

Abstract
Purpose
It is well known that the dopaminergic signaling pathway plays a pivotal role in the control of axial elongation. Much research has shown that retinal dopamine (DA) is decreased in experimental myopia, but the exact alteration in DA quantity underlying the myopia model induced by flickering light (FL) has not yet been fully elucidated. Therefore, in this study, we first attempted to prove the feasibility of the myopia model induced by FL and then to determine whether and how DA and its receptors changed in myopia induced by FL.
Methods
Forty-five 2-week-old guinea pigs were randomly divided into three groups, as follows: the control group, form-deprivation myopia (FDM) group, and FL-induced myopia (FLM) group. Animals in the control and FDM groups were raised under normal illumination, and the right eyes of the FDM group were covered with semitransparent hemispherical plastic shells serving as eye diffusers. Guinea pigs in the FLM group were raised under illumination with a duty cycle of 50% at a flash rate of 0.5 Hz. The refraction, axial length (AL), and corneal radius of curvature (CRC) were measured using streak retinoscopy, A-scan ultrasonography, and keratometry, respectively, before and after 2, 4, 6, and 8 weeks of treatment. The contents of DA, 3,4-dihydroxyphenylacetic acid (DOPAC), and homovanillic acid (HVA) in the retina, vitreous body, and RPE were measured at the end of the 8-week experiment using high-performance liquid chromatography (HPLC). The numbers of retinal D1 DA receptor (D1DR) and D2 DA receptor (D2DR) were evaluated via immunohistofluorescence and western blot assay.
Results
The refraction of the FLM group became more myopic throughout the experimental period, which was mainly indicated by decreased refraction and a longer AL compared with the control group (p<0.05). The contents of DA, DOPAC, and HVA in the retina, vitreous body, and RPE of the FLM group were significantly increased, but decreased in the FDM group, compared with those of the control group (both p<0.05). Like form-deprived eyes, the expressions of retinal D1DR and D2DR in FL eyes were significantly upregulated compared with controls (p<0.05).
Conclusions
Myopia can be induced by 0.5-Hz FL in guinea pigs at puberty. Contrary to FDM, dopaminergic neuron activity and DA release were significantly elevated in FLM. Like in FDM, the expressions of D1DR and D2DR were upregulated in FLM. Thus, the results of our study may further demonstrate that the DA system is associated with the development of myopia.
Introduction
Myopia occurs when the ocular axial length (AL) is too long for the refractive power of the lens and cornea, which in turn causes the image of distant objects to be focused in front of the photoreceptor layer of the retina, so that the person cannot see things in the distance clearly. With the growing prevalence of myopia in many parts of the world, especially in Asia, attention to this issue has been increasing. Two forms of experimental myopia have been established so far, as follows: form-deprivation myopia (FDM) and lens-induced myopia (LIM). Both forms of myopia have been studied using diffusers [1] or lid suture [2] to produce myopia in a variety of animal models, including tree shrews [3], marmosets [4], chicks [5], mice [6], and rhesus monkeys [7]. LIM is induced by placing a negative lens before animals’ eyes to disturb image formation and force the image to be formed behind the retina, which subsequently induces extension of AL, thereby leading to axial myopia [8].
Recently, there has been increasing evidence demonstrating that flickering light (FL) can influence the emmetropization process. Schwahn and Schaeffel [9] reported that the eyes of chicks raised in FL were more hyperopic with significantly flattened corneas compared with those of chicks raised in normal lighting. In contrast, by comparing refraction errors of strobe-reared cats with those of normal cats, Cremieux et al. [10] concluded that the strobe-reared cats were significantly more myopic than the normal cats were. Crewther et al. [11] found that chicks raised under the condition of a non-square wave profile with low-frequency (1, 2, and 4 Hz) temporal luminance modulation exhibited a general myopic shift in refractive compensation and enhanced ocular growth of lens, whether positive, plano, or negative lenses were used. However, these luminance modulations had no effect on refraction or ocular parameters without lenses. After comparing strobe-reared guineas pigs with a control group under steady illumination, Yue et al. [12] reported that in the five FL-induced myopia (FLM) groups established using FL of different frequencies (5, 1, 0.5, 0.25, and 0.1 Hz), the incidence of myopic shift and axial eye length both increased.
Studies on animal models of myopia have concluded that dopamine (DA), a neurotransmitter released exclusively by a population of amacrine or interplexiform cells, is critical during the development of experimental myopia [13,14]. In vivo, once DA is released from the retinal amacrine and interplexiform cells, monoamine oxidase (MAO) can remove the amine group on DA to produce 3,4-dihydroxyphenylacetic acid (DOPAC). Shortly after DOPAC is formed, catechol-0-methyltransferase (COMT) can add a methyl group to DOPAC to make homovanillic acid (HVA). Hence, the release rates of retinal DA from retinal amacrine cells can be reflected by vitreal DA concentrations [15,16]. In 1995, Guo [17] demonstrated a decrease in the levels of DA and its major metabolite, DOPAC, but an increase in AL, after applying negative lenses to chicken eyes. Like in LIM, retinal DA and DOPAC levels were found to drop in FDM [18]. Other research conducted on chickens [19] and rhesus monkeys [20] also demonstrated that DA concentrations in the retina decreased with the induction of FDM. Furthermore, with intravitreal injection of apomorphine (the DA nonselective agonist), the development of deprivation-induced myopia or negative LIM was prevented [21]. Likewise, exogenous DA [22] and its precursor, levodopa [13], had similar protective effects against myopia development induced by form deprivation (FD). Decreased levels of DA and its two metabolites have already been observed in FDM, but the levels of DA and its metabolites in FLM have not been previously studied.
It is considered that the influence of apomorphine on the inhibition of FDM involves binding with the retinal D2-like receptors. Recently, a study successfully indicated that the inhibition of D2 receptors could also slow down the development of spontaneous myopia in albino guinea pigs [23]. Although several studies have displayed that the inhibitory effects of DA agonists on FDM and LIM were mediated through stimulation of D2 receptor, several other reports have further demonstrated that the balance of D2 and D1 receptor activation is also important. Furthermore, multiple examples in the literature have indicated that the number of DA receptors (D1 and D2) in FDM increased [24,25]. Thus, in summary, these studies suggest that binding of DA to its receptors has an important role in the development of experimental myopia.
FDM is a well established myopia model, and it has been employed in many studies on the pathogenesis of myopia; in addition, the establishment of an FLM model can provide a new tool to study the mechanism of myopia from a different perspective. There are little available data concerning the role of the dopaminergic system in FL-induced myopia. Particularly, it remains unclear whether the levels of retinal DA and its metabolites and receptors undergo similar changes in FLM to those in FDM. Based on the abovementioned studies, in our research, the myopia model was first established by applying FL stimulation, and subsequently, the concentrations of these compounds were measured separately in different layers of the eye (including the retina, vitreous body, and RPE) to determine what changes they exhibited in the FLM model.
Methods
Animals
Two-week-old guinea pigs (Cavia porcellus, English short-hair stock, tricolor strain, n=45) were obtained from the Laboratory Animal Center at Fudan University in Shanghai, China. Guinea pigs were raised at a temperature of 20–22 °C, at a relative humidity of 55 to 65%, and they had free access to sufficient and fresh food and water, which was supplied daily.
