

Abstract
The aryl hydrocarbon receptor (AhR) is a highly conserved transcription factor that mediates a broad spectrum of species-, strain-, sex-, age-, tissue-, and cell-specific responses elicited by structurally diverse ligands including 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD). Dose-dependent effects on liver-specific and sexually dimorphic gene expression were examined in male and female mice gavaged with TCDD every 4 days for 28 or 92 days. RNA-seq data revealed the coordinated repression of 181 genes predominately expressed in the liver including albumin (3.7-fold), α-fibrinogen (14.5-fold), and β-fibrinogen (17.4-fold) in males with corresponding AhR enrichment at 2 hr. Liver-specific genes exhibiting sexually dimorphic expression also demonstrated diminished divergence between sexes. For example, male-biased Gstp1 was repressed 3.0-fold in males and induced 4.5-fold in females, which were confirmed at the protein level. Disrupted regulation is consistent with impaired GHR-JAK2-STAT5 signaling and inhibition of female specific CUX2-mediated transcription as well as the repression of other key transcriptional regulators including Ghr, Stat5b, Bcl6, Hnf4a, Hnf6, Foxa1/2/3, and Zhx2. Attenuated liver-specific and sexually dimorphic gene expression was concurrent with the induction of fetal genes such as alpha-fetoprotein. The results suggest AhR activation causes the loss of liver-specific and sexually dimorphic gene expression producing a functionally “de-differentiated” hepatic phenotype.
Introduction
2,3,7,8-Tetrachlorodibenzo-p-dioxin (TCDD) is a persistent environmental contaminant that bioaccumulates and elicits a broad spectrum of biochemical and toxic effects. These effects are largely mediated by the aryl hydrocarbon receptor (AhR), a ligand-activated transcription factor [1]. Following ligand binding, cytosolic AhR dissociates from its chaperone proteins (e.g., HSP90, XAP2, and P23) and translocates to the nucleus where it dimerizes with the aryl hydrocarbon receptor nuclear translocator (ARNT). This complex interacts with dioxin response elements (5’-GCGTG-3’) to elicit differential gene expression. AhR-mediated gene expression changes also involve interactions with non-consensus sequences or tethering to other transcription factors [2, 3].
The liver is a primary target of TCDD-elicited pathologies including the induction of fatty liver disease [4, 5], impaired cellular proliferation [6], altered iron and bile acid homeostasis [7], reprogramming of carbohydrate and amino acid metabolism [8], coagulation cascade activation [9], and development of hepatocellular carcinoma (HCC) [10]. Moreover, AhR null mice show impaired liver development [11]. Although many phenotypic and underlying hepatic transcriptomic responses of AhR activation are conserved across sexes, differences in sensitivities between male and female mice are reported [12, 13]. These differences warrant further investigation as the role of sex in disease development and toxicity has not been adequately investigated, particularly in liver, one of the most sexually dimorphic organs [14].
Sexually dimorphic gene expression is closely associated with liver-specific gene regulation. It is largely mediated by the growth hormone (GH) signaling cascade (GHR-JAK2-STAT5) in mice, rats, and humans [15–18]. In males, the pulsatile secretion of GH by the pituitary is under gonadal steroid control while females exhibit more stable GH levels due to continuous secretion [19]. GH binding to the GH receptor (GHR), JAK2 activation, and STA5b phosphorylation results in intermittent or persistent STA5b activation and DNA binding [19]. The female-specific transcription factor, CUX2, and the expression of other hepatocyte nuclear factors (e.g., Hepatocyte Nuclear Factor 4 α;HNF4α, B-Cell CLL/Lymphoma 6; BCL6, Zinc Fingers and Homeoboxes 2; ZHX2) also influence GHR-JAK2-STAT5 signaling and serves an important role in determining sexually dimorphic xenobiotic metabolism gene expression [19, 20]. These same transcription factors are also implicated in liver development and hepatocyte differentiation [21].
AhR agonists impair GHR-JAK2-STAT5 signaling in mice and rats [22–25] and are reported to cause feminization of male mouse liver gene expression [26]. To further investigate AhR-mediated hepatotoxicity, the effects of TCDD on liver-specific and sexually dimorphic gene expression were examined using published hepatic RNA-seq datasets from male and female mice orally gavaged with 0–30 μg/kg TCDD every 4 days for 28 or 92 days (females only) [7, 27, 28]. Overall, TCDD repressed sexually-dimorphic gene expression in male and female livers yielding a more ambiguous phenotype (e.g. male and female hepatic gene expression converged to sexually ambiguous profiles). In addition, TCDD elicited the loss of liver-specific gene expression while inducing gene expression associated with liver progenitor cells producing a functionally “de-differentiated” hepatocyte phenotype. These changes are consistent with the disruption of GHR-JAK2-STAT5, CUX2, and HNF signaling. The cumulative loss of the associated functions may compromise liver function and contribute to TCDD-elicited hepatotoxicity.
Materials and methods
Animals
The animals, study design, and analysis of transcriptomic and ChIP-seq data have been previously described [29]. Briefly, postnatal day 25 (PND25) C57BL/6 mice were obtained from Charles River Laboratories (Portage, MI). Females were housed in polycarbonate cages with cellulose fiber chips (Aspen Chip Laboratory Bedding, Warrensberg, NY) and males in Innovive cages (Innovive inc., San Diego, CA) with ALPHA-dri (Shepherd Specialty Papers, Chicago, IL) bedding at 30–40% humidity, 23°C, and a 12h light/dark cycle were fed ad libitum (Harlan Teklad 22/5 Rodent Diet 8940, Madison, WI) with free access to water. Housing conditions between sexes differ due to changes within the animal care facilities at Michigan State University. No significant differences in TCDD-elicited responses due to these changes have been observed. On PND 28 and every following 4th day (d), animals were orally gavaged with 0.1 mL sesame oil or 0.01, 0.03, 0.1, 0.3, 1, 3, 10 and 30 μg/kg TCDD (Dow Chemical Company, Midland, MI) for a total of 28 or 92d (females only). Male mice exhibited greater sensitivity to TCDD and therefore were not treated for 92 days to avoid overt toxicity and death. Four days following the final dose mice were sacrificed by cervical dislocation. Liver samples were collected and immediately frozen in liquid nitrogen. Doses used compensate for the short study duration compared to lifelong cumulative human exposure from diverse AhR ligands, their bioaccumulative nature, and differences in metabolism and half-life. Furthermore, doses result in hepatic tissue levels that span human background serum concentrations reported in the United States, Germany, Spain, and the United Kingdom as well as serum levels in Viktor Yushchenko 4–39 months following intentional poisoning [30]. All procedures were approved by the All-University Committee on Animal Use and Care.
