

ABSTRACT
Phagocytosis of the Lyme disease-causing pathogen Borrelia burgdorferi has been shown to be important for generating an inflammatory response to the pathogen. As a result, understanding the mechanisms of phagocytosis has been an area of great interest in the field of Lyme disease. Several cell surface receptors that participate in B. burgdorferi phagocytosis have been reported, including the scavenger receptor MARCO and integrin α3β1. We sought to define the mechanisms by which these receptors mediate phagocytosis and to identify signaling pathways activated downstream of these receptors upon contact with B. burgdorferi. We identified both Syk and Src signaling pathways as ones that participate in B. burgdorferi phagocytosis and the resulting cytokine activation. In our studies, we found that both MARCO and integrin β1 play a role in the activation of the Src kinase pathway. However, only integrin β1 participates in the activation of Syk. Interestingly, the integrin activates Syk without the help of the signaling adaptor Dap12 or FcRγ. Thus, we report that multiple pathways participate in B. burgdorferi internalization and that different cell surface receptors act simultaneously in cooperation and independently to mediate phagocytosis.
KEYWORDS: Borrelia burgdorferi, integrin beta 1, Lyme disease, MARCO, scavenger receptor, Src family kinase, Syk, phagocytosis
INTRODUCTION
Phagocytosis is an important host defense process in response to infection. The control of pathogen burden through internalization and subsequent destruction by innate immune cells is a critical process for the response to a number of pathogens (1, 2). Immune responses activated intracellularly postphagocytosis have also been shown to be critical during host responses (1–4). The strength and repertoire of host immune factors produced in response to infection are dependent on pathogen uptake for a number of bacterial infections, including infection by the pathogen Borrelia burgdorferi, the causative agent of Lyme disease (5–8).
Internalization of B. burgdorferi is important for a robust inflammatory response. The process of phagocytosis is a complex event involving cell surface receptors that participate in the attachment and internalization of the pathogen by initiating membrane-proximal signaling pathways. B. burgdorferi internalization can occur through coiling phagocytosis, with the involvement of GTPases such as Cdc42 and Rac1, which lead to actin rearrangement to engulf the bacteria (9, 10). The interaction between the bacterium and cell surface receptors such as integrin αMβ2 and the Fcγ receptor results in the formation of F-actin structures that engage the Wiskott-Aldrich family protein and the Arp2/3 complex, leading to the internalization of the pathogen (9, 11–15). Additionally, B. burgdorferi has been shown to engage other cell surface receptors that also play a role in the internalization of the pathogen, such as the glycosylphosphatidylinositol (GPI)-anchored protein CD14, integrin α3β1, and the scavenger receptor (SR) MARCO (13, 14, 16–18).
SRs have traditionally been associated with the recognition of various modified low-density lipoproteins (LDLs). However, they have also been shown to respond to microbes and microbial products (19). The engagement of oxidized LDL with the class B SR CD36 causes the activation of Src family kinases (SFKs), Fyn and/or Lyn, and focal adhesion kinase 1 (FAK1), resulting in actin polymerization and increased cell spreading (20, 21). The exact nature of the interaction between the C-terminal tail of SRs and SFKs is unclear, and a specific docking site has not been identified. Without a discernible cytoplasmic signaling domain, SRs are thought to require the cooperation of other cell surface molecules in a “signalosome,” a protein complex that initiates signaling (19). The nature of the ligand likely determines the type of signalosome formed. While it has been shown that the class A SR MARCO is involved in the internalization of ligands, including B. burgdorferi, the mechanism of its involvement is unclear (17).
Integrins are also phagocytic receptors that internalize a variety of ligands and participate in active signaling mediated by the cytoplasmic domain of their β-tails (22–25). Upon engagement by a ligand, a variety of adaptor proteins are recruited to the integrins, forming a signaling complex that assembles into a focal adhesion structure along with kinases, phosphatases, and proteases that regulate integrin function (22–26). Integrin β-chain cytoplasmic tails contain important motifs, such as the NPxY motif, which are phosphorylated at tyrosine residues and important for the binding of adaptor proteins and the activation of downstream signaling molecules (22, 27). SFKs and FAK are some of the adaptor molecules that are known to participate in integrin-mediated signaling (26).
SFKs and the spleen tyrosine kinase (Syk) have been shown to be important for phagocytosis and signaling in response to FcγR-bound particles (28, 29). SFK activity has also been linked to Toll-like receptor 2 (TLR2) inflammatory cytokine signaling (30–32). We hypothesized that SFKs and Syk may be involved in the internalization of B. burgdorferi, contributing to the intracellular immune responses activated by the bacterium. Heit et al. recently examined the signalosome involved in the Syk-mediated internalization of the scavenger receptor CD36 (21). Following activation by oxidized LDL, CD36 associated with β1 and/or β2 integrins and the tetraspanins CD9 and/or CD81. CD36 also engaged the immunoreceptor tyrosine-based activation motif (ITAM)-containing receptor FcRγ, which was an important bridge to Syk activation and, therefore, the internalization of CD36-bound ligands.
In our study, we focused on the roles of the scavenger receptor MARCO and β1 integrin, both of which were previously shown to participate in B. burgdorferi internalization (16, 17). We tested whether MARCO and β1 integrin jointly mediated the activation of Syk, similarly to CD36 and integrins. However, our studies showed that while β1 integrin played a role in Syk activation, MARCO did not. β1 integrin and MARCO instead seem to cooperate in the activation of SFKs and FAK. Surprisingly, the internalization of B. burgdorferi was not dependent on ITAM-containing proteins such as Dap12 and FcRγ, suggesting that they do not act as a bridge to Syk or SFKs. Our data show that multiple signaling pathways are activated upon B. burgdorferi engagement of the host cell and that not all receptors that are engaged and participate in internalization lead to the activation of the same pathways. Thus, there are multiple modes of entry into the cell.
RESULTS
Scavenger receptor MARCO and integrin β1 mediate cytokine activation in response to B. burgdorferi.
