

Abstract
Biofilms help bacteria survive under adverse conditions, and the quorum sensing (QS) system plays an important role in regulating their activities. Quorum sensing inhibitors (QSIs) have great potential to inhibit pathogenic biofilm formation and are considered possible replacements for antibiotics; however, further investigation is required to understand the mechanisms of action of QSIs and to avoid inhibitory effects on beneficial bacteria. Lactobacillus paraplantarum L-ZS9, isolated from fermented sausage, is a bacteriocin-producing bacteria that shows potential to be a probiotic starter. Since exogenous autoinducer-2 (AI-2) promoted biofilm formation of the strain, expression of genes involved in AI-2 production was determined in L. paraplantarum L-ZS9, especially the key gene luxS. D-Ribose was used to inhibit biofilm formation because of its AI-2 inhibitory activity. Twenty-seven differentially expressed proteins were identified by comparative proteomic analysis following D-ribose treatment and were functionally classified into six groups. Real-time quantitative PCR showed that AI-2 had a counteractive effect on transcription of the genes tuf, fba, gap, pgm, nfo, rib, and rpoN. Over-expression of the tuf, fba, gap, pgm, and rpoN genes promoted biofilm formation of L. paraplantarum L-ZS9, while over-expression of the nfo and rib genes inhibited biofilm formation. In conclusion, D-ribose inhibited biofilm formation of L. paraplantarum L-ZS9 by regulating multiple genes involved in the glycolytic pathway, extracellular DNA degradation and transcription, and translation. This research provides a new mechanism of QSI regulation of biofilm formation of Lactobacillus and offers a valuable reference for QSI application in the future.
Keywords: Lactobacillus paraplantarum L-ZS9, quorum sensing inhibitor, biofilms, proteomics, recombinant strains
Introduction
Bacteria live in dense and diverse communities termed biofilms, which is a major mode of microbial life (Nadell et al., 2016). Biofilms are sessile microbial communities that attach to biotic and abiotic surfaces and survive as self-organized, three-dimensional structures by producing an extracellular polymeric matrix (Penesyan et al., 2015). This lifestyle helps microorganisms to survive in unfavorable environments (Taylor et al., 2014; Olsen, 2015). Biofilms of pathogenic bacteria may cause antibiotic-tolerant infections as well as damage to surfaces and flow systems (Hoiby et al., 2010; Bixler and Bhushan, 2012; Drescher et al., 2013). However, similar to some non-pathogenic microorganisms and beneficial bacteria, biofilms enable pathogenic bacteria to resist environmental conditions, leading to successful colonization and maintenance of their population (Lepargneur and Rousseau, 2002).
Within biofilms, bacteria use a quorum sensing (QS) system to communicate with each other. The LuxS/AI-2 QS system exists widely in both Gram-negative and Gram-positive bacteria. The signal molecule, autoinducer-2 (AI-2), has been deemed as a universal language for intra-species and inter-species communication (Schauder et al., 2001; Gospodarek et al., 2009). The luxS AI-2 synthase gene homolog has been found in a wide range of bacteria, with 17% of the phylum Bacteroidetes and 83% of the Firmicutes predicted to have the homolog according to the KEGG database (Thompson et al., 2015). Although most research has focused on the regulation of AI-2 in biofilms of pathogens, some non-pathogenic and beneficial bacteria have been reported to use this ‘universal’ and ‘common’ signaling system to regulate their behavior (Park et al., 2016). For example, AI-2 signaling has been reported to regulate cell growth and metabolism of Lactobacillus (Buck et al., 2009; Moslehi-Jenabian et al., 2009; Park et al., 2016). And in Bifidobacteria, AI-2 activity correlates with biofilm formation and gut colonization (Christiaen et al., 2014; Sun et al., 2014).
Since QS inhibitors (QSIs) hinder biofilm formation and reduce bacterial virulence in the biofilm state, they are considered as an ideal tool to inhibit the biofilm formation of pathogens and provide a good target to control bacterial infection (Pan and Ren, 2009; Harjai et al., 2014). An ideal QSI should significantly interfere with the QS system and social behavior without toxic effects on the bacteria and/or host. Ribose has no toxic effects and shares structural similarity with AI-2, which exhibits a furanosyl borate diether form (Cao and Meighen, 1989). This resemblance has been thought to cause competition between AI-2 and ribose. Thus, ribose has been used as a QSI to inhibit biofilm formation of various pathogenic bacteria (Armbruster et al., 2011; Lee et al., 2015; Cho et al., 2016; Ryu et al., 2016). However, the underlying mechanism of biofilm inhibition of non-pathogenic and beneficial bacteria still needs to be investigated.
Recently, LuxS/AI-2 QSIs have shown great potential to replace antibiotics to control pathogenic biofilm formation and infection (Brackman et al., 2009, 2011; Roy et al., 2013; Ryu et al., 2016). However, adequate considerations should be noted when QS therapy develops rapidly. Because of the common existence of LuxS/AI-2 QS in microorganisms, LuxS/AI-2 QSIs may inhibit biofilm formation of non-pathogenic and beneficial organisms as well. Lactobacillus paraplantarum L-ZS9 was originally isolated from fermented sausage and was proven to produce class IIb bacteriocins to inhibit the growth of pathogenic bacteria in our previous study (Wang et al., 2015; Zhang et al., 2016). It has the potential to be used as a probiotic starter. This study determined AI-2 production in L-ZS9 and AI-2 inhibitory activity by D-ribose. The effects of D-ribose on biofilm formation of L-ZS9 and its underlying mechanism were investigated and analyzed. This study provides new information about the regulation mechanism of this QSI on biofilm formation of Lactobacillus and has value as a reference for the reasonable application of QSI.
Materials and Methods
Bacterial Strains and Culture
Lactobacillus paraplantarum L-ZS9 was incubated at 37°C in de Man-Rogosa-Sharpe (MRS) broth (Bridge, Beijing) and MRS agar aerobically. Escherichia coli DH5α (Takara, Dalian) was incubated in Lennox broth or on solid medium with 1.5% (w/v) agar at 37°C. DH5α and L-ZS9 transformed with pMG76e vector were cultured with 200 and 3 μg/mL erythromycin, respectively. The pMG76e vector was provided by Professor Shangwu Chen from China Agricultural University (CAU, Beijing, China). Vibrio harveyi BB170 and BB152 were provided by Professor Xiangan Han from Shanghai Veterinary Research Institute (CAAS, Shanghai, China) and cultured in Marine Broth 2216 (Difco, United States) or Autoinducer Bioassay (AB) medium (Bassler et al., 1993).
Analysis of Genes Related to AI-2 Production
The complete genome sequence of L. paraplantarum L-ZS9 is deposited in GenBank with accession number CP013130 (Liu and Li, 2016). The pfs, luxS, metE/metH, and metK genes involved in AI-2 production and the sahH gene responsible for metabolism of S-adenosylhomocysteine (SAH) to S-ribosylhomocysteine in one-step were searched for in the genome sequence of L. paraplantarum L-ZS9 according to their gene annotations by using the BLAST program of NCBI. Sizes of these genes were analyzed by Primer Input 3.01, and locations were determined by analyzing the genome map.
Growth-Curve Assay
To determine the effects of AI-2 and D-ribose on the growth of L. paraplantarum L-ZS9, a growth-curve assay was conducted. L. paraplantarum L-ZS9 was cultured in MRS broth in the presence or absence of AI-2 (3.7, 18.5, 37, 185 μM) (initial concentration 3.7 mM, purchased from OMM Scientific, United States). L. paraplantarum L-ZS9 was also cultured in MRS broth in the presence or absence of D-ribose (0, 10, 50, and 100 mM). Cultures were incubated at 37°C for 36 h. After 3, 6, 9, 12, 15, 18, 21, 24, 27, 30, 33, and 36 h, the optical density at 600 nm was determined using a UV-1800 Spectrophotometer (Shanghai Meipuda instrument, China).
Determination of AI-2-Mediated Bioluminescence and AI-2 Inhibitory Activity of D-Ribose
Lactobacillus paraplantarum L-ZS9 was cultured in 12% (w/v) skim milk medium. After culture for 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, and 24 h, cells were collected by centrifuging at 12,000 g, 4°C, for 10 min. The cell-free culture fluid (CF) was obtained by filtering through a 0.22-μm filter (Millipore, Bedford, MA, United States) and adjusted to pH 7.0. The reporter strain V. harveyi BB170 was diluted 1:5000 with AB medium, and the CF sample was added to the diluted BB170 culture at 1:10 (v/v). The mixture was incubated at 28°C for 5 h, and 100 μL aliquots were added to white, flat-bottomed, 96-well plates (Thermo Labsystems, Franklin, MA, United States) to detect AI-2 activity. The CF collected from BB152 and DH5α cultures was used as a positive and negative control, respectively. Luminescence was measured using a Tecan GENios Plus microplate reader in luminescence mode (Tecan Austria GmbH, Grodig, Austria).
