

Abstract
This protocol outlines the procedures for preparing and recording from the isolated whole hippocampus, of WT and transgenic mice, along with recent improvements in methodologies and applications for the study of theta oscillations. A simple characterization of the isolated hippocampal preparation is presented whereby the relationship between internal hippocampal theta oscillators is examined together with the activity of pyramidal cells, and GABAergic interneurons, of the cornu ammonis-1 (CA1) and subiculum (SUB) areas. Overall, we show that the isolated hippocampus is capable of generating intrinsic theta oscillations in vitro and that rhythmicity generated within the hippocampus can be precisely manipulated by optogenetic stimulation of parvalbumin-positive (PV) interneurons. The in vitro isolated hippocampal preparation offers a unique opportunity to use simultaneous field and intracellular patch-clamp recordings from visually-identified neurons to better understand the mechanisms underlying theta rhythm generation.
Keywords: Neuroscience, Issue 126, Hippocampus, neural oscillations, theta rhythm, interneurons, in vitro preparation, electrophysiology, optogenetics.
Introduction
Hippocampal theta oscillations (4 - 12 Hz) are amongst the most predominant forms of rhythmic activity in the mammalian brain and are believed to play key roles in cognitive functions such as processing of spatiotemporal information and formation of episodic memories1,2,3. While several in vivo studies that highlight the relationship of theta-modulated place-cells with spatial navigation and lesion studies, as well as clinical evidence, support the view that hippocampal theta oscillations are involved in memory formation4,5,6, the mechanisms associated with generation of hippocampal theta oscillations are still not fully understood. Early in vivo investigations suggested that theta activity depended mainly on extrinsic oscillators, in particular rhythmic input from afferent brain structures such as the septum and entorhinal cortex7,8,9,10. A role for intrinsic factors - internal connectivity of hippocampal neural networks together with the properties of hippocampal neurons - was also postulated based on in vitro observations11,12,13,14,15,16,17,18. However, apart from a few landmark studies19,20,21, difficulties in developing approaches that could replicate physiologically realistic population activities in simple in vitro slice preparations have, for a long time, delayed more detailed experimental examination of the intrinsic abilities of the hippocampus and related areas to self-generate theta oscillations.
An important downside of the standard in vitro thin-slice experimental setting is that the 3D cellular and synaptic organization of brain structures is usually compromised. This means that many forms of concerted network activities based on spatially distributed cell assemblies, ranging from localized groups (≤1 mm radius) to populations of neurons spread across one or more brain areas (>1 mm), cannot be supported. Given these considerations, a different type of approach was needed to study how theta oscillations emerge in the hippocampus and propagate to related cortical and subcortical output structures.
In recent years, the initial development of the "complete septo-hippocampal" preparation to examine bidirectional interactions of the two structures22, and the ensuing evolution of the "isolated hippocampus" preparation, have revealed that intrinsic theta oscillations occur spontaneously in the hippocampus lacking external rhythmic input23. The value of these approaches lies on the initial insight that the whole functional structure of these regions had to be preserved in order to function as a theta rhythm generator in vitro22.
Protocol
All procedures have been performed according to protocols and guidelines approved by the McGill University Animal Care Committee and the Canadian Council on Animal Care.
1. Acute Hippocampus In Vitro Preparation
NOTE: Isolating the intact hippocampal preparation involves three major steps: (1) Preparation of solutions and equipment, (2) Dissection of the hippocampus and (3) Setting up the fast perfusion rate system necessary for generation of intrinsic theta oscillations. In this protocol, the timely performance of procedures – from dissection to recording – is particularly important because the isolated hippocampus constitutes such a dense, but delicate, preparation that maintaining functional connectivity of the structure in vitro requires great care. Preparing everything beforehand ensures that an adequate level of perfusion is available as early as possible to minimize cell damage and maintain physiological function.
- Sucrose-based artificial Cerebral Spinal Fluid (aCSF) solution
- Prepare 1X stock sucrose solution to be used for dissection and post-dissection incubation. Combine all components listed in Table 1, except for CaCl2, into 1 L of deionized water, and store at 4 °C for up to 2 weeks.
- Standard aCSF solution
- Prepare standard aCSF for high flow rate perfusion of the isolated hippocampus during electrophysiological recordings. To make the concentrated (5X) stock aCSF, dissolve all the components listed in Table 2, except CaCl2, in 2 L of deionized water and store at 4 °C.