The treatment and care of the animals adhered to the Association for Research in Vision and Ophthalmology (ARVO) Statement for the Use of Animals in Ophthalmic and Vision Research. The animal research was approved by the Local Animal Care and Ethics Committee at Jinshan Hospital of Fudan University, Shanghai, China.
Experimental design
Forty-five 2 -week-old guinea pigs were randomly divided into three groups, as follows: the control group, FDM group, and FLM group. A maximum of five animals were housed in standard mouse cages (50 × 40 × 60 cm with a mesh size of 1.5 × 5.0 cm), and each cage was placed in a custom, ventilated carton. Each carton had a corresponding opaque lid that was totally covered to ensure complete darkness inside the carton. All guinea pigs were raised in a dark room with no light from outside. Every cage in the FLM group was totally covered to ensure complete darkness from the outside of the carton. Meanwhile, the animals in the control and FDM groups were maintained on a 12 h:12 h light-dark cycle (600 lx; lights on at 6 AM and off at 6 PM). The experiment lasted for 8 weeks.
For the FLM group (n=15), four light-emitting diode (LED) lights (narrow spectrum, 505 nm; color temperature, 2,850 K) were installed in the four upper corners of each cage at a height of 50 cm from the bottom of each cage to achieve uniform lighting intensity at the cage bottom. The illumination was manipulated via temporal luminance modulation of 0.5 Hz and 600 lux produced by a function generator linked to the LED lights. The function generators (Yinuo Automation Co., LTD, Changsha, China; linear output; analog signal; alternating-current pulse, 220 V) were placed on the outside of the carton. All animals in the FLM group received the illumination of 600 lux with a duty cycle of 50% at a flash rate of 0.5 Hz (i.e., 1 s of dark phase/2 s) from 6 AM–6 PM for 8 weeks. During each flicker cycle, luminance varied between a maximum of 600 lux and minimum of 0 lux.
In the FDM group (n=15), monocular deprivation was induced using translucent plastic diffusers made from white moldable plastics glued to the guinea pigs’ right eyes. The diffuser was placed sufficiently far from the eyelid so that it would not interfere with the normal functions of the eyelid. The transmission of the eye diffusers to light was 60%. The plastic shells were checked at least twice a day to ensure secure attachment of the shells to the right eyes. Finally, the control group (n=15) was kept in the cartons without any additional treatments.
Optical and biometric measurements
All measurements were performed at 2-week intervals, that is, before the start of the treatments (labeled “week 0”) and after 2, 4, 6, and 8 weeks of treatment. All measurements were performed by a research optometrist with help from an assistant. The identities of the different groups were masked. Although data were available for the left eyes, only the right eyes (except the refraction measurement) were examined to avoid mixing independent and dependent data.
The refractive errors were examined with a streak retinoscope at a working distance in a dark environment, using lens bars to neutralize the two principal meridians. One drop of tropicamide ophthalmic solution (1 mg/ml) was administered every 5 min up to four times to achieve cycloplegia and a completely dilated pupil before the measurement. Animals were only slightly restrained by controlling their bodies without anesthesia. The refraction of each eye was obtained by averaging three collected sets of the measurements and expressed as spherical equivalents. Before the AL measurement, the guinea pigs were anesthetized via the administration of 0.5% proparacaine hydrochloride (Alcon, Belgium), and the AL of each right eye was measured using A-scan ultrasonography (11 MHz; SW1000, Suowei Co., Ltd., Tianjin, China). Each value obtained in the final experimental data was the average of five separate measurements. The corneal radius of curvature (CRC) was measured in alert guinea pigs with a keratometer (OM-4; Topcon, Tokyo, Japan), combined with an +8.0 D aspherical lens on the anterior surface. Final data represented the mean of the horizontal and vertical measurements. A set of stainless steel ball bearings was used for calibration, and the CRC was determined by averaging three readings from the balls at different places [26]. The CRC for each animal was measured in triplicate.
High-performance liquid chromatography
After biometric measurements, animals were killed with CO2 at the end of week 8, and their vitreous body, retina, and RPE were carefully dissected from the eyecups under a microscope and quick frozen in liquid nitrogen to await further measurements on the levels of DA, DOPAC, and HVA; these parameters were assayed using high-performance liquid chromatography (HPLC) with electrochemical detection. Chemicals and reagents: All chemicals were of HPLC grade, and ultrapure water was produced using an Ultra-Pure water system (Millipore, Billerica, MA). DOPAC, DA hydrochloride, HVA, and sodium octanesulfonate (OSA) were all purchased from the Sigma-Aldrich company (St. Louis, MO).
HPLC analysis
Preparation of Standard Solutions and Reagents: Stock standard solutions of 1 mmol/l were prepared in a solution of 0.2 N perchloric acid and stored at 4 °C before usage. These stock solutions were diluted to the desired concentration with mobile phase. Instrumentation and Conditions: The Agilent 1200 series neurotransmitter analyzer consists of a G1367B autosampler, a G1312A binary pump, a G1322A degasser, and the ANTEC DECRARD SDC (Antec, Zoeterwoude, Netherlands) electrochemical detector equipped with a Sencell including a 2-mm glassy carbon working electrode.
HPLC Conditions
The samples’ supernatants were injected into an Acclaim C18 column (2.2 μm, 2.1 × 100 mm; Thermo Fisher Scientific, Waltham, MA) at 38 °C. Separations were performed at a flow rate of 0.2 ml/min using a mobile phase of phosphate buffer (PB; 0.05 mM EDTA, 1.7 mM orthosilicic acid [OSA], 90.0 mM Na2HPO4, 50.0 mM citric acid). The voltage for the detection cell was set at +700 mV, with the voltage for the guard cell set at +750 mV. The data were collected and analyzed by ChemStation (Agilent Technologies, Santa Clara, CA). Peaks and relative concentrations were identified by comparison with known external standards.
Immunohistofluorescence
Guinea pigs were deeply anesthetized with pentobarbital and perfused transcardially with physiologic saline followed by 150–200 ml of 4% paraformaldehyde in 0.1 M PB (pH 7.4). The eyes were enucleated quickly, and we used 8–0 silk thread to mark the midperiphery of the eyeball. Subsequently, once anterior segments of the eyes were removed, the remnants of the eyecups were immediately fixed in fresh 4% paraformaldehyde in 0.1 M PB (pH 7.4) for 20 min and chilled sequentially in 10%, 20%, and 30% sucrose (wt/vol) in 0.1 M PB at 4 °C. The eyecups embedded in optimal cutting temperature (OCT; Miles, Elkhart, IN) compound were frozen using liquid nitrogen; they were then sectioned at a thickness of 15 μm along the midperiphery line on a cryostat. Eight slices were obtained for each eyeball. The sections were mounted on gelatin chromium–coated slides.