Transcriptomic, AhR ChIP-seq, and putative dioxin response element data
Transcriptomic, AhR ChIP-seq, and computationally identified putative dioxin response element (pDRE) data were previously published [7, 8, 28]. Genes were considered differentially expressed when |fold-change| ≥ 1.5 and posterior probability values (P1(t)) ≥ 0.8 determined using an empirical Bayes approach [28]. Significant AhR ChIP-seq binding used a FDR ≤ 0.05. pDREs were considered functional when the matrix similarity score (MSS) ≥ 0.85. Sequencing data is available in the Gene Expression Omnibus (GEO; GSE62902, GSE81990, GSE87519, GSE97634, GSE97636). Sexually dimorphic genes were identified using RNA-seq datasets from male and female mice from vehicle treatment groups from the studies described above (GSE62902 and GSE87519) to account for age and other experimental factors. Following read mapping to the mouse reference genome (GRCm38) and gene count determinations [28], counts were transformed through variance stabilizing transformation (VST) using DESeq in R [31]. Data was normalized using a semi-parametric approach in SAS v9.3. An empirical Bayes approach was used to calculate posterior probabilities (P1(t)) values. Genes were considered sexually dimorphic if there was a |fold-change| ≥ 2.0 and P1(t) ≥ 0.8 difference between male and female transcriptomic data sets (S1 Table). Identification of over-represented transcription factor binding sites for ChIP-seq peaks was performed using Pscan-ChIP [32].
Gene set enrichment and neighborhood gene expression regulation analyses
Gene set enrichment analysis (GSEA) [33] of sexually dimorphic and liver-specific genes was performed using a pre-ranked gene list based on the magnitude of the fold-change (largest induction to most repressed). The liver-specific gene set was identified using published microarray data representing 96 different tissues and cell types including adult male liver [34]. Tissue specificity was determined based on the difference of the gene in the tissue of interest and subtracting the highest signal in all other tissues (S1 Fig). Genes were considered liver-specific when the difference in microarray signal was ≥ 5,000 (S2 Table).
Genomic neighborhoods consisting of several nearby genes exhibiting similar differential expression, indicating putative shared regulation, was determined by calculating a running sum of log2 fold-changes. Firstly, the differentially expressed gene (|fold-change| ≥ 1.5) with smallest transcription start site (TSS) genomic coordinate was identified and that fold-change (in log2 space) was set as the initial score. The next differentially expressed gene based on TSS genomic coordinate was then found and that value (in log2 space) was added to the score. When a gene TSS was identified but did not exhibit a change in expression, the existing score was recorded. This process was continued across all genes in the chromosome, then all chromosomes. Scores were plotted based on the TSS coordinate and assessed for neighborhoods identified as dramatic increases (induction) or decreases (repression) revealing genome regions of closely located genes that may be co-regulated.
Histopathology
Paraffin embedded livers were sectioned at 4 μm, placed on slides coated with 2% 3-Aminopropyltriethoxysilane, and dried at 56°C overnight. Slides were deparaffinized in Xylene and hydrated through descending grades of ethyl alcohol to distilled water then placed in Tris Buffered Saline pH 7.4 (Scytek Labs, Logan, UT) for 5 minutes (min). Endogenous peroxidase was blocked using 3% Hydrogen Peroxide/Methanol bath for 30 min followed by running tap and distilled water rinses. Standard micro-polymer complex staining steps were performed at room temperature on the IntelliPath™ Flex Autostainer. All staining steps are followed by TBS Autowash buffer (Biocare Medical, Concord, CA) rinses. After blocking for non-specific protein with Background Punisher (Biocare) for 10 min, sections were incubated with Polyclonal Rabbit anti—AFP (Protein Tech, 14550-1-AP; 1:100) in normal antibody diluent (NAD-Scytek) for 60 min. Following 30 min in ProMark Rabbit anti–Rodent™ Micro-Polymer (Biocare), reaction development was performed with Romulin AEC™ (Biocare) for 5 minutes and counterstained with CAT Hematoxylin (Biocare). Histological processing and staining was performed at the Michigan State University Investigative Histopathology Lab (https://humanpathology.natsci.msu.edu/).
Protein determinations
Protein level were determined using a Wes capillary electrophoresis system (ProteinSimple, San Jose, CA) as previously described [8]. Liver samples were homogenized in radioimmunoprecipitation (RIPA) buffer supplemented with protease inhibitor cocktail. Total protein was quantitated by bicinchoninic acid assay. Equal amounts of protein were analyzed using antibodies for albumin (ALB; Abclonal A0353; 1:50), glutamate dehydrogenase 1 (GLUD1; Abclonal A7631; 1:100), glutathione S-transferase Pi 1 (GSTP1; Abclonal A5691; 1:50), hydroxyacid oxidase 1 (HAO1; Abclonal A6470; 1:50) and signal transducer and activator of transcription 5 (STAT5; Cell Signaling 9363; 1:50). Unlike traditional western blots, the Wes system quantitatively measures the chemiluminescence signal which were used to generate presented figures. Traces are provided as S2 Fig. Statistical analysis of protein levels in vehicle and treated mice were performed using the nonparametric Mann-Whitney’s U-test and considered significantly different when P ≤ 0.05.
Results
TCDD-mediated repression of liver-specific gene expression
To further explore the effect of TCDD on hepatic gene expression, 181 genes primarily expressed in the liver were identified using 96 GeneAtlas MOE430 datasets representing tissue/cell types from naïve male mice [34]. These 181 genes are subsequently referred to as “liver-specific” based on a microarray signal difference of ≥5,000 units when comparing constitutive expression in liver to other non-hepatic tissues/cell types. Signal difference was used instead of fold change to capture only highly expressed genes. Our liver-specific gene set included previously identified liver-specific genes including albumin (Alb), methionine adenosyltransferase (Mat1a), fatty acid binding protein 1 (Fabp1), and fetuin-A (Ahsg). GSEA revealed repression by TCDD in both male and female mice at 28 and 92d suggesting an overall loss of liver-specific expression in both sexes (Fig 1A–1C). Repressed genes included Alb, Mat1a, glutamate dehydrogenase 1 (Glud1), and hydroxyacid oxidase 1 (Hao1) in both sexes. Hepatic HAO1 protein levels were also repressed in males while hepatic ALB levels increased contrary to its transcriptional repression (Fig 2). Liver-specific genes demonstrating increased expression in males and females included Cyp1a2 and Ugt2b35, members of the AhR gene battery [35].
Fig 1. Gene expression changes of liver related genes.