D-Ribose inhibition of BB152 and L-ZS9 AI-2 activity was also determined by using the BB170 reporter strain. D-Ribose (0, 10, 50, and 100 mM) was added to AB medium containing diluted BB170 culture (1:5000, v/v) with the CF of BB152 (1:10, v/v) or L-ZS9 (1:10, v/v). After incubation at 28°C for 5 h, AI-2 activity was detected as described above.
Biofilm Formation Assay
Crystal violet (CV) staining was used to quantify biofilm formation of L. paraplantarum L-ZS9. Briefly, overnight culture of L. paraplantarum L-ZS9 was diluted to an optical density of 0.1 at 600 nm. Diluted culture (200 μL) was transferred to a 96-well plate (Corning, United States). Different concentrations of AI-2 (3.7, 18.5, 37, 185 μM), D-ribose (10, 50, and 100 mM) was added, and the mixture was incubated at 37°C for 36 h. The wells were washed gently three times with phosphate-buffered saline, stained with 0.1% CV for 30 min at room temperature, rinsed with distilled water, air-dried, and 100 μL 95% ethanol added to dissolve the CV. The absorbance at 595 nm was determined using a Synergy 2 microplate reader (Biotek, Winooski, VT, United States).
Proteomic Analysis
Lactobacillus paraplantarum L-ZS9 treated with and without D-ribose (100 mM) was used for gel-based proteome analysis. Briefly, total-cell protein was extracted by trichloroacetic acid / acetone precipitation (Lakshman et al., 2008). Isolated protein was resuspended in lysis buffer (7 M urea, 2 M thiourea, 2%, w/v, CHAPS) and stored at -80°C until analysis. Protein concentration was determined by Bradford protein assay (Bio-Rad).
For electrophoresis, 400 μg of total protein was loaded onto immobilized pH-gradient strips (17 cm, non-linear pH 4-7; Bio-Rad). Isoelectric focusing and sodium dodecyl sulfate polyacrylamide gel electrophoresis were carried out as previously described (Wang et al., 2006). After electrophoresis, gels were stained with Colloidal Coomassie Brilliant Blue G-250, imaged using an ImageScanner (Amersham Biosciences), and analyzed by ImageMaster 2D Platinum 6.0 software (Wang et al., 2006). Protein spots with ≥1.5-fold change in abundance (p < 0.05) were subjected to protein identification.
The selected protein spots were excised, and in-gel digestion was performed with 0.01 μg/μL trypsin (Promega, Madison, WI, United States) in 25 mM ammonium bicarbonate. Tryptic peptides were dissolved in 0.5% (w/v) trifluoroacetic acid. Matrix-assisted laser desorption/ionization time-of-flight mass spectrometry and tandem time-of-flight/time-of-flight mass spectrometry were carried out on a 4800 Proteomics Analyzer (Applied Biosystems, United States) as described previously (Wang et al., 2006). Combined mass with mass/mass spectra was used to interrogate protein sequences in the NCBI database using the MASCOT database search algorithms (version 2.1, Matrix Science, London, United Kingdom). A protein was reported as identified if the Mascot score confidence interval > 95%. Functional analyses of the differentially expressed proteins were conducted using the Universal Protein Resource2 and the Kyoto Encyclopedia of Genes and Genomes3.
Quantitative Real-Time PCR (qRT-PCR)
Lactobacillus paraplantarum L-ZS9 was cultured in MRS with D-ribose (100 mM) or AI-2 (18.5 μM) at 37°C to logarithmic phase. L. paraplantarum L-ZS9 cultured without AI-2 or D-ribose was used as a control. Total RNA was extracted with TRIzol (Invitrogen, United States) according to the manufacturer’s instructions. RNA quality was determined by measuring A260/A280 and A260/A230 and by gel electrophoresis. Isolated RNA was transcribed into single-stranded cDNA using a TUREscript 1st Strand cDNA Synthesis Kit (Aidlab, China). qRT-PCR was performed by using SYBR Green assay kit (Tiangen, China) and a 7500 Fast Real-Time PCR system (Applied Biosystems). Primers were designed by Primer 3 Input4. The 16S rRNA gene was used as an internal reference. The relative expression of specific genes was calculated by using the 2-ΔΔCT method according to Livak and Schmittgen (2001).
Construction of Plasmids and Bacterial Strains
Chromosomal DNA of L. paraplantarum L-ZS9 was isolated using a TIANamp Bacteria DNA Kit (Tiangen Biotech, Beijing). The genes tuf, fba, gap, pgm, nfo, rib, and rpoN were amplified from the chromosomal DNA by PCR and cloned into pMD18T vector (Takara, Dalian). The constructed tuf, fba, gap, pgm, nfo, rib, and rpoN-pMD18T vectors and the pMG76e vector (provided by Professor Shangwu Chen, China Agricultural University) were digested with Fastdigest enzymes XbaI and XhoI (Thermo, United States). The gene fragments were inserted into the linearized pMG76e plasmid using a Rapid DNA Ligation Kit (Thermo, United States), and the constructed vector was transformed into E. coli DH5α. The constructed plasmids harboring the genes of interest were identified by PCR and extracted by using a TIANpure Midi Plasmid Kit (Tiangen, China). Constructed pMG76e and empty pMG76e plasmids were electrotransformed into L. paraplantarum L-ZS9 competent cells, and recombinant strains were selected with erythromycin and confirmed by PCR.
Biofilm Formation of Recombinant Strains
The growth of recombinant strains was analyzed by measuring the optical density at 600 nm to determine the effect of the expression the genes of interest on the growth of L-ZS9. Biofilm formation of recombinant strains over-expressing tuf, fba, gap, pgm, nfo, rib, and rpoN was measured by CV staining. Overnight cultures of recombinant strains and the wild-type strain were diluted in MRS without erythromycin to an optical density of 0.1 at 600 nm, and 200 μL of each suspension was transferred to a 96-well plate (Corning, NY, United States). After incubation at 37°C for 36 h, the biofilm formation of these strains was measured as described above.
Statistical Analysis
The data were analyzed by one-way analysis of variance (ANOVA) using Graphpad Prism 5.0. Data are presented as means ± SEM. A p-value less than 0.05 was considered as significant.
Results
L. paraplantarum L-ZS9 Contains All the Genes Responsible for AI-2 Production But Not sahH
Biosynthesis of AI-2 is part of the methionine catabolism cycle. MetE/MetH is responsible for transforming homocysteine into methionine. Methionine is then converted to S-adenosylmethionine (SAM) in a reaction catalyzed by MetK. Methyltransferase transforms SAM to SAH, which is converted to homocysteine through Pfs and LuxS, and AI-2 is produced (Schauder et al., 2001; Parveen and Cornell, 2011). Complete genome analysis showed that L. paraplantarum L-ZS9 contains all the genes involved in AI-2 production, including pfs, luxS, metE/metH, metK, and the methyltransferase gene, but sahH or its homologous gene, responsible for changing SAH to homocysteine directly, was not found (Figure 1A). These results indicated that L. paraplantarum L-ZS9 metabolized SAH to homocysteine through Pfs and LuxS, and produced AI-2 as a byproduct. The size and location of these genes are shown in Figure 1B.
FIGURE 1.
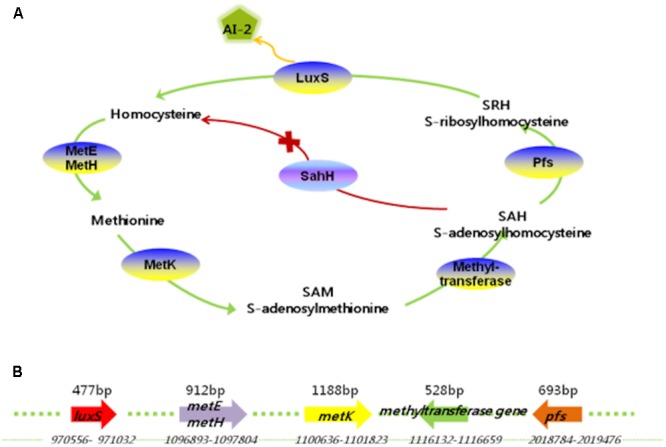
AI-2 production pathway (A) and sizes and locations of related genes (B) in L. paraplantarum L-ZS9.
L. paraplantarum L-ZS9 Produced AI-2 When Cultured in Skim Milk Medium
AI-2 production was measured by using the reporter strain V. harveyi BB170 in a bioassay with CFs of L. paraplantarum L-ZS9 cultured in skim milk medium. V. harveyi BB152 CF with confirmed AI-2 activity (7492.604 relative light units (RLUs)) was used as a positive control, E. coli DH5α CF with no AI-2 activity (321.805 RLUs) was used as a negative control, and V. harveyi BB170 with no CF (307.740 RLUs) was used as a blank control. AI-2 activity was detected in CFs prepared from L. paraplantarum L-ZS9 grown in skim milk medium and increased with increasing incubation time to reach a maximum value (4864.748 RLUs) after 22 h (Figure 2). These results suggested that L. paraplantarum L-ZS9 can produce AI-2 and that the AI-2 concentration depended on the incubation time.
