

Abstract
Suberoylanilide hydroxamic acid (SAHA) has been assessed in clinical trials as part of a “shock and kill” strategy to cure HIV-infected patients. While it was effective at inducing expression of HIV RNA (“shock”), treatment with SAHA did not result in a reduction of reservoir size (“kill”). We therefore utilized a combined analysis of effects of SAHA on the host transcriptome and proteome to dissect its mechanisms of action that may explain its limited success in “shock and kill” strategies. CD4+ T cells from HIV seronegative donors were treated with 1 µM SAHA or its solvent dimethyl sulfoxide (DMSO) for 24 hours. Protein expression and post-translational modifications were measured with iTRAQ proteomics using ultra high-precision two-dimensional liquid chromatography - tandem mass spectrometry. Gene expression was assessed by Illumina microarrays. Using limma package in the R computing environment, we identified 185 proteins, 18 phosphorylated forms, 4 acetylated forms and 2,982 genes, whose expression was modulated by SAHA. A protein interaction network integrating these 4 data types identified the HIV transcriptional repressor HMGA1 to be upregulated by SAHA at the transcript, protein and acetylated protein levels. Further functional category assessment of proteins and genes modulated by SAHA identified gene ontology terms related to NFκB signaling, protein folding and autophagy, which are all relevant to HIV reactivation. In summary, SAHA modulated numerous host cell transcripts, proteins and post-translational modifications of proteins, which would be expected to have very mixed effects on the induction of HIV-specific transcription and protein function. Proteome profiling highlighted a number of potential counter-regulatory effects of SAHA with respect to viral induction, which transcriptome profiling alone would not have identified. These observations could lead to a more informed selection and design of other HDACi with a more refined targeting profile, and prioritization of latency reversing agents of other classes to be used in combination with SAHA to achieve more potent induction of HIV expression.
Keywords: HIV latency, suberoylanilide hydroxamic acid, iTRAQ mass spectrometry, HMGA1, autophagy, protein folding
1. Introduction
The persistent cellular reservoir of HIV provirus is a major obstacle to a cure (Richman et al., 2009). The “shock and kill” treatment strategy has been envisioned as a controlled induction of virus reactivation in the presence of combination antiretroviral therapy (cART) to reveal latently infected cells for immune system recognition and destruction (Ylisastigui et al., 2004). Histone deacetylase (HDAC) inhibitor (HDACi) suberoylanilide hydroxamic acid (SAHA), an FDA-approved compound for treatment of cutaneous T cell lymphoma (Mann et al., 2007), has been used in clinical trials to reactivate HIV to reduce the size of the latent reservoir (Archin et al., 2014; Archin et al., 2012; Elliott et al., 2014). Exposure to HDACis is tightly associated with histone hyperacetylation and chromatin decondensation, which provides a transcriptionally favorable environment for HIV reactivation (Matalon et al., 2011). SAHA was effective at inducing HIV RNA expression in most patients on cART with suppressed viremia (Archin et al., 2012; Elliott et al., 2014); however, treatment with SAHA did not result in a reduction of reservoir size (Archin et al., 2014; Elliott et al., 2014). SAHA is also used as a synergistic agent to screen in vitro for other latency reversing agents (LRAs); therefore, limitations of its activity require further elucidation. Understanding potential counter-regulatory effects of SAHA on HIV reactivation will guide the selection of modifications of this compound and prioritization of LRAs in combination therapies.
SAHA was well tolerated in HIV-infected patients (Archin et al., 2014; Archin et al., 2012; Elliott et al., 2014), and an in vitro treatment of primary CD4+ T cells with a physiological concentration of SAHA elicited only modest effects on gene expression (Beliakova-Bethell et al., 2013). However, genes induced by SAHA may specifically regulate the state of HIV latency (Shirakawa et al., 2013), so that the net effect of SAHA on these genes results in insufficient viral induction to kill a cell. The function of non-histone targets of HDACs (e.g. chaperone protein HSP90) may also be modulated by SAHA (Choudhary et al., 2009). SAHA binds not only HDACs, but other proteins as well (Bantscheff et al., 2011), opening the possibility for direct regulation of additional targets. A non-histone effect of SAHA relevant to HIV reactivation was previously demonstrated by the Peterlin group (Contreras et al., 2009). In this case, SAHA promoted HIV reactivation by causing the release of positive transcription elongation factor (p-TEFb) from its inactive complex, which is required for Tat-mediated transcriptional elongation. Ultimately, multiple steps in the HIV replication cycle have to be successfully completed to reveal the infected cell to the immune system. These include cell signaling leading to proper assembly of transcription factors on the long terminal repeat (LTR), transcription, RNA splicing, RNA nuclear export, protein translation, and membrane trafficking. Systems-wide studies would enhance our understanding of complex effects of SAHA on key cellular pathways and processes required for HIV reactivation.
RNA expression profiling by microarrays and RNA-Seq technology have been the foremost strategies for identifying genome-wide effects of a disease or a treatment. Studies using SAHA demonstrated downregulation of a subset of genes (Beliakova-Bethell et al., 2013; LaBonte et al., 2009; Wozniak et al., 2010), which is consistent with the existence of the secondary mechanisms of action and cannot be explained by chromatin decondensation. Transcriptomic methods are sensitive and capable of detecting the majority of the annotated genes; however, gene expression studies do not uncover the effects at the functional (protein) level. Liquid chromatography - mass spectrometry (LC-MS) based proteomics methods may be used to confirm functionality of transcripts. In addition, despite currently being less sensitive than transcriptomics, LC-MS proteomics can identify the endophenotypic effects not otherwise reflected in the transcriptome including the occurrence of post-transnationally modified proteins. We have therefore performed non-targeted quantitative iTRAQ proteomics experiments by ultra-high precision two-dimensional LC-MS using human primary CD4+ T cells treated with SAHA. By combining proteomic and transcriptomic datasets, we performed integrated data analysis for a more complete characterization of the secondary effects of SAHA. Based on protein function established in published literature, we propose that some of the observed effects of SAHA may have relevance to HIV reactivation, i.e. enhance or inhibit HIV transcription.
2. Materials and Methods
2.1. CD4+ T cell isolation and SAHA treatment
Healthy donor volunteers provided written informed consent using a protocol approved by UCSD IRB. Primary CD4+ T cells were isolated and cultured as described previously (Beliakova-Bethell et al., 2013). All CD4+ T cell samples had >98% purity and <5% activation (HLA-DR+), as assessed by flow cytometry. Prior to treatment, cell concentration was adjusted to 2.5 million per ml with fresh medium. Cells were treated with 1 µM SAHA or its solvent dimethyl sulfoxide (DMSO) and plated into 6-well tissue culture plates at 2 ml/well. Following 24 hours of treatment, 10–15 million cells were collected after washing 4 times with 50 ml phosphate buffered saline to remove all traces of serum proteins. Cell pellets were frozen in dry ice/ethanol bath and stored at −80°C until protein isolation. A separate sample set was treated to validate gene expression by a method independent of high throughput profiling, droplet digital polymerase chain reaction (ddPCR) [(Beliakova-Bethell et al., 2014) and Supplementary Methods].
