potentials; the latency between ANF EPSC and ANF-triggered IPSC onsets was 1.3 ± 0.2 ms (n = 5), and the latency between PF EPSC and PF-triggered IPSC onsets was 2.0 ± 0.1 ms (n = 7), consistent with previous studies (Doiron et al., 2011; Sedlacek and Brenowitz, 2014). ANF-triggered IPSCs showed depression, whereas PF-activated IPSCs showed facilitation (Figures 3A and 3B), reflecting the short-term plasticity of the excitatory ANF and PF inputs onto vertical cells and cartwheel cells, respectively. In particular, ANF to vertical cell excitatory inputs depress, whereas vertical cell to fusiform cell inhibitory inputs do not (Kuo et al., 2012). Together, these data indicate that ANF and PF inputs can drive distinct monosynaptic EPSCs and disynaptic IPSCs onto fusiform cells. This disynaptic inhibition strongly constrained the temporal summation of EPSCs during repetitive afferent activity (Figure 3B).
Figure 3. Enhancement of Feed-Forward Inhibition to Fusiform Cells by 5-HT Is Input-Pathway Specific.

(A) EPSC-IPSC sequence was recorded in fusiform cells in response to ANF or PF stimulation at holding potentials of −60 mV. The disynaptic currents were abolished by 10 µM NBQX (control, black; NBQX, blue), confirming the disynaptic origin of the IPSCs.
(B) Representative average traces of disynaptic currents in responses to ANF or PF stimulation (10 pulses at 50 Hz) from fusiform cells show that 10 µM 5-HT reduced the inward component and enhanced the outward component of ANF disynaptic currents (top) but did not change PF disynaptic currents (bottom).
(C) Superimposed traces of first currents in (B).
(D and E) Summary of inward charge, outward charge, and charge ratio of (D) ANF- and (E) PF-disynaptic currents under the control and 5-HT conditions. The charge ratio is the ratio of excitatory-to-inhibitory charge for each cell, which is then averaged across all cells. Stimulus artifacts were removed for clarity. Error bars represent SEM.
To assess the effects of 5-HT on stimulus-evoked disynaptic IPSCs, we examined 5-HT modulation by measuring the total outward charge in the EPSC/IPSC sequence with and without 5-HT. 5-HT enhanced the disynaptic inhibitory current in response to ANF stimulation, yet had no effects on the disynaptic inhibitory current evoked by PF stimulation in fusiform cells (Figures 3B–3D; first stimulus, control: 213.3 ± 39.9 pA*ms, 5-HT: 397.5 ± 71.1 pA*ms, p < 0.01, n = 6; average of 6–10 stimuli, control: 94.0 ± 6.7 pA*ms, 5-HT: 168.0 ± 25.5 pA*ms, p < 0.05). Moreover, 5-HT shifted the balance of excitation and inhibition (E/I) over the entire course of the stimulus train. 5-HT decreased the average E/I ratio for the first stimulus in a train (control: 0.68 ± 0.17, 5-HT: 0.22 ± 0.04; p < 0.05; n = 6; Figure 3D), and this reduction was maintained throughout the train (average for 6–10 stimuli, control: 1.11 ± 0.15, 5-HT: 0.63 ± 0.15; p < 0.001) despite ongoing synaptic depression. The decrease in inward synaptic current likely results from a reduction of excitatory synaptic transmission by activation of presynaptic 5-HT1ARs (Figure 2). Such a decrease in inward charge must contribute to the enhanced outward charge phase of these waveforms; however, as shown below, a second factor enhancing the outward charge component is enhanced feed-forward inhibition onto fusiform cells. Importantly, 5-HT had no effect on the disynaptic current evoked by PF stimulation (p > 0.05; n = 7; Figures 3B, 3C, and 3E). Thus, serotonergic modulation of inhibition onto fusiform cells is pathway specific.
5-HT Enhances Excitability of Vertical Cells
To understand how 5-HT selectively enhanced inhibition during ANF stimulation, we examined the effects of 5-HT on each element of the inhibitory microcircuit. Enhanced feed-forward inhibition could reflect increased excitability of vertical cells by activation of somatodendritic 5-HT receptors, which would in turn enable recruitment of greater numbers of vertical cells during auditory nerve activity. Alternatively, there might be a direct potentiation of ANF-vertical cell excitatory synapses and/or vertical cell-fusiform cell inhibitory synapses by presynaptic 5-HT receptors. We therefore tested the effects of 5-HT on the excitability of vertical cells, EPSCs in vertical cells evoked by ANF stimulation, and vertical-cell-mediated IPSCs onto fusiform cells.
Vertical cells were identified on the basis of their location and firing pattern (Figure 4A; Kuo et al., 2012). Under current-clamp, 5-HT (10 µM) depolarized the resting membrane potential by 4.3 ± 0.8 mV (p < 0.001, paired t test; n = 9; Figures 4B and 4C) in the presence of synaptic blockers, indicating a direct postsynaptic action of 5-HT. Accompanying the depolarization was an increase of spontaneous spike activity (control: 0.48 ± 0.45 Hz, 5-HT: 13.5 ± 2.7 Hz; p < 0.001; n = 12; Figures 4B–4D). To examine how 5-HT affects the sensitivity of vertical cells to current stimuli, we injected a series of 0.5 s current steps into vertical cells, starting with −20 pA and incrementing by 20 pA, and the spike rate at each current level was measured with and without 5-HT. 5-HT shifted the input-output relation to the left (Figures 4E and 4F), yet did not change the resting membrane potential or spike rate in cartwheel cells (p > 0.05; n = 9; Figures 4B–4D), the primary feed-forward inhibitory neuron of the multisensory pathway.
Figure 4. 5-HT Increases the Excitability of Vertical Cells but Not Cartwheel Cells.

(A) Representative action potential firing patterns of a vertical cell and a cartwheel cell recorded in current-clamp mode.
(B) 5-HT depolarized the membrane potential and increased spontaneous spike rate in a vertical cell (left), but did not change the membrane potential in a cartwheel cell (right). Black dashed lines denote resting membrane potential before application of 5-HT.
