

Abstract
Influenza viruses exhibit a remarkable ability to adapt and evade the host immune response. One way is through antigenic changes that occur on the surface glycoproteins of the virus. The generation of escape variants is a powerful method in elucidating how viruses escape immune detection and in identifying critical residues required for antibody binding. Here, we describe a protocol on how to generate influenza A virus escape variants by utilizing human or murine monoclonal antibodies (mAbs) directed against the viral hemagglutinin (HA). With the use of our technique, we previously characterized critical residues required for the binding of antibodies targeting either the head or stalk of the novel avian H7N9 HA. The protocol can be easily adapted for other virus systems. Analyses of escape variants are important for modeling antigenic drift, determining single nucleotide polymorphisms (SNPs) conferring resistance and virus fitness, and in the designing of vaccines and/or therapeutics.
Keywords: Immunology, Issue 126, Influenza virus, monoclonal antibodies, escape variants, microbiology, virology
Introduction
Similar to other RNA viruses, influenza A viruses possess an error-prone polymerase that allows for the generation of a multitude of antigenic variants with each round of replication1,2,3. The influenza A virus has an astonishing ability to adapt and evade the human immune response via antigenic drift, which is achieved through an accumulation of mutations on the surface glycoproteins that leads to the loss of antibody binding. Antigenic drift of the viral surface glycoproteins, HA and neuraminidase (NA), necessitates the need to reformulate and administer the vaccine annually.
Technological advancements in the isolation and generation of antigen-specific antibodies have yielded a high number of vaccine-induced mAbs4,5,6,7,8. In turn, the characterization of the epitopes of mAbs that broadly neutralize influenza A viruses has greatly aided the development of several universal influenza vaccine candidates9,10,11,12,13,14. Elucidating the antigenic footprint of a mAb reveals the structural determinants of neutralization and allows for an informed approach towards vaccine design. However, it is neither realistic nor cost-effective for laboratories to structurally characterize extensive panels of mAbs through x-ray crystallography or cryo-electron microscopy in order to map epitopes on the viral antigen15,16,17,18.
X-ray crystallography or cryo-electron microscopy requires expensive equipment, specialized techniques and potentially an extensive amount of time to generate data. An alternative and faster approach is utilizing the rapid generation of diverse viral populations via the error-prone RNA-dependent RNA polymerase to generate escape mutants to determine the epitopes of mAbs19,20,21,22,23. The generation of escape variants does not require any special equipment or technique and can be performed with conventional laboratory reagents and equipment.
Here, we describe a method that allows for the mapping of critical residues required for mAb binding that recognize the influenza HA.
Protocol
CAUTION: A number of influenza viruses circulating in the human population (e.g., H1, H3) are biosafety level 2 class pathogens that must be handled with care and proper personal protective equipment. Handling of viruses must be approved by the Institutional Review Board. The following protocol was approved by the Institutional Review Board at Mount Sinai.
NOTE: HA-specific antibodies that inhibit viral replication can generally be categorized into i) ones that bind on or adjacent of the receptor binding site on top of the globular head and ii) ones that bind distal of the receptor binding domain, which includes the lateral side of the globular head and the stalk region of the HA. Antibodies that target the receptor binding site prevent the engagement of sialic acid motifs on the surface of target cells and can be measured using a hemagglutination inhibition (HI) assay. Antibodies that are HI-negative, such as stalk-specific antibodies, can still inhibit viral replication, but can only be assessed using neutralization assays.
1. Categorizing Antibodies Based on HI and Neutralization Activities
- HI assay
- In a 96-well V-bottom plate, add 25 µL of 1x PBS on columns 2 to 12.
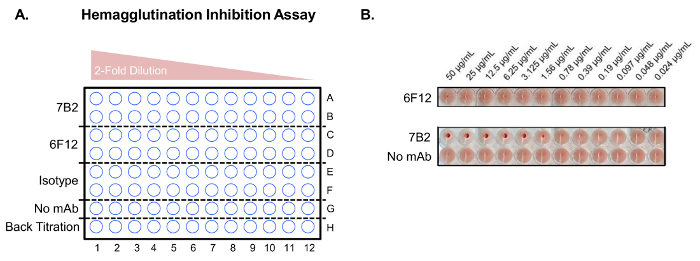
- Perform 2-fold serial dilutions of the antibodies by transferring 25 µL from column 1 to column 2, and so forth. Discard the last 25 µL from column 12 (Figure 1A). NOTE: Make sure to include a row of no antibody control.