2.2. Proteomics and transcriptomics datasets
Protein was isolated and liquid chromatography - tandem mass spectrometry was performed as described previously (Al-Daghri et al., 2014; Manousopoulou et al., 2015; Papachristou et al., 2013). Briefly, 100 µg of protein from each sample was extracted, reduced, alkylated, and proteolysed with trypsin. Peptides were labeled with iTRAQ 8-plex, pooled and subjected to two dimensions of liquid chromatography, and were characterized with nano-capillary ultra-performance liquid chromatography hyphenated with a nanospray ionization hybrid LTQ / FT-Obitrap Elite ultra-high resolution mass spectrometry system. Unprocessed raw data files were searched by Proteome Discoverer for native, phosphorylated and acetylated peptides at a peptide false discovery rate of <1% against the human Uniprot proteome. For more details on protein preparation and proteomics, please refer to Supplementary Methods. The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium (Vizcaíno et al., 2014) via the PRIDE partner repository (PXD002150). Transcriptomic data that assessed the effects of SAHA (1 µM) in CD4+ T cells was obtained from our previously published SAHA dose responsive Illumina microarray dataset (Reardon et al., 2015) at the Gene Expression Omnibus (GSE66994).
2.3 Statistical analyses
To identify detected proteins, a statistical processing approach was used that accounts for key mass spectral features to reduce the effects of peptide co-isolation on the resulting iTRAQ reporter ions and thus increase the accuracy of relative protein expression. Proteins were median-normalized and converted to log2 paired ratios (SAHA/DMSO). Protein expression values were obtained by averaging peptide intensity values and ratio-filtered. Filtering was not performed on acetylated or phosphorylated forms due to their intrinsic low abundance. Determination of differentially expressed proteins (DEPs), their phosphorylated (DPPs) and acetylated (DAPs) forms, and genes (DEGs) was done using limma (Smyth, 2004). Genes were further filtered on fold change (|log2 FC| > 1) for the protein interaction network (PIN). The PIN was constructed using Metacore™ from GeneGo, Inc. and visualized with Cytoscape (Shannon et al., 2003). Node colour was subdivided into sections using the MultiColoredNodes package (Warsow et al., 2010). Gene Ontology (GO) analysis was performed using Functional Analysis of Individual Microarray Expression (FAIME) (Yang et al., 2012). Gene membership for each GO term was determined with BioMart (Kasprzyk, 2011) using the Ensemble 78 Genes database and the GRCh38 Dataset (Flicek et al., 2014). GO term differential expression between the SAHA and DMSO control conditions was determined using a paired Student's t-test. In a discovery driven approach, proteins and protein GO terms with a nominal p-value (p) <0.05 were considered significant. For genes and gene GO terms, a false discovery rate-corrected p-value (FDR) <0.05 (Benjamini and Hochberg, 1995) was considered significant. Genes and proteins are referred to by official gene symbol except where noted. Please refer to the Supplementary Methods for details of data analysis procedures.
3. Results
3.1. Proteins and genes modulated by SAHA
The quantitative proteomics profiled 1,547 proteins, identifying 185 DEPs, 18 DPPs and 4 DAPs (p<0.05) between CD4+ T cells from 4 donors treated with SAHA or the DMSO control (Table S1). Identification of DPPs and DAPs was possible thanks to the in-depth and orthogonal two-dimensional liquid chromatographic separation of the tryptic peptides followed by their ultra-high resolution mass spectrometry. However, their non-targeted detection suggests that these in vivo modified proteins had higher abundance relative to other in vivo modified proteins not detected in this study that typically require prior enrichment for their analysis (Papachristou et al., 2013). To compare the effect of SAHA treatment between the proteome and transcriptome, microarray gene expression data were selected from our previous SAHA dose responsive study (Reardon et al., 2015). A paired analysis identified 2,982 genes modulated by SAHA (FDR<0.05) in CD4+ T cells from 6 donors (see Table S2 for the complete list of DEGs at the probe level). The modulation of a large number of these genes was confirmed at the protein level with 56 up- and 49 downregulated at both levels (Figure 1). Even though an order of magnitude more DEGs were identified compared to DEPs, there was near complete agreement in the direction of modulation by SAHA when overlapped at the RNA and protein levels.
Figure 1. Overlap between differentially expressed proteins (DEPs, p<0.05) and genes (DEGs, FDR<0.05).

DEPs and DEGs were identified using linear modeling in R (package limma). The venn diagram was constructed using the VennDiagram package in R. Up- and downregulated genes and proteins are shown.
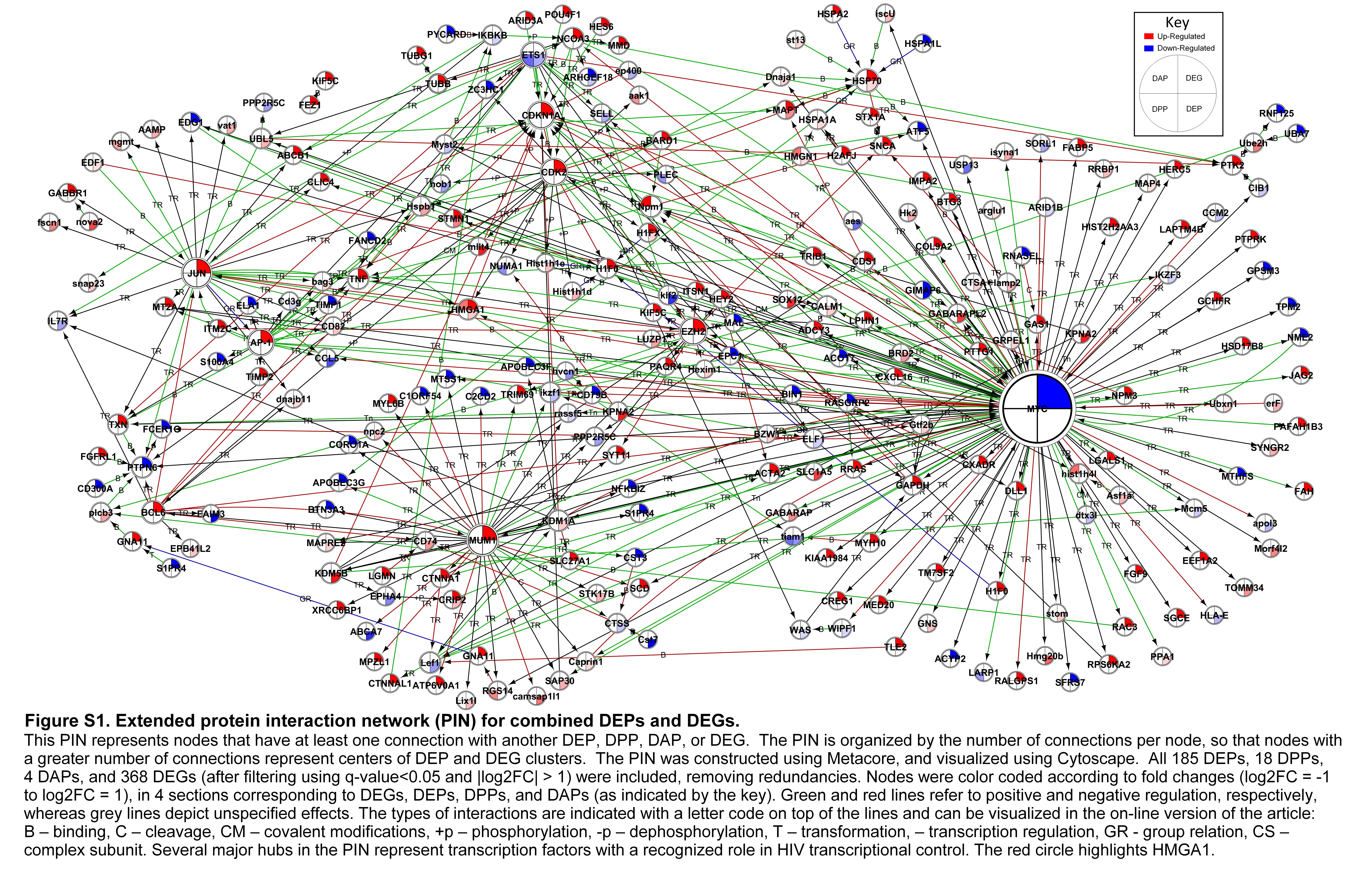
3.2. PINs for proteins and genes modulated by SAHA
