

Abstract
Biatractylolide, isolated from the ethyl acetate extract of Atractylodes macrocephala, has shown various pharmacological activities such as antitumor and antioxidant activities. In this work, we aim to study the protective effect of biatractylolide on glutamate-induced rat adrenal pheochromocytoma cell (PC12) and human bone marrow neuroblastoma cell line (SH-SY5Y) injury and preliminarily explore its mechanism. The results showed that glutamate was cytotoxic with an inhibitory concentration 50% (IC50) of 8.5 mM in PC12 and 10 mM in SH-SY5Y cells. In this work, the preincubation with biatractylolide (10, 15, and 20 μM) observably improved cell viability, inhibited the apoptosis of cells induced by glutamate, and reduced the activity of LDH. AO staining revealed that apoptosis of cells was decreased. Additionally, the results of western blotting manifested that pretreatment with biatractylolide could downregulate GSK3β protein expression and upregulate p-Akt protein expression, thereby protecting PC12 and SH-SY5Y cells from injury. All these findings indicate that biatractylolide has a neuroprotective effect on glutamate-induced injury in PC12 and SH-SY5Y cells through a mechanism of the PI3K-Akt-GSK3β-dependent pathways.
1. Introduction
Alzheimer's disease (AD) is a progressive neurodegenerative disease, used to describe a wide range of conditions, and its appearance was related to advanced age [1]. The main clinical symptoms included memory loss and cognitive impairment. At present, common clinical drugs in treatment of AD are most AChE inhibitors [2] and N-methyl-d-aspartate (NMDA) receptor antagonist [3]. Cholinesterase inhibitors can alleviate cholinergic deficit and improve neurotransmission by blocking the degradation of Ach [4, 5] and are therefore considered to be an effective and efficient way for the treatment and prevention of AD.
PC12 and SH-SY5Y cell, neural cell lines, has become an ideal cell model for exploring the mechanism of nerve cell injury in vitro and the protective effect of drugs on neuronal cells because of its typical characteristics of neuroendocrine cells [6, 7]. A large number of studies indicate [8–10] that abnormal aggregation of glutamate in the cells can lead to degeneration of the nervous system, which can cause nerve excitability toxicity. Several studies have demonstrated that the main reason for the loss of neurons in the central nervous system is due to the overexcitation of glutamate receptors and high concentration of glutamate induces PC12 and SH-SY5Y cell death [11–13]. Therefore, glutamate-induced PC12 and SH-SY5Y cells have been chosen to establish nerve injury model in our study.
The roots of Atractylodis Macrocephalae Rhizoma have been widely used in traditional Chinese medicine due to its various pharmacological activities including antioxidation [14], gastroprotective [15], antitumor [16], anti-AD [17] effects. Biatractylolide is an active ingredient existing in Atractylodis Macrocephalae Rhizoma and its small molecule is double sesquiterpene ester which has a novel symmetrical structure. Our previous study confirmed that biatractylolide has a significant effect on reducing the activity of AChE in the brain and improving the memory ability of mouse dementia induced by aluminum trichloride [18]; moreover, biatractylolide can significantly reduce cholinesterase activity in model rats and improve the behavior and memory of AD model rats induced by Aβ1−40 [19]. Importantly, biatractylolide can inhibit the enzyme activity of AChE and downregulate the expression of AChE in a dose-dependent manner, which has been confirmed in our previous study [20]. However, the specific mechanisms of biatractylolide are not well understood, and there are no reports about protective effect of biatractylolide currently. Therefore, in our study, biatractylolide was applied to explore its protective effects against the glutamate-induced cell damage in PC12 and SH-SY5Y cells and mechanisms, providing a theoretical basis for its great potential application on the treatment of Alzheimer's disease.
2. Materials and Methods
2.1. Reagents
Biatractylolide was isolated from the ethyl acetate extract of Atractylodes macrocephala by multistep chromatographic processing [20] (Figure 1). Glutamate and DMSO were purchased from Sigma Chemicals (USA). Antibodies for the protein characteristics were against total Akt, phosphor-Akt, total GSK3β, phosphor-GSK3β, and β-actin which were purchased from Cell Signaling Technology (Danvers, MA, USA).