The sections prepared according to the procedure mentioned above were first blocked in 2% donkey serum in 0.01M PBS(0.2M Na2HPO4 0.2M NaH2PO4, 0.09% NaCl, pH 7.4) with 0.1% Triton (PBST) and then incubated overnight at 4 °C in a mixture of rabbit anti-D1 DA receptors (D1DR; diluted 1:200; Abcam; CB, UK; catalog reference: ab81296) and mouse anti-D2 DA receptors (D2DR; diluted 1:100; Santa Cruz, Santa Cruz, CA; catalog reference: sc-5303). Following this, they were incubated for another 90 min at room temperature in a mixture of secondary antibodies, namely goat anti-rabbit Alexa 488 (Invitrogen; Carlsbad, CA; catalog reference: A11070) and donkey anti-mouse Alexa 594 (Invitrogen; catalog reference: A21203), which were both diluted to 1:800. After repeating the incubation twice, the sections were rinsed three times in PB, mounted on glass slides, and coverslipped with Vectashield (Vector, Burlingame, CA). Images of D1DR- and D2DR-stained retinal sections were taken using a Leica SP5 confocal laser scanning microscope (Leica, Mannheim, Germany) at 63× magnification. The Image-Pro Plus 6.0 software program was used for the analysis of retinal micrographs (Step 1: Calibration Measure–Calibration–Intensity- new–Grey Level or Std. Optical Density—Close; Step2:Measure Density Measure–Count/Size—measure–select measurements–Density(red)–Edit–Draw/Merge Object—Trace or Rectangular AOI–Choose 10 places—OK—View–Measurement Data–File–Data to clipboard–open an excel file—paste to Excel—Analyze with SPSS Statistics 20).
Western blot
To verify the data obtained with immunohistofluorescence, quantitative western blot assays for D1DR and D2DR expressions in the retinas were also performed. The guinea pigs were sacrificed, and the retinal tissues were quickly collected and stored on ice. Subsequently, the tissues were homogenized in 2% 3-[(3-Cholamidopropyl)-dimethyl- ammonio]-1-propane sulfonate; N,N-Dimethyl-N-(3-sulfopropyl)-3-[[(3a,5b,7a,12a)-3,7,12-trihydroxy-24-oxocholan-24-yl] amino]-1-propanaminium inner salt (CHAPS) buffer containing 10 mM sodium phosphate (pH 7.2), 1% sodium deoxycholate, 0.15 M sodium chloride, and protease inhibitor cocktail; they were then centrifuged at 11,280 × g for 10 min at 4 °C. The protein concentration was determined using a MicroBCA protein assay kit (Beyotime Biotechnology, Shanghai, China). Then, the tissue homogenate (50 μg protein equivalent) of the entire retina of each guinea pig was boiled at 100 °C in sodium dodecyl sulfate (SDS) sample buffer for 5 min, electrophoresed on 10% SDS-polyacrylamide gel, and transferred to a polyvinyldifluoridine membrane (Bio-Rad). The membranes were blocked for 2 h at room temperature in blocking buffer, consisting of 20 mM Tris-HCl (pH 7.4), 137 mM NaCl, 0.1% Tween-20, and 5% nonfat milk. This was followed by overnight incubation in a buffer containing the monoclonal rabbit antibody against D1DR (1:1,000, Abcam; catalog reference: ab81296) or mouse antibody against D2DR (1:1,000, Santa Cruz; catalog reference: sc-5303) overnight at 4 °C. Bext, membranes were washed with 0.1% Tween-20, and then treated with goat anti-rabbit immunoglobulin G (IgG; Invitrogen, USA; catalog reference: A11070) or donkey anti-mouse IgG (Invitrogen, USA; catalog reference: A21203) conjugated to alkaline phosphatase (1:1,000 dilution) for 1 h at 37 °C. Stripping filters and reprobing for glyceraldehyde 3-phosphate dehydrogenase (GAPDH) were performed for normalization. Controls for nonspecific binding were determined by omission of the primary antibody. Films were scanned with a film scanner (Image Master VDS; Amersham Biosciences Inc., Piscataway, NJ) and subsequently analyzed to measure the optical densities of immunostained bands on the film using an image-processing and analysis system (Q570IW; Leica).
Data analysis
Only data from the right eyes were registered for analysis in this study, except refraction measurement. The change was defined as the total amount of variation during the experimental period, and this was determined using the difference between the data measured at the first examination (week 0) and those from the last examination (week 8). Parametric statistical analyses were performed using SPSS Statistics 20 (SPSS, Inc., IBM, Chicago, IL) and GraphPad Prism 5 (GRAPHPAD Software, Inc., San Diego, CA). Multivariate repeated-measurements ANOVA (ANOVA) was applied to compare the refractive error, AL, and CRC among the different groups. One-way ANOVA was used to compare the expression of neurotransmitters and DA receptors. The results were expressed as mean ± standard error of the mean (SEM); p<0.05 was regarded as the indicator of statistical significance.
Results
Optical and biometric measurements
No significant differences in refractive error, AL, or CRC were observed among the groups (one-way ANOVA) before the experiment, nor did these measurements appear in both eyes of any individual guinea pig (p>0.05). Thus, the right eyes were validated as experimental objects.
The refractive error of right eyes (FD eyes) in the FDM group and of the bilateral eyes (FL eyes) in the FLM group became more myopic throughout the experimental period compared to the corresponding eyes of the control group (Figure 1). In week 4, significant differences were observed in FD eyes compared with FL and control eyes (the right eyes of the two groups; Figure 1A, p<0.01). There were statistically significant differences among the right eyes of the FDM, FLM, and control groups from week 6 (all p<0.05). After 8 weeks of treatment, the changes in the refractive status of control right eyes, FD eyes, and FL right eyes were −2.28±0.34 D, −13.18±1.33 D, and −8.14±0.60 D, respectively, and significant differences existed among the three groups (Figure 1B, p<0.01). When considering the left eyes of the three groups, the refractive error in the left eyes of guinea pigs raised in FL was significantly more myopic than that in the other two groups starting from week 6 (Figure 1C, p<0.01). No significant differences were detected between the deprived contralateral eyes and control left eyes during the experimental period (p>0.05). After 8 weeks of treatment, changes in refractive status of the control left eyes, FD contralateral eyes, and FL left eyes were −4.23±0.41 D, −4.31±0.31 D and −7.70±0.74 D, respectively, and the refraction changes in the FL left eyes were much larger than in the left eyes in the other two groups (Figure 1D, p<0.01); however, no significant difference was shown between the control left eyes and FD contralateral eyes (p>0.05).
Figure 1.

Refractive comparisons in bilateral eyes of the control, FLM, and FDM groups during the 8-week treatment period. A: Refraction of the right eyes in the three groups throughout the experiment. B: Refractive changes of the right eyes in the three groups after 8 weeks of treatment. C: Refraction of the left eyes in the three groups throughout the experiment. D: Refractive changes of the left eyes in the three groups after 8 weeks of treatment. The calculation of change was based on the refraction measured at the end of week 8 minus that obtained at week 0. Similar calculations were applied for AL and CRC measurement (FLM: flickering light (FL)-induced myopia, FDM: form-deprivation myopia, AL: axial length, CRC: corneal radius of curvature. *p<0.05 versus control group (n=15 for each group); **p<0.01 versus control group; #p<0.05 versus FDM group; ##p<0.01 versus FDM group; data are expressed as mean ± standard error of the mean [SEM]).