(C and D) 5-HT increased (C) resting membrane potential (n = 9) and (D) spontaneous spike rate of vertical cells (n = 12), but not of cartwheel cells (n = 9).
(E) Representative traces of action potential firing in a vertical cell elicited by somatic injection of current pulses illustrating the effects of 5-HT.
(F) Firing frequency as a function of amplitude of injected current (n = 8) in vertical cells.
Error bars represent SEM. **p < 0.01; ***p < 0.001.
To explore the basis of the postsynaptic effect of 5-HT on vertical cells, we recorded in voltage-clamp configuration at −70 mV, observing a 5-HT-induced slow inward current (Figure S3A). No effect of 5-HT was seen in recordings from cartwheel cells (data not shown). The inward current in vertical cells could be largely abolished by the selective 5-HT2ARs antagonist MDL-11939 (2 µM), and was mimicked by application of 30 µM α-methyl-5-HT, a selective agonist of 5-HT2ARs (Figures S3A and S3B), consistent with the presence of 5-HT2ARs in the DCN (Cornea-Hébert et al., 1999; Pazos et al., 1985; Wright et al., 1995), and labeling of presumptive DCN vertical cells in Htr2a-Cre-tdTomato mice (http://www.brain-map.org). Furthermore, 5-HT-induced current was abolished by intracellular GTP-γ-S (1.5 mM), a non-hydrolysable GTP analog that blocks G protein in its activated state (Figures S3A and S3B), confirming that the 5-HTevoked response requires G protein signaling. Finally, we found that this current could be largely blocked by HCN channel blockers, Cs+ (2 mM) or ZD7288 (20 µM) (Figures S3A and S3B), indicating that HCN channels are the primary downstream targets for these observed excitatory actions of 5-HT. These data are almost identical to results for fusiform cells (Tang and Trussell, 2015) and suggest that fusiform and vertical cells may use the same pathways in their response to 5-HT.
5-HT Does Not Affect Transmitter Release from Vertical Cells
We next tested for effects of 5-HT on other key synapses in the circuit. 5-HT did not change the peak amplitude or the PPR of ANF-to-vertical cell EPSCs (Figures 5A–5C) or PF-to-cartwheel cell EPSCs (Figures 5B and 5C). Although 5-HT did not increase EPSCs in vertical cells, it markedly increased the firing probability of spikes evoked by a train of suprathreshold “strong” stimulation of ANF input (firing probability in control: 0.44 ± 0.04; firing probability in 5-HT: 0.81 ± 0.08; p < 0.001; n = 12; Figures 5D and 5G), indicating that 5-HT increased coupling between EPSPs and action potential generation. Interestingly, 5-HT had no significant effect on the latency to spiking in vertical cells evoked by a train of strong intensity stimulation of ANF (control 2.77 ± 0.33 ms versus 5-HT 2.51 ± 0.29 ms; n = 10; p > 0.05, paired t test) despite the larger spike probability. 5-HT also enhanced the firing probability in response to subthreshold EPSPs evoked by “weak” ANF stimulation (firing probability in control: 0.00 ± 0.00; firing probability in 5-HT: 0.17 ± 0.03; p < 0.01; n = 5 out of 6 cells; Figures 5E–5G). These data illustrate that the way in which 5-HT increases inhibition is specifically by recruitment, allowing vertical cells to fire more consistently during ANF activity, and by extension to allow more vertical cells contacting a given fusiform cell to become active.
Figure 5. Pathway-Specific Enhancement of Orthodromic Firing by 5-HT.

(A) Experimental configuration.
(B) Representative traces from a vertical cell (top) show that 10 µM 5-HT did not affect EPSCs evoked by ANF stimulation. Similarly, 5-HT did not change EPSCs recorded in a cartwheel cell (bottom) evoked by PF stimulation.
(C) Summary of 5-HT’s effects on EPSC1 and PPR in vertical cells (n = 23) and cartwheel cells (n = 11). (D) Top: representative traces of EPSPs and action potentials in a vertical cell in response to ANF stimulation (10 pulses at 50 Hz) with a strong intensity before and during 5-HT. Bottom: the raster plots for the activity of spike during ANF stimulation presentation with and without 5-HT.
(E) Representative traces of EPSPs and action potentials in response to ANF stimulation with a weak intensity in a vertical cell before and during 5-HT.
(F) The first EPSPs or spikes in (E) are shown at enlarged scales.
(G) Summary of 5-HT’s effects on firing probability of spikes triggered by ANF stimulation with strong (n = 12) or weak (n = 5) intensity.
Error bars represent SEM. **p < 0.01; ***p < 0.001.
To test the possibility that 5-HT could directly affect inhibitory transmission between vertical and fusiform cells, we made dual recordings from synaptically connected vertical cell-fusiform cell pairs (Figure S4). Unitary IPSCs in fusiform cells were triggered by trains of spikes at 50 Hz evoked via somatic current injection into presynaptic vertical cells. Although 5-HT depolarized the vertical cells, it did not change the amplitude (p > 0.05; n = 5 pairs; Figures S4B and S4D) or short-term plasticity (p > 0.05) of unitary IPSCs in fusiform cells. Similarly, 5-HT also did not affect the unitary IPSCs in fusiform cells evoked by spikes in connected cartwheel cells (p > 0.05; n = 6 pairs; Figures S4C and S4E). These data suggest that 5-HT regulates feed-forward inhibition to fusiform cells through enhanced excitability of vertical cells, rather than through a direct effect on ANF-vertical cell excitatory synapses or vertical cell-fusiform cell inhibitory synapses.
