

Abstract
Background
While intensive Plasmodium falciparum multidrug resistance surveillance continues in Cambodia, relatively little is known about Plasmodium vivax drug resistance in Cambodia or elsewhere. To investigate P. vivax anti-malarial susceptibility in Cambodia, 76 fresh P. vivax isolates collected from Oddar Meanchey (northern Cambodia) in 2013–2015 were assessed for ex vivo drug susceptibility using the microscopy-based schizont maturation test (SMT) and a Plasmodium pan-species lactate dehydrogenase (pLDH) ELISA. P. vivax multidrug resistance gene 1 (pvmdr1) mutations, and copy number were analysed in a subset of isolates.
Results
Ex vivo testing was interpretable in 80% of isolates using the pLDH-ELISA, but only 25% with the SMT. Plasmodium vivax drug susceptibility by pLDH-ELISA was directly compared with 58 P. falciparum isolates collected from the same locations in 2013–4, tested by histidine-rich protein-2 ELISA. Median pLDH-ELISA IC50 of P. vivax isolates was significantly lower for dihydroartemisinin (3.4 vs 6.3 nM), artesunate (3.2 vs 5.7 nM), and chloroquine (22.1 vs 103.8 nM) than P. falciparum but higher for mefloquine (92 vs 66 nM). There were not significant differences for lumefantrine or doxycycline. Both P. vivax and P. falciparum had comparable median piperaquine IC50 (106.5 vs 123.8 nM), but some P. falciparum isolates were able to grow in much higher concentrations above the normal standard range used, attaining up to 100-fold greater IC50s than P. vivax. A high percentage of P. vivax isolates had pvmdr1 Y976F (78%) and F1076L (83%) mutations but none had pvmdr1 amplification.
Conclusion
The findings of high P. vivax IC50 to mefloquine and piperaquine, but not chloroquine, suggest significant drug pressure from drugs used to treat multidrug resistant P. falciparum in Cambodia. Plasmodium vivax isolates are frequently exposed to mefloquine and piperaquine due to mixed infections and the long elimination half-life of these drugs. Difficulty distinguishing infection due to relapsing hypnozoites versus blood-stage recrudescence complicates clinical detection of P. vivax resistance, while well-validated molecular markers of chloroquine resistance remain elusive. The pLDH assay may be a useful adjunctive tool for monitoring for emerging drug resistance, though more thorough validation is needed. Given high grade clinical chloroquine resistance observed recently in neighbouring countries, low chloroquine IC50 values seen here should not be interpreted as susceptibility in the absence of clinical data. Incorporating pLDH monitoring with therapeutic efficacy studies for individuals with P. vivax will help to further validate this field-expedient method.
Electronic supplementary material
The online version of this article (doi:10.1186/s12936-017-2034-2) contains supplementary material, which is available to authorized users.
Keywords: Drug resistance, Plasmodium vivax, Cambodia, Ex vivo assay, pvmdr1
Background
Cambodia continues to be at the epicenter of globally emerging multidrug resistant malaria. This has prompted intensive elimination efforts accompanied by surveillance to characterize Plasmodium falciparum resistance. Over the past several decades, P. falciparum has developed resistance to numerous drugs, particularly in Southeast Asia where high grade treatment failures have been documented [1–4]. Since 2000, the use of artemisinin (ART)-based combination therapy (ACT) as first-line treatment has been implemented in nearly all malaria endemic areas to overcome resistance developing as a result of monotherapy treatments. However, P. falciparum resistance to ART emerged in 2006, just a few years after introduction of artesunate–mefloquine (AS–MQ) in Cambodia [5, 6], and was later confirmed on a large scale by intensive multidisciplinary surveillance studies [7]. In 2013, the first high grade clinical failures of dihydroartemisinin–piperaquine (DHA–PPQ) were reported [8]. DHA–PPQ had only recently been introduced as first-line therapy in Cambodia. Reduced in vitro P. falciparum susceptibility to PPQ developed on a background of artemisinin resistance here [9, 10], with specific molecular mechanisms elucidated soon thereafter [11, 12].
While intensive surveillance for P. falciparum continues in Cambodia, little is known about Plasmodium vivax drug resistance. Clinical P. vivax resistance is far more difficult to characterize due to difficulties distinguishing true recrudescence of resistant parasites, reinfections with new blood-stage P. vivax infections, and relapsing infections by liver stage hypnozoites. Moreover, inability to maintain long-term culture of P. vivax parasites prevents reproducible assessments of parasite drug susceptibilities. Plasmodium vivax resistance is generally assumed to be less pronounced than P. falciparum, as blood-stage P. vivax infection remains clinically susceptible to most available anti-malarials. CQ resistant P. vivax was first reported in 1989 from Papua New Guinea (PNG) and Papua Indonesia [13–16], where CQ monotherapy remains ineffective. Chloroquine-resistant vivax isolates in this region have been found to harbour polymorphisms in the pvmdr1 (P. vivax multidrug resistance 1) gene, whereas amplification of the gene has been associated with reduced susceptibility to MQ and other drugs in vitro [17–20]. Sporadic cases of CQ failure have since been reported in parts of Southeast Asia and South America [21–24], but there is no clear evidence that these are associated with pvmdr1 mutations in these regions. Thus, the usefulness of this putative drug resistance marker for detecting emerging resistance remains uncertain.
Artemisinin-based combination therapy is now recommended for use as first-line agent for all malaria in areas of multidrug resistant P. falciparum, particularly where clinical failures have been documented [25, 26]. In Cambodia, DHA–PPQ was used as first-line therapy for both vivax and falciparum malaria since 2011 to respond to declining efficacy of CQ for P. vivax treatment in specific northern and western provinces where the clinical cure rate had dropped to 80–90%, though cure rates have remained near 100% elsewhere [27–29]. The approach was also implemented to simplify drug administration and overcome diagnostic difficulties. Limited diagnostic capacity makes distinguishing P. vivax from P. falciparum microscopically challenging, despite overall increases in diagnostic capacity after years of effort. Frequent relapse of latent P. vivax following treatment for blood stage P. falciparum infections, and high prevalence of difficult to distinguish mixed species infections pose further challenges. While the use of DHA–PPQ to treat CQ-sensitive P. vivax is thought to be effective and convenient [26], it may in fact exacerbate resistance to both P. vivax and P. falciparum [30].
To investigate P. vivax anti-malarial susceptibility in Cambodia, fresh P. vivax isolates collected from Northern provinces from 2013 to 2015 were assessed for sensitivity to commonly used drugs in short-term culture. Growth inhibition was measured by both the microscopy-based schizont maturation test (SMT) and Plasmodium pan-species lactate dehydrogenase (pLDH) ELISA. SMT had been considered the conventional method for P. vivax drug testing, but not sensitive for low parasitaemia samples. It is also labour-intensive and interpretation of results is subjective. pLDH ELISA has emerged, offering notable advantages. pLDH has demonstrated higher sensitivity for detection in settings of low P. vivax growth rate and very low parasitaemia, improving yields for IC50 determination [31]. Results from these assays were then compared with those from concurrently collected P. falciparum isolates analysed in both the novel pLDH assay and the previously established HRP-2 assay [32]. Plasmodium vivax multidrug resistance gene 1 (pvmdr1) mutations and copy number, which have been proposed as candidate markers for drug resistance, were analyzed in a subset of isolates. This is the first report that we are aware of that documents ex vivo P. vivax drug susceptibility in Cambodia, and provides baseline data for future surveillance and elimination efforts.
Methods
Study site, sample collection and processing
Plasmodium isolates were obtained from volunteers with uncomplicated malaria enrolled in an anti-malarial drug resistance surveillance study conducted June 2013–October 2015, in Oddar Meanchey province, northern Cambodia. The study was approved by the Cambodian National Ethics Committee for Health Research (NECHR), and the Walter Reed Army Institute of Research (WRAIR) Institutional Review Board (protocol WR1576). All subjects were ≥ 13 years old without a history of anti-malarial drug use within the past 7 days. Diagnosis of malaria was performed using Giemsa-stained peripheral blood smears, and confirmed later by real-time PCR [9]. A total of 76 P. vivax isolates were collected from volunteers with mono P. vivax from June 2013 to October 2015, while 58 P. falciparum isolates were collected for comparison from those with mono P. falciparum infections from January to August 2014. After informed consent, patient venous blood samples were collected in sodium heparin and directly tested fresh for ex vivo drug susceptibility within 6 h of phlebotomy without blood centrifugation, leukocyte depletion or culture adaptation. Additional blood was collected in an EDTA tube for molecular marker analysis for P. vivax drug resistance.
In the first 29 P. vivax isolates, a portion of heparinized blood was centrifuged, and cell pellets including buffy coat were passed through CF11 cellulose (Whatman™, Maidstone, UK) or Plasmodipur filters (EuroProxima, The Netherlands) to remove human white blood cells (WBC). WBC depleted samples were tested for drug susceptibility side by side with whole blood samples. Samples were treated with 45% Percoll in only the first 3 P. vivax isolates to separate late stage parasites and enrich for and synchronize at early stages for the schizont maturation test (SMT) [33]. An overall schematic for the experiment is shown in Fig. 1.