FIGURE 2.
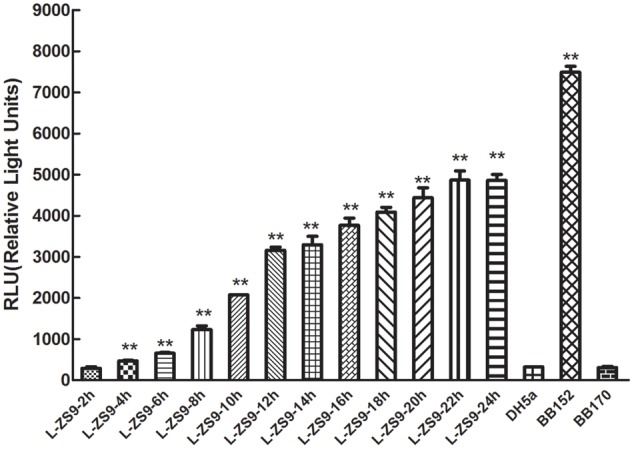
AI-2 activity in cell-free culture fluids (CFs) of L. paraplantarum L-ZS9. V. harveyi BB152 served as a positive control and E. coli DH5α as a negative control. Data are presented as mean ± SEM. n ≥ 3. ∗p < 0.05, ∗∗p < 0.01 compared with the negative control.
Inhibitory Effect of D-Ribose on AI-2 Activity of BB152 and L-ZS9
As shown in Figures 3A,B, exogenous AI-2 and D-ribose had no influence on growth of L. paraplantarum L-ZS9. D-Ribose significantly inhibited the AI-2 activity of V. harveyi BB152 and L. paraplantarum L-ZS9 in a dose-dependent manner (Figures 4A,B); 100 mM D-ribose inhibited AI-2 activity of V. harveyi BB152 and L. paraplantarum L-ZS9 to about 0.10-fold and 0.13-fold, respectively. The result indicated that D-ribose could be used as a QSI against AI-2 activity of L-ZS9.
FIGURE 3.
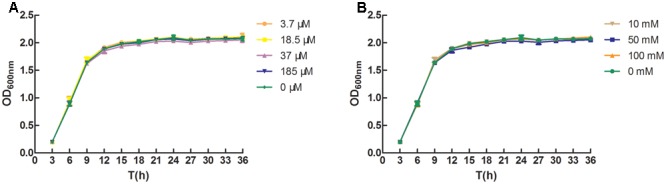
Effects of AI-2 (A) and D-ribose (B) on growth of L. paraplantarum L-ZS9. Growth of the strain in the absence of AI-2 or D-ribose served as a control. Data are presented as mean ± SEM. n ≥ 3. ∗p < 0.05, ∗∗p < 0.01 compared with the control.
FIGURE 4.
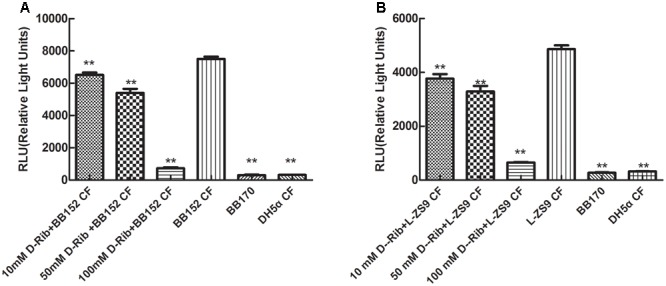
Effect of D-ribose on AI-2 activity of V. harveyi BB152 (A) and L. paraplantarum L-ZS9 (B). E. coli DH5α served as a negative control. Data are presented as mean ± SEM. n ≥ 3. ∗p < 0.05, ∗∗p < 0.01 compared with the positive control.
Inhibitory Effect of D-Ribose on Biofilm Formation of L-ZS9 Induced by AI-2
Figure 5A shows that 3.7–37.0 μM AI-2 increased biofilm formation of L. paraplantarum L-ZS9, which peaked at 18.5 μM AI-2. Different concentrations of AI-2 showed different effects on biofilm formation of L-ZS9. The effect of AI-2 on biofilm formation was not dose-dependent; the highest biofilm formation was observed when cultures were treated with 18.5 μM AI-2, and decreased with increasing concentration of AI-2. On the contrary, D-ribose inhibited the biofilm formation of L-ZS9 in a dose-dependent manner, and 100 mM D-ribose inhibited the biofilm formation of L-ZS9 to about 0.25-fold compared with the control (Figure 5B).
FIGURE 5.
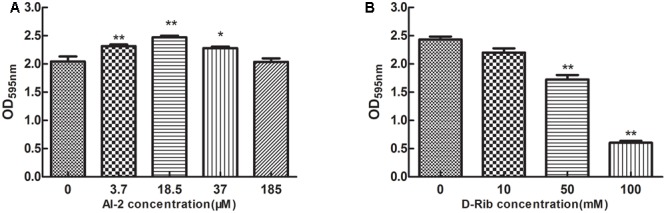
Effect of AI-2 (A) and D-ribose (B) on biofilm formation of L. paraplantarum L-ZS9. The biofilm formation of the strain in the absence of AI-2 or D-ribose served as a control. Data are presented as mean ± SEM. n ≥ 3. ∗p < 0.05, ∗∗p < 0.01 compared with the control.
Protein Identification in L. paraplantarum L-ZS9 Treated with or without D-Ribose
To profile the impact of D-ribose on protein expression, L. paraplantarum L-ZS9 was cultured with or without D-ribose. The proteomes were comparatively studied by gel-based proteomic analysis. Preliminary 2D gel electrophoresis analysis with a wide pH gradient (pH 3–10) showed that the majority of proteins had their isoelectric point (pI) in the acidic region (data not shown). Therefore an immobilized pH-gradient strip with a pH gradient of 4–7 was used in this study. As shown in Figure 6, 27 differentially expressed proteins were detected (P < 0.05) (details of biological replicates are provided in the Supplementary Material). Among them, 15 proteins showed up-regulation and 12 proteins showed down-regulation by at least 1.5-fold following D-ribose treatment. The molecular weight (MW) and pI of these proteins are listed in Table 1. Functionally, these differentially expressed proteins were classified into six groups that related to carbon and carbohydrate metabolism, amino acid metabolism, nucleotide transport and metabolism, fatty acid metabolism, transcription and translation, and other functions. Function classification of these proteins suggested that D-ribose may inhibit biofilm through multiple mechanisms, which include regulating translation and transcriptional activity, carbohydrate and energy metabolism, amino acid synthesis, and protein and enzyme secretion.
FIGURE 6.
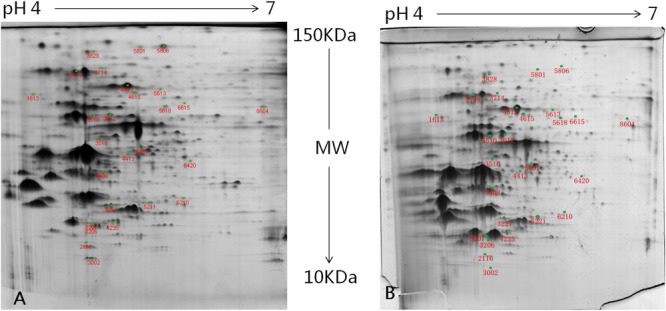
Two-dimensional electrophoresis of the total-cell proteins of L. paraplantarum L-ZS9 treated without (A) or with D-ribose (B). Differentially expressed protein spots of cells treated with or without D-ribose were numbered according to the numbering in Table 1.
Table 1.