It was hypothesized that the most important genes modulated by SAHA would be affected at several different levels (i.e., DEGs, DEPs, DPPs and DAPs). To integrate the 4 datasets, they were superimposed onto a PIN (Figures 2 and S1). For visualization purposes, only genes and proteins with 5 or more connections are presented in Figure 2, whereas the complete PIN is presented in Figure S1. These PINs revealed that high mobility group (HMG) AT-hook 1 (HMGA1) was upregulated at the RNA (DEG), protein (DEP) and acetylated protein (DAP) levels. Heat shock protein 70 (HSP70) was represented in the PIN by 2 genes, HSPA1A, upregulated at the protein level, and HSPA2, upregulated at the RNA level. V-ets avian erythroblastosis virus E26 oncogene homolog 1 (ETS1) was downregulated by SAHA at the protein level, including total and phosphorylated (pS294) forms. A number of well-connected genes were modulated at the protein level only, e.g. lymphoid enhancer-binding factor 1 (LEF1), lysine (K)-specific demethylase 1A (KDM1A), and inhibitor of kappa light polypeptide gene enhancer in B-cells, kinase beta (IKBKB). Another set of well-connected genes was regulated by SAHA only at the RNA level, e.g. v-MYC avian myelocytomatosis viral oncogene homolog (MYC), enhancer of zeste homolog 2 (EZH2), activator protein 1 (AP-1), and nuclear receptor coactivator 3 (NCOA3). While well-connected genes modulated at the protein level by SAHA may have regulatory roles for other genes, the role of genes modulated by SAHA at the RNA level would need further confirmation by more sensitive methods of protein detection.
Figure 2. Protein interaction network (PIN) for combined DEPs, DPPs, DAPs and DEGs.

The PIN was constructed using Metacore, and visualized using Cytoscape. All 185 DEPs, 18 DPPs, 4 DAPs and 368 DEGs (after filtering using FDR<0.05 and |log2FC| > 1) were included, removing redundancies. Nodes were color coded according to fold changes (log2FC = −1 to log2FC = 1), in 4 sections corresponding to DEGs, DEPs, DPPs and DAPs (as indicated by the key). Only nodes that had 5 or more connections to other DEPs or DEGs are shown, while nodes with fewer connections were hidden to improve the quality of the image. Several well-connected DEPs and DEGs in the PIN represent transcription factors with a recognized role in HIV transcriptional control. Green and red lines refer to positive and negative regulation, respectively, whereas grey lines depict unspecified effects. The red circle highlights HMGA1.
3.3. Functional analysis of proteins and genes modulated by SAHA
To better understand the biological processes modulated by the well-connected proteins in the PIN, DEPs and DEGs were subjected to GO analysis using FAIME (Yang et al., 2012). An order of magnitude fewer proteins was detected by quantitative proteomics compared to genes detected by microarrays. Therefore, protein representation across the GO terms was assessed. A good correlation (R2=0.96) was observed between the number of genes and proteins mapping to all the GO terms (Figure S2). The exceptions were the GO terms Plasma membrane proteins and Integral components of membrane, which were under-represented for proteins, consistent with their hydrophobic properties and low abundance that result in poor extraction (Helbig et al., 2010). Overall, 1,484 GO terms were upregulated and 935 downregulated by SAHA at the RNA level. At the protein level, 119 GO terms were upregulated and 146 downregulated. There was good overlap in GO terms (N=40 up and N=20 down) that were significantly modulated by SAHA at the RNA and protein levels (Figure 3 and Table S3). Terms related to chromatin regulation, Negative regulation of chromatin silencing and Nuclear euchromatin, were upregulated by SAHA (Figure 4). Terms related to histone acetylation and histone acetyltransferase complex were downregulated, as was observed previously with a lower dose of SAHA (Beliakova-Bethell et al., 2013). Positive regulation of T cell proliferation and Positive regulation of T cell activation were downregulated. Importantly, a number of terms functionally relevant to HIV replication were identified. Regulation of I-kappaB kinase/NF-kappaB signaling, Protein binding involved in protein folding, Chaperone mediated protein folding requiring cofactor, and Autophagic vacuole were uregulated by SAHA (Figure 4).
Figure 3. Overlap between GO terms modulated by SAHA at the RNA and protein levels.

GO terms modulated by SAHA were identified using FAIME. The venn diagram was constructed using the VennDiagram package in R. Up- and down-regulated GO terms are shown.
Figure 4. GO terms significantly modulated by SAHA.

GO terms modulated by SAHA were identified using FAIME. Terms related to known effects of SAHA and effects of SAHA relevant to HIV reactivation are shown. The heatmap represents values obtained by subtraction of a GO term average FAIME score for DMSO control from a GO term average FAIME score for SAHA (Δ FAIME Score). Red, the difference in FAIME score is greater than 0, and GO term is upregulated as the result of SAHA treatment. Blue, the difference in FAIME score is less than 0, and GO term is downregulated as the result of SAHA treatment. Count refers to the number of cells in the heatmap with the indicated difference in FAIME scores between SAHA and DMSO controls. P1 through P4 indicate samples used for protein analysis; G1 through G6 indicate samples used for gene expression analysis. The same genes and proteins were represented in the GO terms Histone H4-K5 acetylation and Histone H4-K8 acetylation, so these terms are depicted by a single row on the heatmap.
3.4. Validation of gene expression by ddPCR
Six donors, different from the ones who participated in profiling studies, were recruited. Three genes, whose expression was modulated by SAHA both at the RNA and protein levels, were independently validated using ddPCR. Two of the selected genes were upregulated (HMGA1 and ASF1A) and one downregulated (AES) by SAHA. All three genes were significantly modulated by SAHA as determined by ddPCR (Figure 5), in the same direction as in the microarray and quantitative proteomics studies.