Fig. 1.
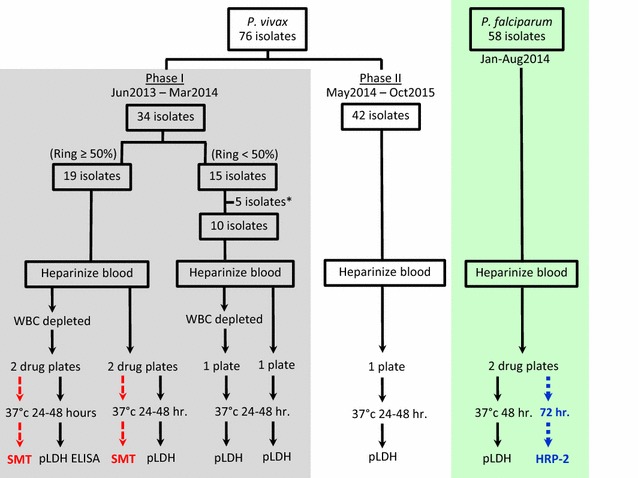
Schematic for ex vivo experiments. Plasmodium vivax ex vivo experiments were divided into 2 phases. In Phase I, the schizont maturation test (SMT) was performed concurrently with the pLDH ELISA. Blood samples were processed to deplete white blood cells, and incubated with drugs of interest in parallel with whole blood samples. The SMT was conducted only in samples with ≥ 50% ring stages. *5 isolates with ring stage < 50% in initial samples were not tested for drug susceptibility, but 3 of them had Percoll treatment applied to separate late stage parasites and enrich for early stages for the SMT. In phase II, only pLDH ELISA was performed on whole blood samples. For P. falciparum testing, pLDH ELISA was performed concurrently with the HRP-2 ELISA with 48 h incubation times used in the pLDH ELISA to ensure meaningful comparison of P. vivax susceptibility, while the previously established 72 h incubation was followed for the HRP-2 method
Preparation of dried drug plates
Dried drug-coated plates were prepared using published methods, and tested against the P. falciparum W2 reference clone for quality control [32, 34]. All drugs were provided by Chemical Repository of the Walter Reed Army Institute of Research (WRAIR). Briefly, dihydroartemisinin (DHA), artesunate (AS), mefloquine hydrochloride (MQ), quinine sulfate hydrate (QN), chloroquine diphosphate (CQ), and piperaquine phosphate (PPQ) were coated onto 96 well plates. Atovaquone (ATQ), lumefantrine (LUM), doxycycline (DOX) and artemisone (ATM) were added only to later plate lots once the initial methods had been established, and as a result were tested in only 76% (ATQ and LUM), 60% (DOX), and 49% (ATM) of isolates, respectively. Final drug concentrations (after adding samples) ranged from 0.095 to 70 nM for DHA, 0.07 to 52 nM for AS, 0.66 to 482 ng/ml for MQ, 2.18 to 1596 nM for QN, 5.31 to 3877 nM for CQ, 0.9 to 674 nM for PPQ, 0.38 to 273 nM for ATQ, 0.13 to 95 nM for LUM, 285 to 207,943 nM for DOX, and 0.02 to 25 ng/ml for ATM. Starting in 2013, some P. falciparum isolates were able to grow in extremely high PPQ concentrations [35]. From 2014 onward, increased PPQ concentration (3.4–53,905 nM) was used in addition to the standard dilutions to ensure accurate inhibitory concentrations could be determined. The top row of each plate served as a drug-free control.
Immediate ex vivo drug susceptibility assay
At the time of diagnosis (before treatment), fresh Plasmodium isolates were tested for sensitivity to anti-malarials by culturing on dried drugs coated plates and measuring growth inhibition. For the P. vivax assay, blood samples were adjusted to 2% haematocrit in McCoy’s 5A medium supplemented with 20% AB blood-group serum, and added to dried drug coated plates. Plates were then incubated at 37 °C in a candle jar for 24–48 h, and parasite maturation in drug-free wells was checked by microscopy at 24 h and then every 6 h until 48 h. Parasite growth inhibition over the established concentration ranges was measured by the microscopy-based schizont maturation test (SMT) using the modified WHO microtest [36, 37] for the first 19 P. vivax isolates with > 50% early stage parasites in initial samples and Plasmodium pan-species lactate dehydrogenase (pLDH) ELISA [31, 38] for all isolates, side by side (Fig. 1). For the SMT, incubation was stopped when ≥ 40% of parasites in drug-free wells had reached the schizont stage. Thick smear examination of samples from each well was performed, and the number of schizonts per 200 asexual stage parasites for each drug concentration was determined. The number of schizonts were plotted against drug concentrations, and IC50s were estimated by nonlinear regression analysis using GraphPad Prism version 6.0. For the pLDH-ELISA assay, incubation was stopped when ≥ 40% schizont stages were reached in drug-free wells, or at 48 h if parasite cultures did not reach at least 40% schizonts. Plates were frozen and later thawed for analysis of growth inhibition using the pLDH-ELISA. pLDH optical density (OD) readings were plotted against drug concentrations, and IC50s were estimated as described above. Samples with poor growth rates, defined as < 40% schizonts in drug-free wells or a pLDH-OD ratio < 1.7 between no-drug control wells and maximum tested drug concentrations, were excluded from data analysis for SMT or pLDH-ELISA, respectively. A “successful” IC50 assay result was defined as achieving a sigmoidal concentration–response when testing serial drug dilutions for at least one of the tested drugs.
For the P. falciparum assay, the pLDH-ELISA [38] and the established histidine-rich protein-2 (HRP-2) ELISA [32] was used to simultaneously test field isolates and reference laboratory clones (W2 and 3D7). Culture conditions used in the P. falciparum assay were adjusted to 1.5% haematocrit in 0.5% Albumax RPMI 1640. Parasite growth inhibition was assessed by pLDH-ELISA and HRP-2 ELISA after 48 and 72 h incubation, respectively. The same incubation period (48 h) for P. vivax and P. falciparum were used in the pLDH ELISA to ensure meaningful comparison of susceptibility results. The previously established 72 h incubation was followed for the HRP-2 method. IC50s were estimated as described above. Isolates with reduced susceptibility to PPQ capable of growing in maximum drug concentrations could not be interpolated using the standard PPQ dilution range (0.9–674 nM). To accurately estimate IC50 dose–response, the curve was replotted by fitting ‘zero-growth’ HRP2 or pLDH OD values at the extrapolated PPQ concentration of 53,905 nM [35, 39].
Plasmodium vivax multi-drug resistance (pvmdr1) SNPs and copy number analysis
Genomic DNA was prepared from EDTA-anticoagulated blood using the QIAamp DNA Mini Kit (Qiagen) and used to analyse pvmdr1 Y976F and F1076L mutations and copy number variation. Multiplex real-time PCR was used to assess pvmdr1 copy number. The pvmdr1 (target) and pv aldolase (reference) genes were both amplified by RT-PCR, following methods described by Lin et al. [40]. Plasmids containing cloned fragments of pvmdr1 and pv aldolase, developed using the TOPO XL PCR Cloning Kit (Invitrogen, Carlsbad, CA), were used as positive controls in each experiment. Reactions were performed in duplicate. Threshold cycle (Ct) values were used to calculate the relative quantitation of pvmdr1 copy number by the Pfaffl method [41]. Copy number was determined by rounding to the nearest integer and was considered increased if > 1. For pvmdr1 Y976F and F1076L analysis, PCR was performed to amplify a 647 bp region of the pvmdr1 gene covering these SNPs, using the conditions previously described in Lin et al. and amplified products were directly sequenced [40].
Statistical analysis
Statistical analysis was performed using Graph-Pad Prism version 6.0 (GraphPad Software, Inc, San Diego, CA, USA). Parasite drug susceptibilities were expressed as median IC50s for all isolates. Differences in susceptibility between groups were determined using non-parametric Mann–Whitney or Kruskal–Wallis tests. Comparison of IC50s attained from pLDH and HRP-2 ELISA were made using Wilcoxon matched pair testing. Assay correlation was evaluated using Spearman’s correlation test. Spearman test was also used to examine the correlation between IC50 and ratio of early/mature stage parasites (EM ratio) at start of assay to determine the effect of initial P. vivax stage on LDH-ELISA IC50 results.