The ALs in FD eyes and FL right eyes were longer compared with those in the control right eyes after 2 weeks of treatment at each time point (Figure 2A, p<0.01), but no significant differences were observed between the FD eyes and FL right eyes (p>0.05). The AL changes in the right eyes of the control, FDM, and FLM groups were 1.81±0.03 mm, 2.85±0.11 mm, and 2.67±0.14 mm, respectively, by the end of treatment (Figure 2B). Compared to the right eyes of the control group, the length alterations in the right eyes of the FDM and FLM groups were more remarkable (p<0.01), but no significant difference was found between the right eyes of the FDM and FLM groups (p>0.05).
Figure 2.

Comparisons of AL and CRC among the right eyes of the control, FLM, and FDM groups during the 8-week treatment period. A: ALs of the right eyes in the three groups throughout the experimental period. B: AL alterations of the right eyes in the three experimental groups after 8 weeks of treatment. C: CRCs of the right eyes in the three groups throughout the experiment. D: CRC changes of the right eyes in the three experimental groups after 8 weeks of treatment (FLM: flickering light (FL)-induced myopia, FDM: form-deprivation myopia, AL: axial length, CRC: corneal radius of curvature. *p<0.05 versus control group; **p<0.01 versus control group; #p<0.05 versus FDM group; ##p<0.01 versus FDM group; data are shown as mean ± standard error of the mean [SEM]).
The CRC increased in a similar manner among all the right eyes in the three groups during the 8-week experimental period, but no significant differences were found at each time point among the groups until the end of week 8: The FD eyes and FL right eyes displayed greater CRC values when compared with the control right eyes (Figure 2C, p<0.01). In addition, significant differences in CRC changes were displayed between the right eyes of the model groups (FDM and FLM) and the control group (p<0.01) during the experimental period (Figure 2D). However, neither the CRC values at each time point nor CRC variations showed significant differences between the right eyes of the FDM and FLM groups (p>0.05).
Contents of DA and its metabolites
Retinal, vitreal, and RPE levels of DA; the principal DA metabolite, DOPAC; and the secondary DA metabolite, HVA, were assessed using HPLC analysis. Among them, the vitreal DOPAC level is a robust index of retinal DA release [27], and the HVA level is an indirect indicator for DA storage [28,29].
In control eyes, the levels of DA in the retina, vitreous body, and RPE were 82.22±4.68 pg/μl, 11.66±0.67 pg/μl, and 11.64±0.95 pg/μl, respectively. Therefore, the DA contents in the vitreous body and RPE were both approximately 14% of that of the retina. As shown in Figure 3A, by comparing the retinal DA and metabolite levels in the three groups, we found that the levels of DA, DOPAC, and HVA in the retinas of FL eyes were significantly increased compared with those of control right eyes and FD eyes (all p<0.05). In contrast, the levels of DA, DOPAC, and HVA in the retinas of the FD eyes were less than those of the control eyes (all p<0.05). When considering the levels of DA and its two metabolites in the vitreous bodies of the three groups, similar trends in DA, DOPAC, and HVA were detected (Figure 3B). These results indicated that not only retinal DA storage but also retinal DA release decreased significantly in the FDM group, but they increased significantly in the FLM group. We also analyzed these substances in the RPE layer (Figure 3C) and discovered that the levels of DA and its metabolites in the FD and FL eyes were significantly increased when compared to those of the control eyes (all p<0.05). However, no significant differences were observed between the FDM and FLM eyes (p>0.05).
Figure 3.

Levels of DA, DOPAC, and HVA in the eyes of the three groups after the 8-week experiment. A: Retinal levels of DA, DOPAC, and HVA increased in the FLM group but decreased in the FDM group compared with controls (p<0.05). B: Expression of these substances in the vitreous body was comparable to their trends in the retina, whereas the vitreal DOPAC level in the FLM group showed no significant difference compared with that in the control group (p>0.05) and showed a significant difference compared with that in the FDM group (p<0.05). C: Levels of DA and its two metabolites in the RPE were significantly increased in the FDM and FLM eyes compared with control eyes (p<0.05; FLM: flickering light (FL)-induced myopia, FDM: form-deprivation myopia, DA: dopamine, DOPAC: 3,4-dihydroxyphenylacetic acid, HVA: homovanillic acid. *p<0.05 versus control group; #p<0.05 versus FDM group; data are presented as mean ± standard error of the mean [SEM]).
D1DR and D2DR levels
D1DR and D2DR were detected on neurons of the inner and outer layers of the retina. D1DRs are mainly found on retinal ganglion cells (RGCs) and the inner nuclear layers (INLs), while D2DRs are mainly found on RGCs and the inner plexiform layers (IPLs; Figure 4). The most intense label of D1DR and D2DR are likely observed in the FDM group, less in the FLM group, and least in the control group.
Figure 4.
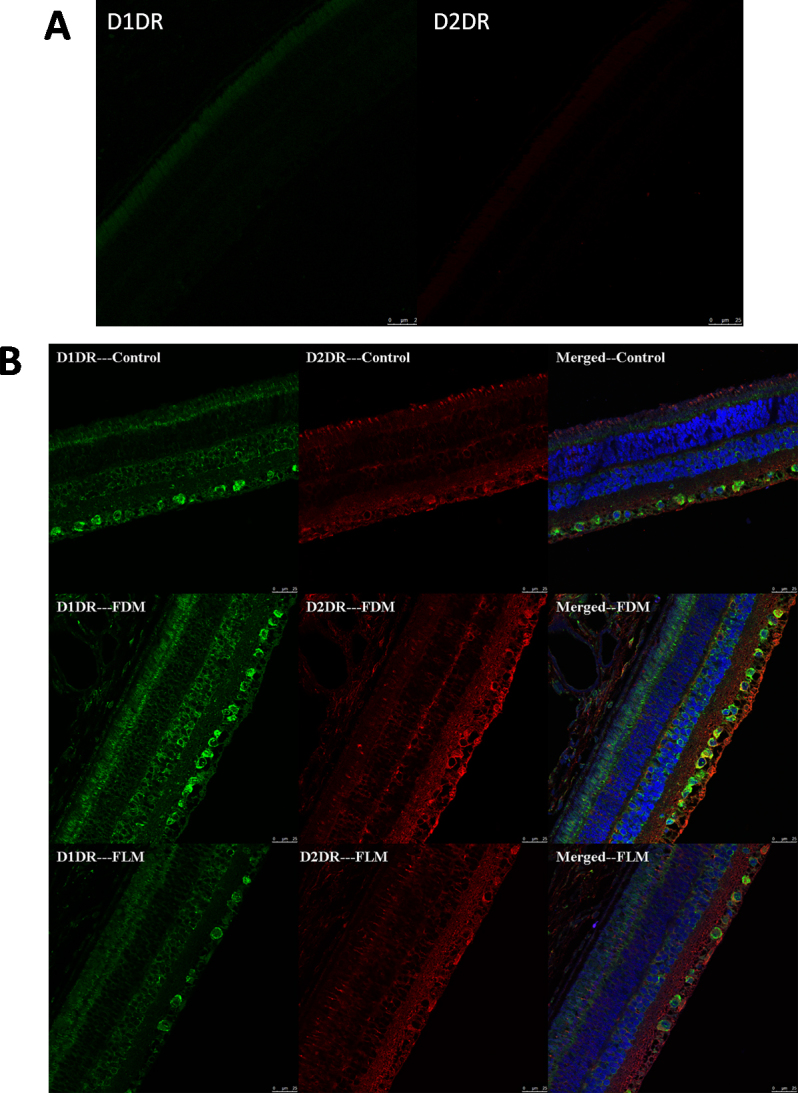
Photographs of retinas stained by immunofluorescence staining of D1 dopamine (DA) receptors (D1DR) and D2 DA receptors (D2DR). Green staining is D1DR, red staining is D2DR, blue staining is the cell nucleus (scale bar: 25 μm). D1DR was mainly found on the retinal ganglion cells (RGCs) and inner nuclear layers (INLs), while D2DR was mainly found on the RGCs and inner plexiform layers (IPL; Figure 4A shows the primary control images for each antibody).