- Dilute the virus stock (a reassortant virus expressing the HA and NA of A/California/04/09 with the internal segments of A/Puerto Rico/8/34) to 8 hemagglutination units/25 µL diluted virus stock. Add 25 µL of the diluted virus stock (8 hemagglutination) to each well (Rows A to G). NOTE: The antibody and virus mixture should have a final volume of 50 µL with a starting final concentration of 50 µg/mL.
- Incubate the plate at room temperature (RT) for 45 min.
- For the back-titration row (H), add 50 µL of 1x PBS on wells H2 to H12. Add 100 µL of the 8 hemagglutination units/25 µL to well H1. Serially dilute two-fold by transferring 50 µL from H1 to H2, and so forth. Discard the last 50 µL from well H12. Finally, add 50 µL of 0.5% chicken red blood cells (RBC) to all wells of the 96-well V-bottom plate. NOTE: The mAb samples in the assay should have a final volume of 100 µL: mAb (25 µL), virus (25 µL) and RBC (50 µL). The final volume of the no mAb control should contain 25 µL of 1x PBS, 25 µL of virus and 50 µL of RBC.
- Incubate the plate at 4 °C for 1 h.
- Visually read the plates for HI activity. If there is a positive readout for a particular antibody, proceed to step 2.1 to generate escape variants. If the antibody is HI-negative, proceed below to step 1.2 to assess if the antibody has neutralizing activity in a cell culture assay. NOTE: A positive readout of an HI-active mAb is indicated by a dark red RBC pellet in the center of a well (Figure 1B; 7B2) that will form a tear drop when the 96-well V-bottom plate is held at a 45° angle. A negative readout will not form a dark red RBC pellet in the well (Figure 1B; 6F12 and no mAb). The isotype control will also not form a dark red RBC pellet and should look identical to the stalk-specific antibody, 6F12 or the no mAb control sample (Figure 1B)23.
- Microneutralization assay
- Plate Madin Darby Canine Kidney (MDCK) cells at a density of 2 x 104 cells/well in a tissue culture treated 96-well plate and incubate at 37 °C and 5% CO2 for 17 to 19 h. NOTE: The cells can also be allowed to adhere to the bottom of the well for a minimum of 4 h before use.
- In a separate 96-well plate, perform seven 3-fold serial dilutions of the human mAb 4D055, CR911417 or isotype IgG control, at a starting concentration of 200 µg/mL in 1X Minimal Essential Medium (MEM) supplemented with tosyl phenylalanyl chloromethyl ketone (TPCK)-treated trypsin (1 µg/mL) (Figure 2). NOTE: Row A should contain 75 µL of diluted antibody (starting concentration of 200 µg/mL). Serially dilute (3-fold) down the plate by transferring 25 µL from Row A to Row B, and so forth. Rows A to H should have a final volume of 50 µL. There is no need to change tips in between dilution transfers.
- Dilute the virus stock (reassortant virus expressing the HA and the NA of A/Shanghai/1/13 with the internal segment of A/Puerto Rico/8/34) to a 100 of 50% tissue culture infectious dose (TCID50)/50 µL in 1x MEM supplemented with TPCK-treated trypsin (1 µg/mL)24. Add 50 µL/well of diluted virus to the antibody preparations (step 1.2.2). Add 50 µL of 1X MEM to the uninfected cell control wells.
- Incubate the virus-antibody mixtures in a 37 °C incubator (with 5% CO2) for 1 h. NOTE: The virus-antibody mixtures should have a total volume of 100 µL: 50 µL of antibody dilution (step 1.1.2) and 50 µL of virus containing 100 TCID50 (step 1.2.3).
- Aspirate the media in the wells and add the entire 100 µL of virus-antibody mixtures to corresponding wells. NOTE: The aspiration is done using an 8-channel aspirator adapter attached to a vacuum. Alternatively, an 8- or 12-well multichannel micro-pipette may be used to manually aspirate. All aspiration during the inoculum removal or washes is done from the highest concentration of antibody to the lowest, without the need to change tips.
- Wash the monolayer with 200 µL of 1x PBS. Aspirate 200 µL of 1x PBS (as in step 1.2.5). Repeat the washing once more for a total of two washes.
- Infect the monolayer of MDCK cells by adding the entire 100 µL/well of virus-antibody mixtures (from step 1.2.4) on the monolayer and infect/incubate at 37 °C (with 5% CO2) for 1 h.
- During the infection, in a separate 96-well plate, prepare another set of antibody dilutions. Add 150 µL of 100 µg/mL of the respective antibody in Row A and 100 µL of 1x MEM supplemented with TPCK-treated trypsin (1 µg/mL) in Rows B to H. Perform 3-fold dilutions by transferring 50 µL from Row A to Row B, and so forth, down to Row H. Discard the last 50 µL from Row H. The total volume for each well should equal 100 µL. Set aside. NOTE: The antibodies are diluted in 1x MEM supplemented with TPCK-treated trypsin (1 µg/mL).