Results
Ex vivo P. vivax assay development using the schizont maturation test
An overall schematic for the experiment is shown in Fig. 1. Of 76 P. vivax isolates collected from Oddar Meanchey Province between June 2013 and October 2015, 71 fresh isolates were tested for ex vivo sensitivity to anti-malarials. Parasite density ranged from 2500 to 38,000 µL−1 with variation in the initial parasite stage. Synchronous (≥ 80%) early and mature trophozoite stages were found in 13 and 19 samples, respectively, while the rest included mixed stages. In an effort to improve suitability for the schizont maturation test (SMT) by synchronizing isolates at the early stage, the first 3 isolates were treated with 45% Percoll to separate late stage parasites and enrich for early stage. Although Percoll treatment helped to increase a proportion of early stages in samples, trophozoites remained, leading to failure to reach 80% early stage for all 3 samples. One explanation is that the density of 45% Percoll normally can separate the schizonts and very late trophozoites, leaving earlier trophozoites and ring stages in the samples. In addition, perfect separation efficiency of Percoll density centrifugation method may not be achievable in practice as even if synchronized late stages are found in interphase layer, mixed stage parasites may still be found in the pellet. The effect of white blood cell (WBC) depletion processes on parasite maturation were compared for the first 29 isolates, but far fewer schizonts in drug-free wells were observed in WBC-depleted samples (0–29% schizonts), compared to whole blood samples (0–72% schizont). None of the WBC depleted samples reached the threshold for successful culture for SMT (≥ 40% schizonts in drug-free wells) [37]. Given that neither Percoll treatment nor WBC depletion improved assay conditions, both were abandoned. To examine the effects of WBC on the assay, the pLDH assay was run on whole blood and packed RBC specimens of these 29 P. vivax isolates, side by side. Good correlations were found for all drugs tested (ρ = 0.5–0.9, P < 0.05), though slightly reduced IC50s were found after removing WBC for DHA, AS, and CQ with median difference of = − 0.6, − 0.8, − 6.2, respectively. No significant differences were found for other drugs. Reasonable correlation of results attained from both approaches suggested assay reliability when using whole blood samples.
In ex vivo assays of whole blood samples, erythrocytic P. vivax maturation was observed for all but 5 isolates in no-drug control wells over 48 h culture resulting in 2–72% schizonts (median = 21%). However, only 22 of 71 isolates were able to develop to ≥ 40% schizonts. The time to maximum ex vivo schizont growth varied from 24 to 48 h, with a median of 48 h, though it tended to be earlier for cultures with mature trophozoite stages taking only 24–30 h. There was a positive correlation between higher ratios of early/mature trophozoites (EM ratio) and time taken to reach maximum schizont growth (Spearman ρ = 0.63; P < 0.001). Of the first 19 P. vivax isolates with > 50% early stage parasites in initial samples, SMT yielded an interpretable IC50 in only 6 isolates (32% success rate). Since the SMT was found to have fewer interpretable results and was significantly more labour-intensive than pLDH-ELISA, the SMT was not performed in the remaining experiments.
Comparability of ex vivo Plasmodium falciparum susceptibility in the HRP-2 and pLDH ELISA assays
Given the challenges of the SMT, the pLDH ELISA was evaluated next. In order to first establish the relevance of the pLDH ELISA assay for comparing P. falciparum to P. vivax resistance profiles, P. falciparum IC50 results in our routinely used HRP-2 assay and the pLDH ELISA were compared. Figure 2 revealed similarities between P. falciparum isolate IC50 in the HRP-2 and pLDH ELISA assays for chloroquine (CQ), lumefantrine (LUM), the artemisinins (ARTs), piperaquine (PPQ), and doxycycline (DOX). Wilcoxon pairwise IC50 comparisons were significantly higher in the pLDH assay for mefloquine (MQ). Figure 3 illustrates global correlation for P. falciparum isolates susceptibility between the 2 assays. There were moderate to strong correlations between results obtained for P. falciparum isolates in the pLDH assay and HRP-2 ELISAs for all drugs tested, with the exception of DOX. The latter findings were despite similarities in median values overall.
Fig. 2.
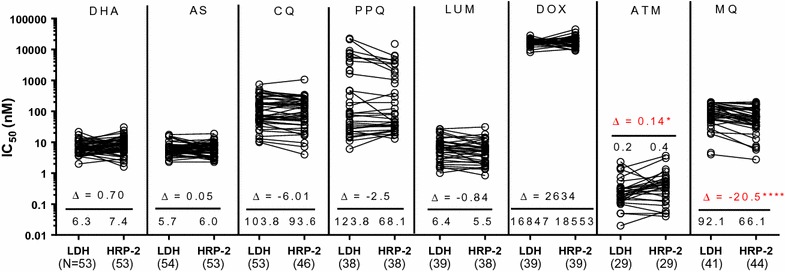
Comparison of P. falciparum IC50 values attained from pLDH and HRP-2 ELISA. Fresh P. falciparum isolates were tested for drug susceptibility using pLDH and HRP-2 ELISA, side by side. Median differences (Δ) are indicated below each pairwise comparison with values from the respective assays for each isolate joined by black lines. Unconnected dots are those where a corresponding value could not be obtained in either the pLDH or HRP-2 ELISA. Median IC50 and numbers of evaluable isolates from the respective assays appear above the X-axis, Significant P-values from the Wilcoxon pair test are indicated as *(P < 0.05), **(P < 0.01), ***(P < 0.001), ****(P < 0.0001)
Fig. 3.
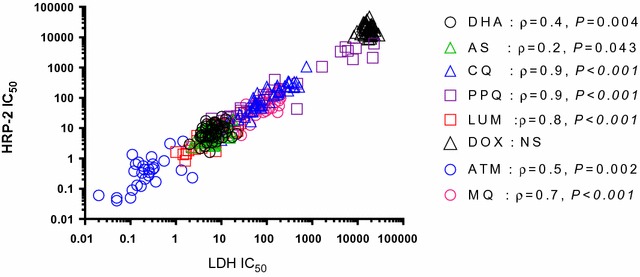
Correlation between P. falciparum IC50 values attained from pLDH and HRP-2 ELISA. There were moderate to strong correlations between results obtained for P. falciparum isolates in the pLDH and HRP-2 ELISAs for all drug tested, with the exception of doxycyline. Significant P-values and correlation coefficients (ρ) from Spearman correlation test were indicated. NS indicates not significant based on a P-value ≥ 0.05
Comparison of ex vivo P. vivax and P. falciparum drug susceptibility in Cambodia 2013–2015 using the pLDH-ELISA method
pLDH-ELISA IC50s to anti-malarials of Cambodian P. vivax isolates collected during 2013–2015 are shown in Fig. 4. Limited SMT data as described above hindered meaningful comparison of results from these 2 assays. pLDH-ELISA IC50 results from P. falciparum isolates collected in the same area, and from laboratory clones are also presented in Fig. 4 for comparison. IC50 results of P. falciparum laboratory clones were able to successfully discriminate expected susceptibility profiles for the 3D7 (CQ-sensitive, MQ-resistant) and W2 strains (CQ-resistant, MQ-sensitive). pLDH-ELISA IC50s were significantly lower for P. vivax than P. falciparum clinical isolates for dihydroartemisinin (DHA), artesunate (AS), and CQ, while no differences were found for LUM where both had low IC50 or DOX where both had high IC50 (Fig. 4). In contrast, pLDH-ELISA IC50s of P. vivax isolates were significantly higher for artemisone (ATM) and MQ than P. falciparum in this assay.
Fig. 4.
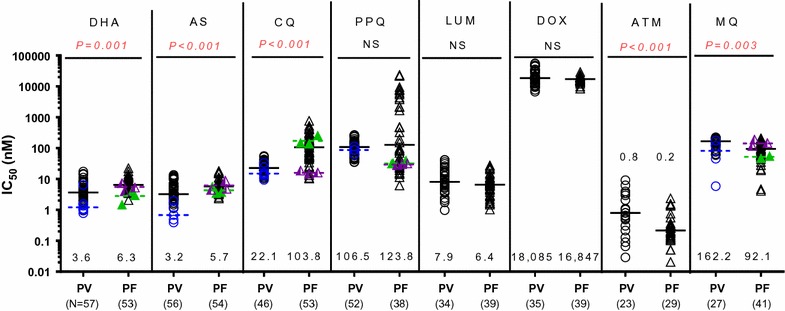
Ex vivo drug susceptibility of P. vivax and P. falciparum isolates collected from Oddar Meanchey Province (northern Cambodia) during 2013–2015. pLDH-ELISA IC50s of fresh P. vivax (PV) and P. falciparum (PF) isolates against commonly used anti-malarials are presented as black dot plots with black median bars, while SMT results for the only six evaluable PV isolates are indicated in blue. Circle represent PV isolates, while triangles represent PF. Median pLDH-ELISA IC50 and numbers of evaluable isolates appear above the X-axis, and the P-values for the Mann–Whitney test comparing IC50s of P. vivax and P. falciparum appear at the top of the graph. NS indicates not significant based on a P-value ≥ 0.05. pLDH-ELISA IC50 of W2 (green) and 3D7 (purple) reference clones obtained from three independent assays are presented in scatter dot plot with bars representing averages
Both P. vivax and P. falciparum had comparable median PPQ IC50 (106.5 vs 123.8 nM respectively), but some P. falciparum isolates were able to grow in much higher concentrations above the normal standard curve used (Fig. 4). Although a statistically significant difference was not detected initially when comparing PPQ susceptibility between P. vivax and P. falciparum based on the pLDH assay, 67% of P. falciparum isolates (39 out of 58 total) were found to survive exposure to maximum standard curve PPQ levels (674 nM). Some isolates were found to have few 100-fold greater IC50s (470–22,000 nM) using a new testing paradigm with increased maximal PPQ concentrations of 53,905 nM, compared to those obtained in previous years (< 50 nM). In contrast, none of the evaluated P. vivax isolates were able to grow in the highest tested PPQ concentration of 674 nM, suggesting greater overall susceptibility compared to P. falciparum. P. vivax susceptibility to PPQ increased over the period 2013–2015, while there were minor fluctuations or no change in parasite susceptibility to the other drugs tested (Additional file 1).