- Prepare the equipment
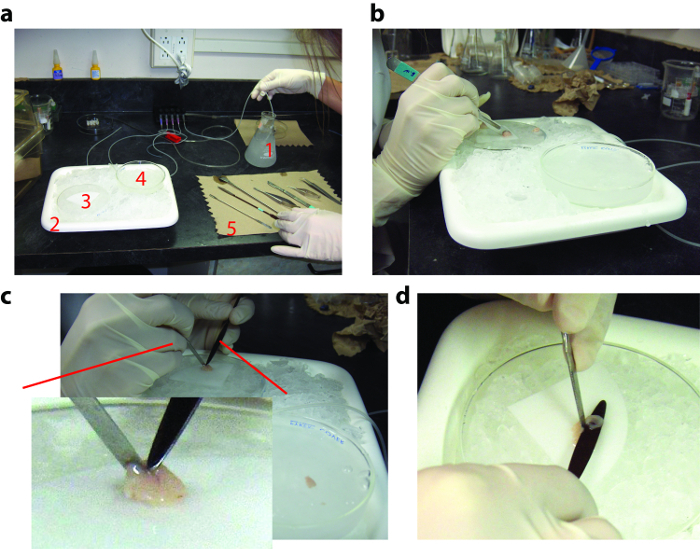
- Before starting dissection, set up the bench area with an ice-filled plastic tray (30 x 20 x 5 cm), carbogen connected tubing lines and three glass Petri dishes (10 x 2 cm) (See Figure 1).
- Add 300 mL of sucrose solution (1X stock) to a 500 mL beaker, bubble it with carbogen (95% O2, 5% CO2) and add Ca2+ [1.2 mM].
- Transfer 60 mL of this sucrose solution to a glass Petri dish and leave at room temperature. Place the remainder in a 4 °C freezer for 10 - 15 min.
- Bubble the ice-cold sucrose solution with carbogen throughout the dissection.
- Fill a second Petri dish (cold holding chamber) with sucrose solution (4 °C) and keep on ice.
- Add 30 mL of ice-cold sucrose solution to a 50 mL beaker.
2. Whole Hippocampus Dissection
NOTE: The method for dissecting the isolated hippocampus is essentially identical to the one developed and described originally22, but with additional details and changes regarding the perfusion rate and recording techniques.
- Brain dissection
- Anesthetize mouse (P20 - 35) under a fume hood with a small paper towel soaked with 1 mL of isoflurane (100%) that is added into the cage.
- Decapitate the anesthetized mouse and quickly submerge the head in ice-cold sucrose solution (50 mL beaker, ~ 5 s). Transfer onto a paper towel.
- Use a scalpel to make a medial skin incision (caudal to rostral) and push skin on the sides to expose the cranium.
- Cut open the skull with a small scissors and use a spatula to rapidly extract the brain and drop it into the Petri dish containing ice-cold sucrose solution. Allow 1-2 min for the brain to cool.
- While the brain is recovering, add 2 - 3 mL of sucrose solution onto a lens paper covered Petri (dissection) dish on ice.
- Isolating the hippocampus
- Place the brain onto the dissection dish in an upright position.
- Remove the cerebellum and frontal cortex with a razor blade. Cut the brain in half along the midsagittal plane and return the two isolated hemispheres to the holding chamber. Allow a further 1 - 2 min for recovery.
- Place single hemisected brain upright onto the dissection dish.
- Rotate the dish until midsagittal structures face the observer and the embedded outline of the septal complex is visible as a thin pear-shaped layer of tissue anterior to the thalamus.
- Hold the hemi-sected brain steady with the microspatula and place the small end of the polytetrafluoroethylene (PTFE)-coated spatula immediately behind (caudal to) the septal area.
- Insert the coated spatula beneath the septum and move the tip down until the dissection dish is reached. Sever the fibers that connect the septal area caudally. Repeat the same operation along the anterior edge of the septum and cut the fibers that connect to the frontal part of the brain.
- With the spatula lightly holding the inner part of the cortex just above the hippocampus in an upright position, use the microspatula to carefully pull down the thalamic, hypothalamic and remaining brain stem nuclei. Use the spatula to cut off and remove the pulled away tissue.
- Turn the isolated hemisphere on its side and identify the outline of the hippocampus.
- Insert the coated spatula in the lateral ventricle, underneath the rostral end of the dorsal hippocampus.
- Hold the spatula horizontally aligned with the midsagittal plane of the hemi-sected brain and slide it through the smooth contour of ventricular walls until the tip emerges caudally.
- Hold the spatula beneath the hippocampus and lightly press it onto the connecting fibers, along the inside layer where the hippocampus joins with the overlying cortex. Apply the microspatula on the external side of this layer and slide against the coated spatula to slice through the connecting fibers.
- To complete the extraction, rotate the dissection dish and insert the coated spatula beneath the ventral hippocampus. Lightly hold the inside of the cortical fold with the spatula and cut through the entorhinal connections using a slicing motion against the microspatula. NOTE: The whole septohippocampal complex can be removed by placing the coated spatula under the entire hippocampus, and pulling out the remaining brain tissue from underneath.
- Once isolation is complete, keep the hippocampus resting on the dish and add a drop of ice-cold sucrose solution to keep it cool. Carefully trim off any remaining cortex and fibers and separate the hippocampus from the septum by gently applying a razor blade through the fornix. NOTE: If patch clamp recordings are to be performed from pyramidal cells (see Section 4.6 Optogenetic control), the isolated hippocampus can be transversally cut at a 45° angle to expose the pyramidal layer of CA1 ("angle-cut" preparation), without compromising the local network circuitry in the remaining portion of the hippocampus.
- Transfer the preparation to room temperature sucrose solution and let it recover for 15 - 30 min before transferring to the recording chamber.