- Aspirate the virus-antibody inoculum from the monolayer in step 1.2.7 and replenish with the entire 100 µL/well of the appropriate antibody dilution prepared in step 1.2.8. NOTE: If a well contained a final antibody concentration of 100 µg/mL during the infection (step 1.2.7), then the replenishing media should also contain a final antibody concentration of 100 µg/mL (step 1.2.8).
- Incubate for 24 h in a 37 °C incubator (with 5% CO2).
- Aspirate the media from the 96-well plates and wash with 200 µL/well of 1x PBS three times.
- Fix the cells with 100 µL of cold 80% acetone for 1 h at -20 °C. NOTE: The 80% acetone solution is diluted in double distilled (dd) H2O (e.g., 80 mL of 100% acetone plus 20 mL of ddH20). The 80% acetone solution can be chilled on ice before use.
- Wash the cells with 200 µL/well of 1X PBS three times.
- Block the plates with 200 µL/well of 5% milk diluted in 1X PBS and incubate plates at RT for 1 h.
- Add 100 µL of biotinylated anti-influenza A nucleoprotein (NP) primary antibody diluted 1:2,000 in 1x PBS/1% bovine serum albumin (BSA) and incubate plates at RT for 1 h. NOTE: For influenza B viruses, an influenza B virus-specific anti-NP antibody should be used.
- Wash the plates with 1x PBS three times.
- Add 100 µL of streptavidin-horse radish peroxidase (HRP) conjugate antibody diluted 1:3,000 in 1x PBS/1% BSA and incubate the plates at RT for 1 h.
- Wash the plates with 200 µL of 1x PBS three times.
- Add HRP substrate reagent at 100 µL/well and incubate in the dark at RT. NOTE: The incubation time should be optimized before the addition of the acidic stopping buffer (below). Generally speaking, 15 to 30 min is sufficient.
- Quench the reaction with 50 µL/well of 5 M HCl. CAUTION: 5M HCl is a highly corrosive reagent that can cause damage to eyes, skin and mucous membranes. Addition of this reagent should be done under a vented hood with proper personal protective equipment.
- Read the plates at 492 nm and subtract the background (uninfected cells) wells.
- Calculate the percentage inhibition with the following formula:
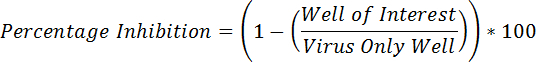
- If an antibody has neutralization activity (and no HI activity), proceed to step 2.2. NOTE: Antibodies that lack in vitro neutralization activity will lack HI activity.

2. Generation of Escape Mutant Variants
NOTE: Neutralizing antibodies that have or lack HI activity are further analyzed with the specific protocols described below.
- Protocol 1: HI-positive/Neutralization-positive antibodies ( Figure 3A )
- Prepare a virus stock of 106 plaque forming units/milliliter (PFU/mL) in 1x PBS in a 400 µL volume.
- Prepare four dilutions of the antibody of interest in increasing concentrations (e.g., 0, 0.5, 0.05 and 0.005 mg/mL) in 1x PBS in a volume of 100 µL per dilution. NOTE: Wild-type virus should always be passaged in parallel and in the absence of antibody. The sequences of these passaged viruses will assist in distinguishing between adaptation to cell culture conditions and escape mutations.
- Mix 100 µL of 106 PFU/mL of virus with 100 µL of each antibody dilution or 100 µL of 1x PBS.
- Incubate for 1 h in a 37 °C incubator (with 5% CO2). Vortex briefly. Inject 200 µL of each mixture into specific-pathogen free (SPF) embryonated chicken eggs.
- Incubate the eggs at 37 °C (without CO2) for 40-44 h.
- Sacrifice the virus infected-embryonated eggs by incubating at 4 °C for a minimum of 6 h.
- Perform the hemagglutination assay, as described above24,26. If all of the allantoic fluid preparations do not have hemagglutination titers, repeat from step 2 with antibody dilutions ranging from 0.005 mg/mL to 0.00005 mg/mL. NOTE: A saturating concentration of an HI-positive antibody may neutralize all the virus particles present. Therefore, it may be necessary to decrease the amount of antibody present in the passaging.