Previously, we have shown that both MARCO and integrin α3, which couples only with the β1 chain, participate in the internalization of B. burgdorferi into macrophages (16, 17). To determine the role of MARCO in downstream cytokine secretion, we stimulated wild-type and MARCO-deficient bone marrow-derived macrophage (BMDMs) with B. burgdorferi at a multiplicity of infection (MOI) of 10 for 6 h and measured the secretion of cytokines such as interleukin-6 (IL-6) and tumor necrosis factor alpha (TNF-α) (Fig. 1A). The secretion of both IL-6 and TNF-α was reduced in MARCO-deficient cells. In addition, we wanted to determine if integrin β1 specifically participates in cytokine activation downstream of B. burgdorferi. Because β1-deficient mice are embryonic lethal, we used β1-deficient mouse embryonic fibroblasts (MEFs) and measured cytokine levels after a 24-h stimulation. Upon B. burgdorferi stimulation, β1-deficient MEFs secrete fewer cytokines than do MEFs reconstituted with β1 integrin (Fig. 1B).
FIG 1.
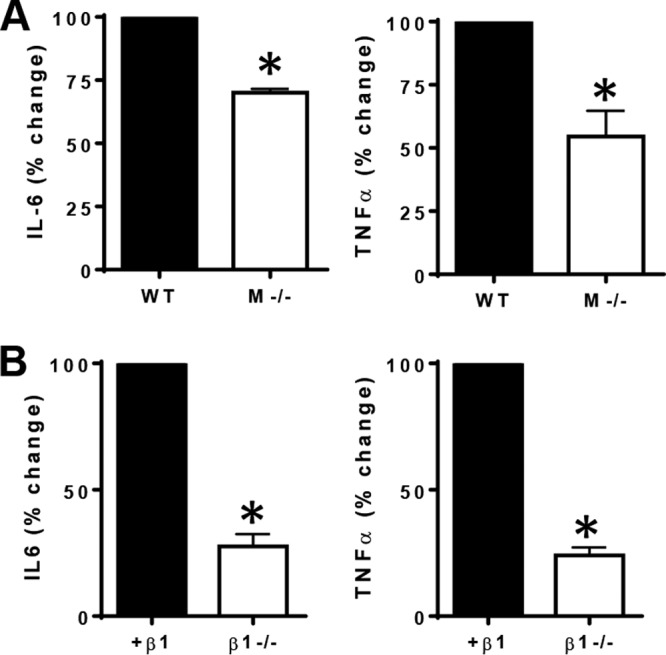
The scavenger receptor MARCO and integrin β1 mediate cytokine activation in response to B. burgdorferi. (A) Bone marrow-derived macrophages (BMDMs) from wild-type (WT) or MARCO-deficient (M−/−) mice were stimulated with B. burgdorferi at an MOI of 10 for 6 h. Supernatants were collected, and the concentrations of secreted IL-6 and TNF-α were determined by an ELISA. Data are shown as means and standard errors of the means of results from three independent experiments. *, P < 0.05. (B) GD25 mouse embryonic fibroblasts (MEFs) deficient in integrin β1 (β1−/−) or reconstituted with full-length integrin β1 (+β1) were stimulated with B. burgdorferi at an MOI of 10 for 24 h. Supernatants were collected, and the concentrations of secreted IL-6 and TNF-α were determined by an ELISA. Data are shown as means and standard errors of the means of results from three independent experiments. *, P < 0.05.
Syk and Src family kinases have a role in phagocytosis and signaling in response to B. burgdorferi.
Src kinase activity has been suggested to be important for the invasion of B. burgdorferi into nonphagocytic cells (33), and Syk kinase has been suggested to play a role in inflammatory signaling (34). Given the role of macrophages in the internalization of B. burgdorferi and the activation of inflammatory immune responses, we were interested in addressing which signaling pathways were necessary for B. burgdorferi phagocytosis in BMDMs. Considering that Syk kinase and Src SFK pathways are known pathways that are activated downstream of scavenger receptors and/or integrins, we first tested their involvement in B. burgdorferi phagocytosis. We used inhibitors of Syk (piceatannol) and Src family (Src-1 and PP1) kinases to measure the effect of the inhibition of these pathways on B. burgdorferi-induced cytokine secretion. In the presence of Syk and SFK inhibitors, the levels of secretion of IL-6 and TNF-α by BMDMs were reduced (Fig. 2A). Syk and SFK inhibitors also resulted in reduced B. burgdorferi phagocytosis, suggesting that these pathways were involved in the internalization of the bacterium (Fig. 2B). To confirm that B. burgdorferi stimulation induces these pathways, we performed phosphoprotein enrichment of cell lysates before and after 30 min of B. burgdorferi stimulation. Lysates were passed through a column, which enriches for proteins that have phosphorylated Ser, Thr, and Tyr residues. The phosphoprotein-containing fractions were then subjected to SDS-PAGE and immunoblotting for total Syk and SFKs. To control for equal protein loading, we also blotted the input lysates prior to phosphoprotein enrichment. We found that B. burgdorferi induces the phosphorylation of both Syk and SFKs in BMDMs (Fig. 2C).
FIG 2.

Src family and Syk kinases have a role in phagocytosis and signaling in response to B. burgdorferi. (A) Wild-type BMDMs were pretreated with Src-1, PP1 (Src family kinase inhibitor), and piceatannol (Syk inhibitor) for 30 min prior to the addition of B. burgdorferi at an MOI of 10. Cells were stimulated for 6 h, supernatants were collected, and the concentrations of secreted IL-6 and TNF-α were determined by an ELISA. Data are shown as means and standard errors of the means of results from three independent experiments. *, P < 0.05. (B) Wild-type BMDMs were stimulated with Bb-GFP for 60 min at 37°C. Any inhibitors or blocking agents used were added for 30 min prior to Bb-GFP stimulation. Following Bb-GFP stimulation, cells were fixed in 1% paraformaldehyde and stained in 2% goat serum and 1% saponin. Noninternalized B. burgdorferi bacteria were stained with anti-B. burgdorferi antibodies followed by a secondary anti-rabbit Alexa Fluor 594 antibody. Phagocytosis is shown as percent internalized bacteria per cell. *, P < 0.05. (C) Wild-type BMDMs were stimulated with B. burgdorferi at an MOI of 10 for 60 min. Cells were lysed and enriched for phosphoproteins. Samples from the input pool and after phosphoenrichment were run on an SDS-PAGE gel and blotted with antibodies to Syk and Src. Shown is a representative blot from three independent experiments.