3. Set Up the Fast Perfusion for Recording the Isolated Hippocampus
- Set up the gravity-fed perfusion system to enable continuous high-speed inflow of oxygenated aCSF at 20 - 25 mL/min.
- Prepare aCSF (1X) flasks in advance and immerse them in a warming bath (~30 °C) at least 15 min before start of experiment. Turn on the in-line solution heater system (30 °C).
- Use aquarium bubblers and tubing lines connected to a carbogen tank to oxygenate aCSF throughout the experiment, and add CaCl2 [2 mM] prior to the start of perfusion.
- Stop aCSF flow and transfer the hippocampus to the recording chamber using the wide end of a glass pipette. Allow the sucrose saturated preparation to sink and settle at the bottom.
- Place the preparation in the center of the recording chamber (in line with the inlet-outlet axis) with the smooth surface of CA1 and SUB on top. Stabilize the hippocampus with small weights at the septal and temporal extremities and restart aCSF flow.
4. Electrophysiology in the Isolated Hippocampus
- Extracellular recording of in vitro hippocampal theta oscillations
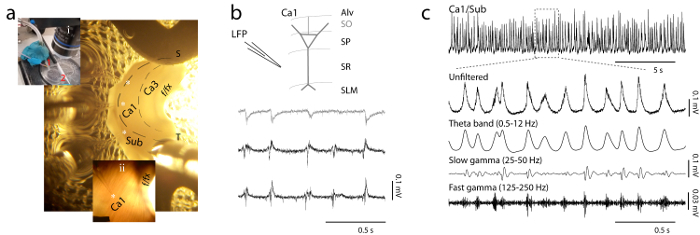
- For extracellular monitoring of Local Field Potential (LFP) or single-unit activity from the in vitro isolated hippocampus, use glass micropipettes (1 - 3 MΩ) filled with aCSF. Record low-noise AC-coupled field potential signals with a patch clamp amplifier of gain x1000, online band-pass filtering 0.1 - 500 Hz and a sampling rate of 5-20 kHz. NOTE: The custom-built microscope used in this protocol is equipped with low- (2.5X) and high-magnification (40X) objectives for low-magnification view of the hippocampus, as well as infrared video microscopy and epifluorescence for single cell recognition. The components of this system include the upright fluorescence microscope, fluorescence filter cubes, an analogue video camera and digital frame grabber with imaging software, an on-stage custom perfusion chamber (Figure 2A, inset i) and high-precision micromanipulators for neuronal recordings and optic fiber placement.
- Use the camera mounted on the microscope and connected to a computer display to inspect the hippocampal preparation under low magnification (2.5X) and rapidly check that there are no undercuts or damage to the external surface. The diagonally oriented arrangement of alveus fibers along the smooth surface of CA1 should be visible (Figure 2A, inset ii) when shining transmitted light through the hippocampus.
- Use the low-magnification wide-field objective to view the isolated hippocampus and the placement of LFP electrodes in specific recording locations. NOTE: A layout of the isolated hippocampal preparation with multiple electrodes placed in different recording locations is illustrated in Figure 2A.
- Lower an LFP electrode into the bath until it touches the surface (alveus) of the CA1 area. NOTE: This usually produces a fast transient in the AC-coupled recording similar to what happens when the electrode comes in contact with the recording media. An audio monitor can be useful to listen to the LFP channel signal after this step. NOTE: Oftentimes, early signs of activity will only appear after 10 - 15 min, therefore it is important to not advance quickly into the preparation. If after this period the surface LFP electrode is still not picking up activity, advance a little further down through the stratum oriens and pause periodically to see if any form of activity arises while approaching the principal cell layer. If nothing is detected after a further 5 - 10 min, carefully retract the electrode and place it elsewhere along the same region.
- Advance the LFP electrode through the pyramidal layer. Observe that extracellular spiking activity initially increases as single-unit discharge from individual neurons (mostly pyramidal and inhibitory basket cells) is detected. Lower the electrode further and note that spiking begins to fade again as the tip crosses into the radiatum. Observe that a clearly visible network oscillation in the theta-frequency range (4 - 12 Hz) becomes apparent and reaches maximum amplitude (~100 - 300 µV) as the recording location is lowered through the radiatum.
- Theta oscillations across a single region
- To test the spatial properties of spontaneous theta oscillations across the CA1 region, place the tip of a reference LFP electrode in a medial CA1 site (medially located along the longitudinal septo-temporal axis) and lower it into the radiatum as described previously.
- Place a second LFP electrode into a CA1 site (200 - 800 µm septal or temporal from the first LFP) and observe that CA1 theta oscillations can synchronize over relatively large distances.
- Theta oscillations across layers
- To test the properties of theta oscillations across hippocampal layers, leave a reference LFP electrode in a CA1 radiatum site. Starting from just above the stratum oriens, lower a second electrode into the pyramidal cell layer, and through the radiatum. Observe a gradual inversion of the LFP signal (relative phase reversal) across the pyramidal layer. NOTE: For representative examples of these types of activity, see Figure 2B. Examples of laminar/depth profiles and Current Source Density (CSD) analysis illustrating the site of phase reversal in isolated hippocampi have been published previously23,24,25.