Gene set enrichment analysis (GSEA) of 181 liver specific-genes in (A) male and (B) female mice gavaged with 30 μg/kg TCDD every 4 days for 28 days, or (C) female mice gavaged with 30 μg/kg TCDD every 4 days for 92 days. The 181 liver-specific genes were identified using published microarray datasets representing 96 tissues/cell types [34]. Identification of liver-specific genes is described in materials and methods. TCDD elicited gene expression changes were ranked from most induced (left—red) to most repressed (right). Vertical black line represents identified liver-specific genes. The top panel (green line) represents a running-sum statistic (enrichment score) based on the lower panel, increasing when a gene is a member of the liver-specific gene set and decreasing when it is not. Enrichment scores increased most dramatically on the right indicating most of the liver-specific genes were repressed by TCDD. (D) Heat map of liver-specific gene expression changes elicited by TCDD. (E) Heatmap of TCDD-elicited repression of hepatokines. For heatmaps (D and E) blue indicates repression while red represents induction. The presence of pDREs (MSS ≥ 0.856) and hepatic AhR enrichment peaks (FDR ≤ 0.05) at 2h are shown as green boxes. Read count represents the maximum raw number of aligned reads to each transcript where yellow represents a lower level of expression (≤ 500 reads) and pink represents a higher level of expression (≥ 10,000).
Fig 2. Hepatic protein levels changes by TCDD.
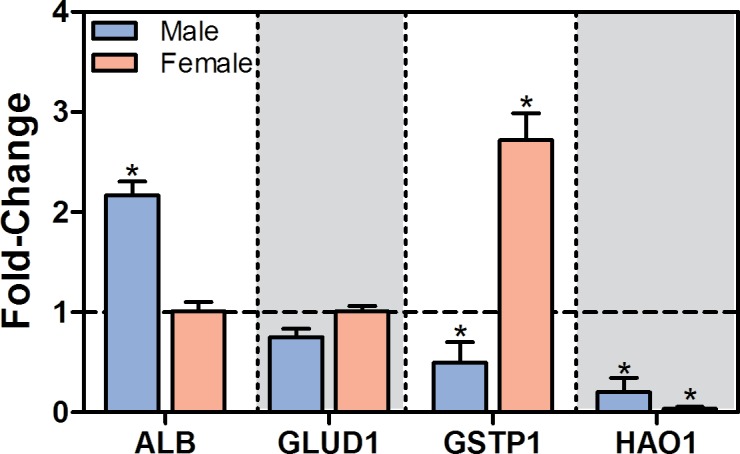
Hepatic protein levels in male and female mice gavaged with sesame oil vehicle or 30 μg/kg TCDD every 4 days for 28 days were measured using the ProteinSimple Wes system. Bars represent mean log2fold-change + SEM for at least 4 animals (N = 4–5). Asterisks (*) indicate a significant difference (P ≤ 0.05) compared to vehicle control determined by Mann-Whitney U-test.
At 30 μg/kg, repressed genes included hepatokines, and secreted liver proteins with autocrine or paracrine activity, (e.g., Lect2–12.8-fold; Igf1–8.0-fold; Ahsg -4.9-fold, Angptl6–2.9-fold; Sepp1–2.1-fold) in male mice (Fig 1E). Ahsg and Sepp1 were also among the 181 liver-specific genes and Lect2, Ahsg, and Sepp1 were repressed 3.1- (92d), 1.5- (92d), and 1.6-fold (28 and 92d), respectively, in females. Interestingly, changes in hepatokine expression have been associated with obesity, type II diabetes, and nonalcoholic fatty liver disease (NAFLD) in rodents and humans consistent with the development and progression of NAFLD by TCDD in mice [1, 4, 5, 36]. Conversely, Fgf21 which improves insulin resistance and reduces lipid accumulation [36] was induced 22.8- and 17.2-fold in male and female mice at 28d, respectively, while Serpina6 (corticosteroid binding globulin; CBG) exhibited an inverted “U”-shaped dose-response in both sexes with 3.4- and 2.1-fold induction in male and female mice, respectively. Fgf21 induction is a characterized AhR-mediated response [37]. Serpina6 shows strong AhR ChIP-seq enrichment upstream of the TSS in male and female mice while Angptl6 also had conserved AhR enrichment in both sexes (Fig 1E).
Induction of liver progenitor cell marker alpha-fetoprotein
The 1.6- and 15.0-fold induction of alpha-fetoprotein (Afp; 30 μg/kg) as well as 7.4- and 7.8-fold induction of TNF receptor superfamily 19 (Tnfrsf19; 30 μg/kg) in female and male mice, respectively, contrasts the overall repression of adult liver-specific gene expression. AFP is typically expressed during development in hepatic progenitor cells or hepatocellular carcinoma cells. Despite the lack of histological evidence of tumorigenesis, liver sections were evaluated for AFP expression. Increased cytosolic and intranuclear AFP staining was observed in male liver sections at 30 μg/kg TCDD including within hypertrophic differentiated hepatocytes showing lipid vacuolization, but not in female sections (Fig 3). The apparent difference in scale of histological images reflect the increased sensitivity of male mice to TCDD (Fig 3D). Expression of other albuminoids which play important roles in systemic transport such as albumin (Alb), afamin (Afm), and vitamin-D binding protein (Gc) were repressed 3.7-, 7.8-, and 4.3-fold, respectively, in male mice at 30 μg/kg. Repression of Alb, Afm and Gc was also observed in females following 92d of exposure, albeit to a lesser extent. Notably, Alb, Afp, Afm and Gc are sequentially co-localized on the same chromosome and arranged in the same transcriptional orientation in all mammals [38]. AhR enrichment was observed at 3 sites without pDREs ~ 13 – 30kb upstream of Alb in both male and female mice 2 h after TCDD treatment suggesting putative AhR-dependent co-regulation (Fig 4). The transcriptional repressor, zinc-finger and homeoboxes 2 (Zhx2), is also known to inhibit the fetal hepatic expression of Afp, H19, and Gpc3 [20]. Accordingly, 30 μg/kg TCDD repressed Zhx2 1.6-fold while fetal markers Afp, H19 and Gpc3 were induced 15.0-, 2.9- and 2.5-fold, respectively.
Fig 3. Immunohistochemical evaluation of hepatic AFP.

Representative photomicrographs for AFP stained liver sections of (A) sesame oil vehicle treated females, (B) sesame oil vehicle treated males, (C) 30 μg/kg TCDD treated females, and (D) 30 μg/kg TCDD treated males. Scale bar represents 50 μm. The portal vein is designated by the letter V, bile ducts with the letter b, and AFP positive stained regions by solid black arrows.
Fig 4. Albuminoid genomic region.

Albuminoid genomic region including Alb, Afp, and Afm. UCSC genome browser tracks show (1) the scale, (2) male AhR ChIP-seq peaks at 2 h, (3) male AhR enriched sites (FDR ≤ 0.05), (4) female AhR ChIP-seq peaks, (5) female AhR enriched sites (FDR ≤ 0.05), (6) location of pDREs (diagonal line indicates pDREs with a matrix similarity score ≥0.85), and (7) location of Alb, Afp and Afm genes within the albuminoid genomic region. Gc, the fourth albuminoid, is located 1 Mb upstream of Alb (not shown). Tracks are available for visualization at http://dbzach.fst.msu.edu/index.php/publications/supplementary-data/.