Differences in the distribution of DA receptors in different layers of the retina between the groups were revealed in Figure 5. In the FDM group, the numbers of both D1DR and D2DR in RGCs, D1DR in the INLs, and D2DR in the IPLs increased when compared with those of the control group (all p<0.05). In the FLM group, D1DR in the RGC layer decreased with a simultaneous increase in D2DR, while the number of RGCs coexpressing D1DR and D2DR increased compared with those in the control group (all p<0.05). However, the expressions of D1DR and D2DR in the RGCs of the FLM group were lower than in the FDM group (both p<0.05). Both in the INLs and IPLs of the FLM group, the quantities of D1DRs and D2DRs increased compared with those in the control group, but less than those in the FDM group (all p<0.05). In conclusion, D1DR and D2DR expressions in FLM were less upregulated than those in FDM when both were compared with the control group.
Figure 5.

Levels of D1 and D2 dopamine (DA) receptors in the three groups. A: The number of positive cells in retinal ganglion cells (RGCs). In the FDM group, the numbers of both D1 DA receptors (D1DR) and D2 DA receptors (D2DR) increased compared with the control group (p<0.05); in the FLM group, the numbers of D1DR and D2DR decreased and increased, respectively, compared with the control group (both p<0.05). The numbers of RGCs that coexpressed D1DR and D2DR increased in both the FDM and FLM groups (p<0.05). There were significant differences among the three groups according to the number of two receptors (p<0.05). B: The average optical density of D1DR-positive cells in inner nuclear layers (INLs) was calculated for the three groups. In the INLs and IPLs, the quantities of D1DR and D2DR increased in both the FDM and FLM groups (FLM: flickering light (FL)-induced myopia, FDM: form-deprivation myopia; p<0.05 versus control group), and significant differences were detected among the three groups (p<0.05). C: The average optical density of D2DR-positive cells in the inner plexiform layers (IPL) was calculated for the three groups (*p<0.05 versus control group; #p<0.05 versus FDM group; data are presented as mean ± standard error of the mean [SEM]).
Western blot analysis
To illustrate the differences reflected by immunofluorescence further, the expressions of D1DR and D2DR proteins in the retinas were assessed quantitatively using western blot analysis (Figure 6 and Figure 7). Figure 6 showed that the expressions of D1DR in the FLM and FDM eyes was significantly upregulated (both p<0.05), and the D1DR level of FDM eyes was significantly increased compared to that of FLM eyes (p<0.05). Figure 7 shows that higher D2DR expression levels were revealed in both the FDM and FLM groups than the level in the control group, but a significant difference was only established between FDM and the controls (p<0.05). Hence, western blot shared similar trends with immunofluorescence.
Figure 6.

Analyses of D1 dopamine (DA) receptor (D1DR) expression in the retinas of three groups by western blot. The expression of D1DR in the FLM eyes was less upregulated than that in the FDM eyes compared with the controls (all p<0.05; FLM: flickering light (FL)-induced myopia, FDM: form-deprivation myopia. *p<0.05 versus control group; #p<0.05 versus FDM group; data were presented as mean ± standard error of the mean [SEM]).
Figure 7.

Analyses of D2 dopamine (DA) receptor (D2DR) expression in the retinas of three groups by western blot. The expression of D2DR was upregulated in the FLM and FDM eyes, but only the FDM eyes showed significant differences when compared with the controls (p<0.05; FLM: flickering light (FL)-induced myopia, FDM: form-deprivation myopia. *p<0.05 versus control group; data are presented as mean ± standard error of the mean [SEM]).
Discussion
In this work, myopic shifts were induced in guinea pigs by 0.5-Hz FL stimulation. In contrast to the results obtained in FDM animal models, however, significant increases in retinal, vitreal, and RPE DA, DOPAC, and HVA levels were detected in FLM eyes. Furthermore, the expression of DA receptors (D1 and D2) was upregulated in both the FDM and FLM eyes.
Myopia was induced less rapidly by FL than by FD (week 6 versus week 4), and the absolute refractive error change after 8 weeks of FL exposure was less than that obtained in FDM (−8.14±0.60 D versus −13.18±1.33 D). These results were comparable to those of Yu et al. [30], who showed that the myopic shift induced in mice by FL was less intense and occurred later than that induced by FD. After 8 weeks’ treatment in this study, we found that CRC was larger in both the FDM and FLM groups compared with that in the control group, which is not in line with the previous studies [12,31,32]. The mechanism underlying such visual changes in the cornea remains unknown.
In the normal control eyes of guinea pigs, the contents of DA in vitreous body and RPE were both approximately 14% of that of the retina. These data are in line with those of Ohngemach et al. [15], who found that the vitreal content of DA was about one-tenth of that of the retina in chickens. Levels of retinal DA, tyrosine hydroxylase (the rate-limiting enzyme in the synthesis of DA), and DOPAC dropped during the development of FDM, indirectly indicating that DA synthesis, release, and metabolism fell [19,20,33]. Our data on the contents of DA and its metabolites in the retina, vitreous body, and RPE layer of the FDM group also supported the similar result that dopaminergic neuron activity and retinal DA release decreased significantly in FDM. A significant finding in this study was that the levels of retinal DA and its metabolites in the FLM guinea pig model were increased, as were those in the vitreous body and RPE layer. This result was completely different from those obtained in the FDM model. DA, as a neurotransmitter in the brain and retina that is sensitive to light exposure [34,35], is widely believed to be involved in the control of eye growth and visual signaling [23]. Animal experiments have indicated that DA release is much higher during the day or in a bright environment than at night or in the dark [36,37]. DA and its main metabolite, DOPAC, could be restored to normal levels once the unilateral translucent goggle-covered eyes were uncovered and re-exposed to normal lighting [38]. Steady light inhibited DA release in cats [39], whereas flashing light stimulated its release [40]. In chick eyes, stroboscopic FL of frequencies of 2, 5, 10, 15, and 20 Hz caused significant accumulation of DA, its primary metabolite, DOPAC, and its secondary metabolite, HVA, in the vitreous body compared with the eyes exposed to constant darkness, and this change is a reliable index of DA release increase. Meanwhile, DA cell activation increased markedly with the onset of these stroboscopic stimulations [37]. The disparity in levels of DA and its metabolites in the three layers (i.e., retina, RPE, and vitreous body) observed in the two myopic models (i.e., FLM and FDM) raises two intriguing possibilities: One is that the light transmission in the covered eyes (FD eyes) was weaker than that in the eyes free from covering under the condition of FL; stronger light may result in higher DA levels. The other possibility is that FL pulses may stimulate the dopaminergic amacrine cells more effectively than the steady light does. Light that flickers ON and OFF triggered strong bursts of spikes in phase with periods of light exposure, whereas steady light evoked an initial transient burst of spikes at the light onset that was rapidly silenced by the depolarizing block and remained silent until the depolarizing excitatory response declined to a level that relieved the spike block [41]. It seems that repetitive firing can stimulate other neuronal cell types to release vesicular and increase intracellular Ca2+ due to summated influx, thereby triggering DA release [42].