- Gamma oscillations and theta-gamma coupling in the intact hippocampus
- Place a field electrode at the CA1/SUB border and lower it until it sits at the interface between the pyramidal and molecular layers. At this level, a field potential displaying clear gamma oscillations with changes in amplitude that phase-lock to the local theta rhythm can be recorded24. Adjust scaling to observe the slow time scale of theta-gamma coupling. Note that gamma bursts occur in two distinct frequency bands, which can be revealed by band-pass filtering the ongoing LFP signal in the slow (25 - 50 Hz) and fast gamma (150 - 250 Hz) range (See Figure 2C for representative filtered traces).
- Whole-cell patch clamp recording during in vitro hippocampal theta oscillations NOTE: In this part of the protocol, the goal is to obtain visually guided whole-cell patch clamp recordings from identified interneurons in isolated hippocampi from transgenic PV-TOM mice expressing the fluorescent protein, tdTomato, under the control of the PV promoter (for more details see materials and Ref 26).
- Use fluorescence video-microscopy with low (2.5X) and high power (40X) magnification to visualize tdTomato-positive interneurons located near the surface of a hippocampal preparation from a PV-TOM mouse (Figure 3). Note that while PV-TOM neurons can be successfully visualized and targeted for patch through the alveus (up to a 100 µm), non-fluorescent pyramidal cells can also be reached relatively easily when the hippocampus is prepared with the angle-cut technique (see Section 2.2.14 note).
- Under low magnification view of the hippocampus, place an LFP electrode in CA1/SUB to monitor theta oscillations while preparing for patch clamp experiments.
- Switch to 40X magnification and immerse the objective over the target region; Lower it until the top layers become visible. Manipulate the position of the microscope to ensure sufficient open access for patch pipette approach from the opposite side.
- Under fluorescence microscopy, select a fluorescent PV-TOM cell and approach it with a patch pipette (2 - 3 MΩ) filled with standard intracellular solution.
- Use a mouth piece to apply light positive pressure to the pipette and monitor the resistance as the electrode is lowered onto the cell. When the tip encounters the target cell, the pipette resistance increases and a clear dimple appears as positive pressure pushes on the membrane. Release the pressure and check that the current pulse flattens as a seal starts to form. Immediately clamp to a hyperpolarized potential (-70 mV), and once a GΩ seal is achieved, rupture the cell membrane with a small amount of negative pressure applied through the pipette holder.
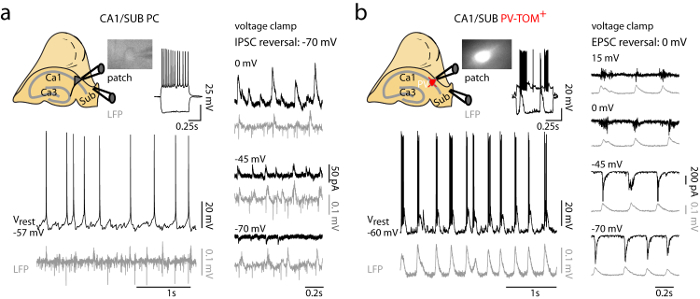
- Once in whole-cell configuration, examine the physiological properties of identified PV cell during spontaneous hippocampal oscillations (Figure 3B).
- Observe that intracellular membrane potential recording from PV cells are characterized by fast-spiking behavior and bursts of action potentials synchronized to the ongoing CA1/SUB theta rhythm.
- Switch to voltage-clamp and hold the cell at different membrane potentials to characterize the ratio of excitatory versus inhibitory synaptic currents generated by the intrinsic rhythmic activity. An example of patch clamp recording from a pyramidal cell is shown in Figure 3A for comparison.
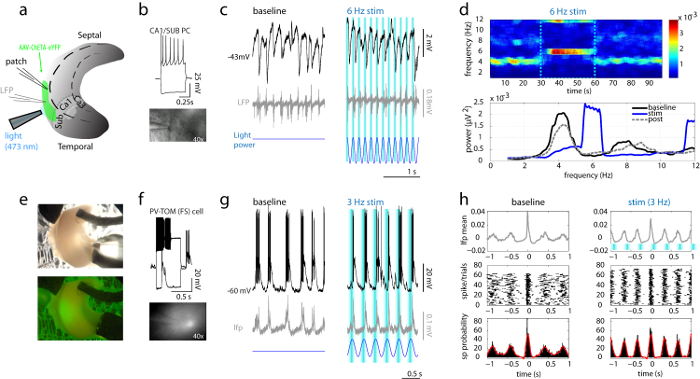
- Optogenetic control in the isolated hippocampus NOTE: For optogenetic control of hippocampal network activity, a cell-type specific strategy involving rhythmic activation of PV-containing GABAergic cells is used here as these cells were shown to play a major role in synchronizing principal cell populations in the isolated hippocampus25. Selective expression of the excitatory channelrhodopsin-2 (ChR2) in PV cells is achieved by crossing the PV-Cre mouse line with mice constitutively expressing ChR2 Cre-dependently (Ai32), thereby generating PV-ChY mice. Alternatively, viral vector constructs (e.g., AAVdj-ChETA-eYFP) can be injected into the hippocampus of PV-Cre or PV-TOM mice (P15-16) to drive the expression of the excitatory opsin in CA1/SUB interneurons (Figure 4A). NOTE: This strategy has been described previously25.