Functional classification of differentially expressed proteins in cells of L. paraplantarum L-ZS9 treated with or without D-ribose.
| Spot no. | Accession no. | Gene | Protein name | Protein MW | Protein pI | Fold changea |
|---|---|---|---|---|---|---|
| Carbon metabolism and carbohydrate metabolism | ||||||
| 2719 | gi| 380033724 | pgm | Phosphoglycerate mutase family protein | 26071.3 | 4.94 | 0.56 |
| 3510 | gi| 545603975 | rib | Ribokinase | 32080.3 | 5.02 | 3.24 |
| 3612 | gi| 545603975 | rib | Ribokinase | 32080.3 | 5.02 | 3.01 |
| 4613 | gi| 380031375 | fba | Fructose-bisphosphate aldolase | 30901.6 | 5.07 | 0.47 |
| 5618 | gi| 254555821 | gap | Glyceraldehyde-3-phosphate dehydrogenase | 36415.6 | 5.3 | 0.54 |
| 5613 | gi| 380031375 | fba | Fructose-bisphosphate aldolase | 30901.6 | 5.07 | 2.34 |
| 3206 | gi| 545606863 | ptk | Phosphoketolase | 88690.1 | 5.05 | 5.12 |
| Amino acid metabolism | ||||||
| 6210 | gi| 545603889 | asnB | Asparagine synthase | 73077.3 | 5.72 | 0.21 |
| 6420 | gi| 545606144 | murD | UDP-N-acetylmuramoyl-L-alanyl-D-glutamate synthetase | 49993.6 | 5.75 | 1.67 |
| Nucleotide transport and metabolism | ||||||
| 3828 | gi| 254556519 | gmk | Guanylate kinase | 23445.7 | 4.99 | 2.33 |
| 1613 | gi| 545605966 | rihC | Ribonucleoside hydrolase RihC | 33030.6 | 4.67 | 2.18 |
| 3516 | gi| 545605039 | guaC | Guanosine 5′-monophosphate oxidoreductase | 39843.3 | 5.21 | 0.3 |
| 4413 | gi| 544928185 | purA | Adenylosuccinate synthetase | 47207.3 | 5.25 | 0.46 |
| 6615 | gi| 545604536 | nfo | Endonuclease IV | 31603.1 | 5.65 | 2.1 |
| Amino acid metabolism | ||||||
| 4615 | gi| 545605392 | smp | S-malonyltransferase | 33242.1 | 5.59 | 1.77 |
| 5211 | gi| 513840005 | lai | Linoleic acid isomerase | 64204.6 | 5.37 | 1.74 |
| Transcription and translation | ||||||
| 2116 | gi| 545606085 | tuf | Elongation factor Tu | 43364.1 | 4.95 | 0.1 |
| 3002 | gi| 550695603 | rpoB | DNA-directed RNA polymerase subunit beta | 131576.3 | 4.87 | 0.02 |
| 3207 | gi| 545606497 | fusA | Elongation factor G | 76921.7 | 4.82 | 9.91 |
| 3227 | gi| 254556435 | thrS | Threonyl-tRNA synthetase | 73773 | 5.1 | 0.43 |
| 3408 | gi| 545606490 | serS | Seryl-tRNA synthetase | 48103.2 | 5.15 | 2.2 |
| 5801 | gi| 545605166 | rpoN | Sigma 54 factor, RpoN | 21724.2 | 5.47 | 0.14 |
| 5806 | gi| 545605166 | rpoN | Sigma 54 factor, RpoN | 21724.2 | 5.47 | 0.03 |
| Other | ||||||
| 3714 | gi| 254556461 | rrg | Response regulator | 26356 | 5.06 | 0.55 |
| 4235 | gi| 545605165 | secA | Preprotein translocase subunit SecA | 89545.2 | 5.16 | 3.13 |
| 5401 | gi| 545603952 | nirD | Assimilatory nitrite reductase, subunit | 44407.8 | 5.51 | 2.2 |
| 8604 | gi| 545604158 | adh | Aryl-alcohol dehydrogenase family enzyme | 36955.8 | 6.71 | 2.03 |
aD-Ribose-treated vs. untreated cells.
Effects of D-Ribose and AI-2 on Gene Transcription of L. paraplantarum L-ZS9
As shown in Figure 7, D-ribose changed the transcription of the tuf, fba, gap, pgm, nfo, rib, and rpoN genes of L. paraplantarum L-ZS9, which was consistent with the effect of D-ribose on their protein levels. These genes were selected from those encoding the differentially expressed proteins of the above 2D polyacrylamide gel electrophoresis results based on their relationship to QS, which has been reported in previous research (Wolfe et al., 2004; Da et al., 2008; Kiedrowski et al., 2011; Iyer and Hancock, 2012; Sahu et al., 2012; Zhou et al., 2012; Hao et al., 2013; Li Z. et al., 2015; Rajendran et al., 2015). D-Ribose inhibited the transcription of tuf, pgm, fab, gap, and rpoN to about 0.39-fold, 0.56-fold, 0.20-fold, 0.27-fold, and 0.78-fold, respectively, while it increased the transcription of nfo and rib to about 2.57-fold and 4.87-fold, respectively.
FIGURE 7.
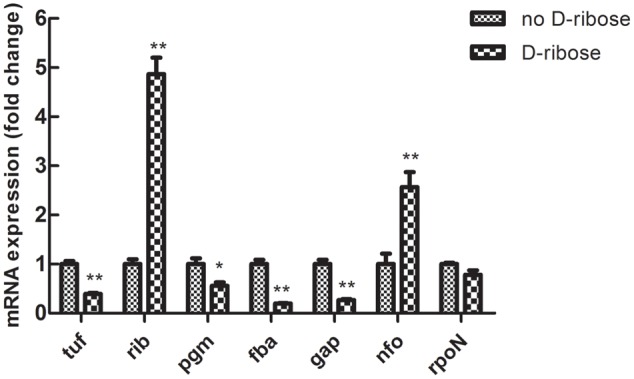
Effects of D-ribose (100 mM) on mRNA expression of tuf, fba, gap, pgm, nfo, rib, and rpoN genes in L. paraplantarum L-ZS9. The mRNA expression of these genes in the absence of D-ribose served as a control. Data are presented as mean ± SEM. n ≥ 3. ∗p < 0.05, ∗∗p < 0.01 compared with the control.
D-Ribose produced inhibition of AI-2 activity and biofilm formation of L-ZS9, which was induced by AI-2. Thus, the effect of AI-2 on gene expression was investigated. As shown in Figure 8, exogenous AI-2 changed the transcription of tuf, fba, gap, pgm, nfo, rib, and rpoN in L. paraplantarum L-ZS9. Opposite to the effect of D-ribose, AI-2 increased transcription of tuf, pgm, fab, gap and rpoN to about 30-fold, 20-fold, 11-fold, 10-fold, and 378-fold, respectively. And AI-2 decreased the transcription of gene nfo and rib to about 0.21-fold and 0.5-fold, respectively. These results showed that AI-2 and D-ribose had opposite effects on the expression of tuf, fba, gap, pgm, nfo, rib, and rpoN.
FIGURE 8.
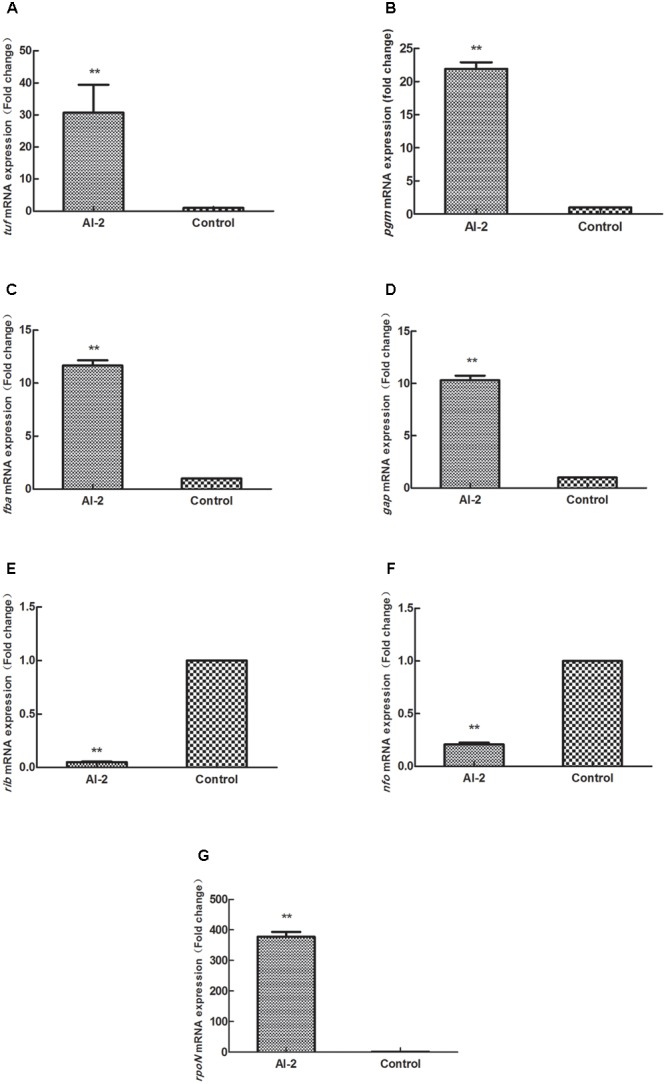
Effects of AI-2 (18.5 μM) on mRNA expression of (A–G) tuf, fba, gap, pgm, nfo, rib, and rpoN genes in L. paraplantarum L-ZS9. The mRNA expression of these genes in the absence of AI-2 served as a control. Data are presented as mean ± SEM. n ≥ 3. ∗p < 0.05, ∗∗p < 0.01 compared with the control.
tuf, fba, gap, pgm, nfo, rib, and rpoN Regulated Biofilm Formation of L. paraplantarum L-ZS9
To validate the effects of tuf, fba, gap, pgm, nfo, rib, and rpoN gene expression on biofilm formation of L. paraplantarum L-ZS9, recombinant strains over-expressing these genes were constructed and their biofilm formation abilities were compared with those of the wild-type strain. Over-expression of these genes had no significant effect on the growth of L. paraplantarum L-ZS9 (Figure 9A). As shown in Figure 9B, over-expression of tuf, pgm, fba, gap, and rpoN increased biofilm formation and over-expression of nfo and rib inhibited biofilm formation of L. paraplantarum L-ZS9 significantly.
