

Abstract
Here we describe a staged, behavioral testing approach that can be used to screen for compounds that exhibit in vivo efficacy on cognitive and functional motor behaviors in transgenic mouse models of β-amyloidosis and tauopathy. The paradigm includes tests for spontaneous alternation in a Y-maze, novel object recognition, and limb clasping. These tests were selected because they: 1) interrogate function of cognitive or motor domains and the correlate neural circuitry relevant to the human disease state, 2) have clearly defined endpoints, 3) have easily implementable quality control checks, 4) can be run in a moderate throughput format, and 5) require little intervention by the investigator. These methods are designed for investigators looking to screen compounds for activity in short-term and working memory tasks, or functional motor behaviors associated with Alzheimer's disease mouse models. The methods described here use behavioral tests that engage a number of different brain regions including hippocampus and various cortical areas. Investigators that desire cognitive tests that specifically assess cognition mediated by a single brain region could use these techniques to supplement other behavioral tests.
Keywords: Medicine, Issue 123, Rodent behavior, spontaneous alternation, novel object recognition, limb clasping, amyloid-β, tau, Alzheimer's disease, neurodegeneration
Introduction
Alzheimer's disease (AD) is a progressive neurodegenerative disorder resulting in debilitating cognitive decline that affects approximately 44 million people worldwide. Currently there are no available treatments for AD that are disease-modifying, emphasizing the urgent need for preclinical discovery of novel therapeutic strategies for this disease. A number of different transgenic mouse models have been created that recapitulate various aspects of AD1,2, including deficits in cognitive domains disrupted in patients3. These mouse models represent a useful tool for facilitating efficient screening in vivo.
When assessing a compound for potential in vivo efficacy, a staged approach must be taken that screens for efficacy in appropriate cognitive domains and also monitors behaviors that could influence the specific endpoints used to assess cognition. Many transgenic mouse models of AD exhibit hyperactivity and other behaviors that may interfere with a particular cognitive test, and prohibit its use in drug screening4. Moreover, for an approach to be implemented in a drug screening environment, the particular tests used should sustain at least a moderate throughput, have clearly defined endpoints, and a procedure that requires minimal intervention by the investigators. Using these criteria, behavioral screens can be implemented that exhibit the reproducibility, low intra- and inter-assay variance and effect sizes needed for compound screening. Detailed here are the methods we have employed to screen for compounds effective in mitigating the cognitive and motor phenotypes present in transgenic mouse models of β-amyloidosis and tauopathy5,6. The methods described are adapted from commonly used behavioral paradigms reported in the literature7, with specific optimizations and quality control checks so that they may be used in transgenic mouse models relevant to AD. This protocol can be used with a variety of data acquisition and analysis systems and assumes that the investigator has a working knowledge of the associated software.
Protocol
The methods detailed in this publication were reviewed by the institutional animal care and use committee (IACUC) at Hilltop Laboratory Animals to ensure proper care, use, and humane treatment of animals in compliance with applicable federal, state, and local laws and regulations, such as the federal Animal Welfare Regulations, or AWRs (CFR 1985), and Public Health Service Policy on Humane Care and Use of Laboratory Animals, or PHS Policy (PHS 1996).
1. General Guidelines for all Behavioral Assessment
Before any animal handling, cover existing cage cards with a new cage card that indicates only the unique, blinded animal identifier. NOTE: Investigators that handle mice during routine compound/placebo dosing are not permitted to handle mice for behavioral assessment.
Dim or turn off the overhead lights and adjust the lighting such that illumination at the floor of the arena or maze is 30-35 lux.
For studies that span several weeks, record body weights weekly as an indirect measure of overall health. NOTE: Additional cage-side checks for coat quality, posture, gait and spontaneous locomotion can be included if more robust health checks are warranted.
2. Habituating Mice to Handling by Investigators
Two days prior to any behavioral test, habituate the mice to handling. Remove the cage from the cage rack and place on a level surface.
Remove the lid from the cage. Handle the mouse exactly as it would be handled during performance of the upcoming behavioral test. Place the mouse into a cupped hand above the home cage.
Measure the latency to jump from the investigator's hand back into the home cage. Hold mice for a maximum of 5 s. NOTE: Mice that exhibit latencies of ≥2 s are deemed "habituated". Mice that exhibit latencies <2 s during the first trial undergo 2 additional habituation sessions that day.
Have the mice undergo 2 consecutive days of handling habituation. Note any mouse that is not habituated by the end of the 2nd day.
3. Assessing Spatial Working Memory by Measuring Spontaneous Alternation in a Y-maze8
Prior to initial use, thoroughly clean the Y-maze with an unscented bleach germicidal wipe, 70% EtOH, followed by dH2O9. Clearly designate the arms of the maze as 'A', 'B' & 'C' or other comparable unique identifiers.
Prior to the start of a testing session, set the data acquisition system or video cameras and set up proper tracking of mice in the maze. Calibrate the distance in the maze using captured video images of a ruler or other object of known length. NOTE: The behavioral methods in this procedure will work with a variety of data acquisition systems and the authors assume anyone performing this procedure is proficient in the use of their chosen data acquisition system. Power analyses indicate that sample sizes of 10-15 mice per group are required for a β ≤0.2.