- Confirm the escape variants by performing the HI assay24 (step 1.1). NOTE: HI-active antibodies block HA engagement of sialic acid motifs on the target cells. Therefore, a virus in the presence of its cognate antibody loses the ability to agglutinate RBCs (presence of RBC pellet). Theoretically, escape variants of HI-active antibodies can still bind sialic acid motifs even in the presence of its cognate antibody and thus can agglutinate RBCs (no RBC pellet). If the HI of the antibody of interest is still detectable, repeat the protocol from step 2.1.2 with a higher starting concentration of the antibody.
- Protocol 2: HI-negative/Neutralization-positive antibodies (Figure 3B) NOTE: In order to generate escape variants against neutralizing antibodies that lack HI activity, the virus must be passaged in the presence of increasing amounts of antibody.
- Plate MDCK cells in a 6-well plate at a density of 1 x 106 cells/well and incubate for a minimum of 4 h in a 37 °C incubator (with 5% CO2).
- Dilute the virus stock to 106 PFU/mL or the virus from previous passage in 1x MEM with TPCK-treated trypsin (1 µg/mL) in a 500 µL volume.
- Prepare a single dilution of antibody (0.02 mg/mL for the original passage or higher for all following passages) in 1x MEM with TPCK-treated trypsin in a 250 µL volume.
- Mix 250 µL of diluted virus with 250 µL of diluted antibody (+antibody) or 250 µL of 1x MEM (no antibody control).
- Incubate the virus-antibody mixture for 30 min in a 37 °C incubator (with 5% CO2).
- Aspirate the media using a glass Pasteur pipette and wash the monolayer of cells with 1 mL of 1X PBS.
- Add 500 µL of the mixtures into the wells and incubate in a 37 °C incubator (with 5% CO2) for 1 h.
- After 1 h, supplement the wells with 2 mL of 1x MEM with TPCK-treated trypsin (1 µg/mL).
- Check the cells at 48 h post-infection for signs of cytopathic effect (CPE) on the microscope or perform a hemagglutination assay to detect viral growth26.
- If there is gross CPE in the cultures supplemented with antibody, harvest the supernatant in multiple cryo-tubes, label with the passage number and store at -80 °C.
- Save 100 µL of the supernatant to infect a fresh monolayer of MDCKs with 2 mL of 1x MEM supplemented with TPCK-trypsin and antibody. Remember to include a no antibody control for every passage. NOTE: Increase the concentration of the antibody by two-fold (or at the discretion of the researcher) in the next passage (every two days).
- Increase the concentration of the antibody in each successive passage until virus growth is still viable even with a final concentration of 0.6 mg/mL of antibody. Freeze the multiple vials of supernatant of each passage and store at -80 °C. NOTE: The no antibody control is crucial in verifying the growth of virus from one passage to another. If there is gross CPE in the no antibody control, but no CPE in the +antibody group, this indicates that the concentration of the antibody was too high and no escape variants were generated. If there is gross CPE in the no antibody control, but only moderate CPE in the +antibody group, this indicates the presence of potential escape variants. In the next passage, increase the passage volume to 200 µL of supernatant and maintain the concentration of antibody to increase the likelihood of generating escape variants.
3. Isolation of Escape Variants Through Plaque Purification
Plate MDCK cells in a 6-well plate at a density of 1 x 106 cells/well and incubate for a minimum of 4 h in a 37 °C incubator (with 5% CO2).
Dilute the antibodies in 1x MEM with TPCK-treated trypsin starting at 300 µg/mL in a volume of 250 µL and mix with 250 µL of corresponding escape mutant virus. Virus passaged in the absence of antibody should also be plaque purified.
Aspirate the media from the cells, wash with 1x PBS three times and add the entire 500 µL of the virus-antibody mixture (step 3.2).
Incubate the plates for 1 h in a 37 °C incubator (with 5% CO2) making sure to rock back and forth every 10 min to prevent drying of the monolayer.
Aspirate the virus-antibody mixtures and replenish the wells with overlay agar media containing corresponding amounts of antibody (300 µg/mL; step 3.2).
Incubate the plate for 40-44 h in a 37 °C incubator (with 5% CO2).
Circle the visible plaques with a blue- or black-colored marker to facilitate plaque picking.
Pick four plaques for each escape mutant virus-antibody combination, as well as wild-type viruses that were passaged in MDCK cells or eggs in the absence of antibody.
Resuspend the plaque in 100 µL of 1x PBS.
Inject the entire 100 µL of plaque purified escape mutant virus into 10-day old SPF embryonated chicken eggs.
Incubate the eggs for 40-44 h in a 37 °C incubator (without 5% CO2).
Perform a hemagglutination assay to confirm the presence of virus (step 1.1).
