

Abstract
Salmonella enterica serovar Typhi (S. Typhi), the causative agent of the typhoid fever, is a pathogen of great public health importance. Typhoid vaccines have the potential to be cost-effective measures towards combating this disease, yet the antigens triggering host protective immune responses are largely unknown. Given the key role of cellular-mediated immunity in S. Typhi protection, it is crucial to identify S. Typhi proteins involved in T-cell responses. Here, cells from individuals immunized with Ty21a typhoid vaccine were collected before and after immunization and used as effectors. We also used an innovative antigen expressing system based on the infection of B-cells with recombinant Escherichia coli (E. coli) expressing one of four S. Typhi gene products (i.e., SifA, OmpC, FliC, GroEL) as targets. Using flow cytometry, we found that the pattern of response to specific S. Typhi proteins was variable. Some individuals responded to all four proteins while others responded to only one or two proteins. We next evaluated whether T-cells responding to recombinant E. coli also possess the ability to respond to purified proteins. We observed that CD4+ cell responses, but not CD8+ cell responses, to recombinant E. coli were significantly associated with the responses to purified proteins. Thus, our results demonstrate the feasibility of using an E. coli expressing system to uncover the antigen specificity of T-cells and highlight its applicability to vaccine studies. These results also emphasize the importance of selecting the stimuli appropriately when evaluating CD4+ and CD8+ cell responses.
Author summary
Salmonella enterica serovar Typhi (S. Typhi) is the causative agent of the life-threatening typhoid fever that affects 11.9–20.6 million individuals annually in low-income and middle-income countries. The T-cells, CD4+ and CD8+ T cells, play a significant role in protection against S. Typhi infection. Yet, the antigens triggering host protective immune responses recognized by these cells are largely unknown. To address this shortcoming, in this study we used an E. coli expression system methodology for identifying immunogenic proteins of S. Typhi. We found that although the pattern of response to individual S. Typhi proteins was variable among the typhoid vaccinees, the E. coli expressing system uncovered the antigen specificity of T-cells, and highlight its applicability to vaccine studies.
Introduction
Typhoid fever is caused by Salmonella enterica serovar Typhi (S. Typhi), a human-restricted pathogen that enters the host through the gut-associated lymphoid tissue. Recent calculations of the typhoid burden estimated that 11.9–20.6 million new cases of typhoid fever occur annually in low-income and middle-income countries with about 129,000–223,000 mortality [1–4]. Based on data provided by the World Health Organization, 90 percent of these typhoid deaths occur in Asia, and most victims are children under five years of age [5]. Furthermore, antimicrobial treatment of enteric fever and asymptomatic carriers has become increasingly complicated due to the emergence of multidrug-resistant strains of S. Typhi [6–8]. Thus, there has been an increased emphasis on control measures, such as vaccination to fight S. Typhi infection [9, 10]. It has also become evident that a better understanding of the host immune responses against S. Typhi will be required to achieve this task. Currently, two typhoid vaccines are licensed in the USA for use in humans, the purified Vi (“virulence”) parenteral polysaccharide vaccine and the oral live attenuated S. Typhi strain Ty21a vaccine. Although these vaccines are moderately protective and able to induce herd immunity [11, 12], they also have some significant shortcomings. Since Vi is a T-cell independent antigen, Vi vaccine does not confer “memory,” and there are no robust data to suggest that the efficacy of Vi persists beyond three years [11, 13, 14]. The Ty21a vaccine, which does not elicit anti-Vi antibodies, requires the administration of three to four doses spaced at 48-hour intervals [12, 13, 15]. Moreover, recently, Vi-protein-conjugate vaccines that consist of the S. Typhi Vi polysaccharide covalently bound to a carrier protein have been developed [5, 16–19]. However, issues have been raised about selective pressure for the development and spread of S. Typhi Vi antigen-negative strains due to the generalized use of Vi and Vi-protein-conjugate vaccines [20, 21]. As a result, novel approaches to typhoid vaccination are critically needed [22].
It is now widely accepted that cellular-mediated immunity (CMI) plays a significant role in protection against S. Typhi infection [8]. These host responses rely mainly on two types of T-cells, CD4+ and CD8+ T cells [23–26]. The presence of both CD4+ helper T-cells and classical class Ia and non-classical HLA-E-restricted S. Typhi-specific CD8+ T cells have been observed in individuals who recover from typhoid fever [25] or immunized with Ty21a and other attenuated leading typhoid vaccine candidates, including CVD 906, CVD 908, CVD 908-htrA and CVD 909 [26–33]. Moreover, our group recently provided the first evidence that S. Typhi-specific CD8+ responses correlate with clinical outcome in humans challenged with wild-type S. Typhi [34]. However, the antigen specificity of these T cells remains largely unknown. Moreover, most of the S. Typhi proteins described as being involved in human protection have been derived from studies using mouse models of Salmonella infection [35, 36]. One of the reasons for this is the inherent problems of working with humans as experimental models.
Here, we used an innovative antigen expressing system, originally developed by the Higgins laboratory [37, 38] and based on the infection of B-cells with recombinant E. coli to evaluate T cell responses to four S. Typhi proteins: SifA, FliC, GroEL, and OmpC (Table 1). These proteins are known to confer survival properties to Salmonella and therefore might be evaluated as vaccine antigens [27, 39–44]. Briefly, in this system, EBV-transformed lymphoblastoid B-cell lines (B-LCL) were used as antigen-presenting cells (APCs). These B-LCL were infected with E. coli expressing both S. Typhi proteins and cytoplasmic listeriolysin O (Hly). Hly is a pore-forming hemolysin from Listeria monocytogenes, which allows leakage of E. coli antigen from the phagolysosomal compartment into the APC cytosol, there gaining access to the MHC class I antigen processing and presentation pathway [37, 38]. This system also allows the identification of S. Typhi-specific CD4+ T cell as the expression on E. coli also results in antigen presentation in the context of MHC class II molecules [45]. Additionally, this approach has the advantage of assessing T-cell responses to full-length proteins before initiating more expensive and time-consuming procedures, such as synthesizing overlapping peptides [46]. Due to HLA diversity in humans, host responses to subunit vaccines have a greater chance to be successful if they encompass specific protein antigens rather than specific epitopes within those proteins [45, 46].
Table 1. S. Typhi proteins evaluated in this manuscript.
| Protein Name | Gene Name | Function | MW (kd) |
|---|---|---|---|
| Putative virulence determinant | SifA | Required for endosomal tubulation and formation of Salmonella-induced filaments (Sifs) | 38.6 |
| Flagellin |
FliC | Bacterial-type flagellum-dependent cell motility | 53.2 |
| Chaperonin |
GroEL | Promotes protein refolding | 57.2 |
| Outer membrane protein C |
OmpC | Forms pores that allow passive diffusion of small molecules across the outer membrane | 41.2 |
By using this innovative antigen expression system, we found that the pattern of response to individual S. Typhi proteins was variable. Some individuals responded to all four proteins while others responded to only one or two proteins. When comparing T cells responses to B-LCL exposed to recombinant E. coli to those to purified proteins from the same genes, we observed that the CD4+ cell responses, but not CD8+ cell responses, to recombinant E. coli were significantly associated with the responses to purified proteins. Thus, our results demonstrate the feasibility of using an E. coli expressing system to uncover the antigen specificity of T-cells, and highlight its applicability to vaccine studies. These results also emphasize the importance of selecting the stimuli appropriately when designing experiments aimed at evaluating CD4+ and CD8+ cell responses.
Results
Expression of recombinant proteins
To show the feasibility of our E. coli expressing system, we evaluate four S. Typhi proteins (i.e., SifA, FliC, GroEL, and OmpC) (Table 1) known to confer survival properties to Salmonella then potentially promising as vaccine antigens [27, 39–44]. As shown in Fig 1, proper E.coli protein expression for all four proteins, SifA, OmpC, FliC, and GroEL, as well as the Hly was detected by Western blot.
Fig 1. Expression of S. Typhi proteins and lysteriolysin on recombinant E. coli.

(A) Anti-HisTag antibody revealing positive control protein Annexin3 (A3) as well as lysteriolysin (Hly) and S. Typhi gene encoded proteins (OmpC, FliC, GroEL and SifA). A negative control with pmark (P, stop codons) clones was used instead of the protein. (B) Anti-lyteriolysin antibody revealing Hly gene expression (red box) in all lanes except in the negative control. Expression of S. Typhi proteins and lysteriolysin on recombinant E. coli were detected by Western blot.
We next evaluated the effect of the recombinant E. coli infection on B-LCL viability. Briefly, we assessed cell viability by measuring the levels of Yevid viability staining on 2-hour-E. coli infected B-LCLs that have been rested overnight in the presence of gentamicin. As shown in Fig 2A, regardless of the protein being expressed, the infection did not adversely affect the viability of E. coli-infected B-LCLs. After infection, the percentage of live cells in cultures with recombinant E. coli was comparable to control cultures with media only (uninfected). By using the same experimental conditions as for determinations of cell viability, we also detected the expression of bacterial antigens on B-LCLs. Similarly to the viability, regardless of the type of protein being expressed in the recombinant E. coli, we found similar levels of E. coli-expressing cells as assessed by surface staining with anti-E. coli antigen polyclonal antibody using flow cytometry (Fig 2B–2D).
Fig 2. Expression of bacterial antigens on B-LCL target cells.