Remove the cage from the rack and gently place it on a table in close proximity to the Y-maze. Remove the mouse from its home cage and gently place it into one arm of the Y-maze, facing the center. Have the investigator step far enough away from the maze so that the mouse cannot see the investigator.
Activate the data/video acquisition system immediately after placement of the mouse into the maze.
Press play and record the spontaneous behavior for each mouse for a period of 10 min. Once a session is complete, gently place the mouse back into its home cage and return the cage to the rack.
Clean the maze thoroughly between each session with an unscented bleach germicidal wipe, 70% EtOH, followed by dH2O. Repeat from step 3.4 to assess all mice.
Once all mice have completed exploration of the Y-maze, analyze the data from the acquisition system or manually score the videos of the sessions. An arm entry occurs when all 4 paws of the mouse cross the threshold of the central zone and into the arm and the animal's snout is oriented toward the end of the arm. NOTE: Endpoints that will be analyzed include: total distance traveled in the maze, total distance traveled within each arm (including the central zone), total time spent in each arm (including the central zone), total number of arm entries, number of entries made into each arm, and a sequential list of arms entered to assess number of alternations made.
A spontaneous alternation occurs when a mouse enters a different arm of the maze in each of 3 consecutive arm entries. Spontaneous alternation % is then calculated with the following formula.
 NOTE: For example, if the order of arm entry was: ABCCBABCABC, investigator would score a total of 6 spontaneous alternations (in order: ABC, CBA, ABC, BCA, CAB, ABC). With a total of 11 arm entries, the spontaneous alternation % would be 67%.
NOTE: For example, if the order of arm entry was: ABCCBABCABC, investigator would score a total of 6 spontaneous alternations (in order: ABC, CBA, ABC, BCA, CAB, ABC). With a total of 11 arm entries, the spontaneous alternation % would be 67%.- Make the following quality control checks to ensure the data represent an unbiased assessment of spontaneous alternation.
- Perform a Pearson's correlation of spontaneous alternation % to both total distance traveled and number of arm entries made. NOTE: If there is a significant correlation of spontaneous alternation % to either parameter, then data should be further scrutinized due to potential influence of hyperdynamic locomotion on the apparent cognitive endpoint10.
- Analyze number of entries made into each arm with a 1-way ANOVA test. NOTE: If this analysis is significant then this would indicate presence of cues in the environment that attracted mice to a particular region of the maze.
4. Assessment of Intermediate-term Recognition Memory by Measuring Novel Object Recognition11,12,13
For each phase of this test, thoroughly clean the open field arena with an unscented bleach germicidal wipe, 70% EtOH followed by dH2O prior to initial use.
- One day prior to object exposure, habituate the mice to the open field arena.
- Prior to the start of the habituation session, set up the data acquisition system or video cameras and confirm proper tracking of mice in the maze. Calibrate the distance in the arena using captured video images of a ruler or other object of known length. Mark the corners of the arena in the software to permit scoring of positional biases. NOTE: The behavioral methods in this procedure will work with a variety of data acquisition systems and the authors assume anyone performing this procedure is proficient in the use of their chosen data acquisition system. Power analyses indicate that sample sizes of 15-20 mice per group are required for a β ≤0.2.
- Remove the cage from the rack and gently place on a table in close proximity to the arena.
- Remove the mouse from the home cage and gently place the mouse in the center of the arena. Turn on the tracking software and/or video recording system immediately after placing the mouse into the arena.
- Allow mice to freely explore the arena for 30 min. NOTE: During this period, investigators will not disturb the mice.
- After the habituation session, place mice back into their home cage and clean the arena thoroughly with an unscented bleach germicidal wipe, 70% EtOH followed by dH2O.
- Repeat from step 4.2.2 until all mice have been habituated to the arena.
- After all mice have been habituated to the arena, analyze the video. NOTE: Endpoints to analyze include total distance traveled in the arena and time spent near each corner. If relevant to the mouse model, stereotyped behaviors are included in these analyses (i.e., myoclonic corner jumping, circling, etc.). Mice exhibiting biases in time spent in particular regions of the arena are excluded from further experimentation as this will influence object exploration. NOTE: The first phase of novel object recognition involves familiarizing mice to an object. Herein this portion of the novel object recognition procedure will be referred to as the Sample phase.
- Prior to the start of a sample phase session, place objects into the arena and fix them to the floor with a mounting putty so that animals cannot move the objects. Align two identical objects to a particular wall with enough distance between the walls and objects so that the mice can freely explore the objects from all angles.
- Set up the data acquisition system or video cameras and confirm proper tracking of mice and objects in the maze. Calibrate the distances in the arena using captured video images of a ruler or other object of known length.
- Mark the corners of the arena in the software to permit scoring of positional biases. Mark objects in software and track their exploratory behavior separately for each object (i.e., "Object A" and "Object B").