In contrast to the SMT, pLDH-ELISA was able to determine IC50 in 83% of P. vivax isolates (58/70) evaluated, though success rate was drug dependent, ranging from 66 to 83% for most drugs. There was < 20% success in achieving sigmoidal IC50 curves for quinine (QN) and atovaquone (ATQ). High background optical density (OD) levels in maximally concentrated wells interfered with interpretation. An OD ratio < 1.7 between drug-free control and maximum concentration wells precluded interpretable concentration–response curves. Therefore, QN and ATQ IC50 results were excluded from the analysis. Likewise MQ had a lower success rate (39%) compared to other drugs. The high pLDH-OD background interference observed during QN, ATQ, and MQ testing was also observed in P. falciparum assays but had less effect on interpretability, and was not detected when P. falciparum was tested in the HRP-2 ELISA. The success rate of pLDH-ELISA for P. falciparum isolates ranged from 62 to 98% for all drugs tested.
Effect of initial P. vivax stage, assay incubation time and ex vivo growth efficiency on pLDH-ELISA IC50 results
To determine the effect of initial P. vivax stage on pLDH-ELISA results, the relationship between initial parasite stage and IC50 was examined (Table 1). There was a negative correlation between early/mature trophozoite (EM) ratio and IC50 for DOX (ρ = − 0.7) with lesser effects observed for DHA, AS and LUM (ρ = − 0.3 to − 0.5). Three to four-fold greater IC50 for these drugs were detected in P. vivax isolates with synchronous mature trophozoites initially (EM ratio ≤ 0.25) than those with synchronous early stages (EM ratio ≥ 4). However, there was only a borderline correlation between EM ratio and IC50 for MQ, and no effect for other drugs.
Table 1.
Effect of initial P. vivax stage on pLDH-ELISA IC50 results
| Drug | Spearman correlation test | Mann–Whitney U test | P value | |||||
|---|---|---|---|---|---|---|---|---|
| for IC50 and EM ratio | EM ratio ≥ 4 | EM ratio ≤ 0.25 | ||||||
| N | coefficient | P-value | N | median IC50 | N | median IC50 | ||
| DHA | 57 | − 0.34* | 0.009* | 11 | 2.6 | 12 | 7.5* | 0.008* |
| AS | 56 | − 0.46* | < 0.001* | 11 | 2.9 | 11 | 9.4* | < 0.001* |
| CQ | 46 | − 0.12 | 0.445 | 12 | 23.8 | 4 | 28.1 | 0.716 |
| PPQ | 52 | − 0.01 | 0.922 | 12 | 104.3 | 7 | 99.5 | 0.612 |
| LUM | 34 | − 0.42* | 0.014* | 4 | 5.6 | 7 | 25.4* | 0.023* |
| DOX | 35 | − 0.70* | < 0.001* | 5 | 12,680 | 9 | 42,667* | 0.003* |
| ATM | 23 | − 0.12 | 0.593 | 3 | 0.61 | 7 | 2.0 | 0.137 |
| MQ | 27 | − 0.39 | 0.046* | 4 | 161.2 | 4 | 203.7 | 0.083 |
* Statistical significance
The influence of assay incubation time on pLDH-ELISA IC50 results was examined, and there were noticeable effects when testing DHA, AS, LUM, and DOX. There were significant increases in IC50 for these drugs in isolates requiring shorter incubations (24–30 h) compared to longer ones (42–48 h) (Kruskall–Wallis P value < 0.001–0.016). To assess the effect of ex vivo growth efficiency on assay results, pLDH-ELISA IC50 of isolates in the drug-free control assay reaching ≥ 40% schizonts (the designated threshold for SMT culture success) was compared with results of isolates having less growth efficiency. There were no differences in IC50 between the 2 groups for any of the drugs tested (Table 2).
Table 2.
There was little effect of ex vivo P. vivax growth on pLDH-ELISA IC50 results
| Drug | Maximum number of schizonts | P value of Mann–Whitney U test | |||
|---|---|---|---|---|---|
| < 40% schizont | ≥ 40% schizont | ||||
| N | median IC50 | N | median IC50 | ||
| DHA | 37 | 3.0 | 20 | 4.1 | 0.108 |
| AS | 37 | 3.0 | 19 | 3.5 | 0.166 |
| CQ | 32 | 22.1 | 14 | 22.1 | 0.981 |
| PPQ | 35 | 108.0 | 17 | 105.0 | 0.513 |
| LUM | 23 | 7.3 | 11 | 9.3 | 0.118 |
| DOX | 24 | 16,021 | 11 | 31,326 | 0.102 |
| ATM | 15 | 0.6 | 8 | 1.0 | 0.175 |
| MQ | 19 | 150.0 | 8 | 179.1 | 0.426 |
Plasmodium vivax drug resistance markers
Plasmodium vivax multi-drug resistance (pvmdr1) Y976F and F1076L mutations and copy number were analysed for 23 P. vivax isolates collected in 2013. Of 23 evaluated isolates, pvmdr1 Y976F and F1076L mutations were found in 78% (17/23) and 83% (19/23), respectively, composing 74% (17/23) double mutants, 9% (2/23) single F1076L mutants, and 17% (4/23) wild type isolates. None had pvmdr1 amplification.
Discussion
This is the first report of ex vivo P. vivax drug susceptibility testing of field isolates in Cambodia, providing important baseline data for ongoing resistance surveillance. The focus of containment and elimination efforts in Cambodia to date has been multidrug resistant P. falciparum malaria. Relatively little is known about P. vivax drug sensitivity due to the inability to culture P. vivax long term. The pLDH method bypasses this critical limitation by using fresh isolates, making it potentially practical and informative for measuring ex vivo P. vivax resistance. While further validation of the assay is needed, the correlations observed with the IC50 results with P. falciparum, in both the pLDH and previously established HRP-2 assays, lend support to its utility as a surveillance tool [32]. The P. falciparum HRP-2 assay has been carefully standardized in an effort to reliably produce interpretable IC50 results in P. falciparum field isolates over time [32]. However, it cannot be used in P. vivax which does not produce HRP-2. Unfortunately, we were unable to establish the P. vivax SMT using previously described methods. Potential explanations for the poor success rate of SMT could include low parasitaemia and drug residue in samples. Nearly 40% of P. vivax samples has parasitaemia < 0.1%, and issues of self-medication and unregulated anti-malarial distribution in Cambodia are well documented [42]. Further, WBC depletion process seems to retard P. vivax growth leading to failure for SMT. The loss of P. vivax and time spent during the WBC filtration process could be possible reasons. However, less effect was observed in pLDH ELISA in which IC50 from whole blood, and WBC depleted samples testing were well correlated. This corresponded with the previous finding on P. falciparum HRP-2 method, suggesting the reliability of assay on whole blood sample without WBC removing [43]. Overall, in addition to being a field expedient method, the pLDH method was able to reveal some important information about P. vivax susceptibility.
With the possible exception of chloroquine, blood stage P. vivax is generally thought to have remained susceptible to a wide variety of anti-malarials, though data is limited. Using the pLDH-ELISA, P. vivax appeared significantly more susceptible to dihydroartemisinin (DHA), artesunate (AS), and chloroquine (CQ) than P. falciparum,but less susceptible to mefloquine (MQ), and artemisone (ATM), and similarly susceptible to lumefantrine (LUM), piperaquine (PPQ) and doxycycline (DOX). It should be noted that although DOX IC50 values were in the micromolar range, they were still below previously proposed values for resistance [44]. The present study reveals Cambodian P. vivax isolates appear to remain sensitive to CQ while resistance to other anti-malarials may be worse than previously assumed, though the absence of baseline values precludes definitive conclusions. Although previously established, well-controlled pLDH methods were used here to test both P. vivax and P. falciparum, inter-species difference and assay bias between 2 species cannot be ruled out. Inter-species comparisons require careful interpretation, especially in the absence of baseline data. In addition, mixed parasite stages found in P. vivax samples may confound pLDH results for drugs with stage-specific activity. It is possible that the pLDH results of P. vivax here represent an average overall susceptibility of mixed parasite stages. As an example, our findings indicated that mixed stage P. vivax remained susceptible to CQ, though CQ has specific ring stage activity [37].
The present study brings to light important methodologic considerations for assessing P. vivax resistance in vitro. The effect of parasite growth efficiency on P. vivax pLDH assay was minimal. There were not significant differences in pLDH-ELISA IC50 for most drugs tested between isolates reaching ≥ 40% schizonts and those with less growth. Thus, the pLDH assay had utility even in P. vivax isolates failing to reach the 40% schizont target required for the SMT. pLDH-ELISA validity was also confirmed by successful discrimination of known susceptibility profiles for 3D7 (CQ sensitive, MQ-resistant) and W2 (CQ-resistant, MQ-sensitive) P. falciparum laboratory strains. When testing P. falciparum clinical isolates, pLDH-ELISA was able to detect PPQ resistance at several 100-fold higher IC50, corresponding with results from the 72 h HRP-2 ELISA. Comparison with these previously well benchmarked assays further supports use of pLDH-ELISA for P. vivax isolate drug susceptibility testing.