FIGURE 9.
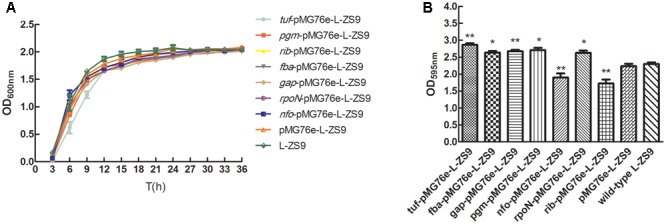
Growth curves (A) and biofilm formation (B) of recombinant L. paraplantarum L-ZS9 strains. The wild-type strain L. paraplantarum L-ZS9 served as a control. Data are presented as mean ± SEM. n ≥ 3. ∗p < 0.05, ∗∗p < 0.01 compared with the control.
Discussion
The LuxS/AI-2 QS system has been found in a variety of bacteria, including both Gram-negative and Gram-positive species. The non-species-specific signal AI-2, which acts as a universal QS signal for interaction between bacterial species, is a byproduct of SAM metabolism with LuxS as the key enzyme. Biosynthesis of AI-2 involves a three-step reaction, which is part of the methionine catabolism cycle. The first step is the removal of a methyl group from SAM, which is catalyzed by SAM-dependent methyltransferases. The resulting product, SAH, is converted to S-ribosylhomocysteine by the enzyme Pfs (Parveen and Cornell, 2011). S-ribosylhomocysteine is hydrolyzed to 4,5-dihydroxy-2,3-pentanedione by LuxS (Schauder et al., 2001). 4,5-Dihydroxy-2,3-pentanedione is further auto-hydrolyzed to form AI-2. SAH exists widely in organisms and is transformed to homocysteine through one-step or two-step conversion. In most bacteria, SAH is metabolized using two-step conversion by the enzymes Pfs and LuxS with concomitant AI-2 production. However, in some prokaryotes and eukaryotes, SAH can be metabolized by one-step conversion using SAH hydrolase (SahH) with no AI-2 production. In this study, genome sequence analysis suggested that L-ZS9 degrades SAH via the enzymes Pfs (MTAN) and LuxS to produce AI-2, without SAH hydrolase (SahH) (Figure 1). In addition, AI-2 activity in CFs of L-ZS9 was determined. AI-2 activity is usually detected by using the reporter strain V. harveyi BB170 (sensor1- sensor2+), which only recognizes AI-2 (Vilchez et al., 2007). However, a high concentration of glucose and acidic pH can inhibit AI-2 activity (DeKeersmaecker and Vanderleyden, 2003; Turovskiy and Chikindas, 2006; Cuadra et al., 2016). Therefore, as for lactic acid bacteria, AI-2 activity detection is difficult under traditional cultural conditions and CF preparation. As a consequence, there was no AI-2 activity detected in L. paraplantarum L-ZS9 when the strain was cultured in MRS broth (data not shown). However, AI-2 activity was detected when L. paraplantarum L-ZS9 was cultured in skim milk medium and CFs were adjusted to pH 7.0 (Figure 2). This preparation method can also be used for AI-2 activity detection of other lactic acid bacteria. According to these results, L. paraplantarum L-ZS9 may have the capacity to communicate with itself and other strains by AI-2. Therefore, LuxS/AI-2 QSI could play an important role in interfering with the social behavior of L-ZS9.
Ribose has been reported to be used as a QSI of the LuxS/AI-2 QS system by competing for AI-2 receptor (James et al., 2006; Armbruster et al., 2011). Our data showed that D-ribose could inhibit AI-2 activity of L-ZS9, suggesting that it can be used as a QSI of the LuxS/AI-2 QS system in L-ZS9. LuxS/AI-2 QS regulates biofilm formation of many bacteria such as Streptococcus mitis, Pseudomonas aeruginosa PAO1, Eikenella corrodens, Candida albicans, Riemerella anatipestifer, Aggregatibacter actinomycetemcomitans, and Bifidobacteria (Shao et al., 2007; Karim et al., 2013; Bachtiar et al., 2014; Christiaen et al., 2014; Han et al., 2015; Li H. et al., 2015; Wang et al., 2016). However, limited studies have been conducted on the regulation of biofilm formation in Lactobacillus by the AI-2/LuxS QS system. It has been reported that luxS of Lactobacillus rhamnosus GG plays a central metabolic role in biofilm formation (Lebeer et al., 2007a). Lactobacillus uses the AI-2 signal to respond to environment stress and to regulate growth and metabolism (Lebeer et al., 2007b, 2008; Moslehi-Jenabian et al., 2009; Yeo et al., 2015). The present study showed that AI-2 increased biofilm formation of L. paraplantarum L-ZS9 (Figure 5A). D-Ribose has been reported to inhibit co-culture biofilm formation of P. aeruginosa PAO1 and Streptococcus mitis significantly (Wang et al., 2016). Ribose could also inhibit streptococcal biofilm formation (Lee et al., 2015). In this study, D-ribose was confirmed to significantly inhibit the biofilm formation of L. paraplantarum L-ZS9 as well (Figure 5B). These results suggest that a universal QSI such as ribose could inhibit biofilm formation of non-pathogenic and beneficial microorganisms, which must be taken into consideration when a universal QSI is used for against pathogens.
To investigate the inhibition mechanism of D-ribose in regulating biofilm formation, relevant genes were firstly identified by comparative proteomic analysis. According to the proteomic analysis and previous research (Wolfe et al., 2004; Da et al., 2008; Kiedrowski et al., 2011; Iyer and Hancock, 2012; Sahu et al., 2012; Zhou et al., 2012; Hao et al., 2013; Li H. et al., 2015; Rajendran et al., 2015), D-ribose may function through gene regulation as shown in Figure 10. The influence of D-ribose and AI-2 on gene transcription (tuf, fba, gap, pgm, nfo, rib, and rpoN) was then further evaluated by qRT-PCR. Our data showed that D-ribose, a LuxS/AI-2 QS system QSI, regulated the expression of tuf, fba, gap, pgm, nfo, rib, and rpoN in an opposite fashion to AI-2. Biofilm assays of recombinant strains further indicated that the overexpression of fba, gap, and pgm could increase the biofilm formation of L-ZS9. The fba, gap, and pgm genes play key roles in the glycolytic pathway (EMP), which is important for energy metabolism. fba is responsible for transformation of frucose-1,6-diphosphate to dihydroxy acetone phosphate, which is transformed to 1,3-diphosphoglycerate by gap, and pgm transforms 3-phosphoglyceride to 2-phosphoglycerate. EMP has been reported to play key roles in biofilm formation and procession of Saccharomyces cerevisiae (Li H. et al., 2015). Proteins involved in EMP were indicated to be up-regulated in biofilm cells of Listeria monocytogenes compared with planktonic cells (Zhou et al., 2012). Our data also showed that EMP in L-ZS9 was positively correlated with biofilm formation. In addition, EMP may play roles in sugar metabolism as well. Previous studies elucidated that extracellular polysaccharide (EPS) is important in biofilm formation (Blackledge et al., 2013). Stoodley et al. (2002) reported the possible role of QS as a signal transduction system to initiate the production of alginate and possibly other types of EPS in P. aeruginosa. Based on these studies, it is logical to deduce that AI-2 and D-ribose could affect biofilm formation of L. paraplantarum L-ZS9 by regulating EPS synthesis.
FIGURE 10.
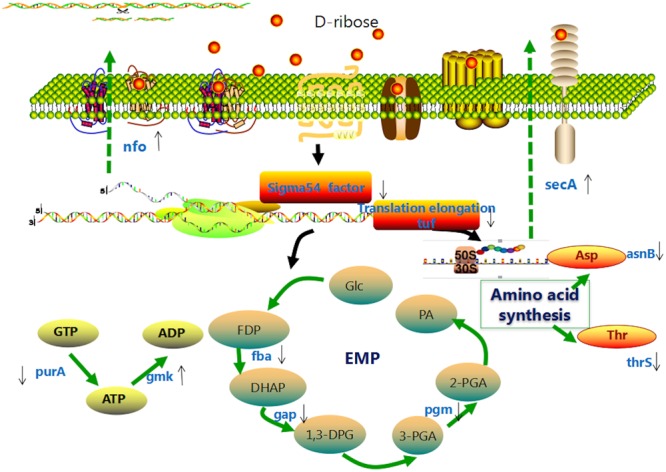
Hypothetical model of the effect of D-ribose on L. paraplantarum L-ZS9 cells. Glc, glucose; FDP, fructose-1,6-diphosphate; DHAP, dihydroxyacetone phosphate; 1,3-DPG, 1,3-diphosphoglyceric acid; 3-PGA, 3-phosphoglyceric acid; 2-PGA, 2-phosphoglyceric acid; PA, pyruvic acid; EMP, glycolysis.