- Remove the cage from the rack and gently place it on a table in close proximity to the arena.
- Remove the mouse from the home cage and gently place it into the center of the arena, facing the objects.
- Allow the mouse to freely explore the objects for 15 min. During this period do not disturb the mice.
- At the end of the session, gently place the mouse back into its home cage. Clean the arena and objects with 70% EtOH and dH2O. Place these objects back into the arena.
- Repeat step 4.2.11 until all mice are familiarized to an object.
- Once all mice have been familiarized to an object, analyze the videos. NOTE: Object explorations are counted once the following criteria have been met: the mouse is oriented toward the object, the snout is within 2 cm of the object, the midpoint of the animal's body is beyond 2 cm from the object, and the previous criteria have been fulfilled for at least 1 s. Additionally, if an animal has satisfied the exploration criteria but exhibits immobility for > 10 s then the exploratory bout is deemed finished.
- Calculate an object bias score for each mouse as follows.
 NOTE: Mice exhibiting an object bias score below 20% or above 80% are excluded from further experimentation.
NOTE: Mice exhibiting an object bias score below 20% or above 80% are excluded from further experimentation.
- The final phase of novel object recognition involves assessing exploratory behavior directed toward both a novel and familiar object in the environment, referred to here as the Test phase. This phase is performed 2-3 h after completion of sample phase.
- Prior to the start of a test phase session, place objects into the arena and fix them to the floor so that the animals cannot move the objects.
- Place the objects in the same position in the arena relative to the sample phase13.
- Balance the relative position of novel and familiar objects across genotypes and treatment groups.
- Ensure there is enough distance between the walls and objects so that the mice can freely explore the objects from all angles.
- Setup the data acquisition system and/or video cameras. Confirm proper tracking of mice and objects in the maze. Calibrate distances in the arena using captured video images of a ruler or other object of known length.
- Mark corners of the arena in the software to permit scoring of positional biases. Mark objects in software and track exploratory behavior for each object individually (i.e., "Novel" and "Familiar").
- Remove the cage from the rack and gently place it on a table in close proximity to the arena.
- Gently place animals into the center of the arena, facing the objects. Record mice freely exploring objects for 10 min.
- At the end of the test session, remove mice from the arena and place mice back into their home cage. Thoroughly clean arena and objects with an unscented bleach germicidal wipe, 70% EtOH and dH2O after each session.
- Repeat from step 4.4.3 until all animals have been assessed.
- Once object exploration is measured for all mice, videos are analyzed. NOTE: Object explorations are counted once the following criteria have been met: the mouse is oriented toward the object, the snout is within 2 cm of the object, the midpoint of the animal's body is beyond 2 cm from the object, and the previous criteria have been fulfilled for at least 1 s. Additionally, if an animal has satisfied the exploration criteria but exhibits immobility for > 10 s then the exploratory bout is deemed finished.
- Assess novel object recognition by comparing time spent exploring the novel to familiar object. Three methods are commonly reported in the literature.
- Analyze raw time spent exploring both novel and familiar objects using a repeated measure test. This method is best used when genotype and/or treatment do not affect total exploration time.
- Calculate novelty preference, using the equation:
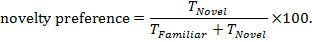 NOTE: This provides the percentage of time spent exploring the novel object relative to the total time exploring objects. Values range from 0% (no exploration of novel object) to 100% (exploration only of the novel object), with a value of 50% indicating equal time spent exploring novel and familiar objects.
NOTE: This provides the percentage of time spent exploring the novel object relative to the total time exploring objects. Values range from 0% (no exploration of novel object) to 100% (exploration only of the novel object), with a value of 50% indicating equal time spent exploring novel and familiar objects. - Calculate discrimination index11, using the equation:
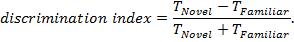 NOTE: This yields the difference in time spent exploring the novel and familiar objects relative to the total time spent exploring objects. Values range from -1 (exploration only of the familiar object) to +1 (exploration only of the novel object, with a value of 0 indicating equal time spent exploring novel and familiar objects.
NOTE: This yields the difference in time spent exploring the novel and familiar objects relative to the total time spent exploring objects. Values range from -1 (exploration only of the familiar object) to +1 (exploration only of the novel object, with a value of 0 indicating equal time spent exploring novel and familiar objects.
- Remove animals that do not participate in the test session due to hyperdynamic locomotion or other stereotypies, from consideration11. NOTE: Criteria used for removal must be objective and determined a priori for the mouse model (i.e., <5th percentile for total exploration time and either >100 average turn angle during test session or >50th percentile time exhibiting myoclonic corner jumping).
5. Assessment of Corticospinal Function in Mice with Limb Clasping14
Video document the entire session. Record the video using a portable hand-held device (i.e., smartphone or equivalent). NOTE: Power analyses indicate that sample sizes of 10-15 mice per group are required for a β ≤0.2.
Remove the home cage from the rack and place it on a table. Document the animal ID in the video prior to the next step.