Figure 1.
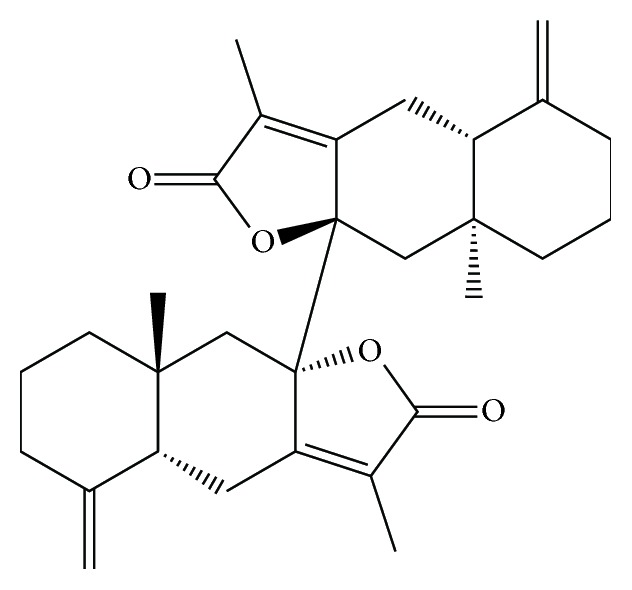
The chemical structure of biatractylolide. Biatractylolide is a kind of internal symmetry double sesquiterpene novel ester compound.
2.2. Cell Culture
PC12 cells and SH-SY5Y cells donated by Dr. James R. Woodgett (China Pharmaceutical University, Nanjing, China) were routinely cultured in DMEM medium in the presence of 10% FBS and 1% double antibody. All cells were cultured at 37°C in a humidified 5% CO2 incubator.
2.3. Experimental Grouping and Drug Treatment
By adding different concentrations of glutamate ranging from 0 to 30 mM in the preliminary experiment, we selected the concentrations (7.5 mM and 10 mM) as the most optimal injured concentrations by determining dose-response curves. The experiment was as follows: (1) blank control group; (2) glutamate injury model group; (3) drug treatment group (low, medium, and high).
2.4. Cell Viability Assay
1 × 104 cells were seeded in 96-well culture plates and cultured overnight. Then the cells were treated with various concentrations of biatractylolide for 30 min before glutamate treatment for 24 hours in incubator. The medium was washed off with PBS and 20 μL MTT Reagent was added to each well and the plate was incubated at 37°C for 4 h. Finally, the medium was removed from each well and 150 μl of DMSO was added to dissolve formazan crystals. The product was quantified by measuring absorbance at 490 nm using a Dynatech MR5000 plate reader.
2.5. Assessment of Apoptosis
6 × 105 PC12 cells and 8 × 105 SH-SY5Y cells were seeded in 6-well plates and cultured overnight. Then the cells were treated with various concentrations of biatractylolide for 30 min before glutamate treatment for 24 hours in incubator. Then treated cells were digested and washed twice with cold PBS. The cell precipitation was resuspended and added to 5 μL FITC Annexin V and 10 μl PI and then incubated for 15 min at room temperature in dark. After adding 300 μL of binding buffer, labeled cells were counted by flow cytometry within 1 h. All early apoptotic cells and necrotic/late apoptotic cells as well as living cells were detected by FACSCalibur flow cytometer and subsequently analyzed by Cell Quest software (Becton Dickinson).
2.6. Acridine Orange/Ethidium Bromide Staining
6 × 105 PC12 cells and 8 × 105 SH-SY5Y cells were seeded in 6-well plates and cultured overnight. Then the cells were treated with various concentrations of biatractylolide for 30 min before glutamate treatment for 24 hours in incubator. The treated cells were washed with cold PBS twice and fixed with formaldehyde for 30 min according to AO/EB staining assay kit instructions. Finally, they were incubated with AO (6 μg/ml) for 10 min at room temperature in the dark and then observed by fluorescence microscopy.