Figure 5. Validation of gene expression by droplet digital PCR (ddPCR).

Number of molecules of each target mRNA was normalized to the number of molecules of the normalizer mRNA (RPL27) in one nanogram of total RNA (expressed as copies per thousand RPL27 molecules). Normality of the distribution and equality of variance in the DMSO and SAHA treated groups were assessed in the R computing environment, and either t-test (HMGA1 and ASF1A) or Wilcoxon signed rank test (AES) were performed to assess the difference of expression induced by SAHA. The experiment was performed with cells from 6 independent donors. Error bars represent standard deviation. **, p<0.01; *, 0.01<p<0.05.
4. Discussion
4.1. SAHA transcriptional and post-transcriptional regulation
Quantitative proteomics identified 185 proteins significantly modulated by SAHA. Over half of these proteins (56%) appear to be regulated at the transcriptional level since their corresponding transcripts were also modulated by SAHA (Figure 1). The remaining proteins were not modulated at the RNA level and would not have been detected in a transcriptomics approach, demonstrating the added value of a proteomics approach. Modulation of a protein may be a result of function of another protein whose transcript was upregulated. For example, upregulation of proteins required for translation (e.g. translation initiation factor EIF5B (Pestova et al., 2000)), may result in increase of translation from existing messenger RNAs (Schwanhausser et al., 2011). Protein expression may also be regulated post-translationally via activity of other proteins modulated by SAHA. For example, 3 proteins that regulate ubiqutination state were modulated by SAHA: E3 ubiqutin ligase DTX3L, E2 ubiquting-cojugating enzyme UBE2H, and a deubiquitinase USP13. It is also possible that the changes in protein expression were the result of earlier transient changes in gene expression, which were not captured in the present study. In addition, even though enrichment for post-translationally modified peptides was not performed, changes in expression of 4 acetylated and 18 phosphorylated proteins after SAHA treatment were detected. Altogether, these data are consistent with the idea that SAHA may have much broader secondary effects beyond chromatin modification than previously recognized.
Among the detected 1,547 proteins, 260 were differentially expressed only at the RNA, and not at the protein level, consistent with the idea that transcriptional effects of SAHA do not always translate into protein production (Mohammadi et al., 2014). However, this conclusion should be interpreted with caution due to the smaller sample size used in the proteomics part of the present study (N=4 for the protein vs N=6 for gene expression analyses). Since cells from different biological donors were used for proteomics and transcriptomics studies, it is also possible that some of the variation between identified DEPs and DEGs was the result of donor-to-donor differences in response to SAHA. However, donor-to-donor variation did not likely play a large role since whenever proteins and transcripts were both detected as differentially expressed, they were modulated in the same direction (Figure 1), and expression of selected genes was confirmed by an independent method in an independent cohort (Figure 5).
4.2. Known effects of SAHA translate from the RNA to the protein level
Identification of GO terms related to previously recognized effects of SAHA at the RNA and protein levels gives confidence that the chosen proteomics methodology provides reliable data. For example, upregulation of genes encoding histones, but downregulation of genes encoding components of acetyltransferase complexes was observed previously by transcriptomics (Beliakova-Bethell et al., 2013). Downregulation of acetyltransferases suggests potential mechanisms by which cells attempt to regain control of acetylation following removal by SAHA of their ability to control acetylation through HDACs. Another process known to be downregulated by SAHA was T cell activation (Mohammadi et al., 2014; Reardon et al., 2015). Both Positive regulation of T cell activation, and Positive regulation of T cell proliferation, were downregulated by SAHA at the RNA and protein levels (Figure 4).
4.3. Effects of SAHA on RNA and proteins with a role in HIV reactivation
A number of HMG proteins (HMGA1, HMGN1, HMG20B, LEF1) were modulated by SAHA at the protein and/or RNA levels. Like histones, HMG proteins regulate chromatin dynamics, dependent on post-translational modifications (Zhang and Wang, 2010). The most remarkable observation, made possible by using integrated proteomics and transcriptomics data, was the upregulation of HMGA1 at the RNA and protein levels (Figure 2, Tables S1 and S2), as well as a consistent upregulation of its acetylated form (Figure S3). Two mechanisms by which HMGA1 interferes with HIV transcription have been demonstrated. First, it competes with Tat for TAR binding and inhibits both basal and Tat-mediated HIV transcription (Eilebrecht et al., 2013). Second, HMGA1 inhibits transcription of host genes, as well as HIV, by recruiting inactive p-TEFb to target promoters (Eilebrecht et al., 2014). Even though the observed increase of HMGA1 protein was relatively small in the present study (1.15-fold), experimental overexpression (20% increase) (Eilebrecht et al., 2013) resulted in a measurable reduction of LTR activity proportionate to levels of HMGA1 expression. Thus, upregulation of HMGA1 by SAHA would appear to have an undesirable negative effect with respect to HIV reactivation. The HMGA1 protein possesses 5 lysine residues that can be acetylated (Zhang et al., 2007). Of these, acK64 and acK70 have a known function in interferon-beta transcriptional switch in response to viral infection (Munshi et al., 2001). In the present study, the acK14 form of HMGA1 was upregulated by SAHA, for which no specific function has yet been demonstrated.
Identification of HMGA1 (Figure 2) prompted a more in-depth analysis of individual proteins that were modulated by SAHA. We performed a literature search on DEPs and “HIV” or “HIV latency” to determine whether any other proteins modulated by SAHA, besides HMGA1, have a role in activation or repression of HIV transcription (Table 1). Three of the proteins that were found in this search were modulated by SAHA at the RNA level as well (Table 1), and were confirmed by ddPCR (Figure 5). More than half of the identified proteins were not modulated by SAHA at the RNA level. Change in RNA may be transient for some genes, as was noted by Elliott and colleagues (Elliott et al., 2014). For example, BRD2 was detected in their study at the RNA level 2 hours post-treatment, while in the present study it was detected at the protein, but not the RNA level, 24 hours post-treatment. Thus, a proteomics approach has added value to the transcriptomic approach by capturing some of the transient effects on genes at the protein level.
Table 1. Proteins with a known role in HIV reactivation, up- or downregulated as the result of SAHA treatment.