Gently remove the mouse from its cage and suspend by the tail for 5-10 s. The video must record the animal's hind and forepaws while suspended.
After capturing at least 5 s of video, place the mouse back into its home cage, and return the cage to the rack.
Clean the table. Repeat from step 5.2 until all mice have been recorded.
- Score limb clasping from videos of mice suspended by their tail on a scale from 0-4 (see Table 1 for description of scoring). Inspect videos of suspended mice and then assign a score based on the following criteria.
- No limb clasping. Normal escape extension. One hind limb exhibits incomplete splay and loss of mobility. Toes exhibit normal splay.
- Both hind limbs exhibit incomplete splay and loss of mobility. Toes exhibit normal splay.
- Both hind limbs exhibit clasping with curled toes and immobility.
- Forelimbs and hind limbs exhibit clasping and are crossed, curled toes and immobility.
- All mice are scored by 2 independent investigators. Any mouse where the 2 scores differ by more than 1 point is rescored once again.
- Scores that differ are averaged.
 Table 1: Description of Limb Clasping Scores.
Table 1: Description of Limb Clasping Scores.
Representative Results
Aged Tg2576 mice exhibit robust deficits in spontaneous alternations made within a Y-maze15,16, a phenotype that can be replicated using the methods detailed here (Figure 1A). While a trend for increased arm entries is observed in these mice (Figure 1B), the hyperactivity observed in this line of mice did not affect the spontaneous alternation rate (Figure 1C). In contrast, aged rTg4510 mice appear to exhibit increased spontaneous alternation when placed into a Y-maze (Figure 1D). This is due to extreme hyperactivity (Figure 1E) and stereotypy10, which significantly interfere with measurement of spontaneous alternation (Figure 1F). When initially assessing mice in this task, it is critical to ensure that arm entries and/or distance traveled is not significantly correlated with the spontaneous alternation rate.
Prior to assessment of novel object recognition, mice are habituated to the arena where the test will be performed. During the habituation, hyperactivity (Figure 2A) and other stereotyped behaviors relevant to the mouse model can be assessed. During the sample phase, it is critical to measure exploration of each object separately so that mice that exhibit significant biases in exploratory behavior can be excluded from further assessment (Figure 2B, open circles). Novel object recognition is assessed by comparing exploration of a familiar and novel object, and is commonly analyzed in three different ways. If total exploratory time is comparable across genotypes and/or treatment groups, then raw time exploring each object and an appropriate repeated measures tests can be used to determine if there were differences in novel object recognition (Figure 2C). If a particular mouse strain exhibits differences in total exploratory time, novel object recognition can be assessed using either novelty preference (Figure 2D) or discrimination index (Figure 2E).
Limb clasping is a functional motor test that quantifies deficits in corticospinal function. Limb clasping, which is not a cognitive measure, is observed in several transgenic tau mouse models6,17,18,19 and recapitulates some of the functional motor deficits observed in late-stage AD patients. Suspension of mice by the tail elicits an escape response (Figure 3A, "0"). Deficits in the ability to splay the hind limbs and extend the toes are scored based on their severity on a scale from 0-4 (Figure 3A). Using the procedure outlined herein, one can observe significant limb clasping in rTg4510 mice (Figure 3B).
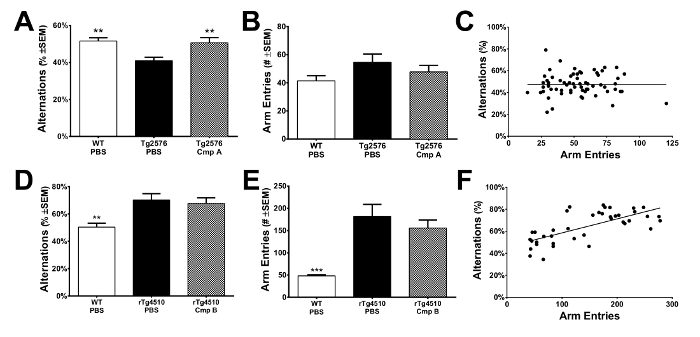 Figure 1: Spontaneous Alternation in the Y-maze. (A) When placed into a Y-maze, mice adopt a lose-shift search strategy that results in a pattern of exploration whereby each arm is explored just once for every 3 arm entries. Aged Tg2576 mice exhibit a significant deficit in spontaneous alternation. Using the procedures outlined in this method, a significant restoration of spontaneous alternation was observed after treatment with a proprietary compound. Data was analyzed using 1-way ANOVA and post-hoc comparisons to Tg-PBS were performed using Dunnett's test. **p <0.01. Error bars indicate SEM. (B) Number of arm entries was not significantly different across any of the groups monitored in this experiment. Data was analyzed using 1-way ANOVA test. Error bars indicate SEM. (C) There was no correlation between spontaneous alternation and the number of arm entries made, indicating that any differences in spontaneous locomotor activity did not impact quantification of spontaneous alternation. Correlation test was performed using Pearson's correlation analysis. (D) When placed into a Y-maze, rTg4510 mice (6 month) appear to exhibit significantly more spontaneous alternation relative to littermate WT mice. Data was analyzed by 1-way ANOVA and post-hoc comparisons to Tg-PBS were performed using Dunnett's test. **p <0.01. Error bars indicate SEM. (E) rTg4510 mice made significantly more arm entries due to their extreme hyperdynamic locomotion. Data was analyzed by 1-way ANOVA and post-hoc comparisons to Tg-PBS were performed using Dunnett's test. ***p <0.001. Error bars indicate SEM. (F) Spontaneous alternation behavior significantly correlated with arm entries, indicating the hyperdynamic locomotor phenotype obscured true spontaneous alternation. Correlation test was performed using Pearson's correlation analysis (r = 0.7, p <0.0001). Please click here to view a larger version of this figure.