4. Extraction of Viral RNA and Analysis of HA Sequence Variation
Extract the viral RNA from 200 µL of escape mutant virus allantoic fluid with a mono-phasic solution of phenol and guanidine isothiocyanate. CAUTION: Phenol is a volatile liquid reagent that can cause cough, shortness of breath and moderately irritate the skin by contact.
Amplify the HA segment from the viral RNA with the use of a reverse transcriptase and gene-specific primers for the influenza A HA segment27. NOTE: The universal primers for influenza B viruses described in Table 2 amplify both the HA (~1,800 bp) and the NA (~ 1,500 bp)28.
Resolve the RT-PCR product in a 1% agarose gel and cut out the correctly sized band (~1,800 bp).
Gel extract the PCR product using a silica-membrane based purification procedure and send the cDNA out for sequencing.
Identify the amino acid residue required for antibody binding by differentiating the mutations found on putative escape variants and passaged wild-type virus due to either cell culture adaptation or immunological selection.
Clone the PCR product containing the wild-type or escape variant HA into a pCAGGs expression vector (NotI and NheI).
Binding of the antibody to the escape variant HA can be assessed with one of two options (or both) described below.
5. Antibody Binding Analyses of Escape Variants
- Immunofluorescence
- Plate 293T cells at a density of 2 x 104 cells/well in a 96-well plate and incubate in the 37 °C incubator (with 5% CO2) for 24 h.
- Transfect the cells with 0.10 µg/well of the pCAGGS plasmids encoding the escape mutant HA, virus passaged HA, and wild-type HA (unpassaged) with the use of a transfection reagent.
- Incubate the 96-well plates for 48 h in 37 °C incubator (with 5% CO2).
- Fix with 100 µL of 0.5% paraformaldehyde for 15 min at RT. CAUTION: Paraformaldehyde is a volatile liquid reagent that can cause cough, shortness of breath and moderately irritate the skin by contact. It has been designated as a potential human carcinogen. Addition of the reagent should be done in a vented chemical hood.
- Wash with PBS 1x three times. Block with 5% milk in 1x PBS for 1 h at RT.
- Wash with 1x PBS three times. Incubate with 5 µg/mL of antibody of interest for 1 h at RT.
- Incubate with 100 µL of secondary antibody (anti-human or anti-mouse Alexa 488) at a dilution of 1:2,000 in 1x PBS/1% BSA for 1 h at RT in the dark.
- Wash with 1x PBS three times. Observe the cells on a fluorescent microscope for positive or negative staining.
- Fluorescence-activated Cell Sorting (FACS)
- Plate 293T cells at a density of 2 x 105 cells/well in a 6-well plate and incubate at 37 °C (with 5% CO2) for 24 h.
- Transfect the cells with 0.50 µg/well with pCAGGS plasmids encoding the escape mutant HA, virus only passaged HA, and wild-type HA with the use of a transfection reagent.
- Incubate the 6-well plates for 48 h at 37 °C (with 5% CO2).
- After 48 h, aspirate the growth media and wash with 1x PBS two times gently (making sure that the monolayer is undisturbed).
- Harvest the transfected 293T cells with 500 µL of FACS buffer (1x PBS/2% fetal calf serum). NOTE: FACS buffer should be pre-chilled at 4 ºC before use.
- Centrifuge the harvested 293T cells at 300 x g for 5 min at 4 °C.
- Aspirate the FACS buffer, and resuspend with 200 µL of FACS buffer containing mAbs of interest (final concentration of 1 to 5 µg/mL). Incubate at RT for 20 min.
- Centrifuge the cells at 300 x g for 5 min at 4 °C. Wash two times with 500 µL of FACS buffer. Aspirate carefully with a glass Pasteur pipette as to not disturb the pellet.
- Resuspend with 200 µL of FACS buffer containing secondary antibody conjugated to Alexa 488 (final dilution of 1:1,000). Incubate in the dark at 4 °C for 20 min.
- Centrifuge the cells at 300 g for 5 min at 4 °C. Wash two times with 500 µL of FACS buffer and carefully aspirate the wash buffer.
- Resuspend in 500 µL of FACS buffer and assess binding of mAbs and/or polyclonal sera to cells transfected with HA by FACS. NOTE: Remember to include a no mAb/polyclonal sera control as well as an untransfected sample in the experiment.