An implied effect of SAHA treatment on HIV transcription, based on the function of a given protein and the direction of its modulation by SAHA, is indicated in the rightmost column.
| Description | Protein (gene symbol) |
Gene name | Reference(s) | Transcript modulated? |
Effect on HIV transcription |
|---|---|---|---|---|---|
| Proteins that act to activate HIV transcription | |||||
| Upregulated by SAHA | HSPA1A | heat shock 70kDa protein 1A | O'Keeffe et al., 2000 | no/yes* | + |
| KDM1A | lysine (K)-specific demethylase 1A | Sakane et al., 2011 | No | + | |
| Downregulated by SAHA | ETS1 | v-ets avian erythroblastosis virus E26 oncogene homolog 1 | Bassuk et al., 1997; Posada et al., 2000; Sieweke et al., 1998; Yang et al., 2009 | No | − |
| LEF1 | lymphoid enhancer-binding factor 1 | Sheridan et al., 1995 | No | − | |
| IKBKB | inhibitor of kappa light polypeptide gene enhancer in B-cells, kinase beta | Mercurio et al., 1997; Nabel and Baltimore, 1987 | No | − | |
|
| |||||
| Proteins that act to inhibit HIV transcription | |||||
| Upregulated by SAHA | HMGA1 | high mobility group AT-hook 1 | Eilebrecht et al., 2014; Eilebrecht et al., 2013 | Yes | − |
| ASF1A | anti-silencing function 1A histone chaperone | Gallastegui et al., 2011 | Yes | − | |
| BRD2 | bromodomain containing 2 | Boehm et al., 2013 | No | − | |
| BRDT | bromodomain, testis-specific | Bisgrove et al., 2007 | No | − | |
| Downregulated by SAHA | AES | amino-terminal enhancer of split | Tetsuka et al., 2000 | Yes | + |
| ARID1B (BAF250) | AT rich interactive domain 1B (SWI1-like) | Mahmoudi, 2012; Rafati et al., 2011 | No | + | |
Plus sign, possible positive effect on HIV transcription, e.g. resulting from upregulation of an activator or downregulation of a repressor. Minus sign, possible negative effect on HIV transcription, e.g. resulting from downregulation of an activator or upregulation of a repressor.
Asterisk indicates that for heat shock protein 70, one gene (HSPA1A) was modulated by SAHA at the protein level only, while another gene (HSPA2) was modulated only at the transcript level.
GO terms, which may be relevant to HIV reactivation and were modulated by SAHA at the RNA and protein levels, included Regulation of I-kappaB kinase/NF-kappaB signaling, Protein binding involved in protein folding, Chaperone mediated protein folding requiring cofactor, and Autophagic vacuole (Figure 4 and Table S3). NFκB signaling is well recognized in HIV transcriptional activation (Nabel and Baltimore, 1987; Osborn et al., 1989). Interestingly, individual genes and proteins significantly upregulated by SAHA and mapping to Regulation of I-kappaB kinase/NF-kappaB signaling had opposite effects on NFκB activity. For example, TNFA, F2RL1/PAR2 (DEGs) and HSPB1 (DEP) activate NFκB (Osborn et al., 1989; Parcellier et al., 2003; Sales et al., 2015) and thus promote HIV reactivation, while ZFAND6 (DEG) represses NFκB (Chang et al., 2011). In addition, IKBKB (DEP) phosphorylates the inhibitor in the inhibitor/NFκB complex (Mercurio et al., 1997), causing dissociation of the inhibitor and activation of NFκB. Its downregulation by SAHA would thus have a negative effect with respect to HIV reactivation. Protein folding may have a role in HIV reactivation, because sequential actions of HSP70 and HSP90 are required for proper folding and stabilization of cyclin-dependent kinase 9 (Cdk9) and assembly of p-TEFb (O'Keeffe et al., 2000). Recently, the role of autophagy in HDACi-induced HIV reactivation and clearance of infected cells has become of interest. In monocyte-derived macrophages, intracellular HIV was shown to be degraded via canonical autophagy pathway upon reactivation with HDACis (Campbell et al., 2015). Lysosomal destabilization following HDACi treatment promoted death of HIV-infected cells, even with incomplete activation of HIV (Stankov et al., 2015). These results indicate the importance of autophagy when using SAHA, and other HDACis, for HIV reactivation.
4.4. Conclusions and implications
The present study is the first to our knowledge to identify proteins and their post-translationally modified forms modulated by SAHA in human primary CD4+ T cells. Combined with the analysis of induced transcriptomic changes, this study demonstrates global regulatory networks affected by SAHA treatment and enhances our understanding of the secondary mechanisms of SAHA action. Expression of a number of genes and proteins with previously reported roles in HIV transcriptional control was modulated by SAHA; some of these effects would appear to be inhibitory for HIV reactivation. Identification of these counter-regulatory effects of SAHA on HIV induction have the potential to strongly impact selection and modification of HDACis and prioritization of other LRAs for future evaluations and advancement to clinical trials. Better potencies for HIV reactivation of the HDACis Romidepsin (Wei et al., 2014) and Panobinostat (Rasmussen et al., 2013) may be due to the lower impact of secondary negative effects possessed by SAHA, which warrants further investigation. Synergistic HIV reactivation when using SAHA with Protein Kinase C (PKC) agonists, such as prostratin and bryostatin (Laird et al., 2015; Williams et al., 2004), may be due to negating adverse effects of SAHA on NFκB signaling pathway by these LRAs. Prostratin and bryostatin are not suitable for use in vivo due to side effects or limited availability; however, other PKCs, such as Ingenol derivatives (Jiang et al., 2014; José et al., 2014), warrant further testing in combination with SAHA using cells from HIV-infected patients ex vivo. Proteome profiling performed in this study revealed a number of potential counter-regulatory effects of SAHA not present at the transcript level. We would therefore recommend using transcriptomic and proteomic profiling as two complementary techniques, transcript profiling being a more sensitive method, and protein profiling for confirmation of transcripts and detection of protein-specific effects.
Supplementary Material
Figure S1. Extended protein interaction network (PIN) for combined DEPs and DEGs.
{kind=link}
Figure S2. Scatterplot correlating number of detected genes and proteins mapping to GO terms.
{kind=link}
Figure S3. The mass spectrometry characterization of the effect of SAHA on HMGA1 expression.
{kind=link}
Supplementary Methods
Table S1. Differentially expressed proteins and their post-translationally modified forms (DEPs, DPPs and DAPs).
Table S2. Differentially expressed genes (DEGs).
Table S3. Gene Ontology (GO) terms modulated by SAHA.
SAHA induces HIV RNA expression, but does not reduce the size of the persistent cellular reservoir of HIV provirus.
We analyzed proteome and transcriptome changes induced by SAHA in human primary CD4+ T cells.
Positive and negative effects of SAHA on genes and proteins with a role in HIV reactivation from latency were identified.
These results may impact selection and modification of HDACis and prioritization of other compounds for future evaluations.
Acknowledgments
We gratefully acknowledge all volunteers at UCSD, who contributed blood, and Merck and Co., Inc., who graciously provided SAHA used in this study. We are indebted to Mr. Roger Allsopp and Mr. Derek Coates for their enthusiasm, fund raising and vision in promoting the FT-MS proteomics platform at the University of Southampton. Thanks to Dr. Xunli Zhang for the kind use of the high-performance liquid chromatography system. We acknowledge the use of the IRIDIS High Performance Computing Facility, and associated support services at the University of Southampton, in the completion of this work. We thank the PRIDE team for the proteomics data processing and repository assistance. This research was supported by the grants from the National Institute of Health, Martin Delaney Collaboratory of AIDS Researchers for Eradication (CARE, U19 AI 096113), through the research infrastructure provided by the UCSD Center for AIDS Research (CFAR, AI 36214) and the James B. Pendleton Charitable Trust. The Wessex Cancer Trust and Medical Research, UK, Hope for Guernsey and the University of Southampton 'Annual Adventures in Research' Grant, made it possible to establish the proteomics infrastructure and its use for this study. This material is based upon work supported in part by the Department of Veterans Affairs (VA), Veterans Health Administration, Office of Research and Development. The views expressed in this article are those of the authors and do not necessarily reflect the position or policy of the Department of Veterans Affairs or the United States government. The sponsors of this research were not involved in the study design, collection or interpretation of the data, manuscript preparation, or the decision to submit the article for publication.