Based on our pLDH-ELISA results, stage-specific drug activity on P. vivax growth was apparent for DOX and AS, but less pronounced for DHA, LUM and MQ (Table 1). Isolates initially at the trophozoite stage had significantly higher IC50s to these drugs than those initially at the ring stage. Specific activity of chloroquine on ring stages, previously described for the schizont maturation test (SMT), was not detected here using the pLDH-ELISA. Comparative P. vivax testing at ring and trophozoite stages for the same isolates may confirm stage-specific activity of these drugs. Duration of drug incubation in the SMT is another factor previously reported to influence in vitro drug responses for P. vivax [37]. A prior statistical modelling study of SMT dose–response data indicated that only assays with initial ring stage parasitaemia ≥ 65% and a duration ≥ 35 h produce robust IC50 values [45]. More data is required to identify the threshold where the association between IC50 assay duration and parasite stage composition disappears in the pLDH ELISA.
Comparing findings of the present study to those reported previously for P. falciparum and P. vivax isolates from the Brazilian Amazon [46, 47] and Indonesia [36, 48, 49] using pLDH ELISA and SMT assays, Cambodian isolates were found to be less susceptible to MQ and PPQ. However, Cambodian and Brazillian P. vivax isolates were more sensitive to CQ than in Papua Indonesia where P. vivax CQ resistance has emerged. Another ex vivo study reported P. vivax CQ resistance in 60% of isolates collected from the Thai-Myanmar border, and higher median IC50 than in Cambodia [50]. Although, comparability of the SMT and pLDH ELISA have yet to be formally established, these regional differences are not surprising. Reduced MQ and PPQ susceptibility of Cambodian isolates reflects higher drug pressure in the region from long term use. High grade PPQ resistance recently emerged in P. falciparum with resultant effects on sympatric P. vivax infection [10, 30]. Chloroquine resistant P. vivax infections have been detected in Indonesia since the 1990s and, in 2008, the national treatment guidelines for P. vivax were changed to ACT [51]. Approximately 60% of P. vivax patients treated with chloroquine experienced a recurrence within 28 days in studies from Malaysia and Vietnam [24, 52]. Yet chloroquine sensitivity was better preserved in studies conducted in Cambodia [29, 44], the Brazilian Amazon [53], Myanmar [54], India [55] and Ethiopia [56].
Piperaquine phosphate IC50s of Cambodian P. vivax isolates were higher than Indonesia, but the median was similar to those of P. falciparum from the same region. However, some Cambodian P. falciparum isolates were able to grow in much higher PPQ concentrations with up to 100-fold greater IC50s than P. vivax, suggesting greater overall susceptibility in P. vivax compared to P. falciparum. In 2012, AS–MQ was replaced with DHA-PPQ as national first-line treatment for both P. falciparum and P. vivax [57]. Since reduced ex vivo P. falciparum PPQ sensitivity of Cambodian isolates has corresponded with high grade failure of DHA–PPQ treatment in this region [10, 35], it raises a concern that DHA–PPQ may also become less effective for P. vivax. Unlike P. falciparum, P. vivax does not exhibit concomitant artemisinin resistance. In the absence of an established baseline value, it is possible that P. vivax has higher intrinsic PPQ IC50 in the assay than P. falciparum. Further, the decline observed in P. vivax IC50 after PPQ was introduced in 2013 may have been the result of a decline in overall parasite fitness in response to developing mutations, similar to previous observations in P. falciparum [58]. Regardless, clinical correlations are necessary to define the impact of increased PPQ IC50s for P. vivax. Also to be explored is whether the change in treatment policy for P. vivax may have relieved chloroquine pressure. However, ongoing P. falciparum chloroquine resistance comparable to previously observed levels [9, 34] argues against this. Resistance in both parasite species would have been expected to subside if exposure had truly been reduced [59]. Definitive demonstration of assay utility will require comparison of ex vivo pLDH-ELISA results with clinical response in well-controlled P. vivax field studies. Limited ability to distinguish recrudescence, relapse, and reinfection clinically may confound interpretation.
While molecular data in the present study was limited by convenience sampling, and available resources, it does offer a few useful observations. The mechanism of CQ resistance in P. vivax remains unclear although a few studies have suggested associations with P. vivax multi-drug resistance gene (pvmdr1) Y976F mutation and pvcrt-o expression [17, 60]. In northern Cambodian isolates, pvmdr1 Y976F mutants were observed at extremely high frequency yet when tested in the ex vivo assay, the isolates are CQ-sensitive. Accordingly, CQ has retained clinical efficacy as a P. vivax rescue agent in trials conducted there over the past several years by the USAMD-AFRIMS [29]. This argues against the usefulness of the Y976F mutation as a CQ resistance marker [17]. No pvmdr1 amplification was detected in Cambodian isolates, precluding correlation with ex vivo drug susceptibility to chloroquine or other anti-malarials. No attempt to measure stage-specific pvcrt-o expression in these clinical isolates was made here. Comparing our findings to those reported previously from other regions revealed geographical differences in pvmdr1 amplification and mutation prevalence. The prevalence of Y976F and F1076L mutations were high in Cambodia, Indonesia, and Papua New Guinea (70–100%) [36, 61, 62], but no mutant was detected in Brazil [47, 60]. Moderate rates of mutation were previously found in Thailand (18–23% for Y976F and 53–61% for F1076L) [50, 62]. Corresponding to previous findings [40], most Cambodian isolates were double mutants (74%), with single F1076L mutants found in < 10%. The prevalence here differed compared to Thailand where a single F1076L mutation was seen in > 60%. No pvmdr1 amplification was detected in northern Cambodian isolates tested in the present study, despite reduced susceptibility to MQ. It is possible that if more sensitive assays targeting the pvmdr1 breakpoint were used [63], minority clones with pvmdr1 amplification may have been detected, given the polyclonal nature of vivax infections in this region [64–66]. Pvmdr1 amplification rates of 4–37% have been observed in other regions of the country [30, 40]. Similarly, a 7–39% amplification rate was reported in Thailand [40, 62]. There was no pvmdr1 amplification in Indonesia [36, 67] and low prevalence in Brazil (0.9–4%) [67, 68]. The lack or low rate of pvmdr1 amplification in some areas of Thailand and Cambodia with intense MQ pressure does not support previous evidence associating pvmdr1 amplification with MQ pressure [67].
Although the pLDH-ELISA IC50 assay as described here represents an important first step to assess P. vivax drug susceptibility, the following caveats must be considered. The confounding factors of mechanism and speed of drug action must be taken into account when interpreting results obtained using different in vitro methods. High optical density (OD) background interference in wells containing maximal concentrations of some drugs tested caused low growth ratios and failure to achieve sigmoidal IC50 curves. This phenomenon was reported previously to depend on the nature of the drug being tested and detection methodology. Relatively higher OD background tends to be observed in non-artemisinin drugs with a more pronounced effect in ELISA-based assays [69, 70]. This may relate to the later onset of action of non-artemisinins in the parasite life cycle, allowing ring stage parasites to continue to produce the proteins being assayed despite high drug concentrations. This may explain paradoxical growth seen at high concentrations of non-artemisinins common in the pLDH-ELISA, but less frequently in the HRP2-ELISA, and not at all in the microscopy-based SMT [69, 70]. This corresponds with the observation of parasite growth in high concentrations of atovaquone, mefloquine, and quinine in the pLDH-ELISA assay for both P. falciparum and P. vivax, but not in the HRP2 ELISA or SMT. The SMT’s relatively low throughput, and challenges interpreting concentration–response curves for parasite cultures unable to develop to the threshold of ≥ 40% schizonts may limit its usefulness. The assay requires extensive experience on stage differentiation by operators to be effective and replicable. However, as the malaria map continues to shrink in Cambodia, it may be possible to better concentrate the expert microscopy skills needed to adequately perform the SMT in areas of greatest need.
Conclusion
Overall, the pLDH-ELISA has the potential to be a useful and replicable method to assess ex vivo P. vivax resistance in field isolates in Cambodia with reasonable throughput. Results correlated well with those observed for P. falciparum in both the pLDH and better established HRP-2 assays. While current malaria elimination efforts are focused on resistant P. falciparum malaria, P. vivax malaria may ultimately prove more difficult to eliminate due to its ease of transmission, dormancy, and only primaquine being effective at preventing P. vivax relapse. Current tools are also inadequate to differentiate relapse and recrudescence from re-infection. Evaluation of ex vivo P. vivax susceptibility using field isolates will likely prove a useful part of the armamentarium to monitor for growing resistance and predict risk of treatment failures. Clinical P. vivax resistance can be difficult to detect due to difficulty distinguishing relapsing disease from recrudescence and/or reinfection, and the pLDH assay may prove a useful adjunct in this regard. Incorporating ex vivo pLDH monitoring with therapeutic efficacy studies for individuals with P. vivax is advised to further validate this field-expedient method.