Representative Results
We have previously used variations of this method to generate escape variants to human and murine mAbs induced by the seasonal influenza virus vaccine, H7N9 vaccination, or sequential DNA/recombinant HA protein vaccination4,5,6,7. As described above, antibodies were first characterized using the HI and microneutralization assays in order to inform us of which specific protocol to continue with next4,5. Antibodies 07-5D03, 07-5F01, 07-5G01, 07-4B03, 07-4E02 and 07-4D05 were found to have HI and neutralization activities against the avian H7N9 virus (A/Shanghai/1/2013) (Table 1), and thus protocol 1 (step 2.1) was utilized. For mAbs with neutralizing that lack HI activity, such as 41-5E04, 045-051310-2B06, 042-100809-2F04 and S6-B01 (Table 1), protocol 2 (step 2.2) was used to generate escape variants. Escape mutant mapping revealed that many of the antibodies recognize critical residues in distinct locations on the viral HA4,5 (Figure 4). While the majority of the HI-positive antibodies have escape mutant residues near previously reported antigenic sites of the H7 HA, the HI-negative antibodies generated escape mutants with point mutations in the stalk region4,5.
| Antibody | HI Activity | NEUT Activity |
| 07-5D03 | + | + |
| 07-5F01 | + | + |
| 07-5G01 | + | + |
| 07-4B03 | + | + |
| 07-4E02 | + | + |
| 07-4D05 | + | + |
| 41-5E04 | - | + |
| 045-051310-2B06 | - | + |
| 042-100809-2F04 | - | + |
| S6-B01 | - | + |
Table 1: Table of antibody HI and neutralization activity. Ten H7-specific mAbs isolated from individuals vaccinated with an experimental H7N9 vaccine exhibit different in vitro antiviral activities5.
| Forward Primer (5' to 3') | Reverse Primer (5' to 3') | Thermocylcer conditions | |
| IAV | TATTCGTCTCAGGGAGCAAAAGCAGGGG | ATATCGTCTCGTATTAGTAGAAACAAGGGTGTTTT | 42 ºC for 60 min, 94 ºC for 2 min/5 cycles of 94 ºC for 20 s, 50 ºC for 30 s and 68 ºC for 3 min 30 s, followed by 40 cycles of 94 ºC for 20 s, 58 ºC for 30 s, and 68 ºC for 3 min 30 s with a final extension time at 68 ºC for 10 min |
| IBV | GGGGGGAGCAGAAGCAGAGC | CCGGGTTATTAGTAGTAACAAGAGC | 45 ºC for 60 min, 55 ºC for 30 min, 94 ºC for 2 min/5 cycles of 94 ºC for 20 s, 40 ºC for 30 s and 68 ºC for 3 min 30 s, followed by 40 cycles of 94 ºC for 20 s, 58 ºC for 30 s, and 68 ºC for 3 min 30 s with a final extension time at 68 ºC for 10 min |
Table 2: Universal influenza virus primers. Primer pairs for the amplification of the HA segments of influenza A27 and B28 viruses and their respective thermocycler conditions.
Figure 1: HI assay. (A) A schematic for the setting up a HI assay to test the activity of two mouse H1-specific mAbs 7B2 (head-specific) and 6F12 (stalk-specific) using a 96-well V-bottom plate, and (B) an example of the results of an HI assay23. Please click here to view a larger version of this figure.
Figure 2: Microneutralization assay. A schematic for setting up a microneutralization assay to test the activity of two human mAbs 4D055 and CR911417. Please click here to view a larger version of this figure.
Figure 3: Generation of escape mutants. The methodology suggested will be dependent on the HI and the microneutralization activity exhibited by the antibody. The generation of escape mutants against (A) neutralizing HI-positive antibodies may require a single passage in eggs, while (B) neutralizing HI-negative antibodies may involve multiple passages with increasing antibody amounts in cell tissue culture. Please click here to view a larger version of this figure.
Figure 4: An example of an epitope map of the novel avian H7N9 HA generated with escape mutant variants. Vaccine-induced antibodies isolated from individuals vaccinated with a candidate H7N9 influenza A vaccine were used to generate escape mutant variants. Each residue indicated in red represents the location of critical amino acids required for efficient binding of a mAb. Data were adapted from Dunand-Henry et al., 20154. Please click here to view a larger version of this figure.
Discussion
Although the majority of residues identified via escape mutants have been accurate, one of the major caveats of this approach is that point mutations of escape variants may not necessarily map within the molecular footprint of the antibody as determined by structural analyses. This is due to the ability of a mutation at a certain residue to lead to a conformational change distal to the location of the mutated residue, analogous to an allosteric effect. Another limitation is that this methodology can only be implemented for neutralizing antibodies; antibodies that lack in vitro selective pressure will not lead to escape mutants. However, this limitation can be overcome with the use of a panel of escape variants generated by previously characterized neutralizing antibodies. Tan et al. used an escape variant of a neutralizing mAb to the H7N9 virus to map the epitope of a non-neutralizing antibody7.