- Place an LFP electrode in the CA1/SUB area and patch a nearby pyramidal cell in the isolated hippocampus of a mouse expressing the blue-light sensitive excitatory opsin ChR2 in PV interneurons.
- Place an optic fiber light guide (0.2 - 1 mm diameter) above the hippocampal preparation and center it on the recorded region. Use blue light (473 nm) from a LED source for optogenetic stimulation, consisting of light pulses (10 - 20 ms, triggered by a transistor-transistor logic signal) or sine wave voltage commands delivered at theta frequencies (4 - 12 Hz). NOTE: The custom LED-based light stimulation assembly has been described elsewhere25.
- Switch to current clamp and characterize the activity of the pyramidal during spontaneous theta oscillations.
- Start the stimulation protocol and record the light responses.Observe that field oscillations and synaptic activity in the recorded neuron become increasingly synchronized during optogenetic stimulation and that rhythmic pacing of PV cells results in a robust control of both frequency and power of theta oscillations (Figure 4).
Representative Results
This section illustrates examples of results that can be obtained by studying theta oscillations in the mouse isolated hippocampal preparation in vitro. The dissection procedure for extracting the isolated hippocampus is illustrated in Figure 1. Using this preparation, intrinsic theta oscillations can be examined during placement of multiple field electrodes, recording overall activity and synchronized synaptic inputs to neuronal populations in different regions and layers of the isolated hippocampus (Figure 2). Representative results from simultaneous whole-cell patch clamp and extracellular recordings are presented to characterize the firing and synaptic properties of specific cell types during spontaneous hippocampal theta oscillations (Figure 3), as well as during optogenetic manipulation of rhythmic activity (Figure 4).
Figure 1: Dissection Procedure for the Isolated Intact Hippocampus Preparation. (a) General view of the dissection setup. Top right: carbogenated ice-cold sucrose solution flask (1); bottom left: ice-filled plastic tray (2) holding the dissection dish covered with lens paper (3); the cold holding chamber containing sucrose solution (4); and a set of surgical tools (5). (b) View of the mouse brain before hemisection on the dissection dish. (c) Recovery of hemisected brain in the cold holding chamber and zoomed view (inset) of the left brain hemisphere before inserting the small end of the coated spatula under the septum. (d) Coated spatula placed under the isolated hippocampus, along the CA1/SUB region, with the remaining brain tissue is pulled out from underneath. Please click here to view a larger version of this figure.
Figure 2: Configuration of the Setup for Recording in vitro Theta Oscillations from the Intact Hippocampal Preparation in the Submerged Recording Chamber. (a) The isolated hippocampus is shown with a layout of hippocampal regions and multiple electrodes placed in four different recording sites distributed septotemporally (indicated by white asterisks). In the view of the recording chamber platform shown above (inset i), the inlet and outlet for fast-perfusion flow are indicated by numbers (1, 2). In the enlarged image of the hippocampus shown below (inset ii), a single electrode is placed in the midsepetotemporal CA1 and fibers of the alveus are readily visible, running diagonally toward the subiculum. S: septal, T: temporal, f/fx: fimbria-fornix. (b) Schematic representation of the organization of CA1 layers with representative LFP traces recorded simultaneously from stratum oriens (grey) and stratum radiatum (black). Note the inverted phase of signals between the two layers. Alv: stratum alveus, PR: stratum pyramidale, SR: stratum radiatum. SLM: Stratum Lacunosum Moleculare. (c) Example LFP trace showing spontaneous theta oscillation recorded from the CA1/SUB area (20 sec segment) and 2 sec expanded segments (below) from unfiltered signal; band-pass filtered for theta frequencies (0.5 - 12 Hz); slow gamma (25 - 55 Hz); and fast gamma (125 - 250 Hz). Please click here to view a larger version of this figure.
Figure 3: Cell-type Specific Activity of Pyramidal Cell and PV Interneuron during Spontaneous Theta Oscillations in the Intact Hippocampal Preparation from a PV-TOM Mouse. (a) Synaptic activity recorded from a pyramidal cell during theta oscillations. Current-clamp traces (left panel) show spontaneous but not rhythmic firing at rest, and inhibitory postsynaptic potentials (iPSPs) that were not clearly synchronized with the slowly emerging LFP oscillation. Voltage-clamp recordings (right panel) show that the corresponding inhibitory postsynaptic currents (iPSCs) have their reversal potential around -70 mV. (b) Synaptic activity recorded from a fast-spiking fluorescent PV interneuron during theta oscillations. In current-clamp (left), this cell was spontaneously firing at rest and was strongly driven by rhythmic excitatory postsynaptic potentials (ePSPs) that were phase-locked with the stable LFP oscillation. In voltage-clamp recordings (right), the excitatory postsynaptic current reversal potential was approximately 0 mV. Schematic drawings on top show the experimental setup together with the placement of patch and field electrodes in the CA1/SUB and high (40X) magnification images of the recorded pyramidal cell and fluorescent PV interneuron. Please click here to view a larger version of this figure.