Figure 1: Spontaneous Alternation in the Y-maze. (A) When placed into a Y-maze, mice adopt a lose-shift search strategy that results in a pattern of exploration whereby each arm is explored just once for every 3 arm entries. Aged Tg2576 mice exhibit a significant deficit in spontaneous alternation. Using the procedures outlined in this method, a significant restoration of spontaneous alternation was observed after treatment with a proprietary compound. Data was analyzed using 1-way ANOVA and post-hoc comparisons to Tg-PBS were performed using Dunnett's test. **p <0.01. Error bars indicate SEM. (B) Number of arm entries was not significantly different across any of the groups monitored in this experiment. Data was analyzed using 1-way ANOVA test. Error bars indicate SEM. (C) There was no correlation between spontaneous alternation and the number of arm entries made, indicating that any differences in spontaneous locomotor activity did not impact quantification of spontaneous alternation. Correlation test was performed using Pearson's correlation analysis. (D) When placed into a Y-maze, rTg4510 mice (6 month) appear to exhibit significantly more spontaneous alternation relative to littermate WT mice. Data was analyzed by 1-way ANOVA and post-hoc comparisons to Tg-PBS were performed using Dunnett's test. **p <0.01. Error bars indicate SEM. (E) rTg4510 mice made significantly more arm entries due to their extreme hyperdynamic locomotion. Data was analyzed by 1-way ANOVA and post-hoc comparisons to Tg-PBS were performed using Dunnett's test. ***p <0.001. Error bars indicate SEM. (F) Spontaneous alternation behavior significantly correlated with arm entries, indicating the hyperdynamic locomotor phenotype obscured true spontaneous alternation. Correlation test was performed using Pearson's correlation analysis (r = 0.7, p <0.0001). Please click here to view a larger version of this figure.
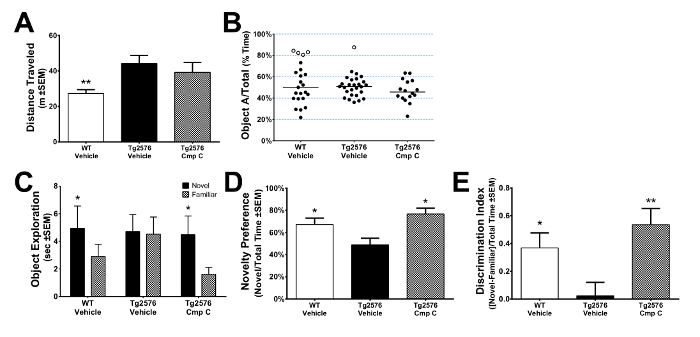 Figure 2: Novel Object Recognition. (A) Arena habituation permits measurement of spontaneous locomotion and other stereotyped behaviors relevant to a particular mouse model. Here, aged Tg2576 mice (22 month) exhibit significantly more spontaneous locomotion relative to littermate WT mice. Data was analyzed by 1-way ANOVA and post-hoc comparisons to Tg-Veh were performed using Dunnett's test. **p <0.01. Error bars indicate SEM. (B) During sample phase, exploration of two identical objects was tracked separately. Mice that exhibit large biases toward exploration of one of the two objects (open circles) were excluded from test phase. (C-E) Novel object recognition was assessed by measuring the exploration of a novel and familiar object. Novel object recognition was assessed using (C) raw exploration time, (D) novelty preference or (E) discrimination index. Data in panel C were analyzed using a 2-way ANOVA with repeated measures and pairwise comparisons were made using Sidak's test. Data in panels D-E were analyzed with a 1-way ANOVA and post-hoc comparisons to Tg-Veh were made using Dunnett's test. *p <0.05, **p <0.01. Error bars indicate SEM. Please click here to view a larger version of this figure.