AhR enrichment at liver-specific genes
AhR enrichment was observed within 136 of the 181 liver-specific genes (79 in both sexes, 56 in males only, and 1 in females only; Fig 5A). pDREs were present in the majority of these binding sites (26 of 56 male sites, 57 of 79 male and female sites, and 0 of 1 female only sites), suggesting direct AhR-DRE-dependent regulation. Over-represented transcription factor motif analysis within AhR enriched regions identified binding sites for HNF4α, COUP-TFI and III, retinoic acid receptor (RXRA, RARG), CCAAT/Enhancer binding proteins (CEBPB, D, E, and G), and vitamin D receptor (VDR) (Fig 5B). In male mice, Cebpd, Rarg, and Vdr were induced 2.0-, 2.8-, and 2.1-fold, respectively, while Cebpe was repressed 4.1-fold at 30 μg/kg TCDD. Only Rarg was differentially expressed in females showing a 1.6-fold induction at 30 μg/kg TCDD. Also implicated in liver-specific gene expression were HNF6 (Onecut1) and HNF3 (Foxa1, Foxa2, and Foxa3) (repressed 5.6-, 1.7-, 1.6-, and 2.3-fold, respectively in males at 30 μg/kg) [39]. HNF6 and HNF3 repression is consistent with the 8.1-fold repression of transthyretin (Ttr), a male target gene. Smaller changes were observed in females after 92d but not at 28d. These results are consistent with proposed AhR-COUP-TFII-mediated repression [2].
Fig 5. AhR enrichment and motif analyses.

Comparison of AhR enrichment at liver-specific genes in male and female mice. (A) Values represent the number of unique genes in each segment of the Venn diagrams while numbers in parentheses show the number of AhR enriched regions containing a pDRE (matrix similarity score ≥ 0.85) for liver-specific genes. (B) Sequences beneath AhR enriched regions were queried for over-represented transcription factor binding motifs using Pscan-ChIP (Zambelli et al. 2013).
TCDD results in loss of sexually dimorphic gene expression
Liver-specific gene expression is also regulated by sexually dimorphic transcription factors [17, 19, 20, 26]. Accordingly, many liver-specific genes are also sexually dimorphic [14]. Hepatic transcription factors implicated in sexual dimorphism such as Onecut1, Stat5a, Stat5b, and Bcl6 were repressed 5.6-, 2.5-, 2.0, and 4.8-fold, respectively, by TCDD in males (Fig 6A and 6B). Cux2, a female-specific transcription factor exhibited dose-dependent repression (6.2-fold) in female mice at 92d. Analysis of male and female RNA-Seq data identified 442 male-enriched genes and 368 female-enriched genes using a |fold-change| ≥ 2.0 and P1(t) ≥ 0.8 criteria. Sulfotransferases (e.g. Sult2a1, 2a2, 2a5, 3a1) exhibit female-specific expression while Cyp4a12 and several mouse urinary proteins (e.g. Mup7, 11, 20, and 21) are primarily expressed in males [40, 41]. A total of 26 genes were classified as sexually dimorphic and liver-specific (S3 Fig). GSEA revealed TCDD repressed male-specific genes while female-specific genes were induced in male mice, and vice-versa in females (Fig 7). For example, TCDD divergently regulated Gstp1 (induced up to 4.5-fold in females; repressed down to 3.0-fold in males) and Cyp2d9 (induced up to 9.8-fold in females; repressed down to 5.8-fold in males). GSTP1 dimorphic expression was confirmed at the protein level (Fig 2). In contrast, highly expressed (>10,000 read counts) major urinary protein genes were repressed in both male (Mup1, 3, 10, 11, 14, 17 and 20 were repressed 167-, 1,538-, 100-, 270-, 179- and 90.9-fold, respectively, at 30 μg/kg) and female (Mup3, 11 and 17 repressed 25.0-, 9.1 and 2.2-fold, respectively, at 30 μg/kg) livers (Fig 8). Female Mup1, 10, 14, and 20 are expressed at lower levels (<1700 counts) and were largely unaffected by TCDD. Mups are located on chromosome 4 between 59,956,806 and 62,150,841 where 44 AhR enrichment peaks may be involved in their coordinated repression. The 1.6-fold inhibition of Zhx2 may also contribute to Mup repression [20, 42].
Fig 6. AhR-mediated changes in sexually dimorphic regulator expression.

(A) GHR-JAK2-STAT5 signaling cascade interactions with liver-specific gene expression regulators [16, 18, 19, 43]. GH-mediated activation of GHR induces STAT5 phosphorylation promoting male-specific gene expression, and induction of female-specific CUX2 (red outline) which regulates female-specific gene expression. CUX2 and STAT5 compete for binding sites resulting in sexually dimorphic gene expression. BCL6, HNF4α, HNF6, and FOXA also regulate sexually dimorphic and liver-specific gene regulation. Blue identifies repressed genes while white represents genes unaffected by TCDD. (B) Heatmap of sexually dimorphic and liver-specific gene expression regulators (A) in males (28d) and females (28 and 92d). Genes in blue were repressed while red indicates induction. STAT5 protein levels were determined in (C) female and (D) male mice gavaged with 30 μg/kg TCDD every 4 days for 28 days. Bars represent mean ± SEM for at least 3 animals (N = 3–4). Asterisks (*) indicate a significant difference compared to vehicle control as determined by Mann-Whitney U-test.
Fig 7. Gene expression changes of sexually dimorphic genes.

Gene set enrichment analysis (GSEA) of sexually dimorphic genes in (A-B) male and (B-C) female mice gavaged with TCDD every 4 days for 28 days. GSEA was performed using male-enriched and female-enriched gene sets determined as described in materials and methods. Gene expression changes were ranked from largest positive to largest negative. The top panel (green line) represents a ‘running-sum statistic’ which increases when a gene is in a gene set (denoted by black bars), and decreasing when it is not. TCDD repressed the majority of male-specific dimorphic genes (A) while inducing many female-specific genes (B) in male mice. Conversely, TCDD induced many male-specific genes (C) while repressing a majority of the female-specific genes (D) in female mice. (E) Heat map demonstrates the gene expression changes in male-specific and female-specific genes at 28 days.
Fig 8. Coordinated repression of Mup genes on chromosome 4.

(A) Running fold-change for chromosome 4 was calculated for male mice gavaged with TCDD every 4 days for 28 days. Points are colored based on maximum read counts with yellow representing lower expression (≤ 500 reads) and pink representing higher expression (≥ 10,000). The presence of AhR ChIP-Seq peaks at 2h in males is indicated by light grey vertical bars behind points. (B) A region demonstrating a dramatic decrease in the Running fold-change was magnified identifying a chromosomal region with multiple genes repressed by TCDD. (C) Genes in the magnified region are show as heat maps.