Recently, several reports have demonstrated that outdoor activities reduced the risk of myopia in both young individuals [43] and animal models [44], which was probably due to light-induced increases in retinal DA release. The notion that the increased DA is accompanied by decreased myopic shift is also supported by findings from the usage of dopaminergic agonists or antagonists in animal eyes. However, the changes in the DA levels found in the three FLM layers seem to be inconsistent with this idea. The different pathogenesis underlying different types of myopia may be attributed to this paradox, although the possibility of an accelerating effect of DA excess in the development of myopia has been raised. Dopaminergic therapy is a main therapy for Parkinson disease today, but it also has been shown to worsen some cognitive functions during treatment; this is presumably due to DA overdose in the brain regions that are relatively DA replete [45,46]. Thus, excess DA may be harmful to the proper functioning of some regions of the body. Because human physiology is maintained in a dynamic balance, with each type of physiologic substance fluctuating within normal ranges, too little or too much is likely to affect normal bodily function. As an example, consider the potassium content; too little or too much potassium can affect the regulation of the acid–base balance and the generation of electrical excitation and conduction. When the eyes are exposed to FL stimulation, DA stores and releases are increased remarkably, which means that an excess of DA is present in the FL eyes, resulting in abnormal ocular growth and development. However, more research should be conducted to shed light on this issue.
We must emphasize that special attention should be paid to the levels of DA and its metabolites in the RPE layer, which were found to be remarkably increased in myopic eyes established either by FD or FL compared with the controls. The hypotheses that local growth modulation is implicated in myopic responses and scleral remodeling and underlies the development of myopia are popular among most researchers [47-49]. The RPE, which separates the choroid from deeper parts of retina, is likely to play a critical role in relaying retinal growth signals to the sclera, as well as to be intimately involved in ocular growth regulation. Research has confirmed that without RPE cells, the inhibitory effect of apomorphine on the proliferation of scleral chondrocytes is small [50]. Based on the current findings, there is a possibility that different retinal signals in the development of FDM and FLM may exist, but at the level of the RPE, there is a common signal that could mediate the common response in ocular growth-regulating signals, thereby ultimately modulating scleral growth.
Dopaminergic signaling in the retina is considered critical during the development of experimental myopia. DA exerts its effects by binding to five different types of receptors (D1–D5) that can be grouped into two different families, namely D1-like receptors (D1 and D5) and D2-like receptors (D2, D3, and D4). DA binding to D1-like receptors activates adenylyl cyclase, leading to an increase in cAMP. In contrast, DA can result in decrease in cAMP by inhibiting adenylyl cyclase via binding to the D2-like receptors [51,52].
To elucidate the possibility that FL may be related to changes in DA activation level by inducing changes in retinal DA receptor levels, immunohistofluorescence and western blot analysis of the samples were performed, and the expressions of retinal DA receptors (D1DR and D2DR) obtained in the deprived eyes and FL-exposed eyes were found to be upregulated. Furthermore, the deprived eyes showed more DA receptor expression than the FL-exposed eyes did. Extensive evidence suggests the important role of D2DR in myopia development [53,54]. However, some recent data have also supported a role for both D1DR and D2DR in the signaling cascades related to the visual control of eye growth [55,56]. Xiao et al. [25] found that deprivation of sharp vision increased the numbers of D1DR and D2DR. Similarly, our research found a role for both D1DR and D2DR in the ocular growth, responding to FD and FL. Thus, it is possible that the number of DA receptors can be regulated during myopia development.
Our finding that myopic shift in guinea pigs induced by FD and by FL produced paradoxical trends in the levels of retinal DA and its metabolites may imply that there are different mechanisms underlying these two types of experimental myopia. However, the exact pathogenesis of FL-stimulated myopia is not yet known. Thus, more research is required to elucidate the mechanisms, especially the dopaminergic signaling, which influences refractive development in FLM, and the common mechanism underlying different experimental myopic models, which may serve as a potential therapeutic target for myopia in the future.
Acknowledgments
This work was supported by Grants from the Shanghai health and family planning committee funded projects of China (No. 201,640,046), Shanghai Municipal Natural Science Foundation of China (13ZR1405800),Shanghai Municipality Health Bureau Youth Project (2013–121) and National Natural Science Foundation of China (No. 81,400,428). The authors convey their thanks to Shuangshuang Zhang, Department of Dermatology, Jinshan Hospital affiliated to Fudan University, Shanghai, China, for the support that improved the efficiency of biometric measurement.