The ITAM-containing adaptors Dap12 and FcRγ do not participate in B. burgdorferi phagocytosis and signaling.
We were interested in determining which signaling molecules may participate in bridging the integrin or the scavenger receptor MARCO to Syk and Src family signaling. Previously reported work with the scavenger receptor CD36 and its interaction with β1 and β2 integrins showed the involvement of ITAM-containing proteins such as Dap12 and FcRγ in bridging the interaction with Syk. Because both Syk and SFKs played a role downstream of B. burgdorferi stimulation, we were interested in determining if these ITAM-containing proteins also played a role in bridging B. burgdorferi-initiated signaling to Syk and SFKs. We isolated BMDMs from Dap12- and FcRγ-deficient mice and stimulated them with B. burgdorferi at an MOI of 10 for 6 h to measure cytokine secretion or for 1 h to measure bacterial internalization. Neither Dap12- nor FcRγ-deficient BMDMs showed a defect in cytokine induction or phagocytosis in response to B. burgdorferi, suggesting that neither of these ITAM-containing proteins participates in bridging cell surface receptor signaling to downstream phagocytic signaling pathways (Fig. 3).
FIG 3.

The ITAM-containing adaptors Dap12 and FcRγ do not participate in B. burgdorferi phagocytosis and signaling. (A) BMDMs from wild-type (WT) and Dap12- or FcRγ-deficient mice were stimulated with B. burgdorferi at an MOI of 10 for 6 h. Supernatants were collected, and the concentrations of secreted IL-6 and TNF-α were determined by an ELISA. Data are shown as means and standard errors of the means of results from three independent experiments. *, P < 0.05. (B) BMDMs from wild-type and Dap12- or FcRγ-deficient mice were stimulated with Bb-GFP for 60 min at 37°C. Any inhibitors or blocking agents used were added for 30 min prior to Bb-GFP stimulation. Following Bb-GFP stimulation, cells were fixed in 1% paraformaldehyde and stained with 2% goat serum and 1% saponin. Noninternalized B. burgdorferi bacteria were stained with anti-B. burgdorferi antibodies followed by a secondary anti-rabbit Alexa Fluor 594 antibody. Phagocytosis is shown as percent internalized bacteria per cell. *, P < 0.05.
Integrin β1 but not MARCO mediates Syk activation in response to B. burgdorferi.
Next, we wanted to determine which cell surface receptors, MARCO and/or integrin β1, link to downstream Syk and Src family kinase pathways. Wild-type and MARCO-deficient BMDMs or β1-deficient and reconstituted MEFs were stimulated with B. burgdorferi at an MOI of 10 for 30 and 60 min, respectively. Cell lysates were enriched for phosphorylated proteins by using a phosphoprotein enrichment column. Samples were run on SDS-PAGE gels and blotted for total Syk. We found that MARCO deficiency did not affect the levels of Syk phosphorylation (Fig. 4A) but that integrin β1 was required for the efficient activation of Syk (Fig. 4B). This suggests that MARCO and integrin β1 do not cooperate to activate Syk and that MARCO utilizes another pathway to internalize B. burgdorferi.
FIG 4.

Integrin β1 but not MARCO mediates Syk activation in response to B. burgdorferi. (A) Wild-type (WT) and MARCO-deficient (M−/−) BMDMs were stimulated with B. burgdorferi at an MOI of 10 for 60 min (+ Bb) or the control (No Bb). Cells were lysed and enriched for phosphoproteins. Samples from the input pool and after phosphoenrichment were run on SDS-PAGE gels and blotted with antibodies to Syk. Shown is a representative blot from three independent experiments. Densitometry was performed by using ImageJ, and data from three independent experiments are shown as mean values (± standard deviations) of the intensities of protein bands of interest normalized to the input. *, P < 0.05. (B) GD25 integrin β1-null MEFs (β1−/−) and integrin β1-reconstituted GD25 MEFs (+β1) were stimulated with B. burgdorferi at an MOI of 10 for 60 min or the control. Cells were lysed and enriched for phosphoproteins. Samples from the input pool and after phosphoenrichment were run on SDS-PAGE gels and blotted with antibodies to Syk. Shown is a representative blot from three independent experiments. Densitometry was performed by using ImageJ, and data from three independent experiments are shown as mean values (± standard deviations) of the intensity of protein bands of interest normalized to the input. *, P < 0.05. (C) GD25 integrin β1-null MEFs, integrin β1-reconstituted GD25 MEFs, and GD25 MEFs reconstituted with β1 integrin mutated at the NPxY motif (+β1 Y-F) were stimulated with B. burgdorferi at an MOI of 10 for 60 min. Cells were lysed and enriched for phosphoproteins. Samples from the input pool and after phosphoenrichment were run on SDS-PAGE gels and blotted with antibodies to Syk. Shown is a representative blot from three independent experiments. Densitometry was performed by using ImageJ, and data are shown as mean values (± standard deviations) of the intensities of protein bands of interest normalized to the input from three independent experiments. (C) Integrin β1-reconstituted GD25 MEFs and GD25 MEFs reconstituted with β1 integrin mutated at the NPxY motif were stimulated with B. burgdorferi at an MOI of 10 for 24 h. Supernatants were collected, and concentrations of secreted IL-6 and TNF-α were determined by an ELISA. Data are shown as means and standard errors of the means of results from three independent experiments. *, P < 0.05.
The β1 integrin cytoplasmic tail contains an NPxY signaling motif. Phosphorylation on the tyrosine residue has been shown to be important for the activation of a number of downstream signaling molecules, including Syk and SFKs. We investigated the role of the NPxY motif in mediating the interaction of integrin with downstream signaling adaptors in response to B. burgdorferi. Integrin β1-deficient MEFs (β1−/−) reconstituted with full-length β1 integrin (+β1) or β1 integrin with mutations in two tyrosine residues in the NPxY motifs (β1 Y-F) were tested to determine the role of the NPxY motif in Syk phosphorylation. Our data suggest that the NPxY motif in the cytoplasmic tail of β1 integrin was required for the efficient phosphorylation of Syk (Fig. 4C). The NPxY motif of β1 integrin was also required for the efficient activation of cytokines in response to B. burgdorferi (Fig. 4D).