Sexually dimorphic gene expression is mediated, in part, by GH, which is secreted intermittently in males and continuously in females. GH signaling is disrupted by AhR-mediated repression of GHR-JAK2-STAT5a/b signaling [19, 22, 25]. In our study, Ghr was repressed 8.3- and 2.4-fold in male and female mice at 30 μg/kg, respectively. Corresponding decreases in STAT5 protein were confirmed (Fig 6C and 6D). Interestingly, HNF4α and STAT5b also regulate a shared set of sexually dimorphic genes [43] including male-specific Gstp1, Cyp2d9, Cyp7b1, Mup4, Cyp4a12, Mup1, Hsd3b5, Elovl3, and Slco1a1 which were repressed (3.0- to 1,186-fold) in both sexes, while Cyp2a4, Sult1e1, and Cyp2b9 were induced (865.4-, 83.8-, and 28.5-fold, respectively). Further highlighting the convergence of sexually dimorphic adult liver gene expression is the coordinated repression of male-enriched adult hemoglobin subunits Hba-a1, Hba-a2, Hbb-bs, and Hbb-bt in males (4.5-, 3.7-, 4.5- and 6.3-fold, respectively), and their induction in females (1.7-, 1.7-, 1.8-, and 2.0-fold, respectively) at 28d. TCDD has been reported to reduce hemoglobin levels in male mice [5].
Discussion
TCDD induces a broad spectrum of hepatic pathologies from hepatic fat accumulation to HCC in a species-, strain-, sex-, age-, tissue- and cell-specific manner [4–10, 26]. Much remains to be understood about the role of sex in liver disease and toxicity [14]. Consequently, we investigated the effect of TCDD on liver-specific and sexually dimorphic gene expression using hepatic RNA-seq datasets from male and female mice [7, 8, 28]. Overall, gene expression changes elicited by TCDD indicate a loss of liver- and sex-specific gene expression producing a sexually ambiguous and functionally de-differentiation liver.
The dose-dependent repression of liver-specific gene expression suggests diminution of the functional hepatic phenotype. Furthermore, AFP induction suggests the TCDD treated liver is retreating to a less differentiated transcriptome consistent with increases in hepatic and serum AFP levels typically associated with HCC or liver regeneration [44]. AFP induction in differentiated hepatocytes marked by vacuolization suggests TCDD caused a loss in hepatocyte differentiation as opposed to progenitor cell proliferation in response to liver regeneration needs. Moreover, AFP staining in liver cancer is only reported to be cytoplasmic [44] while both intranuclear and cytoplasmic staining was observed with TCDD in males at 30 μg/kg, the significance of which is unclear. Paradoxically, Alb, which is typically reciprocally regulated compared to Afp, was repressed as expected, yet hepatic protein levels were increased. Interestingly, plasma ALB levels are lower in TCDD exposed waste incineration workers [45] and PCB exposed transformer repair workers [46].
Further evidence of diminished liver function includes the repression of albuminoids, hemoglobin subunits, major urinary proteins, and hepatokines [36, 47]. In addition to participating in the transport of vitamins, lipids, amino acids, steroids and metal ions, albuminoids also bind heme reducing lipoprotein oxidation and oxidative stress [7, 47]. Alb repression along with hemoglobin Hba and Hbb subunits is also consistent with TCDD elicited decreases in serum proteins [5]. Similarly, fetuin-A (Ashg) and leukocyte derived chemotaxin 2 (Lect2) repression enhances insulin signaling [36] and may support improved glucose tolerance in the absence of altered insulin levels in TCDD treated animals [27]. The physiological significance of Mup repression is unclear as hepatic secretion is largely male specific and implicated in lipid and pheromone binding. Although Mups also regulate glucose and lipid metabolism by repressing gluconeogenic and lipogenic genes [48], they are not expressed in humans.
There was also concomitant loss of sexually dimorphic gene expression consistent with reports of male liver “feminization” and divergent regulation of sexually dimorphic genes [12, 13, 26]. Our comparative study indicates loss of sexually dimorphic expression as opposed to feminization or masculinization. This is likely due to the combined loss of both male gene repression and female gene induction in females, and vice-versa in males. Notably, the number of differentially expressed genes in TCDD treated males (9,314) was much larger than in females (3,766) [7, 12, 13]. This large difference in the number of dysregulated genes between sexes was also reported for HNF4α null mice which resulted in ~1,000 fewer DEGs in female mice [43]. HNF4α regulates liver-specific and GHR-JAK2-STAT5-mediated sexually dimorphic gene expression, the latter of which is impaired by AhR activation through repression of Ghr and Jak2 [17, 19, 22, 26]. Moreover, mouse models lacking hepatic GHR, JAK2, or STAT5 have increased liver damage and develop features of NAFLD [49] suggesting GHR-JAK2-STAT5 impairment contributes to TCDD-elicited NAFLD. The anti-estrogenic effects of AhR agonists in mediating these changes remains unclear [50, 51].
The repression of the negative regulator ZHX2 may also be involved in the loss of liver-specific and sexually dimorphic gene expression [20, 52, 53]. ZHX2 inhibits fetal gene expression in adult liver, namely Afp, H19, and Gpc3, as well as several sex-specific cytochrome P450s [20, 21, 53, 54], all of which were induced by TCDD. Similarly, ZHX2 is required for high expression of Mups [42] most of which were repressed by TCDD. Overall, ZHX2 repression is consistent with the loss of tissue- and sex-specific gene expression and the induction of fetal genes commonly associated with HCC.
Collectively, our studies suggest that TCDD elicits a loss of liver identity by diminishing both tissue-specific and sexually dimorphic gene expression. While the exact mechanism remains to be elucidated, impairment of GHR-JAK2-STAT5 signaling, dysregulation of HNF-mediated transcription, and Zhx2 repression are implicated. These effects may also contribute to the development of NAFLD pathologies and other AhR-mediated toxicities. Indeed, the repression of HNF6 (Onecut1) which plays a regulatory role in sexually dimorphic and liver-specific gene expression, also regulates hepatic lipid metabolism via induction of the transcriptional repressor REV-ERBα (Nr1d1) [55] which was induced by TCDD [7]. In summary, persistent AhR activation promoted the loss of unique hepatic functions and sexual dimorphism that may factor in TCDD-elicited NAFLD and HCC.
Supporting information
(XLSX)
(XLSX)
Microarray datasets for basal gene expression for 96 different male mouse tissues/cell types were obtained from Lattin et al. [34]. The difference between the microarray signal in the liver and the maximum fluorescent signal on a per gene basis in all other tissues/cells was calculated (ΔSignal). A gene was considered liver-specific when ΔSignal ≥ 5,000 units.
(DOCX)
Data was collected and analyzed as described in materials & methods.
(DOCX)
(DOCX)
Acknowledgments
This work was supported by the National Institute of Environmental Health Sciences Superfund Research Program (NIEHS SRP P42ES04911). TRZ is supported by AgBioResearch at MSU. RN was supported by the MSU Barnett Rosenberg Endowed Assistantship and Integrative Training in the Pharmacological Sciences grant (NIH 5T32GM092715). KAF is supported by the Canadian Institutes of Health Research (CIHR) Doctoral Foreign Study Award (DFS-140386).