References
- 1.Wallman J, Turkel J, Trachtman J. Extreme myopia produced by modest change in early visual experience. Science. 1978;201:1249–51. doi: 10.1126/science.694514. [DOI] [PubMed] [Google Scholar]
- 2.Raviola E, Wiesel TN. The mechanism of lid-suture myopia. Acta Ophthalmol Suppl. 1988;185:91–2. doi: 10.1111/j.1755-3768.1988.tb02675.x. [DOI] [PubMed] [Google Scholar]
- 3.Norton TT, Amedo AO, Siegwart JJ. The effect of age on compensation for a negative lens and recovery from lens-induced myopia in tree shrews (Tupaia glis belangeri). Vision Res. 2010;50:564–76. doi: 10.1016/j.visres.2009.12.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Benavente-Perez A, Nour A, Troilo D. The Effect of Simultaneous Negative and Positive Defocus on Eye Growth and Development of Refractive State in Marmosets. Invest Ophthalmol Vis Sci. 2012;53:6479–87. doi: 10.1167/iovs.12-9822. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Mao J, Liu S, Wen D, Tan X, Fu C. Basic fibroblast growth factor suppresses retinal neuronal apoptosis in form-deprivation myopia in chicks. Curr Eye Res. 2006;31:983–7. doi: 10.1080/02713680600910510. [DOI] [PubMed] [Google Scholar]
- 6.Tkatchenko AV, Luo X, Tkatchenko TV, Vaz C, Tanavde VM, Maurer-Stroh S, Zauscher S, Gonzalez P, Young TL. Large-Scale microRNA Expression Profiling Identifies Putative Retinal miRNA-mRNA Signaling Pathways Underlying Form-Deprivation Myopia in Mice. PLoS One. 2016;11:e162541. doi: 10.1371/journal.pone.0162541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Wallman J, Winawer J. Homeostasis of eye growth and the question of myopia. Neuron. 2004;43:447–68. doi: 10.1016/j.neuron.2004.08.008. [DOI] [PubMed] [Google Scholar]
- 8.Troilo D, Quinn N, Baker K. Accommodation and induced myopia in marmosets. Vision Res. 2007;47:1228–44. doi: 10.1016/j.visres.2007.01.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Schwahn HN, Schaeffel F. Flicker parameters are different for suppression of myopia and hyperopia. Vision Res. 1997;37:2661–73. doi: 10.1016/s0042-6989(97)00114-4. [DOI] [PubMed] [Google Scholar]
- 10.Cremieux J, Orban GA, Duysens J, Amblard B, Kennedy H. Experimental myopia in cats reared in stroboscopic illumination. Vision Res. 1989;29:1033–6. doi: 10.1016/0042-6989(89)90117-x. [DOI] [PubMed] [Google Scholar]
- 11.Crewther SG, Barutchu A, Murphy MJ, Crewther DP. Low frequency temporal modulation of light promotes a myopic shift in refractive compensation to all spectacle lenses. Exp Eye Res. 2006;83:322–8. doi: 10.1016/j.exer.2005.12.016. [DOI] [PubMed] [Google Scholar]
- 12.Di Y, Liu R, Chu RY, Zhou XT, Zhou XD. Myopia induced by flickering light in guinea pigs: a detailed assessment on susceptibility of different frequencies. Int J Ophthalmol. 2013;6:115–9. doi: 10.3980/j.issn.2222-3959.2013.02.01. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Mao J, Liu S, Qin W, Li F, Wu X, Tan Q. Levodopa inhibits the development of form-deprivation myopia in guinea pigs. Optom Vis Sci. 2010;87:53–60. doi: 10.1097/OPX.0b013e3181c12b3d. [DOI] [PubMed] [Google Scholar]
- 14.Zhao J, Qu X, Qi Y, Zhou W, Liu X. Study on retinal dopamine transporter in form deprivation myopia using the radiopharmaceutical tracer 99mTc-TRODAT-1. Nucl Med Commun. 2010;31:910–5. doi: 10.1097/MNM.0b013e32833decc0. [DOI] [PubMed] [Google Scholar]
- 15.Ohngemach S, Hagel G, Schaeffel F. Concentrations of biogenic amines in fundal layers in chickens with normal visual experience, deprivation, and after reserpine application. Vis Neurosci. 1997;14:493–505. doi: 10.1017/s0952523800012153. [DOI] [PubMed] [Google Scholar]
- 16.Megaw P, Morgan I, Boelen M. Vitreal dihydroxyphenylacetic acid (DOPAC) as an index of retinal dopamine release. J Neurochem. 2001;76:1636–44. doi: 10.1046/j.1471-4159.2001.00145.x. [DOI] [PubMed] [Google Scholar]
- 17.Guo SS, Sivak JG, Callender MG, Diehl-Jones B. Retinal dopamine and lens-induced refractive errors in chicks. Curr Eye Res. 1995;14:385–9. doi: 10.3109/02713689508999936. [DOI] [PubMed] [Google Scholar]
- 18.Bartmann M, Schaeffel F, Hagel G, Zrenner E. Constant light affects retinal dopamine levels and blocks deprivation myopia but not lens-induced refractive errors in chickens. Vis Neurosci. 1994;11:199–208. doi: 10.1017/s0952523800001565. [DOI] [PubMed] [Google Scholar]
- 19.Stone RA, Lin T, Laties AM, Iuvone PM. Retinal dopamine and form-deprivation myopia. Proc Natl Acad Sci USA. 1989;86:704–6. doi: 10.1073/pnas.86.2.704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Iuvone PM, Tigges M, Fernandes A, Tigges J. Dopamine synthesis and metabolism in rhesus monkey retina: development, aging, and the effects of monocular visual deprivation. Vis Neurosci. 1989;2:465–71. doi: 10.1017/s0952523800012360. [DOI] [PubMed] [Google Scholar]
- 21.Schmid KL, Wildsoet CF. Inhibitory effects of apomorphine and atropine and their combination on myopia in chicks. Optom Vis Sci. 2004;81:137–47. doi: 10.1097/00006324-200402000-00012. [DOI] [PubMed] [Google Scholar]
- 22.Gao Q, Liu Q, Ma P, Zhong X, Wu J, Ge J. Effects of direct intravitreal dopamine injections on the development of lid-suture induced myopia in rabbits. Graefes Arch Clin Exp Ophthalmol. 2006;244:1329–35. doi: 10.1007/s00417-006-0254-1. [DOI] [PubMed] [Google Scholar]
- 23.Feldkaemper M, Schaeffel F. An updated view on the role of dopamine in myopia. Exp Eye Res. 2013;114:106–19. doi: 10.1016/j.exer.2013.02.007. [DOI] [PubMed] [Google Scholar]
- 24.Hu G, Jin GZ. (-)-Stepholidine antagonizes the inhibition by D2 receptor agonists on synaptosomal tyrosine hydroylase in rat corpus striatum. Zhongguo Yao Li Xue Bao. 1995;16:376–9. [PubMed] [Google Scholar]
- 25.Xiao L, Chen R, Guo J. A study on the mechanism of apomorphine for treatment of myopia. Zhonghua Yan Ke Za Zhi. 1998;34:130–3. [PubMed] [Google Scholar]
- 26.Norton TT, McBrien NA. Normal development of refractive state and ocular component dimensions in the tree shrew (Tupaia belangeri). Vision Res. 1992;32:833–42. doi: 10.1016/0042-6989(92)90026-f. [DOI] [PubMed] [Google Scholar]
- 27.Wu X, Qian K, Xu G, Li Y, Ma Y, Huang F, Wang YQ, Zhou X, Qu J, Yang XL, Zhong YM, Weng SJ. The Role of Retinal Dopamine in C57BL/6 Mouse Refractive Development as Revealed by Intravitreal Administration of 6-Hydroxydopamine. Invest Ophthalmol Vis Sci. 2016;57:5393–404. doi: 10.1167/iovs.16-19543. [DOI] [PubMed] [Google Scholar]
- 28.Nicotra A, Parvez SH. Apoptotic molecules and MPTP-induced cell death. Neurotoxicol Teratol. 2002;24:599–605. doi: 10.1016/s0892-0362(02)00213-1. [DOI] [PubMed] [Google Scholar]