Figure 4: Local Field Potential and Simultaneous Patch Clamp Recordings from Single CA1/SUB Pyramidal and PV Neurons during Spontaneous and Optogenetically-driven Theta Oscillations in the Isolated Hippocampus. (a) Diagram showing the LFP and patch electrode recording sites with placement of the light-fiber guide above the CA1/SUB region expressing a blue-light sensitive opsin (ChETA coupled with the fluorophore eYFP). (b) Characterization of a pyramidal neuron showing typical Regular-Spiking (RS) properties (top) and high magnification (40X) image of the recorded cell (bottom). (c) Sample current-clamp recording of the same cell at depolarized membrane potential (black) together with the LFP signal (grey) and showing synchronized rhythmic IPSPs during spontaneous oscillation (left) and during theta-frequency (6 Hz) light stimulation (right). The pattern of light stimulation (blue shading) is depicted on top of the voltage traces. (d) Spectrogram and power spectra of LFP waveform before, during and after a 6 Hz light simulation (baseline, stim, post). (e) Low magnification bright-field and fluorescence images of the isolated hippocampal preparation showing eYFP fluorescence (in green) localized in the CA1/SUB region. (f) Current steps characterization showing Fast-Spiking (FS) behaviour of a recorded PV-TOM interneuron (40X fluorescence image below). (g) Membrane potential recording showing large EPSPs and rhythmic firing of the recorded PV cell synchronized with the LFP signal during spontaneous field oscillation (left) and during light stimulation (right) at 3 Hz. (h) Field Triggered Average (FTA) of PV cell spikes over the CA1/SUB LFP signal. The discharge of the PV cell over multiple trials (centered on the LFP peaks) was converted to a FTA of spikes recorded during spontaneous oscillation (baseline) and during light stimulation (stim). Middle and bottom graphs show the raster plots of spikes and spike probability histograms (average probability is shown in red). Topmost graphs show plots of the average LFP signal which increased in power during light stimulation in parallel with highly synchronized firing of the PV cell phase-locked to the peak of the LFP oscillation. Please click here to view a larger version of this figure.
| SUCROSE SOLUTION (1X) FOR ISOLATED HIPPOCAMPUS | |||
| (Stock solution) | |||
| Compound | MW | Final Conc. (mM) | Amount for 1 L (g) |
| sucrose | 342.3 | 252 | 86.26 g |
| NaHCO3 | 84.01 | 24 | 2.020 g |
| glucose | 180.2 | 10 | 1.800 g |
| KCl | 74.55 | 3 | 0.223 g |
| MgSO4 | 120.4 | 2 | 0.241 g |
| NaH2PO4 | 120 | 1.25 | 0.150 g |
| CaCl2.2H2O [1 M] stock | 147 | 1.2 | 120 μL / 0.1 L * |
| * add 360 μL CaCl2 [1 M] for 0.3 L oxygenated sucrose solution | |||
| pH = 7.4 when oxygenated, Osm 310 - 320 |
Table 1.
| STANDARD ACSF SOLUTION (5X) FOR PERFUSION | |||
| (Stock solution) | |||
| Compound | MW | Final Conc. (mM) | Amount for 2 L 5X |
| NaCl | 58.44 | 126 | 73.6 |
| NaHCO3 | 84.01 | 24 | 20.2 |
| glucose | 180.2 | 10 | 18 |
| KCl | 74.55 | ♦ 4.5 | 3.355 |
| MgSO4 | 120.4 | 2 | 2.41 |
| NaH2PO4 | 120 | 1.25 | 1.5 |
| Ascorbate | 176.1 | 0.4 | 0.705 |
| CaCl2.2H2O [1 M] stock | 147 | 2 | 2 mL / L * |
| * add 2 mL CaCl2 [1 M] for 1 L aCSF (1x) oxygenated solution | |||
| pH = 7.4 when oxygenated, Osm 310 - 320 | |||
| ♦ A slightly elevated [K+]o is used for this aCSF solution (compared to normal aCSF 2.5 mM KCl) to increase excitability of hippocampal networks and facilitate the emergence of theta oscillations. |
Table 2.
Discussion
While electrophysiological recordings from acute hippocampal slices constitute a standard in vitro technique, the methods presented here differ substantially from the classic approach. Unlike the thin slice preparations where specific cell layers are visible at the surface and can be examined directly, the intact hippocampal preparations are more akin to in vivo configurations where electrodes are lowered into targeted brain regions while crossing through individual layers. The integrity of the hippocampus is preserved together with functional connectivity and properties of local neuronal populations. This provides a complex and powerful tool to investigate small and large-scale network oscillations in the hippocampus. For example, the combination of prewired circuitry and cell-type specific properties in the network produces intrinsically and spontaneously generated rhythmic theta and gamma oscillations that mimic important features of hippocampal activity in vivo. Using the methods presented in this protocol, the hippocampus can be extracted quickly from the rodent brain and remain viable for several hours in a fast flowing aCSF environment. Basic electrophysiological techniques are readily applied to investigate how synchronous activity and synaptic function of specific neuronal types influence hippocampal dynamics.