Data Availability
All relevant data are within the paper and its Supporting Information files or available on GEO (https://www.ncbi.nlm.nih.gov/geo/) using accession numbers GSE62902, GSE81990, GSE87519, GSE97634, and GSE97636.
Funding Statement
This work was supported by the National Institute of Environmental Health Sciences Superfund Research Program (NIEHS SRP P42ES04911). TRZ is supported by AgBioResearch at MSU. RN was supported by the MSU Barnett Rosenberg Endowed Assistantship and Integrative Training in the Pharmacological Sciences grant (NIH 5T32GM092715). KAF is supported by the Canadian Institutes of Health Research (CIHR) Doctoral Foreign Study Award (DFS-140386).
References
- 1.Fernandez-Salguero PM, Hilbert DM, Rudikoff S, Ward JM, Gonzalez FJ. Aryl-hydrocarbon receptor-deficient mice are resistant to 2,3,7,8-tetrachlorodibenzo-p-dioxin-induced toxicity. Toxicol Appl Pharmacol. 1996;140(1):173–9. Epub 1996/09/01. doi: 10.1006/taap.1996.0210 . [DOI] [PubMed] [Google Scholar]
- 2.Dere E, Lo R, Celius T, Matthews J, Zacharewski TR. Integration of genome-wide computation DRE search, AhR ChIP-chip and gene expression analyses of TCDD-elicited responses in the mouse liver. BMC genomics. 2011;12:365 Epub 2011/07/19. doi: 10.1186/1471-2164-12-365 ; PubMed Central PMCID: PMC3160422. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Huang G, Elferink CJ. A novel nonconsensus xenobiotic response element capable of mediating aryl hydrocarbon receptor-dependent gene expression. Molecular pharmacology. 2012;81(3):338–47. Epub 2011/11/25. doi: 10.1124/mol.111.075952 ; PubMed Central PMCID: PMC3286297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Boverhof DR, Burgoon LD, Tashiro C, Chittim B, Harkema JR, Jump DB, et al. Temporal and dose-dependent hepatic gene expression patterns in mice provide new insights into TCDD-Mediated hepatotoxicity. Toxicol Sci. 2005;85(2):1048–63. Epub 2005/04/01. doi: 10.1093/toxsci/kfi162 . [DOI] [PubMed] [Google Scholar]
- 5.Vos JG, Moore JA, Zinkl JG. Toxicity of 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) in C57B1/6 mice. Toxicol Appl Pharmacol. 1974;29(2):229–41. Epub 1974/08/01. . [DOI] [PubMed] [Google Scholar]
- 6.Puga A, Xia Y, Elferink C. Role of the aryl hydrocarbon receptor in cell cycle regulation. Chem Biol Interact. 2002;141(1–2):117–30. Epub 2002/09/06. . [DOI] [PubMed] [Google Scholar]
- 7.Fader KA, Nault R, Kirby MP, Markous G, Matthews J, Zacharewski TR. Convergence of hepcidin deficiency, systemic iron overloading, heme accumulation, and REV-ERBalpha/beta activation in aryl hydrocarbon receptor-elicited hepatotoxicity. Toxicol Appl Pharmacol. 2017;321:1–17. Epub 2017/02/19. doi: 10.1016/j.taap.2017.02.006 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Nault R, Fader KA, Kirby MP, Ahmed S, Matthews J, Jones AD, et al. Pyruvate Kinase Isoform Switching and Hepatic Metabolic Reprogramming by the Environmental Contaminant 2,3,7,8-Tetrachlorodibenzo-p-Dioxin. Toxicol Sci. 2016;149(2):358–71. Epub 2015/11/20. doi: 10.1093/toxsci/kfv245 ; PubMed Central PMCID: PMC4900217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Nault R, Fader KA, Kopec AK, Harkema JR, Zacharewski TR, Luyendyk JP. From the Cover: Coagulation-Driven Hepatic Fibrosis Requires Protease Activated Receptor-1 (PAR-1) in a Mouse Model of TCDD-Elicited Steatohepatitis. Toxicol Sci. 2016;154(2):381–91. Epub 2016/09/11. doi: 10.1093/toxsci/kfw175 ; PubMed Central PMCID: PMC5139072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Kociba RJ, Keyes DG, Beyer JE, Carreon RM, Gehring PJ. Long-term toxicologic studies of 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) in laboratory animals. Annals of the New York Academy of Sciences. 1979;320:397–404. Epub 1979/05/31. . [DOI] [PubMed] [Google Scholar]
- 11.Bunger MK, Glover E, Moran SM, Walisser JA, Lahvis GP, Hsu EL, et al. Abnormal liver development and resistance to 2,3,7,8-tetrachlorodibenzo-p-dioxin toxicity in mice carrying a mutation in the DNA-binding domain of the aryl hydrocarbon receptor. Toxicol Sci. 2008;106(1):83–92. Epub 2008/07/29. doi: 10.1093/toxsci/kfn149 ; PubMed Central PMCID: PMC2563146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Lee J, Prokopec SD, Watson JD, Sun RX, Pohjanvirta R, Boutros PC. Male and female mice show significant differences in hepatic transcriptomic response to 2,3,7,8-tetrachlorodibenzo-p-dioxin. BMC genomics. 2015;16:625 Epub 2015/08/21. doi: 10.1186/s12864-015-1840-6 ; PubMed Central PMCID: PMC4546048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Prokopec SD, Watson JD, Lee J, Pohjanvirta R, Boutros PC. Sex-related differences in murine hepatic transcriptional and proteomic responses to TCDD. Toxicol Appl Pharmacol. 2015;284(2):188–96. Epub 2015/02/24. doi: 10.1016/j.taap.2015.02.012 . [DOI] [PubMed] [Google Scholar]
- 14.Della Torre S, Maggi A. Sex Differences: A Resultant of an Evolutionary Pressure? Cell metabolism. 2017;25(3):499–505. Epub 2017/02/14. doi: 10.1016/j.cmet.2017.01.006 . [DOI] [PubMed] [Google Scholar]
- 15.Yang X, Schadt EE, Wang S, Wang H, Arnold AP, Ingram-Drake L, et al. Tissue-specific expression and regulation of sexually dimorphic genes in mice. Genome research. 2006;16(8):995–1004. Epub 2006/07/11. doi: 10.1101/gr.5217506 ; PubMed Central PMCID: PMC1524872. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Zhang Y, Laz EV, Waxman DJ. Dynamic, sex-differential STAT5 and BCL6 binding to sex-biased, growth hormone-regulated genes in adult mouse liver. Molecular and cellular biology. 2012;32(4):880–96. Epub 2011/12/14. doi: 10.1128/MCB.06312-11 ; PubMed Central PMCID: PMC3272977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Oshida K, Vasani N, Waxman DJ, Corton JC. Disruption of STAT5b-Regulated Sexual Dimorphism of the Liver Transcriptome by Diverse Factors Is a Common Event. PloS one. 2016;11(3):e0148308 Epub 2016/03/10. doi: 10.1371/journal.pone.0148308 ; PubMed Central PMCID: PMC4784905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Waxman DJ, O'Connor C. Growth hormone regulation of sex-dependent liver gene expression. Mol Endocrinol. 2006;20(11):2613–29. Epub 2006/03/18. doi: 10.1210/me.2006-0007 . [DOI] [PubMed] [Google Scholar]