To further confirm that it is excitation of vertical cells by 5-HT that accounts for the enhanced feed-forward inhibition in fusiform cells, we made paired recordings from synaptically coupled vertical and fusiform cells, and monitored spike and IPSC activity, respectively, as 5-HT was washed into the bath (Figures 6A and 6B). 5-HT triggered copious firing in the vertical cell and increased the frequency of spontaneous IPSCs in the postsynaptic fusiform cell (Figures 6C–6G). 5-HT increased both the frequency and the amplitude of IPSCs (Figures 6C–6E). Because 5-HT has no direct effect on the inhibitory synapses, the increase in amplitude presumably arose from an increase in spike-evoked, non-uniquantal spontaneous events. In order to determine whether the enhanced IPSCs were due to the vertical cell, we made presynaptic spike-triggered averages, revealing that IPSCs often occurred immediately after a spontaneous (5-HT-induced) presynaptic spike. The latency from the spike to the onset of the IPSC was consistent with monosynaptic transmission from the recorded vertical cell (Figures 6F and 6G; mean latency: 0.48 ± 0.06 ms; n = 5).
Figure 6. Enhanced Excitability of Vertical Cells by 5-HT Leads to Enhanced Inhibitory Tone.

(A) Experimental configuration of a vertical cell-fusiform cell paired recording.
(B) A sample synaptically coupled vertical cell-fusiform cell paired recording showing unitary IPSCs (gray: individual races; black: average trace) recorded in voltage-clamp in a postsynaptic fusiform cell (bottom) in response to train stimulation (5 pulses at 50 Hz) recorded in current-clamp in a presynaptic vertical cell (top).
(C and D) Presynaptic membrane potentials or action potentials were recorded in current-clamp from a vertical cell (C), and IPSCs were recorded in voltage-clamp from a fusiform cell (D) in the absence and presence of 5-HT. Bottom: representative averaged traces of IPSCs before and during 5-HT.
(E) Cumulative fractions of the frequency and amplitude of IPSCs show that 5-HT increased both their frequency and amplitude.
(F) Spikes (top) in the presynaptic vertical cell evoked by 5-HT-generated IPSCs (bottom) (*) in the postsynaptic fusiform cell.
(G) Vertical cell spike-triggered average shows that the vertical cell spike led to short-latency IPSCs (indicated by arrows) in fusiform cell, further confirming that 5-HT evoked spikes in vertical cells generated IPSCs in fusiform cell.
IEI, inter-event interval.
5-HT Reduces Post-excitation Pauses Identically for ANF- and PF-Driven Spike Activity
A well-known feature of fusiform cells recorded in vivo or in vitro is a prolonged HCN-dependent afterhyperpolarization that follows afferent activity, sufficient to eliminate the spontaneous spike activity that is typical of these neurons (Apostolides and Trussell, 2014; Hancock and Voigt, 2002; Rhode et al., 1983). We found that this pause in spiking was also modulated by 5-HT; however, unlike its differential effects on afferent activity, the pause was modulated identically for both sensory pathways, consistent with a postsynaptic site of action of 5-HT. The duration of this pause was defined as the latency from last synaptic stimulation to the first spontaneous spike. 5-HT reduced the pause in response to either ANF or PF stimuli (latency in control: 572 ± 49 ms; latency in 5-HT: 300 ± 44 ms; p < 0.001; n = 11; Figures 7A, 7B, 7D, and 7E). We reasoned that this pause could result from the deactivation of HCN channels during spike trains, leading to a rebound hyperpolarization after spiking ceases. As 5-HT depolarizes the neurons, this deactivation is already partially in place prior to evoked spiking; given that the spontaneous firing rate is enhanced by the depolarization, both effects should reduce the pause duration. We tested this idea by mimicking 5-HT’s depolarizing effect by somatic current injection. As expected, somatic current injections (+50 to +100 pA) fully mimicked the effect of 5-HT (Figures 7C and 7E). Altogether, our data suggest an input pathway/circuit selectivity neuromodulation that biases the sensitivity of the principal cell toward multisensory signals, while simultaneously reducing the quiescent state following signals from any source.
Figure 7. 5-HT Decreases Post-excitation Pauses in Response to Either Auditory Nerve Fiber- or Parallel Fiber-Driven Spikes in Fusiform Cells.

(A and B) Top: representative superimposed traces of spontaneous and (A) ANF or (B) PF stimulation-evoked spiking activity recorded in two fusiform cells before and during application of 5-HT. Both ANF and PF stimulation-evoked spikes were followed by a pause in spontaneous spike activity. 5-HT reduced this transient inhibition. Bottom: the raster plots for the activity of spike in these two fusiform cells during (A) ANF or (B) PF stimulation presentation in the absence and presence of 5-HT (trials 11–30 highlighted by red dashed lines indicate 5-HT application).
(C) Top: representative superimposed traces of spontaneous and ANF-evoked spiking activity recorded in a fusiform cell with and without a +50 pA current injection (control: black traces; current injection: green traces). Current injection reduced this pause. Bottom: an example of raster plots for the activity of spike in the fusiform cell (A) during ANF stimulation presentation under the control and current injection conditions (current injection indicated by two dashed green lines from trials 21–30).
(D) An example of poststimulus time histogram (PSTH; 10 trials) for the activity of the spikes during ANF stimulation presentation under the control and 5-HT conditions from (A) illustrates how the pause was quantified as the latency from the last stimulus artifact to first spontaneous spike.
(E) 5-HT significantly reduced the latency (n = 11). Similarly, a somatic current injection (I injection: +50 to +100 pA) reduced the latency (n = 3).
Thick blue lines indicate the periods with a train of ANF or PF stimulation at 10 pulses and 50 Hz. Error bars represent SEM. *p < 0.05, ***p < 0.001.
DISCUSSION
We identified a neuromodulatory circuit mechanism that suppressed signaling through the auditory pathway in DCN, while simultaneously enhancing transmission through a multisensory pathway. Given that the primary target of the DCN is the inferior colliculus, and given the striking convergence of multiple modalities into the DCN, such pathway-selective neuromodulation could enhance the salience of particular sensory events associated in time with a given acoustic signal, such as somatosensory activity from pinna, head/neck movement, or trigeminal activity (Kanold and Young, 2001; Kanold et al., 2011; Shore, 2005).