Figure 2: Novel Object Recognition. (A) Arena habituation permits measurement of spontaneous locomotion and other stereotyped behaviors relevant to a particular mouse model. Here, aged Tg2576 mice (22 month) exhibit significantly more spontaneous locomotion relative to littermate WT mice. Data was analyzed by 1-way ANOVA and post-hoc comparisons to Tg-Veh were performed using Dunnett's test. **p <0.01. Error bars indicate SEM. (B) During sample phase, exploration of two identical objects was tracked separately. Mice that exhibit large biases toward exploration of one of the two objects (open circles) were excluded from test phase. (C-E) Novel object recognition was assessed by measuring the exploration of a novel and familiar object. Novel object recognition was assessed using (C) raw exploration time, (D) novelty preference or (E) discrimination index. Data in panel C were analyzed using a 2-way ANOVA with repeated measures and pairwise comparisons were made using Sidak's test. Data in panels D-E were analyzed with a 1-way ANOVA and post-hoc comparisons to Tg-Veh were made using Dunnett's test. *p <0.05, **p <0.01. Error bars indicate SEM. Please click here to view a larger version of this figure.
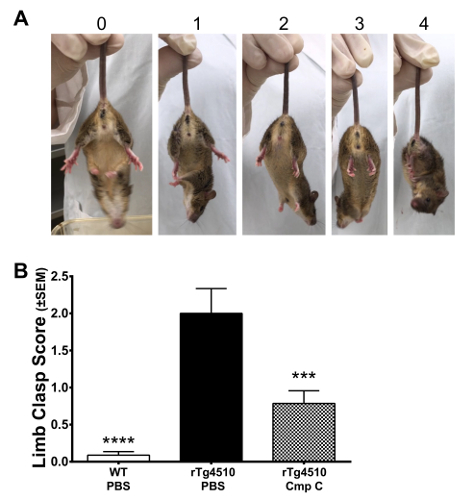 Figure 3: Limb Clasping. (A) Representative images of mice exhibiting various degrees of limb clasping as described in Table 1. (B) rTg4510 mice (6 month) exhibit significant limb clasping relative to littermate WT mice as scored using these methods. Data were analyzed using a 1-way ANOVA and post-hoc comparisons to Tg-PBS were performed using Dunnett's test. ***p <0.001, ****p <0.0001. Error bars indicate SEM. Please click here to view a larger version of this figure.
Figure 3: Limb Clasping. (A) Representative images of mice exhibiting various degrees of limb clasping as described in Table 1. (B) rTg4510 mice (6 month) exhibit significant limb clasping relative to littermate WT mice as scored using these methods. Data were analyzed using a 1-way ANOVA and post-hoc comparisons to Tg-PBS were performed using Dunnett's test. ***p <0.001, ****p <0.0001. Error bars indicate SEM. Please click here to view a larger version of this figure.
Discussion
Significance of the Technique with Respect to Existing Methods This procedure has been designed to screen for in vivo activity of compounds in transgenic mouse models of β-amyloidosis and tauopathy. The staged approach employed here ensures detection of efficacious compounds in cognitive domains relevant to AD3. Moreover, the approach detailed here uses behavioral tests that have clearly defined endpoints, easily implementable quality control checks, can be run in a moderate throughput format, and require little intervention from the investigator. These characteristics result in assays that exhibit good reproducibility within animals and across cohorts, which results in low intra- and inter-assay variance and effect sizes (2 ≤f ≤6) that are robust enough to support behavioral profiling in a drug discovery environment.
Critical Steps within the Protocol Many mouse models in use for AD drug discovery exhibit behaviors consistent with heightened anxiety and aggression. This makes handling habituation essential for performing any of the behavioral tests described here. As these tests rely on unmotivated behaviors, rough handling by the investigator due to a hyperactive and anxious or aggressive mouse can significantly influence performance. Heightened anxiety could result in failure to perform the task, reducing the overall power of the test. Moreover, light levels in the arena are essential for facilitating the spontaneous locomotion needed for each test. Bright light tends to increase anxiety and suppress locomotion in rodents, therefore care should be taken to adjust ambient light levels to 30-35 lux in the arena.
Another critical aspect of the procedure is minimization of strong environmental cues that would interfere with an animal's ability to perform the tasks. Cleaning of the arena and objects in between runs is essential as mice are attracted to novel scents in the environment. Failure to thoroughly clean the arena and objects could result in skewing spontaneous activity of the mouse and masking true cognitive performance. Investigators should also minimize use of personal hygiene products and colognes/perfumes when performing these procedures. Lastly, rodents exhibit robust diurnal and circadian changes in many overt behaviors20 including learning and memory21. Therefore, to minimize variance due to diurnal rhythms in basal behaviors and cognitive performance, all tests should be done at the same time of day across cohorts and studies.
Further, specifically with regard to novel object recognition, the delay interval between sample and test phase, and the selection and placement of objects in the environment are critical parameters. Memory exists in 3 distinct forms: short term memory (STM), intermediate term memory (ITM) and long term memory (LTM)22,23. Changing the interval between sample and test phases from minutes (STM) to hours (ITM) or days (LTM) will change the type of memory tested by the procedure12. Moreover, prior to running the novel object recognition test, many objects should be screened in a test cohort of mice for potential biases in exploration. An object that is excessively attractive or repulsive to the test cohort cannot be used when assessing novel object recognition. Ideally all objects that will be employed in the test, when placed into an arena, will elicit equal exploration times from a naïve cohort of mice. Inadequate testing and optimization of objects can significantly reduce the power of novel object recognition.