- 19.Conforto TL, Zhang Y, Sherman J, Waxman DJ. Impact of CUX2 on the female mouse liver transcriptome: activation of female-biased genes and repression of male-biased genes. Molecular and cellular biology. 2012;32(22):4611–27. Epub 2012/09/12. doi: 10.1128/MCB.00886-12 ; PubMed Central PMCID: PMC3486175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Creasy KT, Jiang J, Ren H, Peterson ML, Spear BT. Zinc Fingers and Homeoboxes 2 (Zhx2) Regulates Sexually Dimorphic Cyp Gene Expression in the Adult Mouse Liver. Gene expression. 2016;17(1):7–17. Epub 2016/05/20. doi: 10.3727/105221616X691712 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Si-Tayeb K, Lemaigre FP, Duncan SA. Organogenesis and development of the liver. Developmental cell. 2010;18(2):175–89. Epub 2010/02/18. doi: 10.1016/j.devcel.2010.01.011 . [DOI] [PubMed] [Google Scholar]
- 22.Nukaya M, Takahashi Y, Gonzalez FJ, Kamataki T. Aryl hydrocarbon receptor-mediated suppression of GH receptor and Janus kinase 2 expression in mice. FEBS letters. 2004;558(1–3):96–100. Epub 2004/02/05. doi: 10.1016/S0014-5793(03)01528-X . [DOI] [PubMed] [Google Scholar]
- 23.Timsit YE, Riddick DS. Stimulation of hepatic signal transducer and activator of transcription 5b by GH is not altered by 3-methylcholanthrene. Endocrinology. 2002;143(9):3284–94. Epub 2002/08/24. doi: 10.1210/en.2002-220212 . [DOI] [PubMed] [Google Scholar]
- 24.Timsit YE, Riddick DS. Interference with growth hormone stimulation of hepatic cytochrome P4502C11 expression in hypophysectomized male rats by 3-methylcholanthrene. Toxicol Appl Pharmacol. 2000;163(2):105–14. Epub 2000/03/04. doi: 10.1006/taap.1999.8862 . [DOI] [PubMed] [Google Scholar]
- 25.Lee C, Riddick DS. Aryl hydrocarbon receptor-dependence of dioxin's effects on constitutive mouse hepatic cytochromes P450 and growth hormone signaling components. Canadian journal of physiology and pharmacology. 2012;90(10):1354–63. Epub 2012/09/18. doi: 10.1139/y2012-099 ; PubMed Central PMCID: PMC3515668. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Oshida K, Waxman DJ, Corton JC. Chemical and Hormonal Effects on STAT5b-Dependent Sexual Dimorphism of the Liver Transcriptome. PloS one. 2016;11(3):e0150284 Epub 2016/03/10. doi: 10.1371/journal.pone.0150284 ; PubMed Central PMCID: PMC4784907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Nault R, Fader KA, Ammendolia DA, Dornbos P, Potter D, Sharratt B, et al. Dose-Dependent Metabolic Reprogramming and Differential Gene Expression in TCDD-Elicited Hepatic Fibrosis. Toxicol Sci. 2016;154(2):253–66. Epub 2016/08/27. doi: 10.1093/toxsci/kfw163 ; PubMed Central PMCID: PMC5139066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Nault R, Fader KA, Zacharewski T. RNA-Seq versus oligonucleotide array assessment of dose-dependent TCDD-elicited hepatic gene expression in mice. BMC genomics. 2015;16(1):373 Epub 2015/05/11. doi: 10.1186/s12864-015-1527-z ; PubMed Central PMCID: PMC4456707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Nault R, Fader KA, Lydic TA, Zacharewski TR. Lipidomic Evaluation of Aryl Hydrocarbon Receptor-Mediated Hepatic Steatosis in Male and Female Mice Elicited by 2,3,7,8-Tetrachlorodibenzo-p-dioxin. Chemical research in toxicology. 2017;30(4):1060–75. Epub 2017/02/28. doi: 10.1021/acs.chemrestox.6b00430 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Nault R, Fader KA, Ammendolia DA, Dornbos P, Potter D, Sharratt B, et al. Dose-dependent metabolic reprogramming and differential gene expression in TCDD-elicited hepatic fibrosis. Toxicol Sci. 2016. Epub 2016/08/27. doi: 10.1093/toxsci/kfw163 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Anders S, Huber W. Differential expression analysis for sequence count data. Genome biology. 2010;11(10):R106 Epub 2010/10/29. doi: 10.1186/gb-2010-11-10-r106 ; PubMed Central PMCID: PMC3218662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Zambelli F, Pesole G, Pavesi G. PscanChIP: Finding over-represented transcription factor-binding site motifs and their correlations in sequences from ChIP-Seq experiments. Nucleic acids research. 2013;41(Web Server issue):W535–43. Epub 2013/06/12. doi: 10.1093/nar/gkt448 ; PubMed Central PMCID: PMC3692095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Subramanian A, Tamayo P, Mootha VK, Mukherjee S, Ebert BL, Gillette MA, et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc Natl Acad Sci U S A. 2005;102(43):15545–50. Epub 2005/10/04. doi: 10.1073/pnas.0506580102 ; PubMed Central PMCID: PMC1239896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Lattin JE, Schroder K, Su AI, Walker JR, Zhang J, Wiltshire T, et al. Expression analysis of G Protein-Coupled Receptors in mouse macrophages. Immunome research. 2008;4:5 Epub 2008/04/30. doi: 10.1186/1745-7580-4-5 ; PubMed Central PMCID: PMC2394514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Yeager RL, Reisman SA, Aleksunes LM, Klaassen CD. Introducing the "TCDD-inducible AhR-Nrf2 gene battery". Toxicol Sci. 2009;111(2):238–46. Epub 2009/05/29. doi: 10.1093/toxsci/kfp115 ; PubMed Central PMCID: PMC2742581. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Lebensztejn DM, Flisiak-Jackiewicz M, Bialokoz-Kalinowska I, Bobrus-Chociej A, Kowalska I. Hepatokines and non-alcoholic fatty liver disease. Acta biochimica Polonica. 2016;63(3):459–67. Epub 2016/06/06. doi: 10.18388/abp.2016_1252 . [DOI] [PubMed] [Google Scholar]