Serotonergic Regulation of Inhibitory Microcircuit Is Cell-Type Specific
Inhibitory interneurons are critical in adjusting relative sensitivity to the diverse inputs a neural circuit receives (Huang et al., 2007; McBain and Fisahn, 2001; Roberts et al., 2013). Accordingly, differential control of interneurons by neuromodulators is a potent mechanism for circuit-level plasticity (Lawrence, 2008). Although vertical cells and cartwheel cells are the most common interneuron in the DCN, their response to 5-HT is sharply different. Indeed, the entire shift in response toward the multimodal input can be accounted for by the actions of the 5-HT2AR→HCN channel signaling pathway in the principal cells and a single type of interneuron, the vertical cell, with an additional small contribution from 5-HT1AR in presynaptic inhibition.
Clearly, there are multiple possible ways in which this circuit could be controlled to effect a shift from auditory to multimodal signaling, for example, by purely presynaptic modulation at the input level. Although 5-HT increased excitability and spontaneous firing of vertical cells, it did not affect strength of EPSCs or unitary IPSCs in the ANF→vertical cell→fusiform cell inhibitory sequence. 5-HT also increased the likelihood of coupling between ANF EPSPs and action potential initiation in vertical cell, again by a postsynaptic mechanism. Finally, dual recordings provided direct evidence that 5-HT increased vertical cell-mediated inhibitory activity in fusiform cells. Given that multiple vertical cells converge upon each fusiform cell (Kuo et al., 2012; Voigt and Young, 1990), and the vertical cell excitability is enhanced by 5-HT, the enhanced inhibition of the fusiform cell’s ANF responses reflects the enhanced recruitment of inhibitory neurons. Intriguingly, this simple mechanism assures that the level of inhibition would scale with the degree of auditory activity. In contrast to 5-HT’s modulation of ANF→fusiform cell feed-forward inhibitory microcircuit, the noradrenergic system selectively regulates the excitability of cartwheel cells and enhances the signal-to-noise ratio of inhibitory signals through the multisensory pathway (Kuo and Trussell, 2011), whereas dopamine acts to adjust spike modes determined by cartwheel cell axons (Bender et al., 2010). Therefore, different neuromodulatory systems can control DCN circuits in distinct ways, yet all appear to have selectivity in their target inhibitory neurons, and thereby may guide distinctly different alterations in processing.
Functional Implications of Serotonergic Modulation
What are the functional outcomes of enhanced multisensory processing in DCN? A similar multisensory enhancement has been reported in superior colliculus and neocortex (Alvarado et al., 2009; Olcese et al., 2013), and proposed as a key mechanism to enhance the salience of context events. Therefore, modulation of integration of sensory signals at the DCN circuit provides flexibility in bi-modal integration (Chabrol et al., 2015; Shore, 2005), allowing multisensory cues potentially to improve detection and discrimination of events in the context environment. In support of this notion, behavioral studies suggest that integration of multisensory input improves the accuracy of orientation of sounds (Kanold and Young, 2001; Young et al., 1995). Therefore, this serotonergic neural circuit could be a key component of the DCN circuitry highlighting the response to salient environmental events.
Studies of 5-HT actions in DCN in vivo show conflicting results (Ebert and Ostwald, 1992; Felix et al., 2017), possibly as a result of variations in where 5-HT is iontophoretically released relative to the location of recorded neurons. In this study, 5-HT was applied uniformly throughout the tissue. If it acts locally, rather than diffusely, the inhibition ANF signals could have a more significant outcome. In vivo studies suggested that vertical-cell-mediated inhibition may regulate frequency and/or intensity tuning (Nelken and Young, 1994; Spirou et al., 1999; Young and Davis, 2002; Zhou et al., 2012). If 5-HT was released locally so as to act on subsets of vertical cells, auditory receptive fields might be sharpened relative to the basal discharge of the cell or may be shifted in their borders. Thus, 5-HT-induced suppression of auditory stimuli might refine sensory receptive fields and minimize potentially distracting effects of competing stimuli, thus providing an efficient strategy to improve sound localization in noisy environments. Future study should explore how 5-HT release regulates the DCN circuitry and consequently modulates its function in living animals.
Beyond the physiological function of 5-HT, our findings may provide insight into the biological mechanisms of tinnitus. Aberrant serotonergic neurotransmission is implicated in tinnitus (Caperton and Thompson, 2011; Norena et al., 1999; Salvinelli et al., 2003; Simpson and Davies, 2000), and enhanced activity (hyperactivity) in fusiform cells of the DCN is also associated with tinnitus (Brozoski et al., 2002; Dehmel et al., 2012; Li et al., 2013). Thus, it will be important to determine whether dysfunctional modulation of the DCN could play a role in central tinnitus. The tonic excitation of fusiform cells is one tinnitus-like outcome. However, studies indicate that the tinnitus can be enhanced in many patients by movement of the head or jaw (Levine and Oron, 2015); such an effect may be related to increased effectiveness of somatosensory signaling in the DCN by 5-HT. Thus, the current study may provide a basis for future work to understand whether dysregulation of 5-HT signaling could affect the experience of tinnitus.
EXPERIMENTAL PROCEDURES
Animals
All procedures were approved by the IACUC of Oregon Health and Science University. Animals used in this study were C57BL/6J wild-type mice or transgenic mice. GlyT2-EGFP mice (labeling glycinergic neurons) were backcrossed into C57BL/6J wild-type mice and maintained and genotyped as previously described (Roberts et al., 2008). NF107-Cre-tdTomato mice (a gift of Chip Gerfen) were generated by crossing NF107-Cre mice with an Ai9 reporter line.
Electrophysiology
Refer to Supplemental Experimental Procedures for detailed methods. For most experiments, fusiform cell, cartwheel cell, and vertical cell were identified based on their location, morphology, and electrophysiological properties. Additionally, GlyT2-EGFP (all glycinergic neurons labeled) transgenic mice in which EGFP is expressed in glycinergic neurons were used to identify vertical and cartwheel cells. Whole-cell recordings were performed in acute conventional or thick brain slices containing the DCN using patch pipettes filled with a K+-based intracellular solution.