B-LCL cells were infected with E. coli at 1:30 MOI with one of the four recombinant E. coli expressing S. Typhi and Hly antigens: Hly/SifA (SifA), Hly/FliC (FliC), Hly/GroEL (GroEL) and Hly/OmpC (OmpC). Uninfected B-LCLs (uninfected) and infected with recombinant E. coli expressing only Hly antigen were used as controls. The (A) viability and the (B) percentage of the E. coli-expressing cells were assessed by flow cytometry. Numbers correspond to the % of positive cells in the indicated quadrant in each histogram followed by mean fluorescence intensity (MFI) of positive cells (in parenthesis). Cumulative data of (C) % and (D) mean fluorescence intensity (MFI) were detected by using anti-E. coli antibody as described in Methods. Average of 3 independent experiments.
Hly functionality
As described above, we reasoned that Hly should promote the phagosomal escape of bacterial antigens thereby improving MHC class I processing of S. Typhi antigens presented by B-LCLs and hence recall immune responses from both CD4+ and CD8+ primed T cells [8, 26, 27, 29, 30, 39, 47]. To test this assumption, Hly-recombinant E. coli strain BL21, or wild type E. coli strain BL21 were used to infect B-LCL cells. Cells were infected for 2 hours using two different multiplicity of infection (MOI, 1:30 and 1:100). After 2 hours, cells were collected, washed to remove extracellular bacteria and cultured in the presence of gentamicin for 2 additional hours. Thus, the ability to detect E. coli proteins in B-LCL infected cells was assessed over time by flow cytometry (up to 120 minutes) using polyclonal anti-E. coli antibodies. As shown in Fig 3, at all-time points evaluated, we observed higher expression of E. coli antigens on B-LCL cells infected with the recombinant E. coli strain expressing Hly as compared to the wild-type E. coli strain. Thus, the hly gene appears functional. These results are very significant since based on our previous study [48], we expect to see background responses against the vector itself (E. coli antigens). Further antigen expression might help to better discriminate T-cell responses to S. Typhi antigens from those directed to E. coli antigens.
Fig 3. Ability of the Hly gene to improve antigen processing.
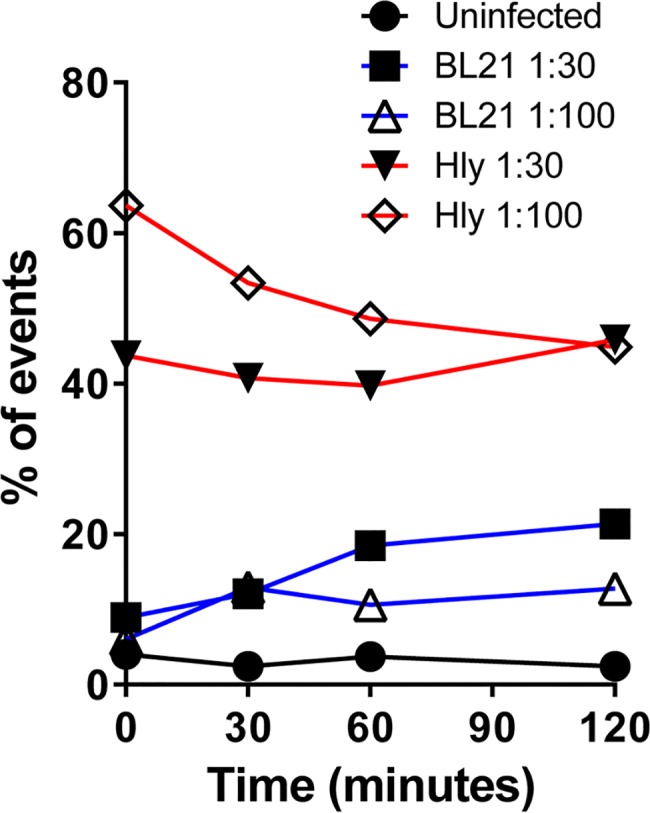
B-LCL cells were infected with either E. coli strain BL21 (BL21) or its Hly-recombinant E. coli (Hly) counterpart at 1:30 or 1:100 multiplicity of infection (MOI). After 2 hours, the cells were washed and exposed to gentamicin for an additional 2 hours to kill and detach extracellular bacteria. After further washings, the ability of the Hly gene to improve antigen processing was assessed by detecting E. coli antigens at the B-cell surface over time by flow cytometry (up to 120 minutes). Cells exposed to media only were used as control (uninfected).
Identification of S. Typhi proteins that are targets of T-cells following oral immunization with Ty21a
In order to demonstrate the feasibility of using an E. coli expression system to uncover the antigen specificity of T-cells, PBMC obtained before and 42 days after immunization were exposed to autologous B-LCL infected with recombinant E. coli expressing Hly only or co-expressing one of the four Salmonella gene products: Hly/SifA, Hly/OmpC, Hly/FliC and Hly/GroEL. Specifically, we studied the ability of ex-vivo PBMC from seven Ty21a-immunized volunteers to express IL-17A, IFN-γ and TNF-α cytokines and/or CD107a and b molecules against autologous infected targets. T-cell responses (i.e., CD4+ and CD8+ T-cells) were evaluated by multichromatic flow cytometry using a 10-color surface/intracellular staining panel. Unstimulated and Staphylococcus enterotoxin B (SEB)-stimulated effector cells were used as negative and positive controls, respectively. We observed that the pattern of response to individual S. Typhi proteins was variable, with some individuals responding to all four proteins while others were responding to only one or two proteins. We also observed differential CD4+ and CD8+ T cells responses to the S. Typhi proteins (Figs 4–7). In six individuals, although the magnitude of responses varied considerably, both CD4+ and CD8+ T cells responded to at least one protein. In one individual, we were unable to detect CD4+ T cells responses to any of the four protein, but CD8+ T cells responded to 3 out of 4 proteins tested (Fig 8). Representative responses from selected volunteers are shown in Figs 4–6.
Fig 4. Antigen presentation of S. Typhi proteins by targets infected with recombinant E. coli.
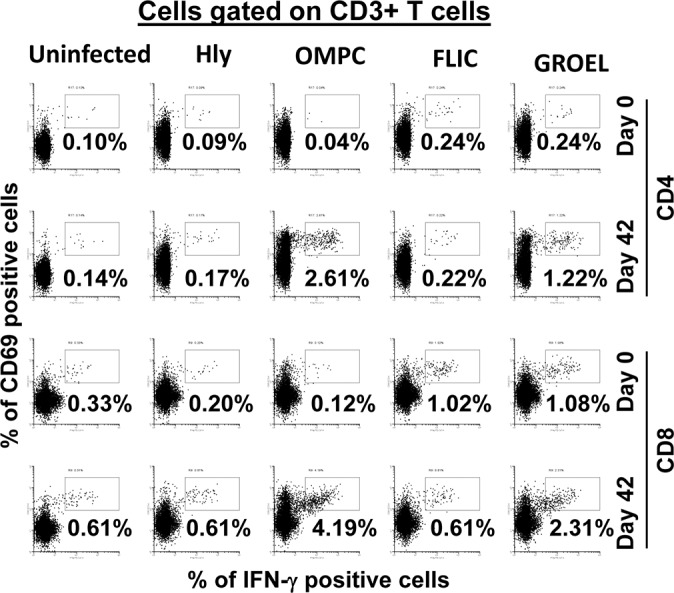
Ex vivo PBMC from a volunteer collected before (day 0) and 42 days after immunization were co-cultured for 16–18 hrs. with autologous B-LCL targets infected at 1:30 MOI with one of the four recombinant E. coli expressing S. Typhi/Hly (Hly/SifA (SifA), Hly/FliC (FliC), Hly/GroEL (GroEL) and Hly/OmpC (OmpC)) or only Hly (control) proteins. After incubation, cells were stained and the ability of the PBMC to express IFN-γ was analyzed by flow cytometry. CD4+ and CD8+ T cells were evaluated. Numbers represent the percentage of positive cells. The data of a representative volunteer are shown.
Fig 7. Percentage of T cell subsets specific to any S. Typhi protein.
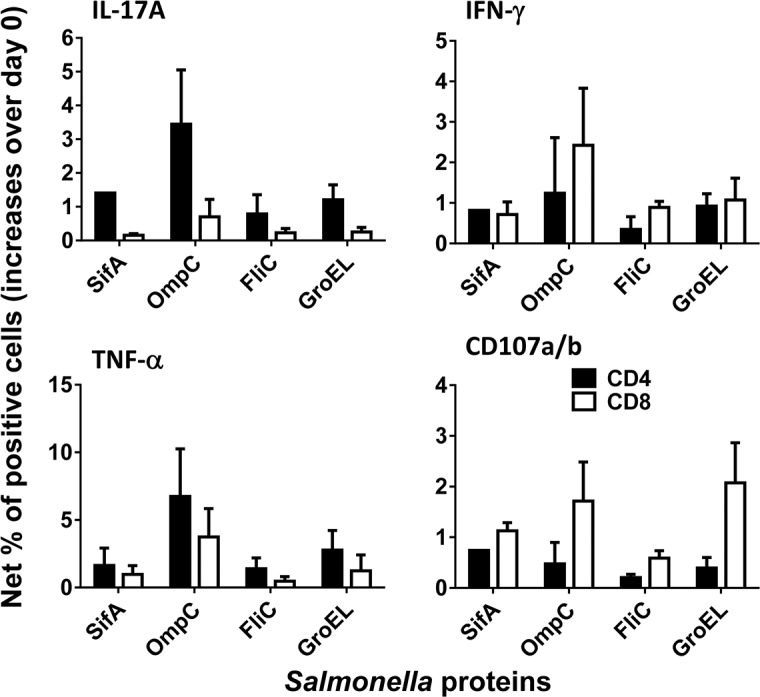
Ex vivo PBMC from 7 volunteers collected before and 42 days after immunization were co-cultured for 16–18 hrs. with autologous B-LCL targets infected at 1:30 MOI with one of the four recombinant E. coli expressing S. Typhi and Hly genes: Hly/SifA (SifA), Hly/FliC (FliC), Hly/GroEL (GroEL) and Hly/OmpC (OmpC). After incubation, cells were stained and the ability of the PBMC to express one or more cytokines (IL-17A, IFN-γ and TNF-α) and/or CD107a/b molecules was analyzed by flow cytometry. Two T cell subset responses (i.e., CD4+ and CD8+ T cells) were evaluated. Net responses were calculated by subtracting the T cell responses to B-LCLs infected with recombinant E. coli expressing S. Typhi/Hly antigens from the responses to the controls (B-LCL expressing Hly only). Increases over day 0 were calculated by subtracting the net responses of the PBMC collected 42 days after immunization from the net responses of PBMC collected before immunization. Bars represent mean ± SE.