Authors’ contributions
Study design Cambodia: SC, CL, DS, and CL; data collection: all; data analysis and interpretation: SC, DS, SS, CL, CL, JJ, JL; wrote manuscript: SC, DS, and CL. All authors read and approved the final manuscript.
Acknowledgements
We are grateful to members of the AFRIMS Immunology and Medicine Department in Thailand and Cambodia not listed as coauthors for their intrepid technical support. We thank Mr. William Ellis at the Walter Reed Army Institute of Research (WRAIR), and the Medicines for Malaria Venture for providing reference drug supplies.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
The datasets generated and/or analysed during the current study are available from the corresponding author on reasonable request.
Consent for publication
All study subjects agreed to publication of the study results in the medical literature without their identity at consent prior to participation.
Disclaimer
The views expressed in this article are those of the authors and do not reflect the official policy of the US Department of the Army, US Department of Defense, or the US or Cambodian Governments.
Ethics approval and consent to participate
All protocols were approved by the Walter Reed Army Institute of Research (WRAIR) and the Cambodian National Ethics Committee for Health Research (NECHR). Protocol approval reference numbers include WR1576 (minimal risk). All study subjects provided informed consent prior to participation.
Funding
Funding for the study was provided by the US Armed Forces Health Surveillance Center and the National Institutes of Health, National Institutes of Allergy and Infectious Disease (Grant K08AI110651 to JTL and R21AI111108 to JJJ). The funding sources did not participate in data analysis or the final decision to publish the manuscript.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Abbreviations
- DHA
dihydroartemisinin
- AS
artesunate
- MQ
mefloquine hydrochloride
- QN
quinine sulfate hydrate
- CQ
chloroquine diphosphate
- PPQ
piperaquine phosphate
- ATQ
atovaquone
- LUM
lumefantrine
- DOX
doxycycline
- ATM
artemisone
- ATRs
artemisinins
- ACT
artemisinin-based combination therapy
- HRP-2
histidine-rich protein 2
- pLDH
Plasmodium pan-species lactate dehydrogenase
- ELISA
enzyme-linked immunosorbent assay
- SMT
schizont maturation test
- IC50
50% inhibitory concentration
- PCR
polymerase chain reaction
- OD
optical density
- Pvmdr1
Plasmodium vivax multi-drug resistance 1
- Ct
threshold cycle
- WHO
World Health Organization
Additional file
Additional file 1. Ex vivo drug susceptibility of P. vivax by years.
Footnotes
Electronic supplementary material
The online version of this article (doi:10.1186/s12936-017-2034-2) contains supplementary material, which is available to authorized users.
Contributor Information
Suwanna Chaorattanakawee, Email: suwann67@yahoo.com, Email: suwanna.cho@mahidol.ac.th.
Chanthap Lon, Email: ChanthapL@afrims.org.
Soklyda Chann, Email: ChannS.ctr@afrims.org.
Kheang Heng Thay, Email: thaykheanghengcnm@gmail.com.
Nareth Kong, Email: kongnarethcnm@gmail.com.
Yom You, Email: youyomcnm@gmail.com.
Siratchana Sundrakes, Email: dao_aj@yahoo.com.
Chatchadaporn Thamnurak, Email: ChatchadapornT.ca@afrims.org.
Sorayut Chattrakarn, Email: SorayutC.ca@afrims.org.
Chantida Praditpol, Email: ChantidaP.ca@afrims.org.
Kritsanai Yingyuen, Email: bomslee_yd@hotmail.com.
Mariusz Wojnarski, Email: mariusz.wojnarski.mil@afrims.org.
Rekol Huy, Email: kolhuy@gmail.com.
Michele D. Spring, Email: Michele.Spring.ctr@afrims.org
Douglas S. Walsh, Email: Douglas.Walsh@va.gov
Jaymin C. Patel, Email: jaymin86@gmail.com
Jessica Lin, Email: jessica_lin@med.unc.edu.
Jonathan J. Juliano, Email: jonathan_juliano@med.unc.edu
Charlotte A. Lanteri, Email: charlotte.a.lanteri.mil@mail.mil
David L. Saunders, Email: david.l.saunders.mil@mail.mil
References
- 1.Wongsrichanalai C, Pickard AL, Wernsdorfer WH, Meshnick SR. Epidemiology of drug-resistant malaria. Lancet Infect Dis. 2002;2:209–218. doi: 10.1016/S1473-3099(02)00239-6. [DOI] [PubMed] [Google Scholar]
- 2.Wongsrichanalai C, Sirichaisinthop J, Karwacki JJ, Congpuong K, Miller RS, Pang L, et al. Drug resistant malaria on the Thai-Myanmar and Thai-Cambodian borders. Southeast Asian J Trop Med Public Health. 2001;32:41–49. [PubMed] [Google Scholar]
- 3.Wernsdorfer WH, Payne D. The dynamics of drug resistance in Plasmodium falciparum. Pharmacol Ther. 1991;50:95–121. doi: 10.1016/0163-7258(91)90074-V. [DOI] [PubMed] [Google Scholar]
- 4.Bjorkman A, Phillips-Howard PA. The epidemiology of drug-resistant malaria. Trans R Soc Trop Med Hyg. 1990;84:177–180. doi: 10.1016/0035-9203(90)90246-B. [DOI] [PubMed] [Google Scholar]
- 5.Dondorp AM, Nosten F, Yi P, Das D, Phyo AP, Tarning J, et al. Artemisinin resistance in Plasmodium falciparum malaria. N Engl J Med. 2009;361:455–467. doi: 10.1056/NEJMoa0808859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Noedl H, Se Y, Schaecher K, Smith BL, Socheat D, Fukuda MM, Artemisinin Resistance in Cambodia 1 Study Consortium Evidence of artemisinin-resistant malaria in western Cambodia. N Engl J Med. 2008;359:2619–2620. doi: 10.1056/NEJMc0805011. [DOI] [PubMed] [Google Scholar]
- 7.Takala-Harrison S, Clark TG, Jacob CG, Cummings MP, Miotto O, Dondorp AM, et al. Genetic loci associated with delayed clearance of Plasmodium falciparum following artemisinin treatment in Southeast Asia. Proc Natl Acad Sci USA. 2013;110:240–245. doi: 10.1073/pnas.1211205110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Saunders DL, Vanachayangkul P, Lon C, U.S. Army Military Malaria Research Program. National Center for Parasitology Entomology and Malaria Control. Royal Cambodian Armed Forces Dihydroartemisinin–piperaquine failure in Cambodia. N Engl J Med. 2014;371:484–485. doi: 10.1056/NEJMc1403007. [DOI] [PubMed] [Google Scholar]
- 9.Chaorattanakawee S, Saunders DL, Sea D, Chanarat N, Yingyuen K, Sundrakes S, et al. Ex Vivo drug susceptibility testing and molecular profiling of clinical Plasmodium falciparum isolates from Cambodia from 2008 to 2013 suggest emerging piperaquine resistance. Antimicrob Agents Chemother. 2015;59:4631–4643. doi: 10.1128/AAC.00366-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Spring MD, Lin JT, Manning JE, Vanachayangkul P, Somethy S, Bun R, et al. Dihydroartemisinin-piperaquine failure associated with a triple mutant including kelch13 C580Y in Cambodia: an observational cohort study. Lancet Infect Dis. 2015;15:683–691. doi: 10.1016/S1473-3099(15)70049-6. [DOI] [PubMed] [Google Scholar]
- 11.Amaratunga C, Lim P, Suon S, Sreng S, Mao S, Sopha C, et al. Dihydroartemisinin–piperaquine resistance in Plasmodium falciparum malaria in Cambodia: a multisite prospective cohort study. Lancet Infect Dis. 2016;16:357–365. doi: 10.1016/S1473-3099(15)00487-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Duru V, Khim N, Leang R, Kim S, Domergue A, Kloeung N, et al. Plasmodium falciparum dihydroartemisinin–piperaquine failures in Cambodia are associated with mutant K13 parasites presenting high survival rates in novel piperaquine in vitro assays: retrospective and prospective investigations. BMC Med. 2015;13:305. doi: 10.1186/s12916-015-0539-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Baird JK, Basri H, Purnomo Bangs MJ, Subianto B, Patchen LC, et al. Resistance to chloroquine by Plasmodium vivax in Irian Jaya, Indonesia. Am J Trop Med Hyg. 1991;44:547–552. doi: 10.4269/ajtmh.1991.44.547. [DOI] [PubMed] [Google Scholar]
- 14.Baird JK, Sustriayu Nalim MF, Basri H, Masbar S, Leksana B, Tjitra E, et al. Survey of resistance to chloroquine by Plasmodium vivax in Indonesia. Trans R Soc Trop Med Hyg. 1996;90:409–411. doi: 10.1016/S0035-9203(96)90526-X. [DOI] [PubMed] [Google Scholar]
- 15.Murphy GS, Basri H, Purnomo Andersen EM, Bangs MJ, Mount DL, et al. Vivax malaria resistant to treatment and prophylaxis with chloroquine. Lancet. 1993;341:96–100. doi: 10.1016/0140-6736(93)92568-E. [DOI] [PubMed] [Google Scholar]
- 16.Rieckmann KH, Davis DR, Hutton DC. Plasmodium vivax resistance to chloroquine? Lancet. 1989;2:1183–1184. doi: 10.1016/S0140-6736(89)91792-3. [DOI] [PubMed] [Google Scholar]