Modifications and Troubleshooting There are several factors that could increase the apparent variability in the cognitive tests described here. Many mouse models of AD exhibit hyperdynamic locomotion3 which can mask or alter behaviors measured as the cognitive endpoint. Moreover, there is growing evidence that sex24,25,26 and even maternal genotype27 can influence development and progression of neuropathology and cognitive phenotypes in AD mouse models. Unexpected variability or failure to implement a behavioral task could be due to any of these factors. When first implementing a particular behavioral test, results should always be stratified by sex, age and if applicable, maternal genotype. Furthermore, the quality checks outlined in this procedure should always be performed to ensure that hyperactivity or other stereotyped behaviors are not interfering with quantification of cognitive endpoints.
Environment can also influence the spontaneous exploratory behavior of rodents. Scents or sounds that are undetectable to researchers could attract or repel mice, skewing results of cognitive tests that rely on spontaneous behavior. When initially establishing Y-maze or novel object recognition, performance of the control measures to ensure that there are no positional biases in exploration of objects and/or the environment is essential. If positional biases are observed then investigators must thoroughly scrutinize the environment and potentially adjust lighting, arena placement, location of testing room relative to other rooms in the facility (i.e., not near a high traffick area or heavy equipment) and arena cleaning procedures.
Habituation to the testing environment is key to achieving optimal performance in the novel object recognition test. For example, low total exploration times may be due to inadequate habituation. As an alternative to the procedures outlined here for handling (Section 2) and arena (Section 4.2) habituation, habituation to handling and the test environment can be performed as 3, 5 min sessions per day for 2 consecutive days.
Limitations of the Technique As with any procedure, these behavioral tests have limitations. These procedures have been employed because they test function of various cortical regions and hippocampus. If the mouse model does not exhibit functional deficits in brain regions probed by these tests, then these techniques will not be useful. Moreover, we have chosen cognitive tests that probe short-term memory. If the mechanism of action of the compound under preclinical assessment is not expected to affect short-term memory then these procedures should be modified accordingly (i.e., increasing the sample-test phase interval to test long-term memory). Lastly, these tests use unmotivated behaviors. Therefore, if a mouse model is excessively hyperactive or displays other stereotyped behaviors that prevent exploration of the environment then these procedures might not be optimal. As an alternative, one could use fear conditioning for Tg2576 or other β-amyloidosis mouse models, or the spatial water maze for rTg4510 or other mouse models of tauopathy3.
Future Applications Once these procedures have been successfully adopted in the lab, several modifications or extensions can be made to assess additional cognitive and functional motor measures. For example, changing the novel object recognition task to determine if a mouse can recognize a change in placement of an object13. Alternatively, instead of using objects, one could use other mice and implement a test of social recognition. With respect to limb clasping and motor function, one could supplement that test with the wire hang and/or grip strength tests. The tests detailed in this method form a solid base to screen for compounds that have in vivo efficacy in translational mouse models for AD, and can be adapted or modified in many ways to best interrogate a particular mouse model or meet the needs of a unique drug discovery program.
Disclosures
J.M. Levenson is employed by Proclara Biosciences, Inc. C. Miedel, J. Patton, A. Miedel, and E. Miedel are employed by Hilltop Laboratory Animals.
Acknowledgments
The authors have no acknowledgements.
References
- Elder GA, Gama Sosa MA, De Gasperi R. Transgenic mouse models of Alzheimer's disease. Mt Sinai J Med. 2010;77(1):69–81. doi: 10.1002/msj.20159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Onos KD, Sukoff Rizzo SJ, Howell GR, Sasner M. Toward more predictive genetic mouse models of Alzheimer's disease. Brain Res Bull. 2016;122:1–11. doi: 10.1016/j.brainresbull.2015.12.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Webster SJ, Bachstetter AD, Nelson PT, Schmitt FA, Van Eldik LJ. Using mice to model Alzheimer's dementia: an overview of the clinical disease and the preclinical behavioral changes in 10 mouse models. Front Genet. 2014;5:88. doi: 10.3389/fgene.2014.00088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodgers SP, Born HA, Das P, Jankowsky JL. Transgenic APP expression during postnatal development causes persistent locomotor hyperactivity in the adult. Mol Neurodegener. 2012;7:28. doi: 10.1186/1750-1326-7-28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hsiao K, et al. Correlative memory deficits, Abeta elevation, and amyloid plaques in transgenic mice. Science. 1996;274(5284):99–102. doi: 10.1126/science.274.5284.99. [DOI] [PubMed] [Google Scholar]
- Santacruz K, et al. Tau suppression in a neurodegenerative mouse model improves memory function. Science. 2005;309(5733):476–481. doi: 10.1126/science.1113694. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crawley JN. What's wrong with my mouse? : behavioral phenotyping of transgenic and knockout mice. 2nd edn. Wiley-Interscience; 2007. [Google Scholar]
- Hughes RN. The value of spontaneous alternation behavior (SAB) as a test of retention in pharmacological investigations of memory. Neurosci Biobehav Rev. 2004;28(5):497–505. doi: 10.1016/j.neubiorev.2004.06.006. [DOI] [PubMed] [Google Scholar]
- Rutala WA, Weber DJ. Guideline for disinfection and sterilization in healthcare facilities. Centers for Disease Control. 2008.