Fig 8. Volunteer responses to S. Typhi proteins.

Ex vivo PBMC from 7 immunized volunteers collected before (day 0) and 42 days after immunization were co-cultured for 16–18 hrs. with autologous B-LCL targets infected at an 1:30 MOI with one of the four recombinant E. coli expressing S. Typhi and Hly antigens: Hly/SifA (SifA), Hly/FliC (FliC), Hly/GroEL (GroEL) and Hly/OmpC (OmpC). After incubation, cells were stained and the ability of the PBMC to express one or more cytokines (IL-17A, IFN-γ and TNF-α) and/or CD107a/b molecules was analyzed by flow cytometry. Two T cell subset responses (i.e., CD4+ and CD8+ T cells) were evaluated. (A) Heat-map of the multifunctionality of CD4+ and CD8+ T cells based on expression of cytokines and CD107a/b antigens. Percentages correspond to the net responses calculated by subtracting the T cell responses to B-LCLs infected with recombinant E. coli expressing S. Typhi/Hly proteins from the responses to the controls (B-LCL expressing Hly only). Volunteers were considered responders if the net responses of the PBMC collected 42 days after immunization were greater than 0.1 from the net responses of PBMC collected before immunization. (B) Cumulative frequency of responders to any functional test.
Fig 6. CD4+ T cell responses to S. Typhi proteins presented by targets infected with recombinant E. coli.
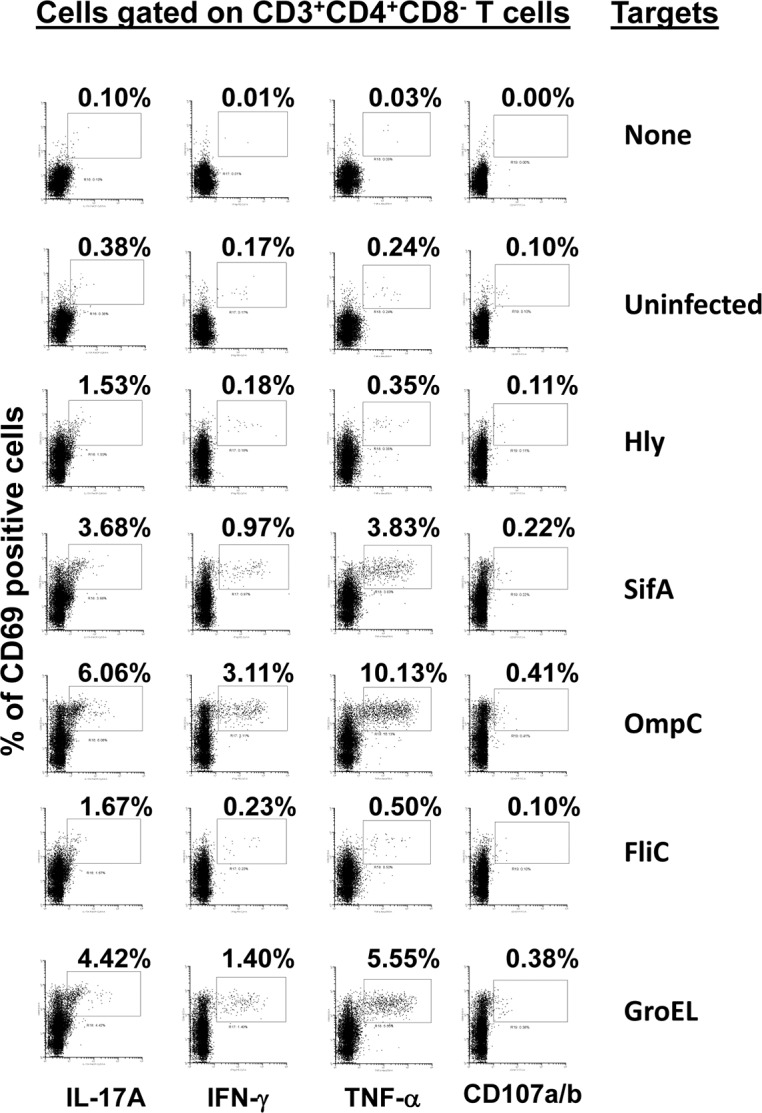
Ex vivo PBMC from a volunteer collected 42 days after immunization were co-cultured for 16–18 hrs. with autologous B-LCL targets infected at 1:30 MOI with one of the four recombinant E. coli expressing S. Typhi/Hly (Hly/SifA (SifA), Hly/FliC (FliC), Hly/GroEL (GroEL) and Hly/OmpC (OmpC)) or only Hly (control) proteins. After incubation, cells were stained and the ability of the PBMC to express one or more cytokines (IL-17A, IFN-γ and TNF-α) and/or CD107a/b molecules was evaluated by flow cytometry. Shown are the CD4+ T cell responses from a representative volunteer. Numbers represent the percentage of positive cells.
Fig 5. CD8+ T cell responses to S. Typhi proteins presented by targets infected with recombinant E. coli.
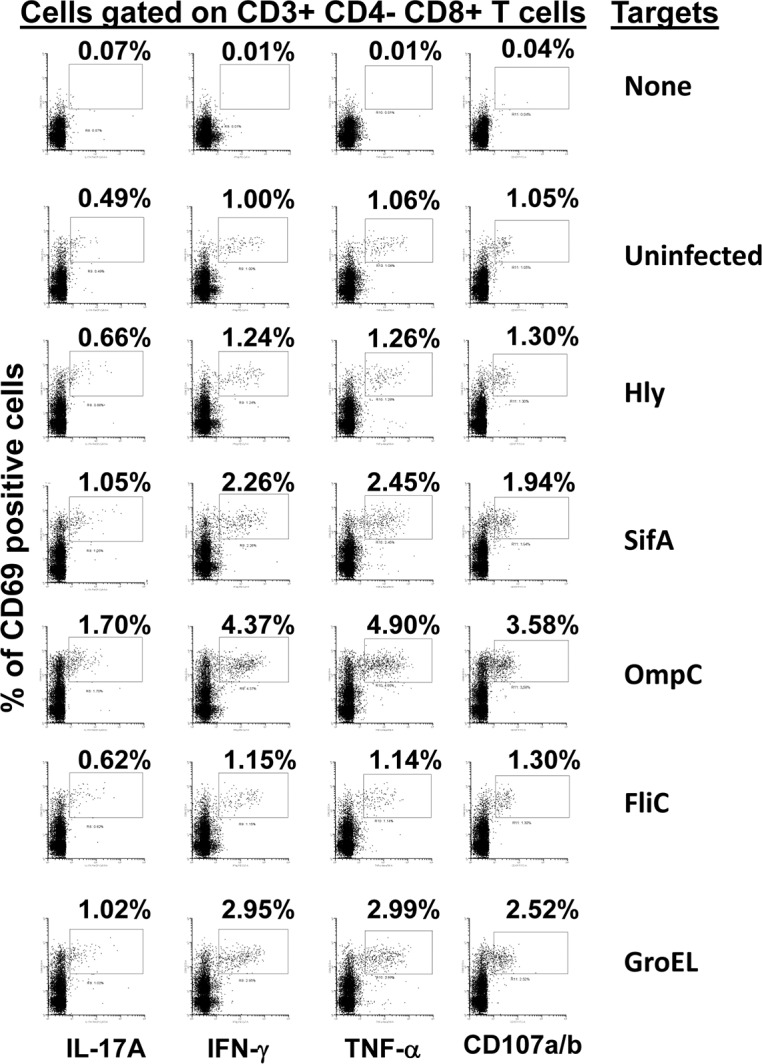
Ex vivo PBMC from a volunteer collected 42 days after immunization were co-cultured for 16–18 hrs. with autologous B-LCL targets infected at 1:30 MOI with one of the four recombinant E. coli expressing S. Typhi/Hly (Hly/SifA (SifA), Hly/FliC (FliC), Hly/GroEL (GroEL) and Hly/OmpC (OmpC)) or only Hly (control) proteins. After incubation, cells were stained and the ability of the PBMC to express one or more cytokines (IL-17A, IFN-γ and TNF-α) and/or CD107a/b molecules was evaluated by flow cytometry. Shown are the CD8+ T cell responses from a representative volunteer. Numbers represent the percentage of positive cells.