Abbreviations
- SAHA
suberoylanilide hydroxamic acid
- DEPs
differentially expressed proteins
- DAPs
differentially expressed acetylated forms of proteins
- DPPs
differentially expressed phosphorylated forms of proteins
- DEGs
differentially expressed genes
- UCSD
University of California, San Diego
Glossary
- cART
combination antiretroviral therapy
- DMSO
dimethyl sulfoxide
- FAIME
Functional Analysis of Individual Microarray Expression
- FDA
Food and Drug Administration
- FDR
false discovery rate-adjusted p-value
- GO
Gene Ontology
- HDAC
histone deacetylase
- HDACi
histone deacetylase inhibitor
- HIV
human immunodeficiency virus
- HMG
high mobility group
- IRB
Institutional Review Board
- iTRAQ
isobaric tags for relative and absolute quantitation
- LTR
long terminal repeat
- PIN
protein interaction network
- PKC
protein kinase C
- p-TEFb
positive transcription elongation factor
- SAHA
suberoylanelide hydroxamic acid
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Al-Daghri NM, Al-Attas OS, Johnston HE, Singhania A, Alokail MS, Alkharfy KM, Abd-Alrahman SH, Sabico Sl, Roumeliotis TI, Manousopoulou-Garbis A, Townsend PA, Woelk CH, Chrousos GP, Garbis SD. Whole Serum 3D LC-nESI-FTMS Quantitative Proteomics Reveals Sexual Dimorphism in the Milieu Intérieur of Overweight and Obese Adults. J Proteome Res. 2014;13:5094–5105. doi: 10.1021/pr5003406. [DOI] [PubMed] [Google Scholar]
- Archin NM, Bateson R, Tripathy MK, Crooks AM, Yang K-H, Dahl NP, Kearney MF, Anderson EM, Coffin JM, Strain MC, Richman DD, Robertson KR, Kashuba AD, Bosch RJ, Hazuda DJ, Kuruc JD, Eron JJ, Margolis DM. HIV-1 Expression Within Resting CD4+ T Cells After Multiple Doses of Vorinostat. J Infect Dis. 2014;210:728–735. doi: 10.1093/infdis/jiu155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Archin NM, Liberty AL, Kashuba AD, Choudhary SK, Kuruc JD, Crooks AM, Parker DC, Anderson EM, Kearney MF, Strain MC, Richman DD, Hudgens MG, Bosch RJ, Coffin JM, Eron JJ, Hazuda DJ, Margolis DM. Administration of vorinostat disrupts HIV-1 latency in patients on antiretroviral therapy. Nature. 2012;487:482–485. doi: 10.1038/nature11286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bantscheff M, Hopf C, Savitski MM, Dittmann A, Grandi P, Michon A-M, Schlegl J, Abraham Y, Becher I, Bergamini G, Boesche M, Delling M, Dumpelfeld B, Eberhard D, Huthmacher C, Mathieson T, Poeckel D, Reader V, Strunk K, Sweetman G, Kruse U, Neubauer G, Ramsden NG, Drewes G. Chemoproteomics profiling of HDAC inhibitors reveals selective targeting of HDAC complexes. Nat Biotech. 2011;29:255–265. doi: 10.1038/nbt.1759. [DOI] [PubMed] [Google Scholar]
- Beliakova-Bethell N, Massanella M, White C, Lada SM, Du P, Vaida F, Blanco J, Spina CA, Woelk CH. The effect of cell subset isolation method on gene expression in leukocytes. Cytometry Part A. 2014;85A:94–104. doi: 10.1002/cyto.a.22352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beliakova-Bethell N, Zhang J, Singhania A, Lee V, Terry V, Richman DD, Spina CA, Woelk CH. Suberoylanilide hydroxamic acid induces limited changes in the transcriptome of primary CD4+ T cells. AIDS. 2013;27:29–37. doi: 10.1097/QAD.0b013e32835b3e26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benjamini Y, Hochberg Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J R Stat Soc, B MET. 1995;57:289–300. [Google Scholar]
- Campbell GR, Bruckman RS, Chu Y-L, Spector SA. Autophagy Induction by Histone Deacetylase Inhibitors Inhibits HIV Type 1. J Biol Chem. 2015;290:5028–5040. doi: 10.1074/jbc.M114.605428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chang E-J, Ha J, Kang S-S, Lee ZH, Kim H-H. AWP1 binds to tumor necrosis factor receptor-associated factor 2 (TRAF2) and is involved in TRAF2-mediated nuclear factor-kappaB signaling. Int J Biochem Cell Biol. 2011;43:1612–1620. doi: 10.1016/j.biocel.2011.07.010. [DOI] [PubMed] [Google Scholar]
- Choudhary C, Kumar C, Gnad F, Nielsen ML, Rehman M, Walther TC, Olsen JV, Mann M. Lysine Acetylation Targets Protein Complexes and Co-Regulates Major Cellular Functions. Science. 2009;325:834–840. doi: 10.1126/science.1175371. [DOI] [PubMed] [Google Scholar]
- Contreras X, Schweneker M, Chen C-S, McCune JM, Deeks SG, Martin J, Peterlin BM. Suberoylanilide Hydroxamic Acid Reactivates HIV from Latently Infected Cells. J Biol Chem. 2009;284:6782–6789. doi: 10.1074/jbc.M807898200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eilebrecht S, Le Douce V, Riclet R, Targat B, Hallay H, Van Driessche B, Schwartz C, Robette G, Van Lint C, Rohr O, Benecke AG. HMGA1 recruits CTIP2-repressed P-TEFb to the HIV-1 and cellular target promoters. Nucleic Acids Res. 2014;42:4962–4971. doi: 10.1093/nar/gku168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eilebrecht S, Wilhelm E, Benecke B-J, Bell B, Benecke AG. HMGA1 directly interacts with TAR to modulate basal and Tat-dependent HIV transcription. RNA Biol. 2013;10:436–444. doi: 10.4161/rna.23686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elliott JH, Wightman F, Solomon A, Ghneim K, Ahlers J, Cameron MJ, Smith MZ, Spelman T, McMahon J, Velayudham P, Brown G, Roney J, Watson J, Prince MH, Hoy JF, Chomont N, Fromentin R, Procopio FA, Zeidan J, Palmer S, Odevall L, Johnstone RW, Martin BP, Sinclair E, Deeks SG, Hazuda DJ, Cameron PU, Sékaly R-P, Lewin SR. Activation of HIV Transcription with Short-Course Vorinostat in HIV-Infected Patients on Suppressive Antiretroviral Therapy. PLoS Pathog. 2014;10:e1004473. doi: 10.1371/journal.ppat.1004473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Flicek P, Amode MR, Barrell D, Beal K, Billis K, Brent S, Carvalho-Silva D, Clapham P, Coates G, Fitzgerald S, Gil L, Girón CG, Gordon L, Hourlier T, Hunt S, Johnson N, Juettemann T, Kähäri AK, Keenan S, Kulesha E, Martin FJ, Maurel T, McLaren WM, Murphy DN, Nag R, Overduin B, Pignatelli M, Pritchard B, Pritchard E, Riat HS, Ruffier M, Sheppard D, Taylor K, Thormann A, Trevanion SJ, Vullo A, Wilder SP, Wilson M, Zadissa A, Aken BL, Birney E, Cunningham F, Harrow J, Herrero J, Hubbard TJP, Kinsella R, Muffato M, Parker A, Spudich G, Yates A, Zerbino DR, Searle SMJ. Ensembl 2014. Nucleic Acids Res. 2014;42:D749–D755. doi: 10.1093/nar/gkt1196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Helbig AO, Heck AJR, Slijper M. Exploring the membrane proteome—Challenges and analytical strategies. J Proteomics. 2010;73:868–878. doi: 10.1016/j.jprot.2010.01.005. [DOI] [PubMed] [Google Scholar]