2.7. Determination of LDH Activity
5 × 105 PC12 cells and 6 × 105 SH-SY5Y cells were seeded in 24-well plates and cultured overnight. Then the cells were treated with various concentrations of biatractylolide for 30 min before glutamate treatment for 24 hours in incubator. Twenty-four hours later, the supernatant was collected and centrifuged at 3500 rpm for 3 min and the OD value of LDH in the medium was determined by colorimetric method according to LDH assay kit.
2.8. Measurement of Intracellular ROS
6 × 105 PC12 cells and 8 × 105 SH-SY5Y cells were seeded in 6-well plates and cultured overnight. Then the cells were treated with various concentrations of biatractylolide for 30 min before glutamate treatment for 24 hours in incubator. Next, the treated cells were washed with cold PBS twice and fixed with formaldehyde for 30 min according to ROS assay kit instructions. The intracellular ROS was observed by fluorescence microscopy.
2.9. Detection of Mitochondrial Membrane Potential by Rhodamine-123 Staining
5 × 105 PC12 cells and 6 × 105 SH-SY5Y cells were seeded in 24-well plates and cultured overnight. Then the cells were treated with various concentrations of biatractylolide for 30 min before glutamate treatment for 24 hours in incubator. The treated cells were harvested and resuspended in the medium and, finally, incubated with Rhodamine-123 (10 μg/ml) for 10 min at room temperature in the dark and then observed by fluorescence microscopy.
2.10. Western Blotting
Western blotting assay was performed using methods described by Su et al. [21]. Primary antibody was added in BSA and allowed to incubate overnight at 4°C, washed with TBS/0.05% Tween-20 for 5 times (10 min per time) before the secondary antibody was added and then incubated for an additional hour at room temperature. The membrane was again washed 3 times before adding Pierce Super Signal chemiluminescent substrate (Rockford, IL, USA) and then immediately imaged on ChemiDoc (Bio-Rad, Hercules, CA, USA). The films were scanned using EPSON PERFECTION V500 PHOTO and quantified by Image J (NIH, Bethesda, MD, USA).
2.11. Statistical Analysis
All data are presented as mean ± SEM and GraphPad Prism statistical software was used for analysis. Comparison between the two groups using Student's t-test and the variance analysis was performed with One-Way ANOVA method and statistical significance was assumed at a value of p < 0.05 or less.
3. Results
3.1. The PC12 and SH-SY5Y Cell Proliferation
Initially, we explore the effects of different concentrations of glutamate (0.94, 1.88, 3.75, 7.5, 15, and 30 mM) on the proliferation of SH-SY5Y cells and PC12 cells using MTT test. The result indicated that the inhibition rate of 7.5 mM glutamate-treated PC12 cells reached 48.2 ± 1.5% (p < 0.001) and 15 mM glutamate-treated SH-SY5Y cells led to 39.12 ± 2.1% decrease of cell viability (Figure 2(a)). Thus, glutamate at 7.5 mM and 10 mM was selected as model concentration. To investigate the impact of biatractylolide on cell damage induced by glutamate, we evaluated cell viability using the MTT approach. The cells were treated with various concentrations of biatractylolide for 30 min before glutamate treatment for 24 h. The result showed that biatractylolide led to a dose-dependent increase on PC12 and SH-SY5Y cells proliferation (Figure 2(b)).
Figure 2.

Protective effect of biatractylolide on the viability of PC12 and SH-SY5Y cells insulted by glutamate. (a) Viability change of PC12 and SH-SY5Y cells treated by glutamate under different concentration circumstances after 24 h. (b) Biatractylolide inhibited glutamate-induced injury in PC12 and SH-SY5Y cells. ∗p < 0.05, ∗∗p < 0.01, ∗∗∗p < 0.005, and ∗∗∗∗p < 0.001 versus control group; #p < 0.05 and ##p < 0.01 versus model group.
3.2. Assessment of Apoptosis
Glutamate-induced cell apoptosis was quantitatively tested by using FITC and PI. As shown in Figures 3(a)-3(b), we found that cell apoptosis rate was considerably increased to 162.8 ± 1.7% and 202.5 ± 3.2% in the model group compared to the control group; however, in the treated group, biatractylolide distinctly decreased glutamate-induced cell apoptosis in a dose-dependent fashion (p < 0.05).