Multifunctional of CD4+ T-cells and CD8+ T- cells
Because previous results from our group have shown that multifunctional T-cells might contribute to S. Typhi immunity [30, 34, 49], we then investigated the multi-functionality patterns of CD4+ T-cells and CD8+ T-cells after exposure to infected B-LCL infected with recombinant E. coli. We measured simultaneously four T-cell functions (i.e., expression of IL-17A, IFN-γ and TNF-α cytokines, or CD107a and b molecules) by multichromatic flow cytometry using the FCOM feature of the WinList software, which provides the % of T-cells expressing each of the possible function combinations. Analyses of multiple function patterns (i.e., single, double, triple or quadruple functions) revealed that, albeit different for different proteins, both CD4+ T-cells and CD8+ T-cells were multi-functional (Fig 8).
T-cell responses to exogenous proteins
An important hypothesis to evaluate is whether the T-cells from volunteers who responded to B-LCL cultured with recombinant E. coli expressing Salmonella gene products also possess the ability to respond to exogenous proteins. To this end, we next investigated whether there was an association between T-cell responses after exposure to autologous B-LCL cultured with recombinant E. coli expressing one of the four Salmonella gene products (Hly/SifA, Hly/OmpC, Hly/FliC and Hly/GroEL) and those T-cell responses after exposure to B-LCL cultured with individual purified recombinant proteins (SifA, OmpC, FliC, and GroEL). To this end, B-LCLs were cultured overnight with purified SifA, OmpC, FliC or GroEL proteins, and used as targets for T-cells. Representative responses of CD8+ and CD4+ cells from one volunteer are shown in Figs 9 & 10 respectively. We found that the CD4+ cell responses to B-LCLs cultured with exogenous proteins were significantly associated with the CD4+ cell responses to B-LCLs cultured with recombinant E. coli expressing one of the four S. Typhi gene products (Fig 11; p = 0.0111, Pearson Product Moment Correlation). However, no significant association was observed between CD8+ cell responses to B-LCLs cultured with recombinant E. coli expressing one of the four Salmonella gene products and CD8+ cell responses to B-LCLs cultured with exogenous recombinant proteins (Fig 11; p = 0.0790, Pearson Product Moment Correlation). On the other hand, CD8+ cell responses to B-LCLs cultured with exogenous proteins were consistently higher than CD4+ cell responses (Figs 9 and 10 & S1 Table). Thus, as expected, CD4+ and CD8+ cell responses against S. Typhi antigens depend on the nature of the stimulant [39, 50]. These results emphasize the importance of selecting the stimuli appropriately when designing experiments aimed at evaluating CD4+ and CD8+ cell responses cell responses [8].
Fig 9. CD8+ T cell responses to S. Typhi proteins presented by targets exposed to one of the four recombinant S. Typhi proteins.

Ex vivo PBMC from a volunteer collected 42 days after immunization were co-cultured for 16–18 hrs. with autologous B-LCL targets exposed to 0.5ug/ml with one of the four recombinant S. Typhi proteins: SifA, OmpC, FliC, and GroEL. Untreated B-LCL targets (media) were used as controls. After incubation, cells were stained and the ability of the PBMC to express one or more cytokines (IL-17A, IFN-γ and TNF-α) and/or CD107a/b molecules was evaluated by flow cytometry. Shown are the CD8+ T cell responses from a representative volunteer. Numbers represent the percentage of positive cells.
Fig 10. CD4+ T cell responses to S. Typhi proteins presented by targets exposed to one of the four recombinant S. Typhi proteins.

Ex vivo PBMC from a volunteer collected 42 days after immunization were co-cultured for 16–18 hrs. with autologous B-LCL targets exposed to 0.5ug/ml with one of the four recombinant S. Typhi proteins: SifA, OmpC, FliC, and GroEL. Untreated B-LCL targets (media) were used as controls. After incubation, cells were stained and the ability of the PBMC to express one or more cytokines (IL-17A, IFN-γ and TNF-α) and/or CD107a/b molecules was evaluated by flow cytometry. Shown are the CD4+ T cell responses from a representative volunteer. Numbers represent the percentage of positive cells.
Fig 11. Correlation between T cell subset responses to B-LCL targets exposed to recombinant S. Typhi proteins or infected with recombinant E. coli expressing S. Typhi proteins.

Ex vivo PBMC were analyzed as described in Figs 1 & 2. Shown are the correlation between T cell subset responses to either of the four proteins (SifA, FliC, GroEL and OmpC) after stimulation by B-LCL targets exposed to recombinant S. Typhi proteins or infected with recombinant E. coli expressing S. Typhi proteins. Samples are representative of two individuals collected 42 days after vaccination. Coefficients of determination “R2” and “p” values are shown. p values of <0.05 were considered statistically significant. Dashed lines represent 95% confidence intervals.
Discussion
One of the characteristics that make the E. coli expression system methodology employed in this manuscript highly relevant for identifying immunogenic proteins of S. Typhi is the use of a translational research approach using human T cells and autologous APC to identify S. Typhi-specific T-cell immune responses. Most published methods have relied heavily on “proof-of-concept” studies performed in mice. However, S. Typhi is a human-restricted pathogen and there are no good animal models that faithfully recapitulate S. Typhi infection [51]. To partially address this shortcoming, the infection of susceptible mice with S. Typhimurium has been used as a model for the pathogenesis of human typhoid fever [51]. Although these murine models have provided considerable knowledge regarding host-pathogen interactions, they do not adequately recapitulate S. Typhi infection in humans [52]. The recent availability of full genome sequences from various Salmonella enterica serovars has uncovered many differences in pseudo genes which can explain, at least in part, the dissimilarities observed in the immune and other host responses to these enteric bacteria [52]. Thus, samples from human participants exposed to the licensed Ty21a oral typhoid vaccine have the potential to provide a better characterization of key Salmonella antigens involved in T cell responses than murine models.
The main novelty of our system is that we engineered the hly gene encoding the pore-forming cytoplasmic listeriolysin O (Hly) protein onto the backbone of the recombinant protein expression plasmid pET-DEST-Hly (see methods). In contrast, Higgin’s E. coli expressing system [37, 38] used two expression plasmids inside individual E. coli cells: one expressing Hly and the other expressing the recombinant protein of interest. The reason for this difference stems from our preliminary results showing that the use of two expression plasmids reduced B-LCL infectivity (S1 Fig). We speculate that this difference is due to the fitness cost for E. coli to maintain, replicate and propagate two plasmids instead of one. There may also have been a negative effect on bacterial cell preparations resulting from growth on media containing two selective antibiotics instead of one.
Herein, using this methodology, we found that all the tested individuals had increased T-cell responses over baseline (before immunization) to at least one of the four S. Typhi proteins evaluated (i.e., SifA, OmpC, FliC, and GroEL). Moreover, multifunctional CD4+ and CD8+ T cells that expressed two or more cytokines (IL-17A, IFN-γ and TNF-α) and/or CD107a/b molecules were detected. These results are particularly significant since we have previously demonstrated that these two T-cell population might play a role in controlling Salmonella infection [39]. These results also support previous data showing that the depletion of either CD4+ or CD8+ T-cells had impaired recall immunity to oral challenge with the virulent S. Typhimurium at different times after vaccination [53].
The reason underlying the observation that the responses to the different antigens are variable among the vaccinees are unclear. However, it is reasonable to speculate that this phenomenon might be due to the HLA‐haplotype variation between individuals. In fact, antigen processing, together with defined MHC genes, are known to shape the individual immune responses to a wide array of pathogens [54]. Furthermore, the differential responsiveness among the participants supports the development of multi-component vaccines by introducing many antigenic determinants into vaccine formulations. Alternatively, these results might encourage a renewed focus on whole-cell live attenuated preparations, especially since they may overcome some of the inherent weaknesses associated with sub-unit vaccines such as the need for considerable amounts of antigens and the use of adjuvants.
We also observed that the magnitude of the responses against S. Typhi SifA, OmpC, FliC and GroEL varied among participants. Since S. Typhi GroEL has a significant degree of homology with self-heat shock proteins in humans [27], these results provide additional information that T-cells can discriminate between self and foreign antigens during the immune response. Interestingly, in contrast to CD8+ cell responses, we found that the CD4+ cell responses to B-LCLs cultured with exogenous proteins were significantly associated with the CD4+ cell responses to B-LCLs cultured with recombinant E. coli expressing S. Typhi genes. These results confirm and extend previous findings from our group and others that the balance of CD4+ and CD8+ cell responses against S. Typhi antigens are likely to depend on the nature of the stimulant [8, 39, 50]. Indeed, previous work from our group has shown that CD4+ cells respond differently to soluble antigen stimulation than CD8+ T cells [39, 50]. Future studies will be directed to use this novel antigen discovery system to evaluate in humans the immune responses to other S. Typhi proteins expressed in this E. coli expression system.
Since these studies were performed in immunized volunteers, it is also important to note that the relative contributions of CD4+ and CD8+ cell responses against these S. Typhi proteins in protection cannot be ascertained. We are directly addressing this critical issue in separate studies in which we are evaluating whether responses to these S. Typhi proteins correlate with protection in volunteers who have been immunized with Ty21a and subsequently challenged with wild-type S. Typhi. Nevertheless, the results presented herein demonstrate the feasibility of using a novel antigen discovery platform. This system could be used, for example, to systematically assess the specificity of T-cell immune responses against the entire proteome of a human pathogen and generate a database of the repertoire of these human T-cell antigen specificities. This should help narrow down the proteins of interest which correlate with defined phenotypes (e.g., responses associated with protection in human challenge studies with S. Typhi, those directed to pathogenic determinants), ultimately leading to the identification of candidate vaccine antigens. Finally, these results emphasize the importance of selecting the appropriate antigens when designing experiments aimed at evaluating CD4+ and CD8+ cell responses.