- Jiang G, Mendes EA, Kaiser P, Sankaran-Walters S, Tang Y, Weber MG, Melcher GP, Thompson GRr, Tanuri A, Pianowski LF, Wong JK, Dandekar S. Reactivation of HIV latency by a newly modified Ingenol derivative via protein kinase Cd-NF-kB signaling. AIDS. 2014;28:1555–1566. doi: 10.1097/QAD.0000000000000289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- José DP, Bartholomeeusen K, da Cunha RD, Abreu CM, Glinski J, da Costa TBF, Rabay AFMB, Filho LFP, Dudycz LW, Ranga U, Peterlin BM, Pianowski LF, Tanuri A, Aguiar RS. Reactivation of latent HIV-1 by new semi-synthetic ingenol esters. Virology. 2014:462–463. 328–339. doi: 10.1016/j.virol.2014.05.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kasprzyk A. BioMart: driving a paradigm change in biological data management. Database (Oxford) 2011;2011:bar049. doi: 10.1093/database/bar049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- LaBonte M, Wilson P, Fazzone W, Groshen S, Lenz H-J, Ladner R. DNA microarray profiling of genes differentially regulated by the histone deacetylase inhibitors vorinostat and LBH589 in colon cancer cell lines. BMC Med Genomics. 2009;2:67. doi: 10.1186/1755-8794-2-67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laird GM, Bullen CK, Rosenbloom DIS, Martin AR, Hill AL, Durand CM, Siliciano JD, Siliciano RF. Ex vivo analysis identifies effective HIV-1 latency–reversing drug combinations. J Clin Invest. 2015;125:1901–1912. doi: 10.1172/JCI80142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mann BS, Johnson JR, Cohen MH, Justice R, Pazdur R. FDA Approval Summary: Vorinostat for Treatment of Advanced Primary Cutaneous T-Cell Lymphoma. Oncologist. 2007;12:1247–1252. doi: 10.1634/theoncologist.12-10-1247. [DOI] [PubMed] [Google Scholar]
- Manousopoulou A, Woo J, Woelk CH, Johnston HE, Singhania A, Hawkes C, Garbis SD, Carare RO. Are you also what your mother eats? Distinct proteomic portrait as a result of maternal high-fat diet in the cerebral cortex of the adult mouse. Int J Obes. 2015 doi: 10.1038/ijo.2015.35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matalon S, Rasmussen TA, Dinarello CA. Histone deacetylase inhibitors for purging HIV-1 from latent reservoir. Mol Med. 2011;17:466–472. doi: 10.2119/molmed.2011.00076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mercurio F, Zhu H, Murray BW, Shevchenko A, Bennett BL, Li Jw, Young DB, Barbosa M, Mann M, Manning A, Rao A. IKK-1 and IKK-2: Cytokine-Activated IκB Kinases Essential for NF-κB Activation. Science. 1997;278:860–866. doi: 10.1126/science.278.5339.860. [DOI] [PubMed] [Google Scholar]
- Mohammadi P, di Iulio J, Muñoz M, Martinez R, Bartha I, Cavassini M, Thorball C, Fellay J, Beerenwinkel N, Ciuffi A, Telenti A. Dynamics of HIV latency and reactivation in a primary CD4+ T cell model. PLoS Pathog. 2014;10:e1004156. doi: 10.1371/journal.ppat.1004156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Munshi N, Agalioti T, Lomardas S, Merika M, Chen G, Thanos D. Coordination of a transcriptional switch by HMGI(Y) actylation. Science. 2001;293:1133–1136. doi: 10.1126/science.293.5532.1133. [DOI] [PubMed] [Google Scholar]
- Nabel G, Baltimore D. An inducible transcription factor activates expression of human immunodeficiency virus in T cells. Nature. 1987;326:711–713. doi: 10.1038/326711a0. [DOI] [PubMed] [Google Scholar]
- O'Keeffe B, Fong Y, Chen D, Zhou S, Zhou Q. Requirement for a Kinase-specific Chaperone Pathway in the Production of a Cdk9/Cyclin T1 Heterodimer Responsible for P-TEFb-mediated Tat Stimulation of HIV-1 Transcription. J Biol Chem. 2000;275:279–287. doi: 10.1074/jbc.275.1.279. [DOI] [PubMed] [Google Scholar]
- Osborn L, Kunkel S, Nabel GJ. Tumor necrosis factor alpha and interleukin 1 stimulate the human immunodeficiency virus enhancer by activation of the nuclear factor kappa B. Proc Natl Acad Sci U S A. 1989;86:2336–2340. doi: 10.1073/pnas.86.7.2336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Papachristou EK, Roumeliotis TI, Chrysagi A, Trigoni C, Charvalos E, Townsend PA, Pavlakis K, Garbis SD. The Shotgun Proteomic Study of the Human ThinPrep Cervical Smear Using iTRAQ Mass-Tagging and 2D LC-FT-Orbitrap-MS: The Detection of the Human Papillomavirus at the Protein Level. J Proteome Res. 2013;12:2078–2089. doi: 10.1021/pr301067r. [DOI] [PubMed] [Google Scholar]
- Parcellier A, Schmitt E, Gurbuxani S, Seigneurin-Berny D, Pance A, Chantôme A, Plenchette S, Khochbin S, Solary E, Garrido C. HSP27 Is a Ubiquitin-Binding Protein Involved in I-κBα Proteasomal Degradation. Mol Cell Biol. 2003;23:5790–5802. doi: 10.1128/MCB.23.16.5790-5802.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pestova TV, Lomakin IB, Lee JH, Choi SK, Dever TE, Hellen CUT. The joining of ribosomal subunits in eukaryotes requires eIF5B. Nature. 2000;403:332–335. doi: 10.1038/35002118. [DOI] [PubMed] [Google Scholar]