Both integrin β1 and MARCO activate the Src family kinases in response to B. burgdorferi.
Next, we wanted to determine the role of integrin and MARCO engagement in the activation of SFKs in response to B. burgdorferi. MARCO-deficient and wild-type BMDMs (Fig. 5A) or β1-deficient and reconstituted MEFs (Fig. 5B) were stimulated with B. burgdorferi at an MOI of 10 for 30 and 60 min, respectively. Cell lysates were enriched for phosphorylated proteins by using a phosphoprotein enrichment column. Samples were run on SDS-PAGE gels and blotted for SFKs. We found that both MARCO and integrin β1 were required for the efficient activation of SFKs (Fig. 5A and B). Downstream of B. burgdorferi activation, MARCO was also required for the activation of the FAK kinase, which participates in SR- and integrin-mediated phagocytosis (26, 34) (Fig. 5C). This suggests that although MARCO and integrin do not cooperate to induce Syk phosphorylation, they both contribute to the induction of SFKs and FAK.
FIG 5.

Both integrin β1 and MARCO activate Src family kinases in response to B. burgdorferi. (A) Wild-type (WT) and MARCO-deficient (M−/−) BMDMs were stimulated with B. burgdorferi at an MOI of 10 for 60 min (+ Bb) or the control (No Bb). Cells were lysed and enriched for phosphoproteins. Samples from the input pool and after phosphoenrichment were run on SDS-PAGE gels and blotted with antibodies to SFKs. Shown is a representative blot from three independent experiments. Densitometry was performed by using ImageJ, and data from three independent experiments are shown as mean values (± standard deviations) of the intensities of protein bands of interest normalized to the input. *, P < 0.05. (B) GD25 integrin β1-null MEFs (β1−/−) and integrin β1-reconstituted GD25 MEFs (+β1) were stimulated with B. burgdorferi at an MOI of 10 for 60 min or the control. Cells were lysed and enriched for phosphoproteins. Samples from the input pool and after phosphoenrichment were run on SDS-PAGE gels and blotted with antibodies to SFKs. Shown is a representative blot from three independent experiments. Densitometry was performed by using ImageJ, and data from three independent experiments are shown as mean values (± standard deviations) of the intensities of protein bands of interest normalized to the input. *, P < 0.05. (C) Wild-type and MARCO-deficient BMDMs were stimulated with B. burgdorferi at an MOI of 10 for 60 min. Cells were lysed and enriched for phosphoproteins. Samples from the input pool and after phosphoenrichment were run on SDS-PAGE gels and blotted with antibodies to FAK. Shown is a representative blot from three independent experiments. Densitometry was performed by using ImageJ, and data from three independent experiments are shown as mean values (± standard deviations) of the intensities of protein bands of interest normalized to the input. *, P < 0.05.
DISCUSSION
Our study focused on understanding how the scavenger receptor MARCO and integrin β1 participate in the internalization of B. burgdorferi and the activation of downstream signaling pathways. We have previously shown that both of these phagocytic receptors participate in B. burgdorferi-mediated phagocytosis; however, in this study, we aimed to elucidate the signaling pathways activated downstream of these two receptors that mediate the phagocytic process (16, 17). We observed that both MARCO and β1 integrin are important for downstream cytokine activation in response to B. burgdorferi. We showed that the Src family and Syk kinases were phosphorylated in cells stimulated with B. burgdorferi and that Src family and Syk kinases were important for phagocytosis and cytokine responses to the bacteria.
Although we focused on MARCO and α3β1 integrin, there are a number of different receptors that have been shown to participate in B. burgdorferi phagocytosis. Coiling phagocytosis of opsonized B. burgdorferi has been the preferred method of internalization of the bacterium (9, 11, 35). Receptors such as the Fcγ receptor; the mannose receptor; the CR3 receptor, also known as integrin αMβ2; and integrin αvβ3 have all been previously shown to participate in attachment to and/or internalization of the pathogen (11, 35–37). Interestingly, αMβ2 was required for the attachment of the bacterium to the host cell but not for its internalization (13, 14). Internalization required the cooperation of the GPI-anchored receptor CD14 and the binding of the integrin to the C-lectin domain of CD14 (14). However, it is not known which phagocytosis-inducing signaling pathways are activated downstream of the integrin-CD14 interaction. Importantly, the internalization of B. burgdorferi into host cells is not fully dependent on any one of these receptors, and thus, there is likely a good degree of redundancy in the phagocytic pathways being activated downstream of these receptors.
Our goal was to identify phagocytic signaling pathways that mediate B. burgdorferi internalization, and we hypothesized that MARCO and β1 integrin may cooperate to induce the Syk kinase pathways using a signalosome and that this cooperation would engage pathways mediating phagocytosis in a manner similar to what has been observed for the scavenger receptor CD36. The model for CD36 and integrin signaling required the incorporation of ITAM-containing proteins such as FcRγ to bridge the complex to Syk signaling (21). We found that the ITAM-containing proteins Dap12 and FcRγ, which are involved in Syk signaling in response to other stimuli, were completely dispensable for the internalization of B. burgdorferi and cytokine activation. Although Dap12 and FcRγ do not seem to be required and thus likely do not bridge integrin signaling to Syk, we cannot exclude the possibility that other signaling molecules may participate in the integrin β1/Syk signaling platform. Interestingly, we show that the NPxY motif of the integrin cytoplasmic tail is important for the integrin-mediated activation of Syk in response to B. burgdorferi. Studies with integrin β3 cytoplasmic tails suggest that the binding of Syk directly to the NPxY motif and its activation are not dependent on the phosphorylation status (38, 39). However, in our hands, NPxY mutants of β1 integrin, which are mutated only at the phosphorylation sites, showed reduced Syk activation, suggesting a role for phosphorylation in the activation of Syk downstream of integrin β1 in the absence of ITAM-containing protein mediators.