Materials and methods
Ethics statement
The human experimentation guidelines of the US Department of Health and Human Services and those of the University of Maryland, Baltimore, were followed in the conduct of the clinical research. All blood specimens were collected from volunteers who participated in the University of Maryland Institutional Review Board approved protocol number HP-00040022 that authorized the collection of blood samples from healthy volunteers for the studies included in this manuscript. Volunteers were explained the purpose and possible consequences of participating in this study and gave informed, signed consent before the blood draws. This protocol has been conducted in accordance with the ethical standards laid down in the 1964 Declaration of Helsinki and the principles of the International Conference on Harmonization Good Clinical Practice guidelines [55].
Participants
Seven healthy adult volunteers, 20–50 (39 ± 10) years old, recruited from the Baltimore-Washington area and the University of Maryland at Baltimore campus, participated in this study. They were immunized with four spaced doses of 2–6 x 109 CFU of oral live attenuated Ty21a at an interval of 48 hours between doses [12, 56]. Blood collection was performed before and 42 days after Ty21a immunization. Peripheral blood mononuclear cells (PBMC) were isolated from the blood by density gradient centrifugation and cryopreserved in liquid N2 following standard techniques [28]. These PBMC were used ex vivo as effector cells or to prepare the target cells.
Gateway cloning vectors
The entire S. Typhi strain Ty2 ORFeome was constructed as a part of the former National Institute of Allergy and Infectious Disease (NIAID)-funded Pathogen Functional Genomics Resource Center (PFGRC). The PFGRC provided a library of entry clones by cloning 3,381 ORFs, out of the 4,323 ORFs predicted in the Ty2 genome (78%), into the pDONR 221 vector using methods described in Peterson et al. [57]. Entry clones can then be shuttled into a variety of destination vectors using in vitro site-specific recombination (http://www.ncbi.nlm.nih.gov/pubmed/11076863). For expression of the S. Typhi proteins, we engineered the pET161-DEST destination vector (Invitrogen, Carlsbad, CA). The pET161-DEST, a commercial plasmid from Invitrogen, is a robust E. coli destination vector that has been used for several years and carries many desirable characteristics for this study. Gene expression is under the tight control of the T7 promoter and lac repressor, and an optimized ribosome binding site is provided by the inserted gene. The vector contains the V5 and His-tag epitopes downstream of the attR sites into which inserts from entry clones were recombined resulting in the addition of these tags as C-terminal fusions to the S. Typhi protein. These tags were used to monitor recombinant protein expression and, if necessary, for protein purification. pET161-DEST also contains the ccdB and chloramphenicol (CmR) genes which help the recovery of correct recombination products after the LR recombinase reaction via selection on ampicillin agar plates. We also inserted the hly gene coding for the cytoplasmic Hly protein from L. monocytogenes into the BglII restriction site. The hly gene was PCR-amplified with a His-tag at the 3’end of the reverse primer, and a BgIll site was added to the forward 5’ end, and reverse 3’ end of each primer, respectively (sense: GCGCAGATCTAGCAAGCATATAATATTGCG, anti-sense: GCGCAGATCTTTAGTGATGGTGATGGTGATGTTCGATTGGATTATCTAC). The resulting vector pET-DEST-Hly was verified by Sanger sequencing. It carries a T7 promoter and the lac operator preceding the gene to be cloned; the cloning site harbors attR1 and attR2 site for cloning from entry clones through the LR clonase reaction (see below). The plasmid was transformed into One Shot ccdB survival competent cells (Invitrogen).
Cloning of S. Typhi proteins from entry clones to pET-DEST-Hly for protein expression
Entry clone glycerol stocks were streaked on LB agar with 50 μg/ml kanamycin and incubated at 37°C overnight. Single colonies were incubated overnight at 37°C in 4ml LB broth with 50 μg/ml kanamycin. Then the entry clone plasmid was extracted using the QIA prep kit (Qiagen, Valencia, CA) and DNA concentration measured using a NanoDrop instrument (Thermo Scientific, Waltham, MA). The pET-DEST-Hly glycerol stock was streaked on LB agar plates with 100 μg/ml carbenicillin and incubated at 37°C. After overnight incubation, single colonies were isolated, incubated overnight in 50 ml LB broth with 100 μg/ml carbenicillin at 37°C and the destination plasmid was extracted with the HiSpeed plasmid preparation kit (Qiagen). DNA concentration was measured by NanoDrop. We chose to evaluate four S. Typhi proteins: SifA, FliC, GroEL, and OmpC (Table 1). Each of these four S. Typhi proteins was then transferred in vitro from the entry clone plasmid into the destination plasmid through LR clonase reactions (Gateway LR clonase II Enzyme mix, Invitrogen) following the manufacturer’s protocol. We also cloned a short non-coding sequence called “pmark” that carried stop codons in all six frames of translation into pET-DEST-Hly as a negative control of protein expression. Finally, 3 μl of LR clonase reaction product was transformed into 50 μl of One Shot BL21(DE3) competent cells (Invitrogen) by chemical transformation.
Verification of expression plasmids
Proper cloning of the target S. Typhi gene was confirmed by PCR amplification using the T7 forward and reverse primers and sequencing of the PCR products. The presence of the hly gene was verified by BgIll digestion of the plasmid.
Protein expression
A single colony from verified BL21 (DE3) expression vector clones was cultured at 37°C overnight in 2 ml LB broth with 100 μg/ml carbenicillin. Then 30 μl of the overnight culture was inoculated to 3 ml fresh LB broth with 100 μg/ml carbenicillin. After ~4h of incubation at 37°C (OD~0.6), the culture was induced for protein expression with 100 μM IPTG and incubated for an additional 2 h. Bacteria were then spun down at 2,800 rpm for 10 min and the supernatant discarded. E. coli were lysed with of 0.3 μg/ml lysozyme in Tris-HCl buffer, pH 7.5 (Thermo Scientific), then proteins were heat-denatured at 100°C for 10 minutes and evaluated on SDS-PAGE gels by Coomassie staining. Specificity of protein expression was confirmed by western blot using mouse anti-His monoclonal antibodies (Sigma, St. Louis, MO), and rabbit anti-mouse IgG conjugated to horseradish peroxidase (HRP) (Sigma) as secondary antibodies; as well as rabbit anti-Listeriolysin polyclonal antibodies (Abcam, Cambridge, MA, USA), and goat anti-rabbit IgG-HRP (Millipore, Billerica, MA) as secondary antibodies. His-tagged Annexin 3 (A3) protein (Abcam) was used as positive control for antibody detection.
Target/stimulator cells
PBMC obtained from Ty21a vaccines before immunization were used to generate autologous Epstein-Barr Virus (EBV)-transformed B-LCLs as previously described [27, 31]. Briefly, B-LCLs were established by using B95-8 cell line (ATCC# CRL1612) supernatants as the EBV source. After transformation, B-LCL were maintained in culture in RPMI 1640 (Gibco, Grand Island, New York) supplemented with 100 U/ml penicillin, 100 μg/ml streptomycin, 50 μg/ml gentamicin, 2 mM L-glutamine, 2.5 mM sodium pyruvate, 10 mM HEPES buffer and 10% heat-inactivated fetal bovine serum (R10) or cryopreserved until used in the experiments.
Infection of target/stimulator cells by E. coli
Target/stimulator cells were infected as previously described [27–30, 39, 47, 48] with slight modifications. Briefly, target cells were infected by incubation in RPMI (without antibiotics) at 37°C for 2 hours with any of the recombinant E. coli strains at 1:30 or 1:100 multiplicity of infection (MOI). After incubation, cells were washed and incubated for an additional 2 or 16–18 hours (overnight) in complete R10 containing gentamicin (100 μg/ml) to kill extracellular and/or to detach cell-bound bacteria. For co-culture experiments, targets were then gamma-irradiated (6,000 rads), surface stained with anti-CD45, a marker abundantly expressed on the surface of hematopoietic cells [58], and used as stimulators. To confirm E. coli infection, aliquots of targets were surface stained with rabbit anti-E. coli antigen polyclonal antibody (1:1000, Abcam).
Exogenous proteins
Four Salmonella purified proteins were tested: SifA, FliC, GroEL, and OmpC (Table 1). The region encoding residues 53–450 of FliC were subcloned from S. Typhi ISP1820 into pET15b. The plasmid was used to transform E. coli Tuner (DE3) for overexpression after being induced with IPTG. The overexpressed protein was purified by standard immobilized metal affinity column chromatography (IMAC) methods. OmpC was purified based on the protocol of Kumar and Krishnaswamy [59]. The sequence encoding OmpC from S. Typhi ISP1820 was subcloned into pET15b, which was used to transform E. coli Tuner (DE3). Overexpression was induced using IPTG, and the OmpC was purified from inclusion bodies using IMAC after solubilizing in buffer containing urea. Refolding was then allowed to occur in 50 mM Tris, pH 8.5, 0.1 M NaCl, 10% (v/v) glycerol and 0.2% (v/v) polyoxyethelene-9-lauryl ether. The sequence for GroEL was introduced into pBAD TOPO TA such that it was in-frame. The protein was overexpressed by the addition of arabinose, and the HT-GroEL purified initially by standard IMAC, which provides good yields at high purity. Removal of contaminating peptides could be achieved by adding acetone to 45% (v/v) to precipitate the GroEl which could then be collected by centrifugation followed by solubilizing using PBS (10 mM phosphate, pH 7.4, 150 mM NaCl). The sequence encoding SifA from Salmonella Typhi ISP1820 was subcloned into pET15b and used to transform E. coli c-41 (DE3). The bacteria were grown to an absorbance of about 0.8 at 600 nm, protein expression induced with IPTG, and the cultures immediately moved to 20°C for overnight growth. The bacteria were then lysed and the SifA purified by standard IMAC methods.