- Wes PD, et al. Tau overexpression impacts a neuroinflammation gene expression network perturbed in Alzheimer's disease. PLoS One. 2014;9(8):106050. doi: 10.1371/journal.pone.0106050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ennaceur A, Delacour J. A new one-trial test for neurobiological studies of memory in rats. 1: Behavioral data. Behav Brain Res. 1988;31(1):47–59. doi: 10.1016/0166-4328(88)90157-x. [DOI] [PubMed] [Google Scholar]
- Taglialatela G, Hogan D, Zhang WR, Dineley KT. Intermediate- and long-term recognition memory deficits in Tg2576 mice are reversed with acute calcineurin inhibition. Behav Brain Res. 2009;200(1):95–99. doi: 10.1016/j.bbr.2008.12.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DeVito LM, Eichenbaum H. Distinct contributions of the hippocampus and medial prefrontal cortex to the "what-where-when" components of episodic-like memory in mice. Behav Brain Res. 2010;215(2):318–325. doi: 10.1016/j.bbr.2009.09.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lalonde R, Strazielle C. Brain regions and genes affecting limb-clasping responses. Brain Res Rev. 2011;67(1-2):252–259. doi: 10.1016/j.brainresrev.2011.02.005. [DOI] [PubMed] [Google Scholar]
- King DL, Arendash GW. Behavioral characterization of the Tg2576 transgenic model of Alzheimer's disease through 19 months. Physiol Behav. 2002;75(5):627–642. doi: 10.1016/s0031-9384(02)00639-x. [DOI] [PubMed] [Google Scholar]
- Lalonde R, Lewis TL, Strazielle C, Kim H, Fukuchi K. Transgenic mice expressing the betaAPP695SWE mutation: effects on exploratory activity, anxiety, and motor coordination. Brain Res. 2003;977(1):38–45. doi: 10.1016/s0006-8993(03)02694-5. [DOI] [PubMed] [Google Scholar]
- Lewis J, et al. Neurofibrillary tangles, amyotrophy and progressive motor disturbance in mice expressing mutant (P301L) tau protein. Nat Genet. 2000;25(4):402–405. doi: 10.1038/78078. [DOI] [PubMed] [Google Scholar]
- Spittaels K, et al. Prominent axonopathy in the brain and spinal cord of transgenic mice overexpressing four-repeat human tau protein. Am J Pathol. 1999;155(6):2153–2165. doi: 10.1016/S0002-9440(10)65533-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Terwel D, et al. Changed conformation of mutant Tau-P301L underlies the moribund tauopathy, absent in progressive, nonlethal axonopathy of Tau-4R/2N transgenic mice. J Biol Chem. 2005;280(5):3963–3973. doi: 10.1074/jbc.M409876200. [DOI] [PubMed] [Google Scholar]
- Merrow M, Spoelstra K, Roenneberg T. The circadian cycle: daily rhythms from behaviour to genes. EMBO Rep. 2005;6(10):930–935. doi: 10.1038/sj.embor.7400541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smarr BL, Jennings KJ, Driscoll JR, Kriegsfeld LJ. A time to remember: the role of circadian clocks in learning and memory. Behav Neurosci. 2014;128(3):283–303. doi: 10.1037/a0035963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kandel ER. The molecular biology of memory storage: a dialogue between genes and synapses. Science. 2001;294(5544):1030–1038. doi: 10.1126/science.1067020. [DOI] [PubMed] [Google Scholar]
- Stough S, Shobe JL, Carew TJ. Intermediate-term processes in memory formation. Curr Opin Neurobiol. 2006;16(6):672–678. doi: 10.1016/j.conb.2006.10.009. [DOI] [PubMed] [Google Scholar]
- Stevens LM, Brown RE. Reference and working memory deficits in the 3xTg-AD mouse between 2 and 15-months of age: a cross-sectional study. Behav Brain Res. 2015;278:496–505. doi: 10.1016/j.bbr.2014.10.033. [DOI] [PubMed] [Google Scholar]
- Yue X, et al. Brain estrogen deficiency accelerates Abeta plaque formation in an Alzheimer's disease animal model. Proc Natl Acad Sci U S A. 2005;102(52):19198–19203. doi: 10.1073/pnas.0505203102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McAllister C, et al. Genetic targeting aromatase in male amyloid precursor protein transgenic mice down-regulates beta-secretase (BACE1) and prevents Alzheimer-like pathology and cognitive impairment. J Neurosci. 2010;30(21):7326–7334. doi: 10.1523/JNEUROSCI.1180-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blaney CE, Gunn RK, Stover KR, Brown RE. Maternal genotype influences behavioral development of 3xTg-AD mouse pups. Behav Brain Res. 2013. pp. 40–48. [DOI] [PubMed]