In contrast to the β1 integrin, MARCO was not required for Syk activation in response to B. burgdorferi. Therefore, we investigated other pathways, such as the Src family kinase pathway, which has been shown to be activated downstream of phagocytic receptors. Both MARCO and integrin β1 were required for the activation of Src family kinases. MARCO was also required for the activation of FAK, a kinase that is also well known to be activated downstream of integrin β1 and requires the phosphorylated NPxY motif (26, 38). This suggests that different receptors cooperate to initiate Src signaling for the internalization of B. burgdorferi. It is as yet unclear whether MARCO and the β1 integrin interact directly at the cell surface or if the complex requires the help of other molecules such as tetraspanins.
Our studies expand our understanding of B. burgdorferi internalization to include specific phagocytic signaling pathways. B. burgdorferi coiling phagocytosis has been shown to include the involvement of GTPases such as Cdc42 and Rac1 (9). The involvement of Syk and SFK signaling and the linking of these pathways to specific receptors contribute to our understanding of the mechanisms of B. burgdorferi internalization. Although MARCO and integrin β1 differ in their activation of Syk kinase, they cooperate in the activation of Src family kinases. The convergence of the two receptors in the activation of pathways such as SFK pathways might explain the partial phagocytic phenotypes observed in the absence of individual receptors. We are now able to build a model where integrin β1 mediates the internalization of the bacterium through the activation of Syk and synergizes with other receptors such as MARCO, at the level of SFKs. This leads to the activation of molecules such as Rac1 and Cdc42, which in turn signal for actin polymerization and rearrangement, leading to phagocytosis (Fig. 6).
FIG 6.
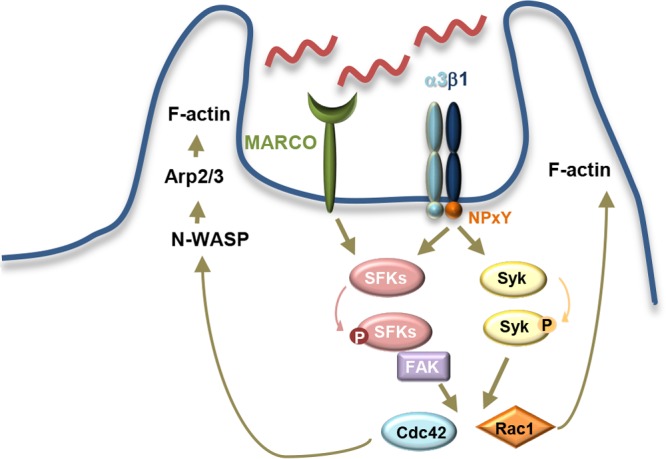
Model of Syk and SFK signaling in B. burgdorferi internalization. B. burgdorferi internalization is mediated by multiple phagocytic receptors, including the scavenger receptor MARCO and integrin β1. In our study, we investigated the roles of these two receptors in activating signaling pathways that lead to phagocytosis. We found that β1 integrin, via its NPxY motif, participates in the activation of Syk, while both MARCO and β1 integrin participate in the activation of the SFK pathway. These pathways have been shown in other systems to lead to the activation of small GTPases such as Rac1 and Cdc42. The small GTPases are important for the phagocytosis of B. burgdorferi via the activation of Wiskott-Aldrich syndrome protein (WASP) and Arp2/3, leading to F-actin polymerization and rearrangement.
The internalization of the pathogen and its localization within the cell are critical for the activation of immune responses (40). Different pathways of internalization triggered by the activation of different receptors have been shown to lead to the differential compartmentalization of the pathogen within the host cell and the differential activation of signaling pathways (24, 27, 41–45). Thus, understanding the pathways of internalization could be significant in understanding the specificity of immune responses.
MATERIALS AND METHODS
Mice and bacterial strains.
FcRγ-deficient (gift from Nora Barrett), Dap12-deficient (gift from Daniel McVicar), and MARCO-deficient mice on a C57BL/6 background were derived as previously described (46–48). All mice were housed in specific-pathogen-free rooms according to institutional guidelines for the humane care and use of laboratory animals approved by the Institutional Animal Care and Use Committee at Tufts University.
Bacteria were cultured in Barbour-Stoenner-Kelly II (BSK-II) complete medium at 37°C to early stationary phase, and the cell density was determined by using a Petroff-Hauser counting chamber. Clonal isolates of infectious, low-passage-number B. burgdorferi sensu stricto (strain N40, clone D10E9) were cultured as described previously (17) and used at an MOI of 10:1 in in vitro experiments.
Reagents and inhibitors.
GD25 mouse embryonic fibroblasts from integrin β1-deficient mice, reconstituted with full-length β1 integrin or with β1 integrin containing YY783/795FF mutations in the NPxY motif, were a kind gift from Deane Mosher at the University of Wisconsin. Cell lines were grown in Dulbecco's modified Eagle's medium (DMEM) plus 10% fetal bovine serum (FBS) with the addition of 10 μg/ml of puromycin for the β1- and YY783/795FF-reconstituted cell lines (26, 27).
Inhibition of Src family and Syk kinases was achieved by using Src-1 (30 μM) (catalog number S2075;), PP1 (10 μM) (catalog number P0040; Sigma), and piceatannol (30 μM) (catalog number P0453; Sigma). Concentrations were chosen based on preliminary experiments and data from previously reported studies (49, 50).
Generation of bone marrow-derived macrophages.
BMDMs were generated as previously described (17, 40). Bone marrow cells were flushed from mouse femurs and tibiae with sterile DMEM and cultured on 100-mm by 15-mm plastic petri dishes for 5 to 7 days in DMEM supplemented with 30% L929 cell-conditioned medium, 20% FBS, and 1% penicillin-streptomycin.
Infection of bone marrow-derived macrophage cultures.
B. burgdorferi bacteria were washed three times with DMEM with 10% FBS in the absence of antibiotics, counted, and resuspended in the same medium. Medium from BMDMs was removed and replaced with the same medium without antibiotics and containing B. burgdorferi at an MOI of 10 in the presence of polymyxin B (Sigma) at 50 μg/ml, to ensure no endotoxin contamination. Cells were harvested at various time points by collecting the cell culture supernatant and then adding TRIzol (Invitrogen) to the remaining cells. In our studies, we did three independent stimulations of BMDMs isolated from three different mice.
Phosphorylated protein enrichment and Western blotting.