Loading of target/stimulator cells with exogenous proteins
B-LCLs were incubated overnight in 24-well plates at a density of 2 x 106/ml/well in the absence or presence of 1 μg/ml of each of the purified proteins at 37°C in a 5% CO2 atmosphere. After incubation, cells were washed and used as stimulators for T-cells.
Monoclonal antibodies for surface and intracellular staining
Cells were stained with monoclonal antibodies (mAbs) to CD69 (clone TPI-55-3) (Beckman-Coulter, Miami, FL), CD4 (clone RPA-T4), CD8 (clone HIT8a), CD107a and b (clones H4A3 and H4B4 respectively), interferon (IFN)-γ (clone B27), tumor necrosis factor (TNF)-α (clone MAb11) (BD Pharmingen, San Diego, CA, USA), CD14 (clone TuK4), CD19 (clone SJ25-C1), CD45 (clone H130) (Invitrogen), interleukin (IL)-17A (clone eBio64DEC17) (eBioscience, San Diego, CA), and CD3 (clone OKT3)(Biolegend, San Diego, CA). Antibodies conjugated to the following fluorochromes were used in these studies: Fluorescein isothiocyanate (FITC), PE-Cy5.5, PE-Cy7, V450, Brilliant Violet (BV)570, BV650, Energy Coupled Dye or PE-Texas-Red conjugate (ECD), allophycocyanin (APC)-Alexa 700 and Quantum Dot (QD) 800.
Effector cells and co-culture
Ex vivo PBMC from immunized volunteers collected before and 42 days after immunization were used as effectors as previously described [48]. Briefly, PBMC were co-cultured with autologous B-LCL cells at an effector to stimulator cell ratio of 5:1 in the presence of mAbs to CD107a and CD107b (15 μl of each/1 x 106 cells in 500 μl of R10 medium). The CD107 a and b antibodies were used to measure degranulation, a mechanism essential for the killing of S. infected targets by the cytotoxic CD8+ cells [60]. PBMC cultured with uninfected target cells or Staphylococcus enterotoxin B (SEB) (10 μg/ml, Sigma) were used as negative and positive controls, respectively. After ~2 hours of stimulation, protein transport blockers, Monensin (1 μg/ml, Sigma) and brefeldin-A (BFA) (2 μg/ml, Sigma), were added to the co-culture. After overnight (16–18 hours) incubation, cells were harvested, stained with a dead-cell discriminator, yellow fluorescent viability dye (Yevid, Invitrogen)[61, 62], followed by surface staining with mAbs against surface antigens (CD3, CD4, CD8, CD14, and CD19) and fixation and permeabilization with Fix & Perm cell buffers (Invitrogen, Carlsbad, CA). Cells were then stained intracellularly for IFN-γ, TNF-α, IL-17A and CD69. Finally, cells were fixed and analyzed by flow cytometry on an LSR-II instrument (BD Biosciences). Data were analyzed with WinList 7.0 (Verity Software House, Topsham, ME). Lymphocytes were gated based on their scatter characteristics. Single lymphocytes were gated based on forward scatter height vs. forward scatter area. A “dump” channel was used to eliminate dead cells (Yevid+) as well as macrophages/monocytes (CD14+), B lymphocytes (CD19+) and targets (CD45+) from analysis. Additional gating on CD3, CD4, and CD8 was performed to identify cytokine-producing (IFN-γ, TNF-α and IL-17A) and CD107 expressing T cell subsets. Net responses were calculated by subtracting the number of positive events of the experimental (Salmonella-Hly proteins) from the negative control (Hly only). Functional responses were considered specific for S. Typhi if the differential in the number of positive and negative events between experimental (Salmonella-Hly proteins) and negative control (Hly only) cultures were significantly increased (P < 0.01) using Z-test. Volunteers were considered responders if the net responses from the PBMC collected 42 days after immunization were greater than 0.1% from the net responses of PBMC collected before immunization.
Statistical analysis
All statistical tests were performed using Prism software (version 7, GraphPad Software, La Jolla, CA). Comparisons between two groups were carried out by Student’s t tests. Correlation analysis was achieved by Pearson Product Moment Correlation tests. P values <0.05 were considered significant.
Supporting information
(TIF)
B-LCL cells were infected with E. coli expressing one (Hly/pmark or Hly/Salmonella protein) or two plasmids (Hly only or Hly and S. Typhi protein) at 1:30 MOI. S. Typhi proteins were FliC, GroEL and OmpC. Uninfected B-LCLs and B-LCLs infected with recombinant E. coli expressing only Hly antigen were used as controls. The percentage of the E. coli-antigen expressing B-LCLs were assessed by flow cytometry using anti-E. coli antibody as described in Methods. Shown are the average of 3 independent experiments. p values of <0.05 were considered statistically significant.
(TIF)
Acknowledgments
We are indebted to the volunteers who allowed us to perform this study. We also thank Mrs. Robin Barnes and the staff from the Recruiting Section of the Center for Vaccine Development for their help in collecting blood specimens and Dr. Haiyan Chen, and Mrs. Regina Harley and Catherine Storrer for excellent technical assistance.
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This work was supported, in part, by National Institute of Allergy and Infectious Diseases (NIAID), National Institutes of Health (NIH), Department of Health and Human Services (DHHS) federal research grants (https://www.niaid.nih.gov) R01 AI036525, U19 AI082655 (Cooperative Center for Human Immunology [CCHI]) and U19-AI109776 (Center of Excellence for Translational Research [CETR] to MS. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institute of Allergy and Infectious Diseases, the National Institutes of Health. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Footnotes
☯ These authors contributed equally to this work.
References
- 1.Mogasale V, Maskery B, Ochiai RL, Lee JS, Mogasale VV, Ramani E, et al. Burden of typhoid fever in low-income and middle-income countries: a systematic, literature-based update with risk-factor adjustment. The Lancet Global health. 2014;2(10):e570–80. Epub 2014/10/12. doi: 10.1016/S2214-109X(14)70301-8 . [DOI] [PubMed] [Google Scholar]
- 2.Mogasale V, Mogasale VV, Ramani E, Lee JS, Park JY, Lee KS, et al. Revisiting typhoid fever surveillance in low and middle income countries: lessons from systematic literature review of population-based longitudinal studies. BMC infectious diseases. 2016;16:35 Epub 2016/01/30. doi: 10.1186/s12879-016-1351-3 ; PubMed Central PMCID: PMCPMC4731936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Antillon M, Warren JL, Crawford FW, Weinberger DM, Kurum E, Pak GD, et al. The burden of typhoid fever in low- and middle-income countries: A meta-regression approach. PLoS neglected tropical diseases. 2017;11(2):e0005376 Epub 2017/02/28. doi: 10.1371/journal.pntd.0005376 ; PubMed Central PMCID: PMCPMC5344533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Crump JA. Updating and refining estimates of typhoid fever burden for public health action. The Lancet Global health. 2014;2(10):e551–3. Epub 2014/10/12. doi: 10.1016/S2214-109X(14)70306-7 ; PubMed Central PMCID: PMCPMC4404498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.DeRoeck D, Jodar L, Clemens J. Putting typhoid vaccination on the global health agenda. N Engl J Med. 2007;357(11):1069–71. Epub 2007/09/15. doi: 10.1056/NEJMp078144 . [DOI] [PubMed] [Google Scholar]
- 6.Connerton P, Wain J, Hien TT, Ali T, Parry C, Chinh NT, et al. Epidemic typhoid in vietnam: molecular typing of multiple-antibiotic-resistant Salmonella enterica serotype typhi from four outbreaks. J Clin Microbiol. 2000;38(2):895–7. Epub 2000/02/03. ; PubMed Central PMCID: PMC86238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Rowe B, Ward LR, Threlfall EJ. Multidrug-resistant Salmonella typhi: a worldwide epidemic. Clin Infect Dis. 1997;24 Suppl 1:S106–9. . [DOI] [PubMed] [Google Scholar]
- 8.Sztein MB, Salerno-Goncalves R, McArthur MA. Complex adaptive immunity to enteric fevers in humans: lessons learned and the path forward. Front Immunol. 2014;5:516 Epub 2014/11/12. doi: 10.3389/fimmu.2014.00516 ; PubMed Central PMCID: PMCPmc4209864. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Buckle GC, Walker CL, Black RE. Typhoid fever and paratyphoid fever: Systematic review to estimate global morbidity and mortality for 2010. Journal of global health. 2012;2(1):010401 Epub 2012/12/01. doi: 10.7189/jogh.02.010401 ; PubMed Central PMCID: PMCPmc3484760. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Levine MM. Typhoid fever vaccines In: Plotkin SA, Mortimer EA, editors. Vaccines. Philadelphia: W.B. Saunders Company; 1994. p. 597–633. [Google Scholar]