Paired Recordings
For paired pre- and postsynaptic recordings (vertical cell-fusiform cell or cartwheel cell-fusiform cell), fusiform cells were voltage-clamped with an intracellular solution containing 20 µM Alexa Fluor 488 so that morphology could be confirmed. Then, nearby (<50 µm intersomatic distance) vertical cells in the deep layer or cartwheel cells in the molecular layer of the fusiform cell were patched in current-clamp mode. After identifying synaptically coupled vertical cell-fusiform cell or cartwheel cell-fusiform cell pairs and demonstrating reliable induction of unitary IPSCs in the postsynaptic fusiform cell upon depolarization of the presynaptic vertical cell or cartwheel cell, the effects of 5-HT on unitary IPSCs were then tested.
Statistical Analysis
All traces were acquired with pClamp 10 software (Molecular Devices) and Axograph X. All figures were made with IgorPro (WaveMetrics). All data are presented as mean ± SEM unless specified otherwise, and statistical significance was assessed using Student’s t tests as appropriate: *p < 0.05; **p < 0.01; ***p < 0.001. Further details are in the Supplemental Experimental Procedures.
Supplementary Material
Highlights.
In a multisensory integrator, 5-HT readjusts the relative strength of different inputs
5-HT enhances excitability of principal cells and auditory input-driven interneurons
Principal cells then respond well to multisensory input, yet have weaker auditory drive
5-HT therefore operates at the microcircuit level to modulate sensory processing
Acknowledgments
We thank Dr. Miloslav Sedlacek for advice in preparing thick slices, Lucille Moore for help with NF107-Cre-tdTomato mouse breeding, all members of the Trussell lab for helpful discussions, and Dr. John Williams for critical comments on the manuscript. This work was supported by NIH grants NS028901 and DC004450 (to L.O.T.) and an Emerging Research grant from the Hearing Health Foundation (to Z.-Q.T.).
Footnotes
Supplemental Information includes Supplemental Experimental Procedures and four figures and can be found with this article online at http://dx.doi.org/10.1016/j.celrep.2017.07.079.
AUTHOR CONTRIBUTIONS
Z.-Q.T. conducted experiments and analyzed data. Z.-Q.T. and L.O.T. designed experiments and wrote the manuscript.
References
- Alvarado JC, Stanford TR, Rowland BA, Vaughan JW, Stein BE. Multisensory integration in the superior colliculus requires synergy among corticocollicular inputs. J. Neurosci. 2009;29:6580–6592. doi: 10.1523/JNEUROSCI.0525-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Apostolides PF, Trussell LO. Control of interneuron firing by subthreshold synaptic potentials in principal cells of the dorsal cochlear nucleus. Neuron. 2014;83:324–330. doi: 10.1016/j.neuron.2014.06.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Babalian AL. Synaptic influences of pontine nuclei on cochlear nucleus cells. Exp. Brain Res. 2005;167:451–457. doi: 10.1007/s00221-005-0178-8. [DOI] [PubMed] [Google Scholar]
- Balakrishnan V, Trussell LO. Synaptic inputs to granule cells of the dorsal cochlear nucleus. J. Neurophysiol. 2008;99:208–219. doi: 10.1152/jn.00971.2007. [DOI] [PubMed] [Google Scholar]
- Bender KJ, Ford CP, Trussell LO. Dopaminergic modulation of axon initial segment calcium channels regulates action potential initiation. Neuron. 2010;68:500–511. doi: 10.1016/j.neuron.2010.09.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berrebi AS, Mugnaini E. Distribution and targets of the cartwheel cell axon in the dorsal cochlear nucleus of the guinea pig. Anat. Embryol. (Berl.) 1991;183:427–454. doi: 10.1007/BF00186433. [DOI] [PubMed] [Google Scholar]
- Brown MC, Ledwith JV., 3rd Projections of thin (type-II) and thick (type-I) auditory-nerve fibers into the cochlear nucleus of the mouse. Hear. Res. 1990;49:105–118. doi: 10.1016/0378-5955(90)90098-a. [DOI] [PubMed] [Google Scholar]
- Brozoski TJ, Bauer CA, Caspary DM. Elevated fusiform cell activity in the dorsal cochlear nucleus of chinchillas with psychophysical evidence of tinnitus. J. Neurosci. 2002;22:2383–2390. doi: 10.1523/JNEUROSCI.22-06-02383.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brunert D, Tsuno Y, Rothermel M, Shipley MT, Wachowiak M. Cell-type-specific modulation of sensory responses in olfactory bulb circuits by serotonergic projections from the raphe nuclei. J. Neurosci. 2016;36:6820–6835. doi: 10.1523/JNEUROSCI.3667-15.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caperton KK, Thompson AM. Activation of serotonergic neurons during salicylate-induced tinnitus. Otol. Neurotol. 2011;32:301–307. doi: 10.1097/MAO.0b013e3182009d46. [DOI] [PubMed] [Google Scholar]
- Chabrol FP, Arenz A, Wiechert MT, Margrie TW, DiGregorio DA. Synaptic diversity enables temporal coding of coincident multisensory inputs in single neurons. Nat. Neurosci. 2015;18:718–727. doi: 10.1038/nn.3974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cornea-Hébert V, Riad M, Wu C, Singh SK, Descarries L. Cellular and subcellular distribution of the serotonin 5-HT2A receptor in the central nervous system of adult rat. J. Comp. Neurol. 1999;409:187–209. doi: 10.1002/(sici)1096-9861(19990628)409:2<187::aid-cne2>3.0.co;2-p. [DOI] [PubMed] [Google Scholar]