Figure 3.

Biatractylolide protected PC12 and SH-SY5Y cells from apoptosis induced by glutamate. (a) Representative flow cytometry scatter plots. (b) Bar chart shows quantitative data of flow cytometry experiments. ∗∗p < 0.01 versus control group; #p < 0.05 and ##p < 0.01 versus model group.
3.3. Acridine Orange/Ethidium Bromide Staining
We observed the morphological characteristic of apoptotic cells using the AO/EB staining test. And Figure 4 displayed that morphological changes including chromatic agglutination, karyopyknosis, and nuclear fragmentation could be observed in glutamate model group by fluorescence microscopy. Compared with the model group, the addition of different concentrations of biatractylolide can markedly inhibit cell damage and improve cell morphology in a concentration-dependent manner.
Figure 4.

Cell morphology of PC12 and SH-SY5Y cells stained with acridine orange (×400). The two cell lines were treated with various concentrations of biatractylolide for 30 min before glutamate treatment for 24 hours. The cell morphology was observed under fluorescence microscope.
3.4. Determination of LDH Activity
We next determined the effects of biatractylolide on LDH activity in PC12 cells and SH-SY5Y cells; we found that the model group has an increase of LDH release, being 280.3 ± 0.3% and 318.22 ± 0.1% as compared to control cells (p < 0.01; Figures 5(a)-5(b)). However, preincubation with biatractylolide at the concentration of 15 μM and 20 μM significantly obstructed LDH release in the PC12 cells, which was decreased from 180.5 ± 0.9% to about 195.3 ± 2.1% as compared to model group (p < 0.01; Figure 5(a)). In addition, biatractylolide (10 μM, 15 μM, and 20 μM) distinctly decreased LDH activity of SH-SY5Y cells, being 174.2 ± 0.4% (p < 0.001), 243.6 ± 0.1% (p < 0.005), and 272.1 ± 2.9% (p < 0.001) as compared to model group (Figure 5(b)).
Figure 5.

Determination of LDH activity. (a)-(b) After the two cell lines were treated with various concentrations of biatractylolide for 30 min before glutamate treatment for 24 hours, the LDH activity was examined by LDH assay kit. ∗∗∗p < 0.01 versus control group; ##p < 0.01, ###p < 0.005, and ####p < 0.001 versus model group.
3.5. Measurement of Intracellular ROS
We also investigated the effects of various concentrations of biatractylolide on the release of ROS in SH-SY5Y cells and PC12 cells. As shown in Figure 6(a), we found that the model group has an increase of relative fluorescence unit in PC12 and SH-SY5Y cells as compared to control cells. However, pretreatment with biatractylolide at the concentration of 20 μM significantly inhibited relative fluorescence intensity of ROS to 112.6 ± 0.7% as compared to model group (p < 0.01) (Figure 6(b)). In addition, biatractylolide (10 μM, 15 μM, and 20 μM) distinctly decreased ROS production of SH-SY5Y cells, being 161.2 ± 1.3% (p < 0.05), 133.6 ± 2.9% (p < 0.01), and 105.9 ± 3.4% (p < 0.01) as compared to model group (Figure 6(c)).
Figure 6.

The glutamate-stimulated ROS was effectively attenuated by biatractylolide in PC12 and SH-SY5Y cell lines. After the two cell lines were treated with various concentrations of biatractylolide for 30 min before glutamate treatment for 24 hours, the ROS production was detected by ROS kit. (a) The fluorescence image. (b)-(c) Bar chart shows quantitative data. ∗p < 0.05 and ∗∗p < 0.01 versus control group; #p < 0.05 and ##p < 0.05 versus model group.
3.6. Detection of Mitochondrial Membrane Potential by Rhodamine-123 Staining
As can be seen in Figure 7, after being treated with 7.5 and 10 mM glutamate, the MMP were considerably decreased compared to the control group (p < 0.001). Pretreatment with biatractylolide (10 μM, 15 μM, and 20 μM) could significantly protect PC12 and SH-SY5Y cells from the glutamate-induced reduction of MMP.
Figure 7.