- 11.Sur D, Ochiai RL, Bhattacharya SK, Ganguly NK, Ali M, Manna B, et al. A cluster-randomized effectiveness trial of Vi typhoid vaccine in India. N Engl J Med. 2009;361(4):335–44. Epub 2009/07/25. doi: 10.1056/NEJMoa0807521 . [DOI] [PubMed] [Google Scholar]
- 12.Levine MM, Ferreccio C, Abrego P, Martin OS, Ortiz E, Cryz S. Duration of efficacy of Ty21a, attenuated Salmonella typhi live oral vaccine. Vaccine. 1999;17(Suppl 2):S22–7. [DOI] [PubMed] [Google Scholar]
- 13.Guzman CA, Borsutzky S, Griot-Wenk M, Metcalfe IC, Pearman J, Collioud A, et al. Vaccines against typhoid fever. Vaccine. 2006;24(18):3804–11. doi: 10.1016/j.vaccine.2005.07.111 . [DOI] [PubMed] [Google Scholar]
- 14.Levine MM, Ferreccio C, Black RE, Tacket CO, Germanier R. Progress in vaccines against typhoid fever. Rev Infect Dis. 1989;11 Suppl 3:S552–67. Epub 1989/05/01. . [DOI] [PubMed] [Google Scholar]
- 15.Ferreccio C, Levine MM, Rodriguez H, Contreras R. Comparative efficacy of two, three, or four doses of Ty21a live oral typhoid vaccine in enteric-coated capsules: a field trial in an endemic area. The Journal of Infectious Diseases. 1989;159(4):766–69. [DOI] [PubMed] [Google Scholar]
- 16.Chinnasami B, Mangayarkarasi V, Prema A, Sadasivam K, Davis M. Safety and immunogenicity of Salmonella Typhi Vi conjugate vaccine (Peda Typh) in children up to five years. International Journal of Scientific and Research Publications,. 2013;3(2):1–5. [Google Scholar]
- 17.Szu SC, Klugman KP, Hunt S. Re-examination of immune response and estimation of anti-Vi IgG protective threshold against typhoid fever-based on the efficacy trial of Vi conjugate in young children. Vaccine. 2014;32(20):2359–63. Epub 2014/03/19. doi: 10.1016/j.vaccine.2014.02.050 ; PubMed Central PMCID: PMCPmc4068246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.van Damme P, Kafeja F, Anemona A, Basile V, Hilbert AK, De Coster I, et al. Safety, immunogenicity and dose ranging of a new Vi-CRM(1)(9)(7) conjugate vaccine against typhoid fever: randomized clinical testing in healthy adults. PLoS One. 2011;6(9):e25398 Epub 2011/10/08. doi: 10.1371/journal.pone.0025398 ; PubMed Central PMCID: PMCPmc3184126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Lin FY, Ho VA, Khiem HB, Trach DD, Bay PV, Thanh TC, et al. The efficacy of a Salmonella typhi Vi conjugate vaccine in two-to-five-year-old children. N Engl J Med. 2001;344(17):1263–9. Epub 2001/04/26. doi: 10.1056/NEJM200104263441701 . [DOI] [PubMed] [Google Scholar]
- 20.Saha MR, Ramamurthy T, Dutta P, Mitra U. Emergence of Salmonella typhi Vi antigen-negative strains in an epidemic of multidrug-resistant typhoid fever cases in Calcutta, India. The National medical journal of India. 2000;13(3):164 Epub 2001/09/18. . [PubMed] [Google Scholar]
- 21.Baker S, Sarwar Y, Aziz H, Haque A, Ali A, Dougan G, et al. Detection of Vi-negative Salmonella enterica serovar typhi in the peripheral blood of patients with typhoid fever in the Faisalabad region of Pakistan. J Clin Microbiol. 2005;43(9):4418–25. Epub 2005/09/08. doi: 10.1128/JCM.43.9.4418-4425.2005 ; PubMed Central PMCID: PMCPmc1234127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Tennant SM, Levine MM. Live attenuated vaccines for invasive Salmonella infections. Vaccine. 2015;33 Suppl 3:C36–41. Epub 2015/04/23. doi: 10.1016/j.vaccine.2015.04.029 ; PubMed Central PMCID: PMCPmc4469493. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Salerno-Goncalves R, Sztein MB. Cell-mediated immunity and the challenges for vaccine development. Trends in microbiology. 2006;14(12):536–42. doi: 10.1016/j.tim.2006.10.004 . [DOI] [PubMed] [Google Scholar]
- 24.Lundin BS, Johansson C, Svennerholm AM. Oral immunization with a Salmonella enterica serovar typhi vaccine induces specific circulating mucosa-homing CD4(+) and CD8(+) T cells in humans. Infect Immun. 2002;70(10):5622–7. Epub 2002/09/14. doi: 10.1128/IAI.70.10.5622-5627.2002 ; PubMed Central PMCID: PMC128315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Sheikh A, Khanam F, Sayeed MA, Rahman T, Pacek M, Hu Y, et al. Interferon-gamma and proliferation responses to Salmonella enterica Serotype Typhi proteins in patients with S. Typhi Bacteremia in Dhaka, Bangladesh. PLoS neglected tropical diseases. 2011;5(6):e1193 Epub 2011/06/15. doi: 10.1371/journal.pntd.0001193 ; PubMed Central PMCID: PMCPmc3110156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Sztein MB. Cell-mediated immunity and antibody responses elicited by attenuated Salmonella enterica Serovar Typhi strains used as live oral vaccines in humans. Clin Infect Dis. 2007;45 Suppl 1:S15–9. doi: 10.1086/518140 . [DOI] [PubMed] [Google Scholar]
- 27.Salerno-Goncalves R, Fernandez-Vina M, Lewinsohn DM, Sztein MB. Identification of a human HLA-E-restricted CD8+ T cell subset in volunteers immunized with Salmonella enterica serovar Typhi strain Ty21a typhoid vaccine. J Immunol. 2004;173(9):5852–62. [DOI] [PubMed] [Google Scholar]
- 28.Salerno-Goncalves R, Pasetti MF, Sztein MB. Characterization of CD8(+) Effector T Cell Responses in Volunteers Immunized with Salmonella enterica Serovar Typhi Strain Ty21a Typhoid Vaccine. J Immunol. 2002;169(4):2196–203. [DOI] [PubMed] [Google Scholar]
- 29.Salerno-Goncalves R, Wahid R, Sztein MB. Immunization of volunteers with Salmonella enterica serovar Typhi strain Ty21a elicits the oligoclonal expansion of CD8+ T cells with predominant Vbeta repertoires. Infect Immun. 2005;73(6):3521–30.; PubMed Central PMCID: PMC1111837. doi: 10.1128/IAI.73.6.3521-3530.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Salerno-Goncalves R, Wahid R, Sztein MB. Ex Vivo kinetics of early and long-term multifunctional human leukocyte antigen E-specific CD8+ cells in volunteers immunized with the Ty21a typhoid vaccine. Clinical and vaccine immunology: CVI. 2010;17(9):1305–14. Epub 2010/07/28. CVI.00234-10 [pii] doi: 10.1128/CVI.00234-10 ; PubMed Central PMCID: PMC2944457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Sztein MB, Tanner MK, Polotsky Y, Orenstein JM, Levine MM. Cytotoxic T lymphocytes after oral immunization with attenuated vaccine strains of Salmonella typhi in humans. J Immunol. 1995;155(8):3987–93. [PubMed] [Google Scholar]
- 32.Wahid R, Salerno-Goncalves R, Tacket CO, Levine MM, Sztein MB. Generation of specific effector and memory T cells with gut- and secondary lymphoid tissue- homing potential by oral attenuated CVD 909 typhoid vaccine in humans. Mucosal Immunology. 2008;1(5):389–98. PubMed Central PMCID: PMC3215293. doi: 10.1038/mi.2008.30 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Viret JF, Favre D, Wegmuller B, Herzog C, Que JU, Cryz SJ Jr., et al. Mucosal and systemic immune responses in humans after primary and booster immunizations with orally administered invasive and noninvasive live attenuated bacteria. Infect Immun. 1999;67(7):3680–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Fresnay S, McArthur MA, Magder L, Darton TC, Jones C, Waddington CS, et al. Salmonella Typhi-specific multifunctional CD8+ T cells play a dominant role in protection from typhoid fever in humans. Journal of translational medicine. 2016;14(1):62 Epub 2016/03/02. doi: 10.1186/s12967-016-0819-7 ; PubMed Central PMCID: PMCPmc4772330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Bumann D. Identification of Protective Antigens for Vaccination against Systemic Salmonellosis. Front Immunol. 2014;5:381 Epub 2014/08/27. doi: 10.3389/fimmu.2014.00381 ; PubMed Central PMCID: PMCPmc4127814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Barat S, Willer Y, Rizos K, Claudi B, Maze A, Schemmer AK, et al. Immunity to intracellular Salmonella depends on surface-associated antigens. PLoS Pathog. 2012;8(10):e1002966 Epub 2012/10/25. doi: 10.1371/journal.ppat.1002966 ; PubMed Central PMCID: PMCPmc3475680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Bouwer HG, Alberti-Segui C, Montfort MJ, Berkowitz ND, Higgins DE. Directed antigen delivery as a vaccine strategy for an intracellular bacterial pathogen. Proc Natl Acad Sci U S A. 2006;103(13):5102–7. Epub 2006/03/22. doi: 10.1073/pnas.0509381103 ; PubMed Central PMCID: PMCPmc1458801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Hu PQ, Tuma-Warrino RJ, Bryan MA, Mitchell KG, Higgins DE, Watkins SC, et al. Escherichia coli expressing recombinant antigen and listeriolysin O stimulate class I-restricted CD8+ T cells following uptake by human APC. J Immunol. 2004;172(3):1595–601. Epub 2004/01/22. . [DOI] [PubMed] [Google Scholar]