Our procedure has provided the first opportunity to systematically explore the dynamics of local field potential oscillations across the whole septo-temporal (dorso-ventral) axis of the hippocampus in vitro (see Refs 22,23,24,25,26,27,28). At the network and physiological levels, the intact preparation preserves: 1) The functional synaptic interactions between cells required to reliably support intrinsic network oscillations with the frequency range and profile of theta waves in vivo, 2) The sharp change in relative phase of theta oscillation at the level of stratum pyramidale, 3) The spatial distribution and coherence of theta oscillation in the hippocampus and its interaction with local gamma frequency rhythms, 4) The amplitude-phase relationship of theta/gamma waves and 5) The specific temporal association of principal cell and interneuron firing with field potential theta oscillations.
Whereas the absence of external input to the hippocampus may appear as a limitation to the technique, it also allows the study of the internal dynamics of the hippocampus in a manner that cannot be done in vivo. In addition, external inputs can be mimicked by optogenetic stimulation of specific fiber terminals and afferent pathways. Intact preparations offer new possibilities to study how network activities propagate and interact in large-scale networks involving the hippocampus and other connected structures. As such, one main application of the intact preparation technique is the investigation of functional network connectivity within or between brain regions. Using the in vitro intact hippocampus should therefore not only provide further information on cellular and network properties of the hippocampus, but also shed light on how these interact to shape processing, integration and generation of information within the brain.
Rhythmic network oscillations engaging the coordinated electro-chemical signaling of large neuronal ensembles act as a central mechanism for encoding, storing and transferring information within and across brain areas. Hippocampal oscillations are thought to be crucial for processing of spatiotemporal information, encoding, and memory. Conversely, hippocampal dysfunction is thought to underlie disorders such as Alzheimer's disease and schizophrenia, which are associated with altered oscillation patterns29. As such, the study of self-generated hippocampal oscillations using intact hippocampal preparations has become a renewed means for elucidating basic properties of nervous system function and dysfunction.
Disclosures
The authors declare no competing commercial or financial interests.
Acknowledgments
This work was supported by the Canadian Institutes of Health Research and Natural Sciences.
References
- Buzsaki G. Theta rhythm of navigation: link between path integration and landmark navigation, episodic and semantic memory. Hippocampus. 2005;15(7):827–840. doi: 10.1002/hipo.20113. [DOI] [PubMed] [Google Scholar]
- Sanders H, Renno-Costa C, Idiart M, Lisman J. Grid Cells and Place Cells: An Integrated View of their Navigational and Memory Function. Trends Neurosci. 2015;38(12):763–775. doi: 10.1016/j.tins.2015.10.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O'Keefe J, Recce ML. Phase relationship between hippocampal place units and the EEG theta rhythm. Hippocampus. 1993;3(3):317–330. doi: 10.1002/hipo.450030307. [DOI] [PubMed] [Google Scholar]
- Winson J. Loss of hippocampal theta rhythm results in spatial memory deficit in the rat. Science. 1978;201(4351):160–163. doi: 10.1126/science.663646. [DOI] [PubMed] [Google Scholar]
- M'Harzi M, Jarrard LE. Strategy selection in a task with spatial and nonspatial components: effects of fimbria-fornix lesions in rats. Behav Neural Biol. 1992;58(3):171–179. doi: 10.1016/0163-1047(92)90426-5. [DOI] [PubMed] [Google Scholar]
- Osipova D, et al. Theta and gamma oscillations predict encoding and retrieval of declarative memory. J Neurosci. 2006;26(28):7523–7531. doi: 10.1523/JNEUROSCI.1948-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stumpf C, Petsche H, Gogolak G. The significance of the rabbit's septum as a relay station between the midbrain and the hippocampus. II. The differential influence of drugs upon both the septal cell firing pattern and the hippocampus theta activity. Electroencephalogr Clin Neurophysiol. 1962;14:212–219. doi: 10.1016/0013-4694(62)90031-7. [DOI] [PubMed] [Google Scholar]
- Mitchell SJ, Ranck JB. Generation of theta rhythm in medial entorhinal cortex of freely moving rats. Brain Res. 1980;189(1):49–66. doi: 10.1016/0006-8993(80)90006-2. [DOI] [PubMed] [Google Scholar]
- Alonso A, Garcia-Austt E. Neuronal sources of theta rhythm in the entorhinal cortex of the rat. I. Laminar distribution of theta field potentials. Exp Brain Res. 1987;67(3):493–501. doi: 10.1007/BF00247282. [DOI] [PubMed] [Google Scholar]