Nonetheless, elucidating the epitopes of antibodies through the generation of escape variants provides a viable alternative to crystallography and cryo-electron microscopy, both of which require an extensive investment of equipment. Other alternatives are to determine the minimal binding region of mAbs using alanine scanning or peptide scanning/truncation mutants. Alanine scanning mutagenesis may require a significant amount of work in generating a large number of variants during screening29, while peptide scanning is limited to linear epitopes30. The method described in this protocol requires no special equipment or technique and in fact, makes use of existing in vitro neutralization assays modified to generate escape variants of the antibodies of interest.
The protocol for generating escape variants that require multiple passages (e.g., stalk-specific antibodies) is highly dependent on the starting concentration of antibody in passage 0. It is better to err on the side of caution and start at a log to half a log lower than the half maximal inhibitory concentration of an antibody and allow for robust virus growth. The researcher can speculate that a high titer virus culture in the presence of low immunological pressure will have a large genetic variation in the viral population. Escape variants can be selected for by gradually increasing the antibody concentration in the following passages. In the event that the virus growth decreases, the amount of viral supernatant can be increased in the next passage while maintaining the same amount of antibody concentration in the previous passage.
The aim of a majority of universal influenza vaccines is to elicit a robust antibody response towards the stalk region of the HA. The analyses of escape variants to stalk-specific antibodies are important in defining the relationship between influenza virus fitness and immunological pressure. Interestingly, escape mutant viruses resulting from stalk-specific mAbs were all attenuated in vivo in murine LD50 studies4. These studies provide a strong case for stalk-based vaccination platforms. Additionally, this protocol could be used to identify escape mutants to other anti-viral compounds, such as small molecule inhibitors. Finally, this methodology is not limited to influenza virus surface glycoproteins, but may also be more widely applied to determine the epitopes of other viral glycoproteins.
Disclosures
The authors declare no conflicts of interest.
Acknowledgments
This project has been funded in part with federal funds from the National Institute of Allergy and Infectious Diseases, National Institutes of Health, Department of Health and Human Services, under CEIRS contract HHSN272201400008C (F.K.); NIH U19AI109946-01 (F.K.); and P01AI097092-04S1 (P.E.L.).
References
- Shaw ML, Palese P. Orthomyxoviridae: the viruses and their replication. 2013.
- Nelson MI, Holmes EC. The evolution of epidemic influenza. Nat Rev Genet. 2007;8(3):196–205. doi: 10.1038/nrg2053. [DOI] [PubMed] [Google Scholar]
- Lauring AS, Andino R. Quasispecies Theory and the Behavior of RNA Viruses. PLoS Pathog. 2010;6(7):e1001005. doi: 10.1371/journal.ppat.1001005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Henry Dunand CJ, Leon PE, et al. Preexisting human antibodies neutralize recently emerged H7N9 influenza strains. J Clin Invest. 2015;125(3):1255–1268. doi: 10.1172/JCI74374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dunand CJH, Leon PE, et al. Both Neutralizing and Non-Neutralizing Human H7N9 Influenza Vaccine-Induced Monoclonal Antibodies Confer Protection. Cell Host Microbe. 2016;19(6):800–813. doi: 10.1016/j.chom.2016.05.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tan GS, Lee PS, et al. Characterization of a Broadly Neutralizing Monoclonal Antibody That Targets the Fusion Domain of Group 2 Influenza A Virus Hemagglutinin. J Virol. 2014;88(23):13580–13592. doi: 10.1128/JVI.02289-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tan GS, Leon PE, et al. Broadly-Reactive Neutralizing and Non-neutralizing Antibodies Directed against the H7 Influenza Virus Hemagglutinin Reveal Divergent Mechanisms of Protection. PLoS Pathog. 2016;12(4):e1005578. doi: 10.1371/journal.ppat.1005578. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith K, Garman L, et al. Rapid generation of fully human monoclonal antibodies specific to a vaccinating antigen. Nat Protoc. 2009;4(3):372–384. doi: 10.1038/nprot.2009.3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steel J, Lowen AC, et al. Influenza virus vaccine based on the conserved hemagglutinin stalk domain. mBio. 2010;1(1) doi: 10.1128/mBio.00018-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krammer F, Pica N, Hai R, Margine I, Palese P. Chimeric hemagglutinin influenza virus vaccine constructs elicit broadly protective stalk-specific antibodies. J Virol. 