D-Ribose and AI-2 increased and decreased the expression of the nfo gene, respectively, which encodes an endonuclease. Extracellular DNA is one of the important components of biofilm matrix, which could be degraded by various nucleases. DNase is able to break biofilm formation of Gram-positive and Gram-negative bacteria rapidly (Nijland et al., 2010). It was reported that comEB regulated biofilm formation of Staphylococcus lugdunensis by influencing production of DNA (Rajendran et al., 2015). As for Acinetobacter baumannii AIIMS 7, extracellular DNA was degraded and biofilm was decreased to 59.41% after treatment with DNase I (Sahu et al., 2012). In Staphylococcus aureus and Neisseria gonorrhoeae, nuclease modulated biofilm formation by regulating extracellular DNA (Kiedrowski et al., 2011; Steichen et al., 2011). Expression of extracellular endonucleases also influenced the biofilm formation of Shewanella oneidensis MR-1 (Godeke et al., 2011), and DNase 1L2 suppressed biofilm formation of P. aeruginosa and Staphylococcus aureus (Eckhart et al., 2007). Similar to other research, over-expression of endonuclease also inhibited biofilm formation of L-ZS9 in our study.
Our data also showed that the biofilm formation of strain tuf-76e-L-ZS9 was increased compared with the wild-type strain. The tuf gene encodes elongation factor thermo unstable (EF-Tu) and functions in translation elongation, protein folding, and protection from stress (Caldas et al., 1998). Although no research has reported that EF-Tu modulates biofilm formation directly, it has been identified as an adherence-related factor that may affect biofilm formation. EF-Tu plays important roles in adherence of Lactobacillus, such as Lactobacillus acidophilus ATCC 4356, Lactobacillus plantarum 423, and Lactobacillus johnsonii NCC533 (La1) (Ramiah et al., 2008, 2009; Dhanani and Bagchi, 2013). This is the first time to our knowledge that EF-Tu has been reported to modulate biofilm formation directly, which may be due to regulation of the initial adherence of L-ZS9. Over-expression of rpoN that encodes σ54 factor also promoted the biofilm formation of L-ZS9. The σ54 factor is structurally and functionally distinct from all other σ factors and is required for initiation of transcription (Shingler, 2011). In addition, σ54 regulates utilization of alternative carbon sources, detoxification systems, assembly of motility organs, and production of extracellular alginate (Maxson and Darwin, 2006). Although the role of σ54 remains unclear in Lactobacillus, it has been reported that σ54 correlates with biofilm formation directly or indirectly in other bacteria (Belik et al., 2008; Saldias et al., 2008; Iyer and Hancock, 2012; Hao et al., 2013). For example, σ54 controls motility, biofilm formation, and colonization in Vibrio fischeri (Wolfe et al., 2004; Yip et al., 2005) and regulates genes involved in type I and type IV pili biogenesis to influence the biofilm formation of Xylella fastidiosa (Da et al., 2008). Based on the results of these studies, σ54 may play similar roles to regulate biofilm formation of L-ZS9. In addition, it was identified that rib also regulated biofilm formation of L-ZS9, and the underlying mechanism still needs to be further investigated. By exploring the effects of D-ribose on L-ZS9, this study identified the importance of serious consideration of LuxS/AI-2 QSI application due to its effects on non-pathogenic or beneficial bacteria. Modification of specific genes (for example genes identified in this research) should be conducted in order to protect these organisms from the QSI.
Conclusion
Lactobacillus paraplantarum L-ZS9 contains key genes related to the LuxS/AI-2 QS system and is able to produce AI-2. D-Ribose can be used as a QSI of L-ZS9 and inhibits biofilm formation of L-ZS9 in contrast to exogenous AI-2 by regulating expression of the tuf, fba, gap, pgm, nfo, rib, and rpoN genes. This research provides new information about regulation mechanisms of the LuxS/AI-2 QS system in Lactobacillus and will be useful for determining the reasonable application of QSI.
Author Contributions
LL and PL designed the experiments. LL, RW, and JZ performed the experiments. LL, NS, and PL analyzed the results and wrote the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Funding. This work was supported by grants from the National Natural Science Foundation of China (31671831 and 31471707).
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2017.01860/full#supplementary-material
References
- Armbruster C. E., Pang B., Murrah K., Juneau R. A., Perez A. C., Weimer K. E., et al. (2011). RbsB (NTHI_0632) mediates quorum signal uptake in nontypeable Haemophilus influenzae strain 86-028NP. Mol. Microbiol. 82 836–850. 10.1111/j.1365-2958.2011.07831.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bachtiar E. W., Bachtiar B. M., Jarosz L. M., Amir L. R., Sunarto H., Ganin H., et al. (2014). AI-2 of Aggregatibacter actinomycetemcomitans inhibits Candida albicans biofilm formation. Front. Cell. Infect. Microbiol. 4:94 10.3389/fcimb.2014.00094 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bassler B. L., Wright M., Showalter R. E., Silverman M. R. (1993). Intercellular signalling in Vibrio harveyi: sequence and function of genes regulating expression of luminescence. Mol. Microbiol. 9 773–786. 10.1111/j.1365-2958.1993.tb01737.x [DOI] [PubMed] [Google Scholar]
- Belik A. S., Tarasova N. N., Khmel’ I. A. (2008). Regulation of biofilm formation in Escherichia coli K12: effect of mutations in HNS, StpA, lon, and rpoN genes. Mol. Gen. Mikrobiol. Virusol. 23 159–162. 10.3103/S0891416808040010 [DOI] [PubMed] [Google Scholar]
- Bixler G. D., Bhushan B. (2012). Biofouling: lessons from nature. Philos. Trans. A Math. Phys. Eng. Sci. 370 2381–2417. 10.1098/rsta.2011.0502 [DOI] [PubMed] [Google Scholar]
- Blackledge M. S., Worthington R. J., Melander C. (2013). Biologically inspired strategies for combating bacterial biofilms. Curr. Opin. Pharmacol. 13 699–706. 10.1016/j.coph.2013.07.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brackman G., Celen S., Baruah K., Bossier P., Van Calenbergh S., Nelis H. J., et al. (2009). AI-2 quorum-sensing inhibitors affect the starvation response and reduce virulence in several Vibrio species, most likely by inferring with LuxPQ. Microbiology 155 4114–4122. 10.1099/mic.0.032474-0 [DOI] [PubMed] [Google Scholar]
- Brackman G., Celen S., Hillaert U., Van Calenbergh S., Cos P., Maes L., et al. (2011). Structure-activity relationship of cinnamaldehyde analogs as inhibitors of AI-2 based quorum sensing and their effect on virulence of Vibrio spp. PLOS ONE 6:e16084 10.1371/journal.pone.0016084 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buck B. L., Azcarate-Peril M. A., Klaenhammer T. R. (2009). Role of autoinducer-2 on the adhesion ability of Lactobacillus acidophilus. J. Appl. Microbiol. 107 269–279. 10.1111/j.1365-2672.2009.04204.x [DOI] [PubMed] [Google Scholar]
- Caldas T. D., El Y. A., Richarme G. (1998). Chaperone properties of bacterial elongation factor EF-Tu. J. Biol. Chem. 273 11478–11482. 10.1074/jbc.273.19.11478 [DOI] [PubMed] [Google Scholar]
- Cao J. G., Meighen E. A. (1989). Purification and structural identification of an autoinducer for the luminescence system of Vibrio harveyi. J. Biol. Chem. 264 21670–21676. [PubMed] [Google Scholar]
- Cho Y. J., Song H. Y., Ben A. H., Choi B. K., Eunju R., Cho Y. A., et al. (2016). In vivo inhibition of Porphyromonas gingivalis growth and prevention of periodontitis with quorum-sensing inhibitors. J. Periodontol. 87 1075–1082. 10.1902/jop.2016.160070 [DOI] [PubMed] [Google Scholar]