Phosphoprotein enrichment was performed by using a phosphoenrichment kit (Pierce Thermo Fisher) according to the manufacturer's instructions. Briefly, 1 × 106 cells were stimulated with B. burgdorferi at an MOI of 10 for 30 min. Cells were washed in PBS and lysed by using Pierce lysis phosphoenrichment buffer. Cells were sonicated and incubated with the phosphoenrichment resin at 4°C overnight. The resin was washed in wash buffer, and the resin was resuspended in 50 μl of 4× lithium dodecyl sulfate (LDS) buffer. Samples were boiled and loaded onto a Tris-glycine–10% polyacrylamide gel. Total Syk antibody (Cell Signaling), total Src family kinase antibody (Cell Signaling), and total FAK antibody (Cell Signaling) were used at 1:1,000 dilutions in a 5% bovine serum albumin (BSA) block.
Phagocytosis assays.
BMDMs were plated at 1 × 105 cells per ml on glass coverslips in non-tissue-culture-treated 24-well plates and were incubated in complete DMEM overnight at 37°C. The following day, medium was replaced with antibiotic-free medium. Cells were stimulated with green fluorescent protein-expressing B. burgdorferi (Bb-GFP) at an MOI of 10:1 for 60 min at 37°C (5). Any inhibitors or blocking agents used were added for 30 min prior to Bb-GFP stimulation. Following Bb-GFP stimulation, cells were fixed in 1% paraformaldehyde prior to staining for immunofluorescence microscopy. We performed three independent phagocytosis assays for each comparison of the wild type and knockout or stimulation with inhibitors.
ELISA measurements.
Supernatants were collected from BMDM cultures at 6 h poststimulation. Cytokine levels were measured by an ELISA using TNF-α (R&D systems) and IL-6 (e-Bioscience) kits according to the manufacturers' instructions.
Statistics.
For ELISAs, the mean percentage of cytokine expression relative to the control is reported, with statistical significance being determined by Mann-Whitney U analysis. For phagocytosis experiments, statistical significance was also determined by Mann-Whitney U analysis. Densitometry significance was determined by using Student's t test.
ACKNOWLEDGMENTS
This work was supported by NIH grants R21AI097971 (to L.T.H.), R21AI097855 (to T.P.-O.), R56AI80846 (to L.T.H.), R01AI076575 (to S.C.B.), R21AI126757 (to L.T.H.), and R01AI103022 (to S.C.B.) and NIGMS grant K12GM074869 (to T.L.K.).
We thank members of the Hu laboratory for helpful discussions and critical review of the manuscript. We thank Dean Mosher at the University of Wisconsin–Madison for providing the GD25 cell lines, Nora Barrett at the Dana-Farber Cancer Institute for providing FcRγ-deficient femurs, and Daniel McVicar at the National Institutes of Health for providing Dap12-deficient femurs for BMDM isolation.
REFERENCES
- 1.Jutras I, Desjardins M. 2005. Phagocytosis: at the crossroads of innate and adaptive immunity. Annu Rev Cell Dev Biol 21:511–527. doi: 10.1146/annurev.cellbio.20.010403.102755. [DOI] [PubMed] [Google Scholar]
- 2.Ip WK, Sokolovska A, Charriere GM, Boyer L, Dejardin S, Cappillino MP, Yantosca LM, Takahashi K, Moore KJ, Lacy-Hulbert A, Stuart LM. 2010. Phagocytosis and phagosome acidification are required for pathogen processing and MyD88-dependent responses to Staphylococcus aureus. J Immunol 184:7071–7081. doi: 10.4049/jimmunol.1000110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Moretti J, Blander JM. 2014. Insights into phagocytosis-coupled activation of pattern recognition receptors and inflammasomes. Curr Opin Immunol 26:100–110. doi: 10.1016/j.coi.2013.11.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Wolf AJ, Arruda A, Reyes CN, Kaplan AT, Shimada T, Shimada K, Arditi M, Liu G, Underhill DM. 2011. Phagosomal degradation increases TLR access to bacterial ligands and enhances macrophage sensitivity to bacteria. J Immunol 187:6002–6010. doi: 10.4049/jimmunol.1100232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Cervantes JL, Dunham-Ems SM, La Vake CJ, Petzke MM, Sahay B, Sellati TJ, Radolf JD, Salazar JC. 2011. Phagosomal signaling by Borrelia burgdorferi in human monocytes involves Toll-like receptor (TLR) 2 and TLR8 cooperativity and TLR8-mediated induction of IFN-beta. Proc Natl Acad Sci U S A 108:3683–3688. doi: 10.1073/pnas.1013776108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Cruz AR, Moore MW, La Vake CJ, Eggers CH, Salazar JC, Radolf JD. 2008. Phagocytosis of Borrelia burgdorferi, the Lyme disease spirochete, potentiates innate immune activation and induces apoptosis in human monocytes. Infect Immun 76:56–70. doi: 10.1128/IAI.01039-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Filgueira L, Nestle FO, Rittig M, Joller HI, Groscurth P. 1996. Human dendritic cells phagocytose and process Borrelia burgdorferi. J Immunol 157:2998–3005. [PubMed] [Google Scholar]