- Vertes RP, Kocsis B. Brainstem-diencephalo-septohippocampal systems controlling the theta rhythm of the hippocampus. Neuroscience. 1997;81(4):893–926. doi: 10.1016/s0306-4522(97)00239-x. [DOI] [PubMed] [Google Scholar]
- Bland BH, Colom LV, Konopacki J, Roth SH. Intracellular records of carbachol-induced theta rhythm in hippocampal slices. Brain Res. 1988;447(2):364–368. doi: 10.1016/0006-8993(88)91141-9. [DOI] [PubMed] [Google Scholar]
- Cobb SR, Buhl EH, Halasy K, Paulsen O, Somogyi P. Synchronization of neuronal activity in hippocampus by individual GABAergic interneurons. Nature. 1995;378(6552):75–78. doi: 10.1038/378075a0. [DOI] [PubMed] [Google Scholar]
- Williams JH, Kauer JA. Properties of carbachol-induced oscillatory activity in rat hippocampus. J Neurophysiol. 1997;78(5):2631–2640. doi: 10.1152/jn.1997.78.5.2631. [DOI] [PubMed] [Google Scholar]
- Chapman CA, Lacaille JC. Cholinergic induction of theta-frequency oscillations in hippocampal inhibitory interneurons and pacing of pyramidal cell firing. J Neurosci. 1999;19(19):8637–8645. doi: 10.1523/JNEUROSCI.19-19-08637.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strata F. Intrinsic oscillations in CA3 hippocampal pyramids: physiological relevance to theta rhythm generation. Hippocampus. 1998;8(6):666–679. doi: 10.1002/(SICI)1098-1063(1998)8:6<666::AID-HIPO9>3.0.CO;2-A. [DOI] [PubMed] [Google Scholar]
- Kocsis B, Bragin A, Buzsaki G. Interdependence of multiple theta generators in the hippocampus: a partial coherence analysis. J Neurosci. 1999;19(14):6200–6212. doi: 10.1523/JNEUROSCI.19-14-06200.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fellous JM, Sejnowski TJ. Cholinergic induction of oscillations in the hippocampal slice in the slow (0.5-2 Hz), theta (5-12 Hz), and gamma (35-70 Hz) bands. Hippocampus. 2000;10(2):187–197. doi: 10.1002/(SICI)1098-1063(2000)10:2<187::AID-HIPO8>3.0.CO;2-M. [DOI] [PubMed] [Google Scholar]
- Gillies MJ, et al. A model of atropine-resistant theta oscillations in rat hippocampal area CA1. J Physiol. 2002;543(Pt 3):779–793. doi: 10.1113/jphysiol.2002.024588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gloveli T, et al. Orthogonal arrangement of rhythm-generating microcircuits in the hippocampus. Proc Natl Acad Sci U S A. 2005;102(37):13295–13300. doi: 10.1073/pnas.0506259102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Konopacki J, Eckersdorf B, Kowalczyk T, Golebiewski H. Firing cell repertoire during carbachol-induced theta rhythm in rat hippocampal formation slices. Eur J Neurosci. 2006;23(7):1811–1818. doi: 10.1111/j.1460-9568.2006.04679.x. [DOI] [PubMed] [Google Scholar]
- Hajos N, et al. Maintaining network activity in submerged hippocampal slices: importance of oxygen supply. Eur J Neurosci. 2009;29(2):319–327. doi: 10.1111/j.1460-9568.2008.06577.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manseau F, Goutagny R, Danik M, Williams S. The hippocamposeptal pathway generates rhythmic firing of GABAergic neurons in the medial septum and diagonal bands: an investigation using a complete septohippocampal preparation in vitro. J Neurosci. 2008;28(15):4096–4107. doi: 10.1523/JNEUROSCI.0247-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goutagny R, Jackson J, Williams S. Self-generated theta oscillations in the hippocampus. Nat Neurosci. 2009;12(12):1491–1493. doi: 10.1038/nn.2440. [DOI] [PubMed] [Google Scholar]
- Jackson J, Goutagny R, Williams S. Fast and slow gamma rhythms are intrinsically and independently generated in the subiculum. J Neurosci. 2011;31(34):12104–12117. doi: 10.1523/JNEUROSCI.1370-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Amilhon B, et al. Parvalbumin Interneurons of Hippocampus Tune Population Activity at Theta Frequency. Neuron. 2015;86(5):1277–1289. doi: 10.1016/j.neuron.2015.05.027. [DOI] [PubMed] [Google Scholar]
- Huh CY, et al. Excitatory Inputs Determine Phase-Locking Strength and Spike-Timing of CA1 Stratum Oriens/Alveus Parvalbumin and Somatostatin Interneurons during Intrinsically Generated Hippocampal Theta Rhythm. J Neurosci. 2016;36(25):6605–6622. doi: 10.1523/JNEUROSCI.3951-13.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gu N, et al. NMDA-dependent phase synchronization between septal and temporal CA3 hippocampal networks. J Neurosci. 2013;33(19):8276–8287. doi: 10.1523/JNEUROSCI.0179-13.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jackson J, et al. Reversal of theta rhythm flow through intact hippocampal circuits. Nat Neurosci. 2014;17(10):1362–1370. doi: 10.1038/nn.3803. [DOI] [PubMed] [Google Scholar]
- Gonzalez-Burgos G, Lewis DA. GABA neurons and the mechanisms of network oscillations: implications for understanding cortical dysfunction in schizophrenia. Schizophr Bull. 2008;34(5):944–961. doi: 10.1093/schbul/sbn070. [DOI] [PMC free article] [PubMed] [Google Scholar]