MMP suppressed by glutamate-insult in PC12 and SH-SY5Y cell lines was effectively ameliorated by biatractylolide. After the two cell lines were treated with various concentrations of biatractylolide for 30 min before glutamate treatment for 24 hours, the mitochondrial membrane potential was examined using the Rhodamine-123. ∗∗∗p < 0.005 and ∗∗∗∗p < 0.001 versus control group; #p < 0.05, ##p < 0.01, and ###p < 0.005 versus model group.
3.7. Protein Characterization
PI3K-Akt signaling pathway is essential for the survival of neurons and the effect of PI3K-Akt-GSK3β signal transduction pathway on GSK3β is most important in the regulation of apoptosis. Studies have revealed that activation of PI3K-Akt-GSK3β pathway can have neuroprotective effects [22]. To validate whether this pathway was involved in protective effects of biatractylolide on glutamate-induced PC12 cells and SH-SY5Y cells, we further analyze protein characterization using western blotting. Figures 8(a)–8(d) showed that the expression of GSK3β was observably increased after glutamate treatment compared with control group (p < 0.05) and pretreatment with biatractylolide led to marked decreases in level of GSK3β of compared to the model group. In comparison, treatment of cells with glutamate induced a significant decrease in the level of p-Akt compared to control group (p < 0.01) and pretreatment with biatractylolide led to significant increases in level of p-Akt compared to the model group.
Figure 8.

Effects of biatractylolide on Akt and Gsk3β expressions of PC12 and SH-SY5Y cells induced by glutamate. (a)-(b) represent protein characterization of Akt and Gsk3β after PC12 and SH-SY5Y cells were treated with various concentrations of biatractylolide for 30 min before glutamate treatment for 24 hours. Blots were also probed for β-actin as loading controls. (c)-(d) showed the ratio of different proteins to β-actin was calculated by the band density of each cell line using Image J software. ∗p < 0.05 and ∗∗p < 0.01 versus control group; #p < 0.05, ##p < 0.01, and ###p < 0.005 versus model group.
4. Discussion
Natural drug has been widely used in the development and research of neuroprotective drugs because of its low side effects, multiple targets, and high efficiency. Some studies have reported that biatractylolide could slow down the isolated guinea pig right atrium heart rate and reduce shrinkage force [23]. Furthermore, this effect could be offset by atropine, indicating that biatractylolide might inhibit cholinesterase effect. However, studies of biatractylolide in neurodegenerative diseases are rare currently; thus, the present study evaluated the neuroprotective effect of biatractylolide against glutamate-induced cell injury and its underlying mechanism. MTT assay is a common method to detect the number of viable cells. In our study, different concentrations of biatractylolide significantly increased the survival of PC12 and SH-SY5Y cells in a dose-dependent fashion. When the cell membrane was damaged, the intracellular LDH was released into the culture medium, and the content of LDH was an important index to detect the cell death. In this study, we found that the level of LDH was significantly increased in the model group, while the LDH level in the supernatant was significantly reduced in a concentration-dependent manner after treating with different concentrations of biatractylolide.
Glutamate-induced neurotoxicity may be caused by glutamate and NMDA receptors, which could produce oxidative damage, thus leading to apoptosis [24]. Acridine orange (AO) staining and PI staining [25–27] are important methods for detecting apoptosis. In our study, PI staining and acridine orange staining showed that biatractylolide could reduce the apoptosis of PC12 and SH-SY5Y cells induced by glutamate. We also found that some proportion of necrotic cells appeared in SH-SY5Y cells line, while it was not apparent in the PC12 cell line. We think that the reason for these differences may be that the sensitivity of the two cell lines to glutamate is different but the difference is reasonable and inevitable. So we conclude that biatractylolide has a protective effect on glutamate-induced apoptosis.
Oxidative stress refers to the imbalance of generation and clearance of oxygen free radicals in the body, leading to oxidative damage caused by accumulation of active oxygen and active nitrogen in the body. Numerous studies have shown that there are increases in the level of ROS in neurodegenerative diseases [28, 29], which is implicated in glutamate-induced neurotoxicity and the progression of AD, so the ROS assay kit was used to detect the release of intracellular ROS in this study. Biatractylolide significantly increased the ROS levels compared to the model group in our study, which indicated that biatractylolide significantly reduced the production of glutamate-induced ROS.