- 8.Shin OS, Isberg RR, Akira S, Uematsu S, Behera AK, Hu LT. 2008. Distinct roles for MyD88 and Toll-like receptors 2, 5, and 9 in phagocytosis of Borrelia burgdorferi and cytokine induction. Infect Immun 76:2341–2351. doi: 10.1128/IAI.01600-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Linder S, Heimerl C, Fingerle V, Aepfelbacher M, Wilske B. 2001. Coiling phagocytosis of Borrelia burgdorferi by primary human macrophages is controlled by CDC42Hs and Rac1 and involves recruitment of Wiskott-Aldrich syndrome protein and Arp2/3 complex. Infect Immun 69:1739–1746. doi: 10.1128/IAI.69.3.1739-1746.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Rittig MG, Krause A, Haupl T, Schaible UE, Modolell M, Kramer MD, Lutjen-Drecoll E, Simon MM, Burmester GR. 1992. Coiling phagocytosis is the preferential phagocytic mechanism for Borrelia burgdorferi. Infect Immun 60:4205–4212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Cinco M, Murgia R, Presani G, Perticarari S. 1997. Integrin CR3 mediates the binding of nonspecifically opsonized Borrelia burgdorferi to human phagocytes and mammalian cells. Infect Immun 65:4784–4789. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Shin OS, Miller LS, Modlin RL, Akira S, Uematsu S, Hu LT. 2009. Downstream signals for MyD88-mediated phagocytosis of Borrelia burgdorferi can be initiated by TRIF and are dependent on PI3K. J Immunol 183:491–498. doi: 10.4049/jimmunol.0900724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Hawley KL, Martin-Ruiz I, Iglesias-Pedraz JM, Berwin B, Anguita J. 2013. CD14 targets complement receptor 3 to lipid rafts during phagocytosis of Borrelia burgdorferi. Int J Biol Sci 9:803–810. doi: 10.7150/ijbs.7136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Hawley KL, Olson CM Jr, Iglesias-Pedraz JM, Navasa N, Cervantes JL, Caimano MJ, Izadi H, Ingalls RR, Pal U, Salazar JC, Radolf JD, Anguita J. 2012. CD14 cooperates with complement receptor 3 to mediate MyD88-independent phagocytosis of Borrelia burgdorferi. Proc Natl Acad Sci U S A 109:1228–1232. doi: 10.1073/pnas.1112078109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Belperron AA, Liu N, Booth CJ, Bockenstedt LK. 2014. Dual role for Fcgamma receptors in host defense and disease in Borrelia burgdorferi-infected mice. Front Cell Infect Microbiol 4:75. doi: 10.3389/fcimb.2014.00075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Marre ML, Petnicki-Ocwieja T, DeFrancesco AS, Darcy CT, Hu LT. 2010. Human integrin alpha(3)beta(1) regulates TLR2 recognition of lipopeptides from endosomal compartments. PLoS One 5:e12871. doi: 10.1371/journal.pone.0012871. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Petnicki-Ocwieja T, Chung E, Acosta DI, Ramos LT, Shin OS, Ghosh S, Kobzik L, Li X, Hu LT. 2013. TRIF mediates Toll-like receptor 2-dependent inflammatory responses to Borrelia burgdorferi. Infect Immun 81:402–410. doi: 10.1128/IAI.00890-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Sahay B, Patsey RL, Eggers CH, Salazar JC, Radolf JD, Sellati TJ. 2009. CD14 signaling restrains chronic inflammation through induction of p38-MAPK/SOCS-dependent tolerance. PLoS Pathog 5:e1000687. doi: 10.1371/journal.ppat.1000687. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Canton J, Neculai D, Grinstein S. 2013. Scavenger receptors in homeostasis and immunity. Nat Rev Immunol 13:621–634. doi: 10.1038/nri3515. [DOI] [PubMed] [Google Scholar]
- 20.Bernasconi MV, Valsangiacomo C, Balmelli T, Peter O, Piffaretti JC. 1997. Tick zoonoses in the southern part of Switzerland (Canton Ticino): occurrence of Borrelia burgdorferi sensu lato and Rickettsia sp. Eur J Epidemiol 13:209–215. doi: 10.1023/A:1007394901846. [DOI] [PubMed] [Google Scholar]
- 21.Heit B, Kim H, Cosio G, Castano D, Collins R, Lowell CA, Kain KC, Trimble WS, Grinstein S. 2013. Multimolecular signaling complexes enable Syk-mediated signaling of CD36 internalization. Dev Cell 24:372–383. doi: 10.1016/j.devcel.2013.01.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Berrier AL, Martinez R, Bokoch GM, LaFlamme SE. 2002. The integrin beta tail is required and sufficient to regulate adhesion signaling to Rac1. J Cell Sci 115:4285–4291. doi: 10.1242/jcs.00109. [DOI] [PubMed] [Google Scholar]
- 23.Caswell PT, Norman JC. 2006. Integrin trafficking and the control of cell migration. Traffic 7:14–21. doi: 10.1111/j.1600-0854.2005.00362.x. [DOI] [PubMed] [Google Scholar]
- 24.Caswell PT, Vadrevu S, Norman JC. 2009. Integrins: masters and slaves of endocytic transport. Nat Rev Mol Cell Biol 10:843–853. doi: 10.1038/nrm2799. [DOI] [PubMed] [Google Scholar]
- 25.Isberg RR, Leong JM. 1990. Multiple beta 1 chain integrins are receptors for invasin, a protein that promotes bacterial penetration into mammalian cells. Cell 60:861–871. doi: 10.1016/0092-8674(90)90099-Z. [DOI] [PubMed] [Google Scholar]
- 26.Wennerberg K, Armulik A, Sakai T, Karlsson M, Fassler R, Schaefer EM, Mosher DF, Johansson S. 2000. The cytoplasmic tyrosines of integrin subunit beta1 are involved in focal adhesion kinase activation. Mol Cell Biol 20:5758–5765. doi: 10.1128/MCB.20.15.5758-5765.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Maginnis MS, Mainou BA, Derdowski A, Johnson EM, Zent R, Dermody TS. 2008. NPXY motifs in the beta1 integrin cytoplasmic tail are required for functional reovirus entry. J Virol 82:3181–3191. doi: 10.1128/JVI.01612-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Crowley MT, Costello PS, Fitzer-Attas CJ, Turner M, Meng F, Lowell C, Tybulewicz VL, DeFranco AL. 1997. A critical role for Syk in signal transduction and phagocytosis mediated by Fcgamma receptors on macrophages. J Exp Med 186:1027–1039. doi: 10.1084/jem.186.7.1027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Fitzer-Attas CJ, Lowry M, Crowley MT, Finn AJ, Meng F, DeFranco AL, Lowell CA. 2000. Fcgamma receptor-mediated phagocytosis in macrophages lacking the Src family tyrosine kinases Hck, Fgr, and Lyn. J Exp Med 191:669–682. doi: 10.1084/jem.191.4.669. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Toubiana J, Rossi AL, Belaidouni N, Grimaldi D, Pene F, Chafey P, Comba B, Camoin L, Bismuth G, Claessens YE, Mira JP, Chiche JD. 2015. Src-family-tyrosine kinase Lyn is critical for TLR2-mediated NF-kappaB activation through the PI 3-kinase signaling pathway. Innate Immun 21:685–697. doi: 10.1177/1753425915586075. [DOI] [PubMed] [Google Scholar]