2013;87(12):6542–6550. doi: 10.1128/JVI.00641-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang TT, Tan GS, et al. Vaccination with a synthetic peptide from the influenza virus hemagglutinin provides protection against distinct viral subtypes. Proc Natl Acad Sci U S A. 2010;107(44):18979–18984. doi: 10.1073/pnas.1013387107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Impagliazzo A, Milder F, et al. A stable trimeric influenza hemagglutinin stem as a broadly protective immunogen. Science. 2015;349(6254):1301–1306. doi: 10.1126/science.aac7263. [DOI] [PubMed] [Google Scholar]
- Krammer F, Palese P, Steel J. Advances in Universal Influenza Virus Vaccine Design and Antibody Mediated Therapies Based on Conserved Regions of the Hemagglutinin. Current Topics in Microbiology and Immunology. 2014. [DOI] [PubMed]
- Wohlbold TJ, Nachbagauer R, Margine I, Tan GS, Hirsh A, Krammer F. Vaccination with soluble headless hemagglutinin protects mice from challenge with divergent influenza viruses. Vaccine. 2015;33(29):3314–3321. doi: 10.1016/j.vaccine.2015.05.038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ekiert DC, Bhabha G, et al. Antibody recognition of a highly conserved influenza virus epitope. Science. 2009;324(5924):246–251. doi: 10.1126/science.1171491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ekiert DC, Friesen RHE, et al. A Highly Conserved Neutralizing Epitope on Group 2 Influenza A Viruses. Science. 2011;333(6044):843–850. doi: 10.1126/science.1204839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dreyfus C, Laursen NS, et al. Highly Conserved Protective Epitopes on Influenza B Viruses. Science. 2012;337(6100):1343–1348. doi: 10.1126/science.1222908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tran EEH, Podolsky KA, et al. Cryo-electron Microscopy Structures of Chimeric Hemagglutinin Displayed on a Universal Influenza Vaccine Candidate. mBio. 2016;7(2):e00257. doi: 10.1128/mBio.00257-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wiley DC, Wilson IA, Skehel JJ. Structural identification of the antibody-binding sites of Hong Kong influenza haemagglutinin and their involvement in antigenic variation. Nature. 1981;289(5796):373–378. doi: 10.1038/289373a0. [DOI] [PubMed] [Google Scholar]
- Gerhard W, Yewdell J, Frankel ME, Webster R. Antigenic structure of influenza virus haemagglutinin defined by hybridoma antibodies. Nature. 1981;290(5808):713–717. doi: 10.1038/290713a0. [DOI] [PubMed] [Google Scholar]
- Jackson DC, Murray JM, White DO, Gerhard WU. Enumeration of antigenic sites of influenza virus hemagglutinin. Infect Immun. 1982;37(3):912–918. doi: 10.1128/iai.37.3.912-918.1982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matsuzaki Y, Sugawara K, et al. Epitope Mapping of the Hemagglutinin Molecule of A/(H1N1)pdm09 Influenza Virus by Using Monoclonal Antibody Escape Mutants. J Virol. 2014;88(21):12364–12373. doi: 10.1128/JVI.01381-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tan GS, Krammer F, Eggink D, Kongchanagul A, Moran TM, Palese P. A pan-H1 anti-hemagglutinin monoclonal antibody with potent broad-spectrum efficacy in vivo. J Virol. 2012;86(11):6179–6188. doi: 10.1128/JVI.00469-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geneva: World Health Organization. WHO manual on animal influenza diagnosis and surveillance. 2002.
- Brauer R, Chen P. Influenza Virus Propagation in Embryonated Chicken Eggs. J Vis Exp. 2015. p. e52421. [DOI] [PMC free article] [PubMed]
- Martínez-Sobrido L, García-Sastre A. Generation of Recombinant Influenza Virus from Plasmid DNA. J Vis Exp. 2010. p. e2057. [DOI] [PMC free article] [PubMed]
- Hoffmann E, Stech J, Guan Y, Webster RG. Universal primer set for the full-length amplification of all influenza A viruses. ArchVirol. 2001;146(12):2275–2289. doi: 10.1007/s007050170002. [DOI] [PubMed] [Google Scholar]
- Zhou B, Lin X, et al. Universal Influenza B Virus Genomic Amplification Facilitates Sequencing, Diagnostics, and Reverse Genetics. J Clin Microbiol. 2014;52(5):1330–1337. doi: 10.1128/JCM.03265-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sidhu SS, Kossiakoff AA. Exploring and designing protein function with restricted diversity. Curr Opin Chem Biol. 2007;11:347–354. doi: 10.1016/j.cbpa.2007.05.001. [DOI] [PubMed] [Google Scholar]
- Chen C-W, Chang C-Y. Peptide Scanning-assisted Identification of a Monoclonal Antibody-recognized Linear B-cell Epitope. J Vis Exp. 2017. p. e55417. [DOI] [PMC free article] [PubMed]