- Correia PA, Lottem E, Banerjee D, Machado AS, Carey MR, Mainen ZF. Transient inhibition and long-term facilitation of locomotion by phasic optogenetic activation of serotonin neurons. eLife. 2017;6:e20975. doi: 10.7554/eLife.20975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davis KA, Miller RL, Young ED. Effects of somatosensory and parallel-fiber stimulation on neurons in dorsal cochlear nucleus. J. Neurophysiol. 1996;76:3012–3024. doi: 10.1152/jn.1996.76.5.3012. [DOI] [PubMed] [Google Scholar]
- Dehmel S, Pradhan S, Koehler S, Bledsoe S, Shore S. Noise overexposure alters long-term somatosensory-auditory processing in the dorsal cochlear nucleus—possible basis for tinnitus-related hyperactivity? J. Neurosci. 2012;32:1660–1671. doi: 10.1523/JNEUROSCI.4608-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doiron B, Zhao Y, Tzounopoulos T. Combined LTP and LTD of modulatory inputs controls neuronal processing of primary sensory inputs. J. Neurosci. 2011;31:10579–10592. doi: 10.1523/JNEUROSCI.1592-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ebert U, Ostwald J. Serotonin modulates auditory information processing in the cochlear nucleus of the rat. Neurosci. Lett. 1992;145:51–54. doi: 10.1016/0304-3940(92)90201-h. [DOI] [PubMed] [Google Scholar]
- Felix RA, 2nd, Elde CJ, Nevue AA, Portfors CV. Serotonin modulates response properties of neurons in the dorsal cochlear nucleus of the mouse. Hear. Res. 2017;344:13–23. doi: 10.1016/j.heares.2016.10.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Froemke RC, Carcea I, Barker AJ, Yuan K, Seybold BA, Martins ARO, Zaika N, Bernstein H, Wachs M, Levis PA, et al. Long-term modification of cortical synapses improves sensory perception. Nat. Neurosci. 2013;16:79–88. doi: 10.1038/nn.3274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Golding NL, Oertel D. Physiological identification of the targets of cartwheel cells in the dorsal cochlear nucleus. J. Neurophysiol. 1997;78:248–260. doi: 10.1152/jn.1997.78.1.248. [DOI] [PubMed] [Google Scholar]
- Hancock KE, Voigt HF. Intracellularly labeled fusiform cells in dorsal cochlear nucleus of the gerbil. I. Physiological response properties. J. Neurophysiol. 2002;87:2505–2519. doi: 10.1152/jn.2002.87.5.2505. [DOI] [PubMed] [Google Scholar]
- Huang ZJ, Di Cristo G, Ango F. Development of GABA innervation in the cerebral and cerebellar cortices. Nat. Rev. Neurosci. 2007;8:673–686. doi: 10.1038/nrn2188. [DOI] [PubMed] [Google Scholar]
- Huot P, Fox SH, Brotchie JM. The serotonergic system in Parkinson’s disease. Prog. Neurobiol. 2011;95:163–212. doi: 10.1016/j.pneurobio.2011.08.004. [DOI] [PubMed] [Google Scholar]
- Hurley LM, Pollak GD. Serotonin modulates responses to species-specific vocalizations in the inferior colliculus. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 2005;191:535–546. doi: 10.1007/s00359-005-0623-y. [DOI] [PubMed] [Google Scholar]
- Imig TJ, Bibikov NG, Poirier P, Samson FK. Directionality derived from pinna-cue spectral notches in cat dorsal cochlear nucleus. J. Neurophysiol. 2000;83:907–925. doi: 10.1152/jn.2000.83.2.907. [DOI] [PubMed] [Google Scholar]
- Itoh K, Kamiya H, Mitani A, Yasui Y, Takada M, Mizuno N. Direct projections from the dorsal column nuclei and the spinal trigeminal nuclei to the cochlear nuclei in the cat. Brain Res. 1987;400:145–150. doi: 10.1016/0006-8993(87)90662-7. [DOI] [PubMed] [Google Scholar]
- Kanold PO, Young ED. Proprioceptive information from the pinna provides somatosensory input to cat dorsal cochlear nucleus. J. Neurosci. 2001;21:7848–7858. doi: 10.1523/JNEUROSCI.21-19-07848.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kanold PO, Davis KA, Young ED. Somatosensory context alters auditory responses in the cochlear nucleus. J. Neurophysiol. 2011;105:1063–1070. doi: 10.1152/jn.00807.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kapoor V, Provost AC, Agarwal P, Murthy VN. Activation of raphe nuclei triggers rapid and distinct effects on parallel olfactory bulb output channels. Nat. Neurosci. 2016;19:271–282. doi: 10.1038/nn.4219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kayser C, Petkov CI, Logothetis NK. Visual modulation of neurons in auditory cortex. Cereb. Cortex. 2008;18:1560–1574. doi: 10.1093/cercor/bhm187. [DOI] [PubMed] [Google Scholar]
- Klepper A, Herbert H. Distribution and origin of noradrenergic and serotonergic fibers in the cochlear nucleus and inferior colliculus of the rat. Brain Res. 1991;557:190–201. doi: 10.1016/0006-8993(91)90134-h. [DOI] [PubMed] [Google Scholar]
- Kuo SP, Trussell LO. Spontaneous spiking and synaptic depression underlie noradrenergic control of feed-forward inhibition. Neuron. 2011;71:306–318. doi: 10.1016/j.neuron.2011.05.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuo SP, Lu H-W, Trussell LO. Intrinsic and synaptic properties of vertical cells of the mouse dorsal cochlear nucleus. J. Neurophysiol. 2012;108:1186–1198. doi: 10.1152/jn.00778.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lawrence JJ. Cholinergic control of GABA release: emerging parallels between neocortex and hippocampus. Trends Neurosci. 2008;31:317–327. doi: 10.1016/j.tins.2008.03.008. [DOI] [PubMed] [Google Scholar]
- Levine RA, Oron Y. Tinnitus. Handb. Clin. Neurol. 2015;129:409–431. doi: 10.1016/B978-0-444-62630-1.00023-8. [DOI] [PubMed] [Google Scholar]