- 31.Klarstrom Engstrom K, Brommesson C, Kalvegren H, Bengtsson T. 2014. Toll like receptor 2/1 mediated platelet adhesion and activation on bacterial mimetic surfaces is dependent on src/Syk-signaling and purinergic receptor P2X1 and P2Y12 activation. Biointerphases 9:041003. doi: 10.1116/1.4901135. [DOI] [PubMed] [Google Scholar]
- 32.Sharma N, Akhade AS, Qadri A. 2016. Src kinases central to T-cell receptor signaling regulate TLR-activated innate immune responses from human T cells. Innate Immun 22:238–244. doi: 10.1177/1753425916632305. [DOI] [PubMed] [Google Scholar]
- 33.Wu J, Weening EH, Faske JB, Hook M, Skare JT. 2011. Invasion of eukaryotic cells by Borrelia burgdorferi requires beta(1) integrins and Src kinase activity. Infect Immun 79:1338–1348. doi: 10.1128/IAI.01188-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Oosting M, Buffen K, Cheng SC, Verschueren IC, Koentgen F, van de Veerdonk FL, Netea MG, Joosten LA. 2015. Borrelia-induced cytokine production is mediated by spleen tyrosine kinase (Syk) but is Dectin-1 and Dectin-2 independent. Cytokine 76:465–472. doi: 10.1016/j.cyto.2015.08.005. [DOI] [PubMed] [Google Scholar]
- 35.Talkington J, Nickell SP. 2001. Role of Fc gamma receptors in triggering host cell activation and cytokine release by Borrelia burgdorferi. Infect Immun 69:413–419. doi: 10.1128/IAI.69.1.413-419.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Cinco M, Cini B, Murgia R, Presani G, Prodan M, Perticarari S. 2001. Evidence of involvement of the mannose receptor in adhesion of Borrelia burgdorferi to monocyte/macrophages. Infect Immun 69:2743–2747. doi: 10.1128/IAI.69.4.2743-2747.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Coburn J, Magoun L, Bodary SC, Leong JM. 1998. Integrins alpha(v)beta3 and alpha5beta1 mediate attachment of Lyme disease spirochetes to human cells. Infect Immun 66:1946–1952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Jakus Z, Fodor S, Abram CL, Lowell CA, Mocsai A. 2007. Immunoreceptor-like signaling by beta 2 and beta 3 integrins. Trends Cell Biol 17:493–501. doi: 10.1016/j.tcb.2007.09.001. [DOI] [PubMed] [Google Scholar]
- 39.Legate KR, Fassler R. 2009. Mechanisms that regulate adaptor binding to beta-integrin cytoplasmic tails. J Cell Sci 122:187–198. doi: 10.1242/jcs.041624. [DOI] [PubMed] [Google Scholar]
- 40.Petnicki-Ocwieja T, Kern A, Killpack TL, Bunnell SC, Hu LT. 2015. Adaptor protein-3-mediated trafficking of TLR2 ligands controls specificity of inflammatory responses but not adaptor complex assembly. J Immunol 195:4331–4340. doi: 10.4049/jimmunol.1501268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Henault J, Martinez J, Riggs JM, Tian J, Mehta P, Clarke L, Sasai M, Latz E, Brinkmann MM, Iwasaki A, Coyle AJ, Kolbeck R, Green DR, Sanjuan MA. 2012. Noncanonical autophagy is required for type I interferon secretion in response to DNA-immune complexes. Immunity 37:986–997. doi: 10.1016/j.immuni.2012.09.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Mantegazza AR, Guttentag SH, El-Benna J, Sasai M, Iwasaki A, Shen H, Laufer TM, Marks MS. 2012. Adaptor protein-3 in dendritic cells facilitates phagosomal Toll-like receptor signaling and antigen presentation to CD4(+) T cells. Immunity 36:782–794. doi: 10.1016/j.immuni.2012.02.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Sasai M, Linehan MM, Iwasaki A. 2010. Bifurcation of Toll-like receptor 9 signaling by adaptor protein 3. Science 329:1530–1534. doi: 10.1126/science.1187029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Bonifacino JS, Traub LM. 2003. Signals for sorting of transmembrane proteins to endosomes and lysosomes. Annu Rev Biochem 72:395–447. doi: 10.1146/annurev.biochem.72.121801.161800. [DOI] [PubMed] [Google Scholar]
- 45.Di Guglielmo GM, Le Roy C, Goodfellow AF, Wrana JL. 2003. Distinct endocytic pathways regulate TGF-beta receptor signalling and turnover. Nat Cell Biol 5:410–421. doi: 10.1038/ncb975. [DOI] [PubMed] [Google Scholar]
- 46.Dahl M, Bauer AK, Arredouani M, Soininen R, Tryggvason K, Kleeberger SR, Kobzik L. 2007. Protection against inhaled oxidants through scavenging of oxidized lipids by macrophage receptors MARCO and SR-AI/II. J Clin Invest 117:757–764. doi: 10.1172/JCI29968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Jozefowski S, Sulahian TH, Arredouani M, Kobzik L. 2006. Role of scavenger receptor MARCO in macrophage responses to CpG oligodeoxynucleotides. J Leukoc Biol 80:870–879. doi: 10.1189/jlb.0705357. [DOI] [PubMed] [Google Scholar]
- 48.Tjota MY, Hrusch CL, Blaine KM, Williams JW, Barrett NA, Sperling AI. 2014. Signaling through FcRgamma-associated receptors on dendritic cells drives IL-33-dependent TH2-type responses. J Allergy Clin Immunol 134:706.e8–713.e8. doi: 10.1016/j.jaci.2014.06.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Lopez-Contreras L, Hernandez-Ramirez VI, Flores-Garcia Y, Chavez-Munguia B, Talamas-Rohana P. 2013. Src and PI3 K inhibitors affect the virulence factors of Entamoeba histolytica. Parasitology 140:202–209. doi: 10.1017/S0031182012001540. [DOI] [PubMed] [Google Scholar]
- 50.Majeed M, Caveggion E, Lowell CA, Berton G. 2001. Role of Src kinases and Syk in Fcgamma receptor-mediated phagocytosis and phagosome-lysosome fusion. J Leukoc Biol 70:801–811. [PubMed] [Google Scholar]