- 37.Cheng X, Vispute SG, Liu J, Cheng C, Kharitonenkov A, Klaassen CD. Fibroblast growth factor (Fgf) 21 is a novel target gene of the aryl hydrocarbon receptor (AhR). Toxicol Appl Pharmacol. 2014;278(1):65–71. Epub 2014/04/29. doi: 10.1016/j.taap.2014.04.013 ; PubMed Central PMCID: PMC4090247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Jin L, Long L, Green MA, Spear BT. The alpha-fetoprotein enhancer region activates the albumin and alpha-fetoprotein promoters during liver development. Developmental biology. 2009;336(2):294–300. Epub 2009/09/29. doi: 10.1016/j.ydbio.2009.09.026 ; PubMed Central PMCID: PMC2783961. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Samadani U, Costa RH. The transcriptional activator hepatocyte nuclear factor 6 regulates liver gene expression. Molecular and cellular biology. 1996;16(11):6273–84. Epub 1996/11/01. ; PubMed Central PMCID: PMC231630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Norstedt G, Palmiter R. Secretory rhythm of growth hormone regulates sexual differentiation of mouse liver. Cell. 1984;36(4):805–12. Epub 1984/04/01. . [DOI] [PubMed] [Google Scholar]
- 41.Hurst JL, Beynon RJ, Armstrong SD, Davidson AJ, Roberts SA, Gomez-Baena G, et al. Molecular heterogeneity in major urinary proteins of Mus musculus subspecies: potential candidates involved in speciation. Scientific reports. 2017;7:44992 Epub 2017/03/25. doi: 10.1038/srep44992 ; PubMed Central PMCID: PMC5364487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Jiang J, Creasy KT, Purnell J, Peterson ML, Spear BT. Zhx2 (zinc fingers and homeoboxes 2) regulates major urinary protein gene expression in the mouse liver. J Biol Chem. 2017;292(16):6765–74. Epub 2017/03/05. doi: 10.1074/jbc.M116.768275 ; PubMed Central PMCID: PMC5399123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Holloway MG, Miles GD, Dombkowski AA, Waxman DJ. Liver-specific hepatocyte nuclear factor-4alpha deficiency: greater impact on gene expression in male than in female mouse liver. Mol Endocrinol. 2008;22(5):1274–86. Epub 2008/02/16. doi: 10.1210/me.2007-0564 ; PubMed Central PMCID: PMC2366185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Mu X, Espanol-Suner R, Mederacke I, Affo S, Manco R, Sempoux C, et al. Hepatocellular carcinoma originates from hepatocytes and not from the progenitor/biliary compartment. The Journal of clinical investigation. 2015;125(10):3891–903. Epub 2015/09/09. doi: 10.1172/JCI77995 ; PubMed Central PMCID: PMC4607132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Kang MJ, Lee DY, Joo WA, Kim CW. Plasma protein level changes in waste incineration workers exposed to 2,3,7,8-tetrachlorodibenzo-p-dioxin. Journal of proteome research. 2005;4(4):1248–55. Epub 2005/08/09. doi: 10.1021/pr049756d . [DOI] [PubMed] [Google Scholar]
- 46.Emmett EA. Polychlorinated biphenyl exposure and effects in transformer repair workers. Environmental health perspectives. 1985;60:185–92. Epub 1985/05/01. ; PubMed Central PMCID: PMC1568555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Fasano M, Fanali G, Leboffe L, Ascenzi P. Heme binding to albuminoid proteins is the result of recent evolution. IUBMB life. 2007;59(7):436–40. Epub 2007/07/27. doi: 10.1080/15216540701474523 . [DOI] [PubMed] [Google Scholar]
- 48.Zhou Y, Rui L. Major urinary protein regulation of chemical communication and nutrient metabolism. Vitamins and hormones. 2010;83:151–63. Epub 2010/09/14. doi: 10.1016/S0083-6729(10)83006-7 ; PubMed Central PMCID: PMC4034056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Mueller KM, Themanns M, Friedbichler K, Kornfeld JW, Esterbauer H, Tuckermann JP, et al. Hepatic growth hormone and glucocorticoid receptor signaling in body growth, steatosis and metabolic liver cancer development. Molecular and cellular endocrinology. 2012;361(1–2):1–11. Epub 2012/05/09. doi: 10.1016/j.mce.2012.03.026 ; PubMed Central PMCID: PMC3419266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Fernandez-Perez L, Guerra B, Diaz-Chico JC, Flores-Morales A. Estrogens regulate the hepatic effects of growth hormone, a hormonal interplay with multiple fates. Frontiers in endocrinology. 2013;4:66 Epub 2013/06/14. doi: 10.3389/fendo.2013.00066 ; PubMed Central PMCID: PMC3670000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Safe S. Molecular biology of the Ah receptor and its role in carcinogenesis. Toxicology letters. 2001;120(1–3):1–7. Epub 2001/04/27. . [DOI] [PubMed] [Google Scholar]
- 52.Peterson ML, Ma C, Spear BT. Zhx2 and Zbtb20: novel regulators of postnatal alpha-fetoprotein repression and their potential role in gene reactivation during liver cancer. Seminars in cancer biology. 2011;21(1):21–7. Epub 2011/01/11. doi: 10.1016/j.semcancer.2011.01.001 ; PubMed Central PMCID: PMC3313486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Gargalovic PS, Erbilgin A, Kohannim O, Pagnon J, Wang X, Castellani L, et al. Quantitative trait locus mapping and identification of Zhx2 as a novel regulator of plasma lipid metabolism. Circulation Cardiovascular genetics. 2010;3(1):60–7. Epub 2010/02/18. doi: 10.1161/CIRCGENETICS.109.902320 ; PubMed Central PMCID: PMC2846770. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Morford LA, Davis C, Jin L, Dobierzewska A, Peterson ML, Spear BT. The oncofetal gene glypican 3 is regulated in the postnatal liver by zinc fingers and homeoboxes 2 and in the regenerating liver by alpha-fetoprotein regulator 2. Hepatology. 2007;46(5):1541–7. Epub 2007/08/03. doi: 10.1002/hep.21825 . [DOI] [PubMed] [Google Scholar]
- 55.Zhang Y, Fang B, Damle M, Guan D, Li Z, Kim YH, et al. HNF6 and Rev-erbalpha integrate hepatic lipid metabolism by overlapping and distinct transcriptional mechanisms. Genes & development. 2016;30(14):1636–44. Epub 2016/07/23. doi: 10.1101/gad.281972.116 ; PubMed Central PMCID: PMC4973293. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(XLSX)
(XLSX)
Microarray datasets for basal gene expression for 96 different male mouse tissues/cell types were obtained from Lattin et al. [34]. The difference between the microarray signal in the liver and the maximum fluorescent signal on a per gene basis in all other tissues/cells was calculated (ΔSignal). A gene was considered liver-specific when ΔSignal ≥ 5,000 units.
(DOCX)
Data was collected and analyzed as described in materials & methods.
(DOCX)
(DOCX)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files or available on GEO (https://www.ncbi.nlm.nih.gov/geo/) using accession numbers GSE62902, GSE81990, GSE87519, GSE97634, and GSE97636.