- 29.Blum D, Torch S, Lambeng N, Nissou M, Benabid A, Sadoul R, Verna JM. Molecular pathways involved in the neurotoxicity of 6-OHDA, dopamine and MPTP: contribution to the apoptotic theory in Parkinson’s disease. Prog Neurobiol. 2001;65:135–72. doi: 10.1016/s0301-0082(01)00003-x. [DOI] [PubMed] [Google Scholar]
- 30.Yu Y, Chen H, Tuo J, Zhu Y. Effects of flickering light on refraction and changes in eye axial length of C57BL/6 mice. Ophthalmic Res. 2011;46:80–7. doi: 10.1159/000323179. [DOI] [PubMed] [Google Scholar]
- 31.Di Y, Lu N, Li B, Liu R, Chu RY, Zhou XT, Zhou XD. Effects of chronic exposure to 0.5 Hz and 5 Hz flickering illumination on the eye growth of guinea pigs. Curr Eye Res. 2013;38:1182–90. doi: 10.3109/02713683.2013.807931. [DOI] [PubMed] [Google Scholar]
- 32.Howlett MHC, McFadden SA. Form-deprivation myopia in the guinea pig (Cavia porcellus). Vision Res. 2006;46:267–83. doi: 10.1016/j.visres.2005.06.036. [DOI] [PubMed] [Google Scholar]
- 33.Megaw PL, Morgan IG, Boelen MK. Dopaminergic behaviour in chicken retina and the effect of form deprivation. Aust N Z J Ophthalmol. 1997;25(Suppl 1):S76–8. doi: 10.1111/j.1442-9071.1997.tb01764.x. [DOI] [PubMed] [Google Scholar]
- 34.Umino O, Lee Y, Dowling JE. Effects of light stimuli on the release of dopamine from interplexiform cells in the white perch retina. Vis Neurosci. 1991;7:451–8. doi: 10.1017/s0952523800009743. [DOI] [PubMed] [Google Scholar]
- 35.Witkovsky P. Dopamine and retinal function. Doc Ophthalmol. 2004;108:17–40. doi: 10.1023/b:doop.0000019487.88486.0a. [DOI] [PubMed] [Google Scholar]
- 36.Doyle SE, Grace MS, McIvor W, Menaker M. Circadian rhythms of dopamine in mouse retina: the role of melatonin. Vis Neurosci. 2002;19:593–601. doi: 10.1017/s0952523802195058. [DOI] [PubMed] [Google Scholar]
- 37.Luft WA, Iuvone PM, Stell WK. Spatial, temporal, and intensive determinants of dopamine release in the chick retina. Vis Neurosci. 2004;21:627–35. doi: 10.1017/S0952523804214110. [DOI] [PubMed] [Google Scholar]
- 38.Pendrak K, Nguyen T, Lin T, Capehart C, Zhu X, Stone RA. Retinal dopamine in the recovery from experimental myopia. Curr Eye Res. 1997;16:152–7. doi: 10.1076/ceyr.16.2.152.5090. [DOI] [PubMed] [Google Scholar]
- 39.Hamasaki DI, Trattler WB, Hajek AS. Light ON depresses and light OFF enhances the release of dopamine from the cat’s retina. Neurosci Lett. 1986;68:112–6. doi: 10.1016/0304-3940(86)90239-9. [DOI] [PubMed] [Google Scholar]
- 40.Kramer SG. Dopamine: A retinal neurotransmitter. I. Retinal uptake, storage, and light-stimulated release of H3-dopamine in vivo. Invest Ophthalmol. 1971;10:438–52. [PubMed] [Google Scholar]
- 41.Newkirk GS, Hoon M, Wong RO, Detwiler PB. Inhibitory inputs tune the light response properties of dopaminergic amacrine cells in mouse retina. J Neurophysiol. 2013;110:536–52. doi: 10.1152/jn.00118.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Fioravante D, Regehr WG. Short-term forms of presynaptic plasticity. Curr Opin Neurobiol. 2011;21:269–74. doi: 10.1016/j.conb.2011.02.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Rose KA, Morgan IG, Ip J, Kifley A, Huynh S, Smith W, Mitchell P. Outdoor activity reduces the prevalence of myopia in children. Ophthalmology. 2008;115:1279–85. doi: 10.1016/j.ophtha.2007.12.019. [DOI] [PubMed] [Google Scholar]
- 44.Smith EL, Hung L, Huang J. Protective Effects of High Ambient Lighting on the Development of Form-Deprivation Myopia in Rhesus Monkeys. Invest Ophthalmol Vis Sci. 2012;53:421–8. doi: 10.1167/iovs.11-8652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Macdonald PA, Monchi O. Differential effects of dopaminergic therapies on dorsal and ventral striatum in Parkinson’s disease: implications for cognitive function. Parkinsons Dis. 2011;2011:572743–818. doi: 10.4061/2011/572743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Cools R, Barker RA, Sahakian BJ, Robbins TW. Enhanced or impaired cognitive function in Parkinson’s disease as a function of dopaminergic medication and task demands. Cereb Cortex. 2001;11:1136–43. doi: 10.1093/cercor/11.12.1136. [DOI] [PubMed] [Google Scholar]
- 47.McBrien NA, Cornell LM, Gentle A. Structural and ultrastructural changes to the sclera in a mammalian model of high myopia. Invest Ophthalmol Vis Sci. 2001;42:2179–87. [PubMed] [Google Scholar]
- 48.McBrien NA, Lawlor P, Gentle A. Scleral remodeling during the development of and recovery from axial myopia in the tree shrew. Invest Ophthalmol Vis Sci. 2000;41:3713–9. [PubMed] [Google Scholar]
- 49.Jobling AI, Wan R, Gentle A, Bui BV, McBrien NA. Retinal and choroidal TGF-beta in the tree shrew model of myopia: isoform expression, activation and effects on function. Exp Eye Res. 2009;88:458–66. doi: 10.1016/j.exer.2008.10.022. [DOI] [PubMed] [Google Scholar]
- 50.Seko Y, Tanaka Y, Tokoro T. Apomorphine inhibits the growth-stimulating effect of retinal pigment epithelium on scleral cells in vitro. Cell Biochem Funct. 1997;15:191–6. doi: 10.1002/(SICI)1099-0844(199709)15:3<191::AID-CBF738>3.0.CO;2-2. [DOI] [PubMed] [Google Scholar]
- 51.Seeman P, Van Tol HH. Dopamine receptor pharmacology. Trends Pharmacol Sci. 1994;15:264–70. doi: 10.1016/0165-6147(94)90323-9. [DOI] [PubMed] [Google Scholar]
- 52.Pescosolido N, Parisi F, Russo P, Buomprisco G, Nebbioso M. Role of Dopaminergic Receptors in Glaucomatous Disease Modulation. BioMed Res Int. 2013;2013:1–5. doi: 10.1155/2013/193048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.McCarthy CS, Megaw P, Devadas M, Morgan IG. Dopaminergic agents affect the ability of brief periods of normal vision to prevent form-deprivation myopia. Exp Eye Res. 2007;84:100–7. doi: 10.1016/j.exer.2006.09.018. [DOI] [PubMed] [Google Scholar]
- 54.Huang F, Yan T, Shi F, An J, Xie R, Zheng F, Li Y, Chen J, Qu J, Zhou X. Activation of dopamine D2 receptor is critical for the development of form-deprivation myopia in the C57BL/6 mouse. Invest Ophthalmol Vis Sci. 2014;55:5537–44. doi: 10.1167/iovs.13-13211. [DOI] [PubMed] [Google Scholar]
- 55.Nickla DL, Totonelly K, Dhillon B. Dopaminergic agonists that result in ocular growth inhibition also elicit transient increases in choroidal thickness in chicks. Exp Eye Res. 2010;91:715–20. doi: 10.1016/j.exer.2010.08.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Jiang L, Long K, Schaeffel F, Zhou X, Zheng Y, Ying H, Lu F, Stell WK, Qu J. Effects of dopaminergic agents on progression of naturally occurring myopia in albino guinea pigs (Cavia porcellus). Invest Ophthalmol Vis Sci. 2014;55:7508–19. doi: 10.1167/iovs.14-14294. [DOI] [PubMed] [Google Scholar]