- Christiaen S. E., O’Connell M. M., Bottacini F., Lanigan N., Casey P. G., Huys G., et al. (2014). Autoinducer-2 plays a crucial role in gut colonization and probiotic functionality of Bifidobacterium breve UCC2003. PLOS ONE 9:e98111 10.1371/journal.pone.0098111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cuadra G. A., Frantellizzi A. J., Gaesser K. M., Tammariello S. P., Ahmed A. (2016). Autoinducer-2 detection among commensal oral streptococci is dependent on pH and boric acid. J. Microbiol. 54 492–502. 10.1007/s12275-016-5507-z [DOI] [PubMed] [Google Scholar]
- Da S. N. J., Koide T., Abe C. M., Gomes S. L., Marques M. V. (2008). Role of sigma54 in the regulation of genes involved in type I and type IV pili biogenesis in Xylella fastidiosa. Arch. Microbiol. 189 249–261. 10.1007/s00203-007-0314-x [DOI] [PubMed] [Google Scholar]
- DeKeersmaecker S. C., Vanderleyden J. (2003). Constraints on detection of autoinducer-2 (AI-2) signalling molecules using Vibrio harveyi as a reporter. Microbiology 149 1953–1956. 10.1099/mic.0.C0117-0 [DOI] [PubMed] [Google Scholar]
- Dhanani A. S., Bagchi T. (2013). The expression of adhesin EF-Tu in response to mucin and its role in Lactobacillus adhesion and competitive inhibition of enteropathogens to mucin. J. Appl. Microbiol. 115 546–554. 10.1111/jam.12249 [DOI] [PubMed] [Google Scholar]
- Drescher K., Shen Y., Bassler B. L., Stone H. A. (2013). Biofilm streamers cause catastrophic disruption of flow with consequences for environmental and medical systems. Proc. Natl. Acad. Sci. U.S.A. 110 4345–4350. 10.1073/pnas.1300321110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eckhart L., Fischer H., Barken K. B., Tolker-Nielsen T., Tschachler E. (2007). DNase1L2 suppresses biofilm formation by Pseudomonas aeruginosa and Staphylococcus aureus. Br. J. Dermatol. 156 1342–1345. 10.1111/j.1365-2133.2007.07886.x [DOI] [PubMed] [Google Scholar]
- Godeke J., Heun M., Bubendorfer S., Paul K., Thormann K. M. (2011). Roles of two Shewanella oneidensis MR-1 extracellular endonucleases. Appl. Environ. Microbiol. 77 5342–5351. 10.1128/AEM.00643-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gospodarek E., Bogiel T., Zalas-Wiecek P. (2009). Communication between microorganisms as a basis for production of virulence factors. Pol. J. Microbiol. 58 191–198. [PubMed] [Google Scholar]
- Han X., Liu L., Fan G., Zhang Y., Xu D., Zuo J., et al. (2015). Riemerella anatipestifer lacks luxS, but can uptake exogenous autoinducer-2 to regulate biofilm formation. Res. Microbiol. 166 486–493. 10.1016/j.resmic.2015.06.004 [DOI] [PubMed] [Google Scholar]
- Hao B., Mo Z. L., Xiao P., Pan H. J., Lan X., Li G. Y. (2013). Role of alternative sigma factor 54 (RpoN) from Vibrio anguillarum M3 in protease secretion, exopolysaccharide production, biofilm formation, and virulence. Appl. Microbiol. Biotechnol. 97 2575–2585. 10.1007/s00253-012-4372-x [DOI] [PubMed] [Google Scholar]
- Harjai K., Gupta R. K., Sehgal H. (2014). Attenuation of quorum sensing controlled virulence of Pseudomonas aeruginosa by cranberry. Indian J. Med. Res. 139 446–453. [PMC free article] [PubMed] [Google Scholar]
- Hoiby N., Bjarnsholt T., Givskov M., Molin S., Ciofu O. (2010). Antibiotic resistance of bacterial biofilms. Int. J. Antimicrob. Agents 35 322–332. 10.1016/j.ijantimicag.2009.12.011 [DOI] [PubMed] [Google Scholar]
- Iyer V. S., Hancock L. E. (2012). Deletion of sigma(54) (rpoN) alters the rate of autolysis and biofilm formation in Enterococcus faecalis. J. Bacteriol. 194 368–375. 10.1128/JB.06046-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- James D., Shao H., Lamont R. J., Demuth D. R. (2006). The Actinobacillus actinomycetemcomitans ribose binding protein RbsB interacts with cognate and heterologous autoinducer 2 signals. Infect. Immun. 74 4021–4029. 10.1128/IAI.01741-05 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karim M. M., Hisamoto T., Matsunaga T., Asahi Y., Noiri Y., Ebisu S., et al. (2013). LuxS affects biofilm maturation and detachment of the periodontopathogenic bacterium Eikenella corrodens. J. Biosci. Bioeng. 116 313–318. 10.1016/j.jbiosc.2013.03.013 [DOI] [PubMed] [Google Scholar]
- Kiedrowski M. R., Kavanaugh J. S., Malone C. L., Mootz J. M., Voyich J. M., Smeltzer M. S., et al. (2011). Nuclease modulates biofilm formation in community-associated methicillin-resistant Staphylococcus aureus. PLOS ONE 6:e26714 10.1371/journal.pone.0026714 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lakshman D. K., Natarajan S. S., Lakshman S., Garrett W. M., Dhar A. K. (2008). Optimized protein extraction methods for proteomic analysis of Rhizoctonia solani. Mycologia 100 867–875. 10.3852/08-065 [DOI] [PubMed] [Google Scholar]
- Lebeer S., Claes I. J., Verhoeven T. L., Shen C., Lambrichts I., Ceuppens J. L., et al. (2008). Impact of luxS and suppressor mutations on the gastrointestinal transit of Lactobacillus rhamnosus GG. Appl. Environ. Microbiol. 74 4711–4718. 10.1128/AEM.00133-08 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lebeer S., De Keersmaecker S. C., Verhoeven T. L., Fadda A. A., Marchal K., Vanderleyden J. (2007a). Functional analysis of luxS in the probiotic strain Lactobacillus rhamnosus GG reveals a central metabolic role important for growth and biofilm formation. J. Bacteriol. 189 860–871. 10.1128/JB.01394-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lebeer S., Verhoeven T. L., Perea V. M., Vanderleyden J., De Keersmaecker S. C. (2007b). Impact of environmental and genetic factors on biofilm formation by the probiotic strain Lactobacillus rhamnosus GG. Appl. Environ. Microbiol. 73 6768–6775. 10.1128/AEM.01393-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee H. J., Kim S. C., Kim J., Do A., Han S. Y., Lee B. D., et al. (2015). Synergistic inhibition of streptococcal biofilm by ribose and xylitol. Arch. Oral Biol. 60 304–312. 10.1016/j.archoralbio.2014.11.004 [DOI] [PubMed] [Google Scholar]
- Lepargneur J. P., Rousseau V. (2002). Protective role of the Doderlein flora. J. Gynecol. Obstet. Biol. Reprod. 31 485–494. [PubMed] [Google Scholar]
- Li H., Li X., Wang Z., Fu Y., Ai Q., Dong Y., et al. (2015). Autoinducer-2 regulates Pseudomonas aeruginosa PAO1 biofilm formation and virulence production in a dose-dependent manner. BMC Microbiol. 15:192 10.1186/s12866-015-0529-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li Z., Chen Y., Liu D., Zhao N., Cheng H., Ren H., et al. (2015). Involvement of glycolysis/gluconeogenesis and signaling regulatory pathways in Saccharomyces cerevisiae biofilms during fermentation. Front. Microbiol. 6:139 10.3389/fmicb.2015.00139 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu L., Li P. (2016). Complete genome sequence of Lactobacillus paraplantarum L-ZS9, a probiotic starter producing class II bacteriocins. J. Biotechnol. 222 15–16. 10.1016/j.jbiotec.2016.02.003 [DOI] [PubMed] [Google Scholar]
- Livak K. J., Schmittgen T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 25 402–408. 10.1006/meth.2001.1262 [DOI] [PubMed] [Google Scholar]
- Maxson M. E., Darwin A. J. (2006). Multiple promoters control expression of the Yersinia enterocolitica phage-shock-protein a (PspA) operon. Microbiology 152 1001–1010. 10.1099/mic.0.28714-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moslehi-Jenabian S., Gori K., Jespersen L. (2009). AI-2 signalling is induced by acidic shock in probiotic strains of Lactobacillus spp. Int. J. Food Microbiol. 135 295–302. 10.1016/j.ijfoodmicro.2009.08.011 [DOI] [PubMed] [Google Scholar]
- Nadell C. D., Drescher K., Foster K. R. (2016). Spatial structure, cooperation and competition in biofilms. Nat. Rev. Microbiol. 14 589–600. 10.1038/nrmicro.2016.84 [DOI] [PubMed] [Google Scholar]