Rh123 [30, 31] is a lipophilic cationic fluorescent dye that is permeable to cell membranes and can be selectively enriched in mitochondria. When the cells are in the survival state, Rh123 is accumulated in mitochondria and emits green fluorescence through the cell membrane, but in apoptosis, the ability of cell mitochondria to accumulate Rh123 was lost and the fluorescence intensity was decreased. In this study, the ROS and MMP results suggest that biatractylolide may have neuroprotective effects.
The process of glutamate-induced nerve cell damage is related to multiple signal transduction pathways. Numerous studies show that PI3K-Akt-GSK3β signal transduction pathway plays important roles in the neuroprotective process [32, 33]. Our present study demonstrated that the two cell lines were exposed to glutamate, the levels of p-Akt decreased, and GSK3β increased significantly compared with the control group while the level of t-Akt was almost unchanged. After treating with biatractylolide, the levels of p-Akt increased and GSK3β decreased in a concentration-dependent manner, suggesting that these changes were abolished by pretreatment with biatractylolide and biatractylolide may play a neuroprotective role by activating PI3K-Akt-GSK3β pathway.
As we mentioned above, biatractylolide effectively protects PC12 and SH-SY5Y cells from the glutamate-induced neuronal damage and it is suggested that the mechanism of protective effect of biatractylolide may be mainly related to PI3K-Akt-GSK3β pathway.
Acknowledgments
This research was supported by Hunan Provincial Natural Science Foundation of China (no. 2017JJ2188), Advanced Fund of Hunan Provincial Education Department (no. 16A128), and Traditional Chinese Medicine of Hunan Province, China (no. 201204) (Xing Feng).
Conflicts of Interest
The authors declare that they have no conflicts of interest.
Authors' Contributions
Li Zhu and Ning Ning contributed equally to this paper.
References
- 1.Haapasalo A., Pikkarainen M., Soininen H. Alzheimer's disease: a report from the 7th Kuopio Alzheimer symposium. Neurodegenerative disease management. 2015;5(5):379–382. doi: 10.2217/nmt.15.31. [DOI] [PubMed] [Google Scholar]
- 2.Darras F. H., Pockes S., Huang G., et al. Synthesis, biological evaluation, and computational studies of Tri- and tetracyclic nitrogen-bridgehead compounds as potent dual-acting AChE inhibitors and h H3 receptor antagonists. ACS Chemical Neuroscience. 2014;5(3):225–242. doi: 10.1021/cn4002126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Nakazawa K., McHugh T. J., Wilson M. A., Tonegawa S. NMDA receptors, place cells and hippocampal spatial memory. Nature Reviews Neuroscience. 2004;5(5):361–372. doi: 10.1038/nrn1385. [DOI] [PubMed] [Google Scholar]
- 4.Matsunaga S., Kishi T., Iwata N. Combination therapy with cholinesterase inhibitors and memantine for alzheimer's disease: systematic review and meta-analysis. Alzheimer's & Dementia. 2014;10(4):P859–P860. doi: 10.1016/j.jalz.2014.05.1710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Joubert J., Foka G. B., Repsold B. P., Oliver D. W., Kapp E., Malan S. F. Synthesis and evaluation of 7-substituted coumarin derivatives as multimodal monoamine oxidase-B and cholinesterase inhibitors for the treatment of Alzheimer's disease. European Journal of Medicinal Chemistry. 2017;125:853–864. doi: 10.1016/j.ejmech.2016.09.041. [DOI] [PubMed] [Google Scholar]