- Li S, Choi V, Tzounopoulos T. Pathogenic plasticity of Kv7.2/3 channel activity is essential for the induction of tinnitus. Proc. Natl. Acad. Sci. USA. 2013;110:9980–9985. doi: 10.1073/pnas.1302770110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lucki I. The spectrum of behaviors influenced by serotonin. Biol. Psychiatry. 1998;44:151–162. doi: 10.1016/s0006-3223(98)00139-5. [DOI] [PubMed] [Google Scholar]
- May BJ. Role of the dorsal cochlear nucleus in the sound localization behavior of cats. Hear. Res. 2000;148:74–87. doi: 10.1016/s0378-5955(00)00142-8. [DOI] [PubMed] [Google Scholar]
- McBain CJ, Fisahn A. Interneurons unbound. Nat. Rev. Neurosci. 2001;2:11–23. doi: 10.1038/35049047. [DOI] [PubMed] [Google Scholar]
- Meltzer CC, Smith G, DeKosky ST, Pollock BG, Mathis CA, Moore RY, Kupfer DJ, Reynolds CF., 3rd Serotonin in aging, late-life depression, and Alzheimer’s disease: the emerging role of functional imaging. Neuropsychopharmacology. 1998;18:407–430. doi: 10.1016/S0893-133X(97)00194-2. [DOI] [PubMed] [Google Scholar]
- Mu Y, Li XQ, Zhang B, Du JL. Visual input modulates audiomotor function via hypothalamic dopaminergic neurons through a cooperative mechanism. Neuron. 2012;75:688–699. doi: 10.1016/j.neuron.2012.05.035. [DOI] [PubMed] [Google Scholar]
- Mugnaini E, Warr WB, Osen KK. Distribution and light microscopic features of granule cells in the cochlear nuclei of cat, rat, and mouse. J. Comp. Neurol. 1980;191:581–606. doi: 10.1002/cne.901910406. [DOI] [PubMed] [Google Scholar]
- Nelken I, Young ED. Two separate inhibitory mechanisms shape the responses of dorsal cochlear nucleus type IV units to narrowband and wideband stimuli. J. Neurophysiol. 1994;71:2446–2462. doi: 10.1152/jn.1994.71.6.2446. [DOI] [PubMed] [Google Scholar]
- Norena A, Cransac H, Chéry-Croze S. Towards an objectification by classification of tinnitus. Clin. Neurophysiol. 1999;110:666–675. doi: 10.1016/s1388-2457(98)00034-0. [DOI] [PubMed] [Google Scholar]
- Oertel D, Young ED. What’s a cerebellar circuit doing in the auditory system? Trends Neurosci. 2004;27:104–110. doi: 10.1016/j.tins.2003.12.001. [DOI] [PubMed] [Google Scholar]
- Oh CK, Drescher MJ, Hatfield JS, Drescher DG. Selective expression of serotonin receptor transcripts in the mammalian cochlea and its subdivisions. Brain Res. Mol. Brain Res. 1999;70:135–140. doi: 10.1016/s0169-328x(99)00110-2. [DOI] [PubMed] [Google Scholar]
- Olcese U, Iurilli G, Medini P. Cellular and synaptic architecture of multisensory integration in the mouse neocortex. Neuron. 2013;79:579–593. doi: 10.1016/j.neuron.2013.06.010. [DOI] [PubMed] [Google Scholar]
- Parikh V, Kozak R, Martinez V, Sarter M. Prefrontal acetylcholine release controls cue detection on multiple timescales. Neuron. 2007;56:141–154. doi: 10.1016/j.neuron.2007.08.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pazos A, Cortés R, Palacios JM. Quantitative autoradiographic mapping of serotonin receptors in the rat brain. II. Serotonin-2 receptors. Brain Res. 1985;346:231–249. doi: 10.1016/0006-8993(85)90857-1. [DOI] [PubMed] [Google Scholar]
- Pinto L, Goard MJ, Estandian D, Xu M, Kwan AC, Lee S-H, Harrison TC, Feng G, Dan Y. Fast modulation of visual perception by basal forebrain cholinergic neurons. Nat. Neurosci. 2013;16:1857–1863. doi: 10.1038/nn.3552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rhode WS. Vertical cell responses to sound in cat dorsal cochlear nucleus. J. Neurophysiol. 1999;82:1019–1032. doi: 10.1152/jn.1999.82.2.1019. [DOI] [PubMed] [Google Scholar]
- Rhode WS, Smith PH, Oertel D. Physiological response properties of cells labeled intracellularly with horseradish peroxidase in cat dorsal cochlear nucleus. J. Comp. Neurol. 1983;213:426–447. doi: 10.1002/cne.902130407. [DOI] [PubMed] [Google Scholar]
- Roberts MT, Bender KJ, Trussell LO. Fidelity of complex spike-mediated synaptic transmission between inhibitory interneurons. J. Neurosci. 2008;28:9440–9450. doi: 10.1523/JNEUROSCI.2226-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roberts MT, Seeman SC, Golding NL. A mechanistic understanding of the role of feedforward inhibition in the mammalian sound localization circuitry. Neuron. 2013;78:923–935. doi: 10.1016/j.neuron.2013.04.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ryugo DK, May SK. The projections of intracellularly labeled auditory nerve fibers to the dorsal cochlear nucleus of cats. J. Comp. Neurol. 1993;329:20–35. doi: 10.1002/cne.903290103. [DOI] [PubMed] [Google Scholar]
- Salvinelli F, Casale M, Paparo F, Persico AM, Zini C. Subjective tinnitus, temporomandibular joint dysfunction, and serotonin modulation of neural plasticity: causal or casual triad? Med. Hypotheses. 2003;61:446–448. doi: 10.1016/s0306-9877(03)00194-4. [DOI] [PubMed] [Google Scholar]
- Sedlacek M, Brenowitz SD. Cell-type specific short-term plasticity at auditory nerve synapses controls feed-forward inhibition in the dorsal cochlear nucleus. Front. Neural Circuits. 2014;8:78. doi: 10.3389/fncir.2014.00078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shore SE. Multisensory integration in the dorsal cochlear nucleus: unit responses to acoustic and trigeminal ganglion stimulation. Eur. J. Neurosci. 2005;21:3334–3348. doi: 10.1111/j.1460-9568.2005.04142.x. [DOI] [PubMed] [Google Scholar]