- 17.Suwanarusk R, Russell B, Chavchich M, Chalfein F, Kenangalem E, Kosaisavee V, et al. Chloroquine resistant Plasmodium vivax: in vitro characterisation and association with molecular polymorphisms. PLoS ONE. 2007;2:e1089. doi: 10.1371/journal.pone.0001089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Marfurt J, de Monbrison F, Brega S, Barbollat L, Muller I, Sie A, et al. Molecular markers of in vivo Plasmodium vivax resistance to amodiaquine plus sulfadoxine–pyrimethamine: mutations in pvdhfr and pvmdr1. J Infect Dis. 2008;198:409–417. doi: 10.1086/589882. [DOI] [PubMed] [Google Scholar]
- 19.Suwanarusk R, Chavchich M, Russell B, Jaidee A, Chalfein F, Barends M, et al. Amplification of pvmdr1 associated with multidrug-resistant Plasmodium vivax. J Infect Dis. 2008;198:1558–1564. doi: 10.1086/592451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Imwong M, Pukrittayakamee S, Pongtavornpinyo W, Nakeesathit S, Nair S, Newton P, et al. Gene amplification of the multidrug resistance 1 gene of Plasmodium vivax isolates from Thailand, Laos, and Myanmar. Antimicrob Agents Chemother. 2008;52:2657–2659. doi: 10.1128/AAC.01459-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Marlar T, Myat Phone K, Aye YuS, Khaing Khaing G, Ma S, Myint O. Development of resistance to chloroquine by Plasmodium vivax in Myanmar. Trans R Soc Trop Med Hyg. 1995;89:307–308. doi: 10.1016/0035-9203(95)90556-1. [DOI] [PubMed] [Google Scholar]
- 22.Soto J, Toledo J, Gutierrez P, Luzz M, Llinas N, Cedeno N, et al. Plasmodium vivax clinically resistant to chloroquine in Colombia. Am J Trop Med Hyg. 2001;65:90–93. doi: 10.4269/ajtmh.2001.65.90. [DOI] [PubMed] [Google Scholar]
- 23.Phillips EJ, Keystone JS, Kain KC. Failure of combined chloroquine and high-dose primaquine therapy for Plasmodium vivax malaria acquired in Guyana, South America. Clin Infect Dis. 1996;23:1171–1173. doi: 10.1093/clinids/23.5.1171. [DOI] [PubMed] [Google Scholar]
- 24.Thanh PV, Hong NV, Van NV, Louisa M, Baird K, Xa NX, et al. Confirmed Plasmodium vivax resistance to chloroquine in Central Vietnam. Antimicrob Agents Chemother. 2015;59:7411–7419. doi: 10.1128/AAC.00791-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.World Health Organization . Guidelines for the treatment of malaria. 3. Geneva: World Health Organization; 2015. [PubMed] [Google Scholar]
- 26.Gogtay N, Kannan S, Thatte UM, Olliaro PL, Sinclair D. Artemisinin-based combination therapy for treating uncomplicated Plasmodium vivax malaria. Cochrane Database Syst Rev. 2013;10:CD008492. doi: 10.1002/14651858.CD008492.pub3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Poravuth Y, Socheat D, Rueangweerayut R, Uthaisin C, Pyae Phyo A, Valecha N, et al. Pyronaridine-artesunate versus chloroquine in patients with acute Plasmodium vivax malaria: a randomized, double-blind, non-inferiority trial. PLoS ONE. 2011;6:e14501. doi: 10.1371/journal.pone.0014501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Leang R, Barrette A, Bouth DM, Menard D, Abdur R, Duong S, et al. Efficacy of dihydroartemisinin–piperaquine for treatment of uncomplicated Plasmodium falciparum and Plasmodium vivax in Cambodia, 2008 to 2010. Antimicrob Agents Chemother. 2013;57:818–826. doi: 10.1128/AAC.00686-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Amaratunga C, Sreng S, Mao S, Tullo GS, Anderson JM, Chuor CM, et al. Chloroquine remains effective for treating Plasmodium vivax malaria in Pursat province, Western Cambodia. Antimicrob Agents Chemother. 2014;58:6270–6272. doi: 10.1128/AAC.03026-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Khim N, Andrianaranjaka V, Popovici J, Kim S, Ratsimbasoa A, Benedet C, Barnadas C, et al. Effects of mefloquine use on Plasmodium vivax multidrug resistance. Emerg Infect Dis. 2014;20:1637–1644. doi: 10.3201/eid2010.140411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Druilhe P, Brasseur P, Blanc C, Makler M. Improved assessment of Plasmodium vivax response to antimalarial drugs by a colorimetric double-site Plasmodium lactate dehydrogenase antigen capture enzyme-linked immunosorbent assay. Antimicrob Agents Chemother. 2007;51:2112–2116. doi: 10.1128/AAC.01385-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Rutvisuttinunt W, Chaorattanakawee S, Tyner SD, Teja-Isavadharm P, Se Y, Yingyuen K, et al. Optimizing the HRP-2 in vitro malaria drug susceptibility assay using a reference clone to improve comparisons of Plasmodium falciparum field isolates. Malar J. 2012;11:325. doi: 10.1186/1475-2875-11-325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Russell B, Suwanarusk R, Malleret B, Costa FT, Snounou G, Baird JK, et al. Human ex vivo studies on asexual Plasmodium vivax: the best way forward. Int J Parasitol. 2012;42:1063–1070. doi: 10.1016/j.ijpara.2012.08.010. [DOI] [PubMed] [Google Scholar]
- 34.Tyner SD, Lon C, Se Y, Bethell D, Socheat D, Noedl H, et al. Ex vivo drug sensitivity profiles of Plasmodium falciparum field isolates from Cambodia and Thailand, 2005 to 2010, determined by a histidine-rich protein-2 assay. Malar J. 2012;11:198. doi: 10.1186/1475-2875-11-198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Chaorattanakawee S, Lon C, Jongsakul K, Gawee J, Sok S, Sundrakes S, et al. Ex vivo piperaquine resistance developed rapidly in Plasmodium falciparum isolates in northern Cambodia compared to Thailand. Malar J. 2016;15:519. doi: 10.1186/s12936-016-1569-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Marfurt J, Chalfein F, Prayoga P, Wabiser F, Wirjanata G, Sebayang B, et al. Comparative ex vivo activity of novel endoperoxides in multidrug-resistant Plasmodium falciparum and P. vivax. Antimicrob Agents Chemother. 2012;56:5258–5263. doi: 10.1128/AAC.00283-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Russell B, Chalfein F, Prasetyorini B, Kenangalem E, Piera K, Suwanarusk R, et al. Determinants of in vitro drug susceptibility testing of Plasmodium vivax. Antimicrob Agents Chemother. 2008;52:1040–1045. doi: 10.1128/AAC.01334-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Druilhe P, Moreno A, Blanc C, Brasseur PH, Jacquier P. A colorimetric in vitro drug sensitivity assay for Plasmodium falciparum based on a highly sensitive double-site lactate dehydrogenase antigen-capture enzyme-linked immunosorbent assay. Am J Trop Med Hyg. 2001;64:233–241. doi: 10.4269/ajtmh.2001.64.233. [DOI] [PubMed] [Google Scholar]
- 39.Saunders DL, Chaorattanakawee S, Gosi P, Lanteri C, Somethy S, Kuntawunginn W, et al. Atovaquone-proguanil remains a potential stopgap therapy for multidrug-resistant Plasmodium falciparum in areas along the Thai-Cambodian border. Antimicrob Agents Chemother. 2015;60:1896–1898. doi: 10.1128/AAC.02302-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Lin JT, Patel JC, Kharabora O, Sattabongkot J, Muth S, Ubalee R, et al. Plasmodium vivax isolates from Cambodia and Thailand show high genetic complexity and distinct patterns of P. vivax multidrug resistance gene 1 (pvmdr1) polymorphisms. Am J Trop Med Hyg. 2013;88:1116–1123. doi: 10.4269/ajtmh.12-0701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Pfaffl MW. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001;29:e45. doi: 10.1093/nar/29.9.e45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Bourdier F. Health inequalities, public sector involvement and malaria control in Cambodia. Sojourn. 2016;31:81–115. doi: 10.1353/soj.2016.0018. [DOI] [Google Scholar]