- Rasmussen TA, Søgaard OS, Brinkmann C, Wightman F, Lewin SR, Melchjorsen J, Dinarello C, Østergaard L, Tolstrup M. Comparison of HDAC inhibitors in clinical development: Effect on HIV production in latently infected cells and T-cell activation. Hum Vaccin Immunother. 2013;9:993–1001. doi: 10.4161/hv.23800. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reardon B, Beliakova-Bethell N, Spina CA, Zhang JX, Singhania A, Richman DD, Woelk CH. Dose-dependent gene expression changes in suberoylanilide hydroxamic acid (SAHA) treated resting CD4 positive T cells. AIDS. 2015 doi: 10.1097/QAD.0000000000000839. In press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richman DD, Margolis DM, Delaney M, Greene WC, Hazuda D, Pomerantz RJ. The challenge of finding a cure for HIV infection. Science. 2009;323:1304–1307. doi: 10.1126/science.1165706. [DOI] [PubMed] [Google Scholar]
- Sales KU, Friis S, Konkel JE, Godiksen S, Hatakeyama M, Hansen KK, Rogatto SR, Szabo R, Vogel LK, Chen W, Gutkind JS, Bugge TH. Non-hematopoietic PAR-2 is essential for matriptase-driven pre-malignant progression and potentiation of ras-mediated squamous cell carcinogenesis. Oncogene. 2015;34:346–356. doi: 10.1038/onc.2013.563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schwanhausser B, Busse D, Li N, Dittmar G, Schuchhardt J, Wolf J, Chen W, Selbach M. Global quantification of mammalian gene expression control. Nature. 2011;473:337–342. doi: 10.1038/nature10098. [DOI] [PubMed] [Google Scholar]
- Shannon P, Markiel A, Ozier O, Baliga NS, Wang JT, Ramage D, Amin N, Schwikowski B, Ideker T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003;13:2498–2504. doi: 10.1101/gr.1239303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shirakawa K, Chavez L, Hakre S, Calvanese V, Verdin E. Reactivation of latent HIV by histone deacetylase inhibitors. Trends Microbiol. 2013;21:277–285. doi: 10.1016/j.tim.2013.02.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smyth K. Linear models and empirical Bayes methods for assessing differential expression in microarray experiments. Stat Appl Genet Mol Biol. 2004;3:1–25. doi: 10.2202/1544-6115.1027. [DOI] [PubMed] [Google Scholar]
- Stankov M, Suhr C, Lin H, Panayotova-Dimitrova D, Goffinet C, Behrens G. Latent HIV-1 reactivation and lysosomal destabilization synergize to host cell death. Program and abstracts of the 22nd Conference on Retroviruses and Opportunistic Infections; Seattle, WA. 2015. [Google Scholar]
- Vizcaíno JA, Deutsch EW, Wang R, Csordas A, Reisinger F, Ríos D, Dianes JA, Sun Z, Farrah T, Bandeira N, Binz P-A, Xenarios I, Eisenacher M, Mayer G, Gatto L, Campos A, Chalkley RJ, Kraus H-J, Albar JP, Martinez-Bartolomé S, Apweiler R, Omenn GS, Martens L, Jones AR, Hermjakob H. ProteomeXchange provides globally co-ordinated proteomics data submission and dissemination. Nat Biotechnol. 2014;32:223–226. doi: 10.1038/nbt.2839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Warsow G, Greber B, Falk SSI, Harder C, Siatkowski M, Schordan S, Som A, Endlich N, Schöler H, Repsilber D, Endlich K, Fuellen G. ExprEssence - Revealing the essence of differential experimental data in the context of an interaction/regulation network. BMC Syst Biol. 2010;4:164–164. doi: 10.1186/1752-0509-4-164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wei DG, Chiang V, Fyne E, Balakrishnan M, Barnes T, Graupe M, Hesselgesser J, Irrinki A, Murry JP, Stepan G, Stray KM, Tsai A, Yu H, Spindler J, Kearney M, Spina CA, McMahon D, Lalezari J, Sloan D, Mellors J, Geleziunas R, Cihlar T. Histone Deacetylase Inhibitor Romidepsin Induces HIV Expression in CD4 T Cells from Patients on Suppressive Antiretroviral Therapy at Concentrations Achieved by Clinical Dosing. PLoS Pathog. 2014;10:e1004071. doi: 10.1371/journal.ppat.1004071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williams S, Samuel A, Chen L-F, Kwon H, Fenard D, Bisgrove D, Verdin E, Greene WC. Prostratin antagonizes HIV latency by activating NF-κB. J Biol Chem. 2004;279:42008–42017. doi: 10.1074/jbc.M402124200. [DOI] [PubMed] [Google Scholar]
- Wozniak MB, Villuendas R, Bischoff JR, Aparicio CB, Martínez Leal JF, de La Cueva P, Rodriguez ME, Herreros B, Martin-Perez D, Longo MI, Herrera M, Piris MÁ, Ortiz-Romero PL. Vorinostat interferes with the signaling transduction pathway of T-cell receptor and synergizes with phosphoinositide-3 kinase inhibitors in cutaneous T-cell lymphoma. Haematologica. 2010;95:613–621. doi: 10.3324/haematol.2009.013870. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang X, Regan K, Huang Y, Zhang Q, Li J, Seiwert TY, Cohen EEW, Xing HR, Lussier YA. Single Sample Expression-Anchored Mechanisms Predict Survival in Head and Neck Cancer. PLoS Comput Biol. 2012;8:e1002350. doi: 10.1371/journal.pcbi.1002350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ylisastigui L, Archin NM, Lehrman G, Bosch RJ, Margolis DM. Coaxing HIV-1 from resting CD4 T cells: histone deacetylase inhibition allows latent viral expression. AIDS. 2004;18:1101–1108. doi: 10.1097/00002030-200405210-00003. [DOI] [PubMed] [Google Scholar]
- Zhang Q, Wang Y. HMG Modifications and Nuclear Function. Biochim Biophys Acta. 2010;1799:28. doi: 10.1016/j.bbagrm.2009.11.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang Q, Zhang K, Zou Y, Perna A, Wang Y. A Quantitative Study on the in-vitro and in-vivo Acetylation of High Mobility Group A1 Proteins. J Am Soc Mass Spectrom. 2007;18:1569–1578. doi: 10.1016/j.jasms.2007.05.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Figure S1. Extended protein interaction network (PIN) for combined DEPs and DEGs.
Figure S2. Scatterplot correlating number of detected genes and proteins mapping to GO terms.
Figure S3. The mass spectrometry characterization of the effect of SAHA on HMGA1 expression.
Supplementary Methods
Table S1. Differentially expressed proteins and their post-translationally modified forms (DEPs, DPPs and DAPs).
Table S2. Differentially expressed genes (DEGs).
Table S3. Gene Ontology (GO) terms modulated by SAHA.