- 39.Salerno-Goncalves R, Wyant TL, Pasetti MF, Fernandez-Vina M, Tacket CO, Levine MM, et al. Concomitant Induction of CD4(+) and CD8(+) T Cell Responses in Volunteers Immunized with Salmonella enterica Serovar Typhi Strain CVD 908-htrA. J Immunol. 2003;170(5):2734–41. [DOI] [PubMed] [Google Scholar]
- 40.Moreno-Eutimio MA, Tenorio-Calvo A, Pastelin-Palacios R, Perez-Shibayama C, Gil-Cruz C, Lopez-Santiago R, et al. Salmonella Typhi OmpS1 and OmpS2 porins are potent protective immunogens with adjuvant properties. Immunology. 2013;139(4):459–71. Epub 2013/02/26. doi: 10.1111/imm.12093 ; PubMed Central PMCID: PMCPmc3719063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Zhao W, Moest T, Zhao Y, Guilhon AA, Buffat C, Gorvel JP, et al. The Salmonella effector protein SifA plays a dual role in virulence. Scientific reports. 2015;5:12979 Epub 2015/08/14. doi: 10.1038/srep12979 ; PubMed Central PMCID: PMCPmc4534788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Figueira R, Watson KG, Holden DW, Helaine S. Identification of salmonella pathogenicity island-2 type III secretion system effectors involved in intramacrophage replication of S. enterica serovar typhimurium: implications for rational vaccine design. mBio. 2013;4(2):e00065 Epub 2013/04/18. doi: 10.1128/mBio.00065-13 ; PubMed Central PMCID: PMCPmc3634603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Salazar-Gonzalez RM, Maldonado-Bernal C, Ramirez-Cruz NE, Rios-Sarabia N, Beltran-Nava J, Castanon-Gonzalez J, et al. Induction of cellular immune response and anti-Salmonella enterica serovar typhi bactericidal antibodies in healthy volunteers by immunization with a vaccine candidate against typhoid fever. Immunol Lett. 2004;93(2–3):115–22. Epub 2004/05/26. doi: 10.1016/j.imlet.2004.01.010 . [DOI] [PubMed] [Google Scholar]
- 44.Carreno JM, Perez-Shibayama C, Gil-Cruz C, Lopez-Macias C, Vernazza P, Ludewig B, et al. Evolution of Salmonella Typhi outer membrane protein-specific T and B cell responses in humans following oral Ty21a vaccination: A randomized clinical trial. PLoS One. 2017;12(6):e0178669 Epub 2017/06/02. doi: 10.1371/journal.pone.0178669 ; PubMed Central PMCID: PMCPmc5453566. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Grubaugh D, Flechtner JB, Higgins DE. Proteins as T cell antigens: methods for high-throughput identification. Vaccine. 2013;31(37):3805–10. Epub 2013/06/29. doi: 10.1016/j.vaccine.2013.06.046 . [DOI] [PubMed] [Google Scholar]
- 46.Rueckert C, Guzman CA. Vaccines: from empirical development to rational design. PLoS Pathog. 2012;8(11):e1003001 Epub 2012/11/13. doi: 10.1371/journal.ppat.1003001 ; PubMed Central PMCID: PMCPmc3493475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Salerno-Goncalves R, Sztein MB. Priming of Salmonella enterica serovar Typhi-specific CD8(+) T cells by suicide dendritic cell cross-presentation in humans. PLoS ONE. 2009;4(6):e5879 doi: 10.1371/journal.pone.0005879 ; PubMed Central PMCID: PMC2691582. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Salerno-Goncalves R, Rezwan T, Sztein MB. B cells modulate mucosal associated invariant T cell immune responses. Front Immunol. 2014;4:511 Epub 2014/01/17. doi: 10.3389/fimmu.2013.00511 ; PubMed Central PMCID: PMCPmc3882667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.McArthur MA, Sztein MB. Heterogeneity of multifunctional IL-17A producing S. Typhi-specific CD8+ T cells in volunteers following Ty21a typhoid immunization. PLoS One. 2012;7(6):e38408 Epub 2012/06/09. doi: 10.1371/journal.pone.0038408 ; PubMed Central PMCID: PMC3367967. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Wahid R, Salerno-Goncalves R, Tacket CO, Levine MM, Sztein MB. Cell-mediated immune responses in humans after immunization with one or two doses of oral live attenuated typhoid vaccine CVD 909. Vaccine. 2007;25(8):1416–25. doi: 10.1016/j.vaccine.2006.10.040 ; PubMed Central PMCID: PMC1840048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Pasetti MF, Levine MM, Sztein MB. Animal models paving the way for clinical trials of attenuated Salmonella enterica serovar Typhi live oral vaccines and live vectors. Vaccine. 2003;21(5–6):401–18. [DOI] [PubMed] [Google Scholar]
- 52.de Jong HK, Parry CM, van der Poll T, Wiersinga WJ. Host-pathogen interaction in invasive Salmonellosis. PLoS pathogens. 2012;8(10):e1002933 doi: 10.1371/journal.ppat.1002933 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Mastroeni P, Villarreal-Ramos B, Hormaeche CE. Role of T cells, TNF alpha and IFN gamma in recall of immunity to oral challenge with virulent salmonellae in mice vaccinated with live attenuated aro- Salmonella vaccines. Microbial pathogenesis. 1992;13(6):477–91. Epub 1992/12/01. . [DOI] [PubMed] [Google Scholar]
- 54.McConnell SC, Hernandez KM, Wcisel DJ, Kettleborough RN, Stemple DL, Yoder JA, et al. Alternative haplotypes of antigen processing genes in zebrafish diverged early in vertebrate evolution. Proc Natl Acad Sci U S A. 2016;113(34):E5014–23. Epub 2016/08/06. doi: 10.1073/pnas.1607602113 ; PubMed Central PMCID: PMCPMC5003237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.International Conference on Harmonisation of Technical Requirements for Registration of Pharmaceuticals for Human Use (ICH) adopts Consolidated Guideline on Good Clinical Practice in the Conduct of Clinical Trials on Medicinal Products for Human Use. International digest of health legislation. 1997;48(2):231–4. Epub 1997/01/01. . [PubMed] [Google Scholar]
- 56.Levine MM, Ferreccio C, Black RE, Germanier R. Large-scale field trial of Ty21a live oral typhoid vaccine in enteric-coated capsule formulation. Lancet. 1987;1(8541):1049–52. . [DOI] [PubMed] [Google Scholar]
- 57.P S.N., B P, T G., V P. High-Throughput Gene Cloning Using the Gateway® Technology In: P P.F., editor. Functional Protein Microarrays in Drug Discovery: CRC Press; 2007. [Google Scholar]
- 58.Coren LV, Shatzer T, Ott DE. CD45 immunoaffinity depletion of vesicles from Jurkat T cells demonstrates that exosomes contain CD45: no evidence for a distinct exosome/HIV-1 budding pathway. Retrovirology. 2008;5:64 Epub 2008/07/18. doi: 10.1186/1742-4690-5-64 ; PubMed Central PMCID: PMC2490705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Kumar PD, Krishnaswamy S. Overexpression, refolding, and purification of the major immunodominant outer membrane porin OmpC from Salmonella typhi: characterization of refolded OmpC. Protein expression and purification. 2005;40(1):126–33. Epub 2005/02/22. doi: 10.1016/j.pep.2004.12.023 . [DOI] [PubMed] [Google Scholar]
- 60.Betts MR, Brenchley JM, Price DA, De Rosa SC, Douek DC, Roederer M, et al. Sensitive and viable identification of antigen-specific CD8+ T cells by a flow cytometric assay for degranulation. J Immunol Methods. 2003;281(1–2):65–78. . [DOI] [PubMed] [Google Scholar]
- 61.Lamoreaux L, Roederer M, Koup R. Intracellular cytokine optimization and standard operating procedure. Nature protocols. 2006;1(3):1507–16. Epub 2007/04/05. doi: 10.1038/nprot.2006.268 . [DOI] [PubMed] [Google Scholar]
- 62.Booth JS, Toapanta FR, Salerno-Goncalves R, Patil S, Kader HA, Safta AM, et al. Characterization and functional properties of gastric tissue-resident memory T cells from children, adults, and the elderly. Front Immunol. 2014;5:294 Epub 2014/07/06. doi: 10.3389/fimmu.2014.00294 ; PubMed Central PMCID: PMCPmc4062881. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(TIF)
B-LCL cells were infected with E. coli expressing one (Hly/pmark or Hly/Salmonella protein) or two plasmids (Hly only or Hly and S. Typhi protein) at 1:30 MOI. S. Typhi proteins were FliC, GroEL and OmpC. Uninfected B-LCLs and B-LCLs infected with recombinant E. coli expressing only Hly antigen were used as controls. The percentage of the E. coli-antigen expressing B-LCLs were assessed by flow cytometry using anti-E. coli antibody as described in Methods. Shown are the average of 3 independent experiments. p values of <0.05 were considered statistically significant.
(TIF)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.