- 43.Chaorattanakawee S, Lanteri CA, Sundrakes S, Yingyuen K, Gosi P, Chanarat N, et al. Attenuation of Plasmodium falciparum in vitro drug resistance phenotype following culture adaptation compared to fresh clinical isolates in Cambodia. Malar J. 2015;14:486. doi: 10.1186/s12936-015-1021-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Javelle E, Madamet M, Gaillard T, Velut G, Surcouf C, Michel R, et al. Delayed Onset of Plasmodium falciparum malaria after doxycycline prophylaxis in a soldier returning from the Central African Republic. Antimicrob Agents Chemother. 2016;60:2592–2593. doi: 10.1128/AAC.01858-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Kerlin DH, Boyce K, Marfurt J, Simpson JA, Kenangalem E, Cheng Q, et al. An analytical method for assessing stage-specific drug activity in Plasmodium vivax malaria: implications for ex vivo drug susceptibility testing. PLoS Negl Trop Dis. 2012;6:e1772. doi: 10.1371/journal.pntd.0001772. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Pratt-Riccio LR, Chehuan YF, Siqueira MJ, das Gracas Alecrim M, Bianco-Junior C, Druilhe P, et al. Use of a colorimetric (DELI) test for the evaluation of chemoresistance of Plasmodium falciparum and Plasmodium vivax to commonly used anti-plasmodial drugs in the Brazilian Amazon. Malar J. 2013;12:281. doi: 10.1186/1475-2875-12-281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Aguiar AC, Pereira DB, Amaral NS, De Marco L, Krettli AU. Plasmodium vivax and Plasmodium falciparum ex vivo susceptibility to anti-malarials and gene characterization in Rondonia, West Amazon, Brazil. Malar J. 2014;13:73. doi: 10.1186/1475-2875-13-73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Pava Z, Handayuni I, Wirjanata G, To S, Trianty L, Noviyanti R, et al. Expression of Plasmodium vivax crt-o is related to parasite stage but not ex vivo chloroquine susceptibility. Antimicrob Agents Chemother. 2015;60:361–367. doi: 10.1128/AAC.02207-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Wirjanata G, Sebayang BF, Chalfein F, Prayoga Handayuni I, Noviyanti R, et al. Contrasting ex vivo efficacies of “reversed chloroquine” compounds in chloroquine-resistant Plasmodium falciparum and P. vivax isolates. Antimicrob Agents Chemother. 2015;59:5721–5726. doi: 10.1128/AAC.01048-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Rungsihirunrat K, Muhamad P, Chaijaroenkul W, Kuesap J, Na-Bangchang K. Plasmodium vivax drug resistance genes; Pvmdr1 and Pvcrt-o polymorphisms in relation to chloroquine sensitivity from a malaria endemic area of Thailand. Korean J Parasitol. 2015;53:43–49. doi: 10.3347/kjp.2015.53.1.43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Sutanto I, Endawati D, Ling LH, Laihad F, Setiabudy R, Baird JK. Evaluation of chloroquine therapy for vivax and falciparum malaria in southern Sumatra, western Indonesia. Malar J. 2010;9:52. doi: 10.1186/1475-2875-9-52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Grigg MJ, William T, Menon J, Barber BE, Wilkes CS, Rajahram GS, et al. Efficacy of Artesunate–mefloquine for chloroquine-resistant Plasmodium vivax malaria in Malaysia: an open-label, randomized, controlled trial. Clin Infect Dis. 2016;62:1403–1411. doi: 10.1093/cid/ciw121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Siqueira AM, Alencar AC, Melo GC, Magalhaes BL, Machado K, Alencar Filho AC, et al. Fixed-dose artesunate–amodiaquine combination vs chloroquine for treatment of uncomplicated blood stage P. vivax infection in the Brazilian Amazon: an open-label randomized, controlled trial. Clin Infect Dis. 2017;64:166–174. doi: 10.1093/cid/ciw706. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Nyunt MH, Han JH, Wang B, Aye KM, Aye KH, Lee SK, et al. Clinical and molecular surveillance of drug resistant vivax malaria in Myanmar (2009–2016) Malar J. 2017;16:117. doi: 10.1186/s12936-017-1770-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Singh L, Rathi S, Kondekar S, Kondekar A, Dongare S. Per protocol analysis of therapeutic efficacy of chloroquine for the treatment of Plasmodium vivax malaria in children. Int J Contemp Pediatr. 2016;3:725–729. doi: 10.18203/2349-3291.ijcp20161909. [DOI] [Google Scholar]
- 56.Seifu S, Zeynudin A, Zemene E, Suleman S, Biruksew A. Therapeutic efficacy of chloroquine for the treatment of Plasmodium vivax malaria among outpatients at Shawa Robit Health Care Centre, North-East Ethiopia. Acta Trop. 2017;171:44–51. doi: 10.1016/j.actatropica.2017.02.027. [DOI] [PubMed] [Google Scholar]
- 57.World Health Organization . National treatment guidelines for malaria in Cambodia. Geneva: World Health Organization; 2012. [Google Scholar]
- 58.Hayward R, Saliba KJ, Kirk K. pfmdr1 mutations associated with chloroquine resistance incur a fitness cost in Plasmodium falciparum. Mol Microbiol. 2005;55:1285–1295. doi: 10.1111/j.1365-2958.2004.04470.x. [DOI] [PubMed] [Google Scholar]
- 59.Henry M, Alibert S, Orlandi-Pradines E, Bogreau H, Fusai T, Rogier C, et al. Chloroquine resistance reversal agents as promising antimalarial drugs. Curr Drug Targets. 2006;7:935–948. doi: 10.2174/138945006778019372. [DOI] [PubMed] [Google Scholar]
- 60.Melo GC, Monteiro WM, Siqueira AM, Silva SR, Magalhaes BM, Alencar AC, et al. Expression levels of pvcrt-o and pvmdr-1 are associated with chloroquine resistance and severe Plasmodium vivax malaria in patients of the Brazilian Amazon. PLoS ONE. 2014;9:e105922. doi: 10.1371/journal.pone.0105922. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Barnadas C, Timinao L, Javati S, Iga J, Malau E, Koepfli C, et al. Significant geographical differences in prevalence of mutations associated with Plasmodium falciparum and Plasmodium vivax drug resistance in two regions from Papua New Guinea. Malar J. 2015;14:399. doi: 10.1186/s12936-015-0879-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Lu F, Lim CS, Nam DH, Kim K, Lin K, Kim TS, et al. Genetic polymorphism in pvmdr1 and pvcrt-o genes in relation to in vitro drug susceptibility of Plasmodium vivax isolates from malaria-endemic countries. Acta Trop. 2011;117:69–75. doi: 10.1016/j.actatropica.2010.08.011. [DOI] [PubMed] [Google Scholar]
- 63.Auburn S, Serre D, Pearson RD, Amato R, Sriprawat K, To S, et al. Genomic analysis reveals a common breakpoint in amplifications of the Plasmodium vivax multidrug resistance 1 locus in Thailand. J Infect Dis. 2016;214:1235–1242. doi: 10.1093/infdis/jiw323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Givens MB, Lin JT, Lon C, Gosi P, Char MC, Lanteri CA, et al. Development of a capillary electrophoresis-based heteroduplex tracking assay to measure in-host genetic diversity of initial and recurrent Plasmodium vivax infections in Cambodia. J Clin Microbiol. 2014;52:298–301. doi: 10.1128/JCM.02274-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Lin JT, Ubalee R, Lon C, Balasubramanian S, Kuntawunginn W, Rahman R, et al. Microscopic Plasmodium falciparum gametocytemia and infectivity to mosquitoes in Cambodia. J Infect Dis. 2016;213:1491–1494. doi: 10.1093/infdis/jiv599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Parobek CM, Lin JT, Saunders DL, Barnett EJ, Lon C, Lanteri CA, et al. Selective sweep suggests transcriptional regulation may underlie Plasmodium vivax resilience to malaria control measures in Cambodia. Proc Natl Acad Sci USA. 2016;113:E8096–E8105. doi: 10.1073/pnas.1608828113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Costa GL, Amaral LC, Fontes CJF, Carvalho LH, de Brito CFA, de Sousa TN. Assessment of copy number variation in genes related to drug resistance in Plasmodium vivax and Plasmodium falciparum isolates from the Brazilian Amazon and a systematic review of the literature. Malar J. 2017;16:152. doi: 10.1186/s12936-017-1806-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Vargas-Rodriguez Rdel C, da Silva Bastos M, Menezes MJ, Orjuela-Sanchez P, Ferreira MU. Single-nucleotide polymorphism and copy number variation of the multidrug resistance-1 locus of Plasmodium vivax: local and global patterns. Am J Trop Med Hyg. 2012;87:813–821. doi: 10.4269/ajtmh.2012.12-0094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Woodrow CJ, Dahlstrom S, Cooksey R, Flegg JA, Le Nagard H, Mentre F, et al. High-throughput analysis of antimalarial susceptibility data by the WorldWide Antimalarial Resistance Network (WWARN) in vitro analysis and reporting tool. Antimicrob Agents Chemother. 2013;57:3121–3130. doi: 10.1128/AAC.02350-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Woodrow CJ, Wangsing C, Sriprawat K, Christensen PR, Nosten F, Renia L, et al. Comparison between flow cytometry, microscopy, and lactate dehydrogenase-based enzyme-linked immunosorbent assay for Plasmodium falciparum drug susceptibility testing under field conditions. J Clin Microbiol. 2015;53:3296–3303. doi: 10.1128/JCM.01226-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets generated and/or analysed during the current study are available from the corresponding author on reasonable request.