- Nijland R., Hall M. J., Burgess J. G. (2010). Dispersal of biofilms by secreted, matrix degrading, bacterial DNase. PLOS ONE 5:e15668 10.1371/journal.pone.0015668 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Olsen I. (2015). Biofilm-specific antibiotic tolerance and resistance. Eur. J. Clin. Microbiol. Infect. Dis. 34 877–886. 10.1007/s10096-015-2323-z [DOI] [PubMed] [Google Scholar]
- Pan J., Ren D. (2009). Quorum sensing inhibitors: a patent overview. Expert Opin. Ther. Pat. 19 1581–1601. 10.1517/13543770903222293 [DOI] [PubMed] [Google Scholar]
- Park H., Shin H., Lee K., Holzapfel W. (2016). Autoinducer-2 properties of kimchi are associated with lactic acid bacteria involved in its fermentation. Int. J. Food Microbiol. 225 38–42. 10.1016/j.ijfoodmicro.2016.03.007 [DOI] [PubMed] [Google Scholar]
- Parveen N., Cornell K. A. (2011). Methylthioadenosine/S-adenosylhomocysteine nucleosidase, a critical enzyme for bacterial metabolism. Mol. Microbiol. 79 7–20. 10.1111/j.1365-2958.2010.07455.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Penesyan A., Gillings M., Paulsen I. T. (2015). Antibiotic discovery: combatting bacterial resistance in cells and in biofilm communities. Molecules 20 5286–5298. 10.3390/molecules20045286 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rajendran N. B., Eikmeier J., Becker K., Hussain M., Peters G., Heilmann C. (2015). Important contribution of the novel locus comEB to extracellular DNA-dependent Staphylococcus lugdunensis biofilm formation. Infect. Immun. 83 4682–4692. 10.1128/IAI.00775-15 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramiah K., van Reenen C. A., Dicks L. M. (2008). Surface-bound proteins of Lactobacillus plantarum 423 that contribute to adhesion of Caco-2 cells and their role in competitive exclusion and displacement of Clostridium sporogenes and Enterococcus faecalis. Res. Microbiol. 159 470–475. 10.1016/j.resmic.2008.06.002 [DOI] [PubMed] [Google Scholar]
- Ramiah K., van Reenen C. A., Dicks L. M. (2009). Expression of the mucus adhesion gene mub, surface layer protein slp and adhesion-like factor EF-TU of Lactobacillus acidophilus ATCC 4356 under digestive stress conditions, as monitored with real-time PCR. Probiotics Antimicrob. Proteins 1 91 10.1007/s12602-009-9009-8 [DOI] [PubMed] [Google Scholar]
- Roy V., Meyer M. T., Smith J. A., Gamby S., Sintim H. O., Ghodssi R., et al. (2013). AI-2 analogs and antibiotics: a synergistic approach to reduce bacterial biofilms. Appl. Microbiol. Biotechnol. 97 2627–2638. 10.1007/s00253-012-4404-6 [DOI] [PubMed] [Google Scholar]
- Ryu E. J., Sim J., Sim J., Lee J., Choi B. K. (2016). D-Galactose as an autoinducer 2 inhibitor to control the biofilm formation of periodontopathogens. J. Microbiol. 54 632–637. 10.1007/s12275-016-6345-8 [DOI] [PubMed] [Google Scholar]
- Sahu P. K., Iyer P. S., Oak A. M., Pardesi K. R., Chopade B. A. (2012). Characterization of eDNA from the clinical strain Acinetobacter baumannii AIIMS 7 and its role in biofilm formation. ScientificWorldJournal 2012:973436 10.1100/2012/973436 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saldias M. S., Lamothe J., Wu R., Valvano M. A. (2008). Burkholderia cenocepacia requires the RpoN sigma factor for biofilm formation and intracellular trafficking within macrophages. Infect. Immun. 76 1059–1067. 10.1128/IAI.01167-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schauder S., Shokat K., Surette M. G., Bassler B. L. (2001). The LuxS family of bacterial autoinducers: biosynthesis of a novel quorum-sensing signal molecule. Mol. Microbiol. 41 463–476. 10.1046/j.1365-2958.2001.02532.x [DOI] [PubMed] [Google Scholar]
- Shao H., Lamont R. J., Demuth D. R. (2007). Autoinducer 2 is required for biofilm growth of Aggregatibacter (Actinobacillus) actinomycetemcomitans. Infect. Immun. 75 4211–4218. 10.1128/IAI.00402-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shingler V. (2011). Signal sensory systems that impact sigma54-dependent transcription. FEMS Microbiol. Rev. 35 425–440. 10.1111/j.1574-6976.2010.00255.x [DOI] [PubMed] [Google Scholar]
- Steichen C. T., Cho C., Shao J. Q., Apicella M. A. (2011). The Neisseria gonorrhoeae biofilm matrix contains DNA, and an endogenous nuclease controls its incorporation. Infect. Immun. 79 1504–1511. 10.1128/IAI.01162-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stoodley P., Sauer K., Davies D. G., Costerton J. W. (2002). Biofilms as complex differentiated communities. Annu. Rev. Microbiol. 56 187–209. 10.1146/annurev.micro.56.012302.160705 [DOI] [PubMed] [Google Scholar]
- Sun Z., He X., Brancaccio V. F., Yuan J., Riedel C. U. (2014). Bifidobacteria exhibit LuxS-dependent autoinducer 2 activity and biofilm formation. PLOS ONE 9:e88260 10.1371/journal.pone.0088260 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taylor P. K., Yeung A. T., Hancock R. E. (2014). Antibiotic resistance in Pseudomonas aeruginosa biofilms: towards the development of novel anti-biofilm therapies. J. Biotechnol. 191 121–130. 10.1016/j.jbiotec.2014.09.003 [DOI] [PubMed] [Google Scholar]
- Thompson J. A., Oliveira R. A., Djukovic A., Ubeda C., Xavier K. B. (2015). Manipulation of the quorum sensing signal AI-2 affects the antibiotic-treated gut microbiota. Cell Rep. 10 1861–1871. 10.1016/j.celrep.2015.02.049 [DOI] [PubMed] [Google Scholar]
- Turovskiy Y., Chikindas M. L. (2006). Autoinducer-2 bioassay is a qualitative, not quantitative method influenced by glucose. J. Microbiol. Methods 66 497–503. 10.1016/j.mimet.2006.02.001 [DOI] [PubMed] [Google Scholar]
- Vilchez R., Lemme A., Thiel V., Schulz S., Sztajer H., Wagner-Dobler I. (2007). Analysing traces of autoinducer-2 requires standardization of the Vibrio harveyi bioassay. Anal. Bioanal. Chem. 387 489–496. 10.1007/s00216-006-0824-4 [DOI] [PubMed] [Google Scholar]
- Wang Y., Cheung Y. H., Yang Z., Chiu J. F., Che C. M., He Q. Y. (2006). Proteomic approach to study the cytotoxicity of dioscin (saponin). Proteomics 6 2422–2432. 10.1002/pmic.200500595 [DOI] [PubMed] [Google Scholar]
- Wang Y., Sun Y., Zhang X., Zhang Z. C., Song J. Y., Gui M., et al. (2015). Bacteriocin-producing probiotics enhance the safety and functionality of sturgeon sausage. Food Control 50 729–735. 10.1016/j.foodcont.2014.09.045 [DOI] [Google Scholar]
- Wang Z., Xiang Q., Yang T., Li L., Yang J., Li H., et al. (2016). Autoinducer-2 of Streptococcus mitis as a target molecule to inhibit pathogenic multi-species biofilm formation in vitro and in an endotracheal intubation rat model. Front. Microbiol. 7:88 10.3389/fmicb.2016.00088 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wolfe A. J., Millikan D. S., Campbell J. M., Visick K. L. (2004). Vibrio fischeri sigma54 controls motility, biofilm formation, luminescence, and colonization. Appl. Environ. Microbiol. 70 2520–2524. 10.1128/AEM.70.4.2520-2524.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yeo S., Park H., Ji Y., Park S., Yang J., Lee J., et al. (2015). Influence of gastrointestinal stress on autoinducer-2 activity of two Lactobacillus species. FEMS Microbiol. Ecol. 91:fiv065 10.1093/femsec/fiv065 [DOI] [PubMed] [Google Scholar]
- Yip E. S., Grublesky B. T., Hussa E. A., Visick K. L. (2005). A novel, conserved cluster of genes promotes symbiotic colonization and sigma-dependent biofilm formation by Vibrio fischeri. Mol. Microbiol. 57 1485–1498. 10.1111/j.1365-2958.2005.04784.x [DOI] [PubMed] [Google Scholar]
- Zhang X., Wang Y., Liu L., Wei Y., Shang N., Zhang X., et al. (2016). Two-peptide bacteriocin PlnEF causes cell membrane damage to Lactobacillus plantarum. Biochim. Biophys. Acta 1858 274–280. 10.1016/j.bbamem.2015.11.018 [DOI] [PubMed] [Google Scholar]
- Zhou Q., Feng X., Zhang Q., Feng F., Yin X., Shang J., et al. (2012). Carbon catabolite control is important for Listeria monocytogenes biofilm formation in response to nutrient availability. Curr. Microbiol. 65 35–43. 10.1007/s00284-012-0125-4 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.