- Shore SE, Vass Z, Wys NL, Altschuler RA. Trigeminal ganglion innervates the auditory brainstem. J. Comp. Neurol. 2000;419:271–285. doi: 10.1002/(sici)1096-9861(20000410)419:3<271::aid-cne1>3.0.co;2-m. [DOI] [PubMed] [Google Scholar]
- Simpson JJ, Davies WE. A review of evidence in support of a role for 5-HT in the perception of tinnitus. Hear. Res. 2000;145:1–7. doi: 10.1016/s0378-5955(00)00093-9. [DOI] [PubMed] [Google Scholar]
- Smith PH, Rhode WS. Electron microscopic features of physiologically characterized, HRP-labeled fusiform cells in the cat dorsal cochlear nucleus. J. Comp. Neurol. 1985;237:127–143. doi: 10.1002/cne.902370110. [DOI] [PubMed] [Google Scholar]
- Spirou GA, Davis KA, Nelken I, Young ED. Spectral integration by type II interneurons in dorsal cochlear nucleus. J. Neurophysiol. 1999;82:648–663. doi: 10.1152/jn.1999.82.2.648. [DOI] [PubMed] [Google Scholar]
- Stein BE, Stanford TR. Multisensory integration: current issues from the perspective of the single neuron. Nat. Rev. Neurosci. 2008;9:255–266. doi: 10.1038/nrn2331. [DOI] [PubMed] [Google Scholar]
- Steinbusch HW. Distribution of serotonin-immunoreactivity in the central nervous system of the rat-cell bodies and terminals. Neuroscience. 1981;6:557–618. doi: 10.1016/0306-4522(81)90146-9. [DOI] [PubMed] [Google Scholar]
- Sutherland DP, Glendenning KK, Masterton RB. Role of acoustic striae in hearing: discrimination of sound-source elevation. Hear. Res. 1998;120:86–108. doi: 10.1016/s0378-5955(98)00056-2. [DOI] [PubMed] [Google Scholar]
- Tang Z-Q, Trussell LO. Serotonergic regulation of excitability of principal cells of the dorsal cochlear nucleus. J. Neurosci. 2015;35:4540–4551. doi: 10.1523/JNEUROSCI.4825-14.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thompson AM, Thompson GC. Serotonin projection patterns to the cochlear nucleus. Brain Res. 2001;907:195–207. doi: 10.1016/s0006-8993(01)02483-0. [DOI] [PubMed] [Google Scholar]
- Thompson AM, Wiechmann AF. 5-HT(1A) receptor subtype mRNA expression in cochlear nucleus. Hear. Res. 2002;164:77–81. doi: 10.1016/s0378-5955(01)00413-0. [DOI] [PubMed] [Google Scholar]
- Thompson GC, Thompson AM, Garrett KM, Britton BH. Serotonin and serotonin receptors in the central auditory system. Otolaryngol. Head Neck Surg. 1994;110:93–102. doi: 10.1177/019459989411000111. [DOI] [PubMed] [Google Scholar]
- Thompson AM, Moore KR, Thompson GC. Distribution and origin of serotoninergic afferents to guinea pig cochlear nucleus. J. Comp. Neurol. 1995;351:104–116. doi: 10.1002/cne.903510110. [DOI] [PubMed] [Google Scholar]
- Voigt HF, Young ED. Cross-correlation analysis of inhibitory interactions in dorsal cochlear nucleus. J. Neurophysiol. 1990;64:1590–1610. doi: 10.1152/jn.1990.64.5.1590. [DOI] [PubMed] [Google Scholar]
- Weinberg RJ, Rustioni A. A cuneocochlear pathway in the rat. Neuroscience. 1987;20:209–219. doi: 10.1016/0306-4522(87)90013-3. [DOI] [PubMed] [Google Scholar]
- Willard FH, Ho RH, Martin GF. The neuronal types and the distribution of 5-hydroxytryptamine and enkephalin-like immunoreactive fibers in the dorsal cochlear nucleus of the North American opossum. Brain Res. Bull. 1984;12:253–266. doi: 10.1016/0361-9230(84)90053-4. [DOI] [PubMed] [Google Scholar]
- Wood WE, Roseberry TK, Perkel DJ. HTR2 receptors in a songbird premotor cortical-like area modulate spectral characteristics of zebra finch song. J. Neurosci. 2013;33:2908–2915. doi: 10.1523/JNEUROSCI.4291-12.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wright DE, Seroogy KB, Lundgren KH, Davis BM, Jennes L. Comparative localization of serotonin1A, 1C, and 2 receptor subtype mRNAs in rat brain. J. Comp. Neurol. 1995;351:357–373. doi: 10.1002/cne.903510304. [DOI] [PubMed] [Google Scholar]
- Wu C, Stefanescu RA, Martel DT, Shore SE. Tinnitus: maladaptive auditory-somatosensory plasticity. Hear. Res. 2016;334:20–29. doi: 10.1016/j.heares.2015.06.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Young ED, Davis KA. Circuitry and function of the dorsal cochlear nucleus. In: Oertel D, Popper AN, Fay RR, editors. Integrative Functions in the Mammalian Auditory Pathway. Springer; 2002. pp. 160–206. [Google Scholar]
- Young ED, Nelken I, Conley RA. Somatosensory effects on neurons in dorsal cochlear nucleus. J. Neurophysiol. 1995;73:743–765. doi: 10.1152/jn.1995.73.2.743. [DOI] [PubMed] [Google Scholar]
- Zhang S, Oertel D. Tuberculoventral cells of the dorsal cochlear nucleus of mice: intracellular recordings in slices. J. Neurophysiol. 1993a;69:1409–1421. doi: 10.1152/jn.1993.69.5.1409. [DOI] [PubMed] [Google Scholar]
- Zhang S, Oertel D. Cartwheel and superficial stellate cells of the dorsal cochlear nucleus of mice: intracellular recordings in slices. J. Neurophysiol. 1993b;69:1384–1397. doi: 10.1152/jn.1993.69.5.1384. [DOI] [PubMed] [Google Scholar]
- Zhou J, Shore S. Projections from the trigeminal nuclear complex to the cochlear nuclei: a retrograde and anterograde tracing study in the guinea pig. J. Neurosci. Res. 2004;78:901–907. doi: 10.1002/jnr.20343. [DOI] [PubMed] [Google Scholar]
- Zhou M, Tao HW, Zhang LI. Generation of intensity selectivity by differential synaptic tuning: fast-saturating excitation but slow-saturating inhibition. J. Neurosci. 2012;32:18068–18078. doi: 10.1523/JNEUROSCI.3647-12.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.