- 6.Yang X., Wang Y., Luo J., Liu S., Yang Z. Protective effects of YC-1 against glutamate induced PC12 cell apoptosis. Cellular and Molecular Neurobiology. 2011;31(2):303–311. doi: 10.1007/s10571-010-9622-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Wang K., Zhu X., Zhang K., Zhou F., Zhu L. Gas1 knockdown increases the neuroprotective effect of glial cell-derived neurotrophic factor against glutamate-induced cell injury in human sh-sy5y neuroblastoma cells. Cellular and Molecular Neurobiology. 2016;36(4):603–611. doi: 10.1007/s10571-015-0241-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Sheldon A. L., Robinson M. B. The role of glutamate transporters in neurodegenerative diseases and potential opportunities for intervention. Neurochemistry International. 2007;51(6-7):333–355. doi: 10.1016/j.neuint.2007.03.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Doble A. The role of excitotoxicity in neurodegenerative disease: implications for therapy. Pharmacology and Therapeutics. 1999;81(3):163–221. doi: 10.1016/S0163-7258(98)00042-4. [DOI] [PubMed] [Google Scholar]
- 10.Llorente-Folch I., Rueda C. B., Pérez-Liébana I., Satrústegui J., Pardo B. L-lactate-mediated neuroprotection against glutamate- induced excitotoxicity requires ARALAR/AGC1. Journal of Neuroscience. 2016;36(16):4443–4456. doi: 10.1523/JNEUROSCI.3691-15.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Jiang G., Wu H., Hu Y., Li J., Li Q. Gastrodin inhibits glutamate-induced apoptosis of pc12 cells via inhibition of CaMKII/ASK-1/p38 MAPK/p53 signaling cascade. Cellular and Molecular Neurobiology. 2014;34(4):591–602. doi: 10.1007/s10571-014-0043-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Chang C. H., Chen H. X., Yü G., et al. Curcumin-Protected PC12 Cells Against Glutamate-Induced Oxidative Toxicity. Food Technology and Biotechnology. 2014;52(4):468–478. doi: 10.17113/ftb.52.04.14.3622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Li R., Zhou P., Guo Y., Lee J.-S., Zhou B. Tris (1, 3-dichloro-2-propyl) phosphate induces apoptosis and autophagy in SH-SY5Y cells: Involvement of ROS-mediated AMPK/mTOR/ULK1 pathways. Food and Chemical Toxicology. 2017;100:183–196. doi: 10.1016/j.fct.2016.12.029. [DOI] [PubMed] [Google Scholar]
- 14.Wang C., Wang K., Chen L., Chou D., Liang W. Anti-oxidative abilities of essential oils from atractylodes ovata rhizome. Evidence-Based Complementary and Alternative Medicine. 2011;2011:8. doi: 10.1093/ecam/neq006.204892 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Wang K.-T., Chen L.-G., Wu C.-H., Chang C.-C., Wang C.-C. Gastroprotective activity of atractylenolide III from Atractylodes ovata on ethanol-induced gastric ulcer in vitro and in vivo. Journal of Pharmacy and Pharmacology. 2010;62(3):381–388. doi: 10.1211/jpp.62.03.0014. [DOI] [PubMed] [Google Scholar]
- 16.Liu Y., Jia Z. W., Dong L., Wang R., Qiu G. Q. A randomized pilot study of atractylenolide I on gastric cancer cachexia patients. Evidence-Based Complementary and Alternative Medicine. 2008;5(3):337–344. doi: 10.1093/ecam/nem031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Li W., Wen H.-M., Cui X.-B., Zhang F.-C., Dong J. Chemical constituents in rhizome of Atractylodes macrocephala. Chinese Traditional and Herbal Drugs. 2007;38(10):1460–1462. [Google Scholar]
- 18.Zhou C., Bao M. H., Liu W. P. Effects of biatractylolide on the AD rats induced by Al Cl3. Journal of Changsha Medical University. 2008;7:26–28. [Google Scholar]
- 19.Feng X., Wang Z. L., Lin Y. C. Biatractylolide’s effect on Aβ1-40-induced dementia model rats. Chinese Pharmacological Bulletin. 2009;25(7):949–951. [Google Scholar]
- 20.Xie Y. C., Ning N., Zhu L., et al. Primary the from Atractylodis Macrocephalae Rhizoma as an acetylcholinesterase inhibitor. Evid Based Complement Alternat Med. 2016 doi: 10.1155/2016/7481323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Su Q., Peng M., Zhang Y., et al. Quercetin induces bladder cancer cells apoptosis by activation of AMPK signaling pathway. American Journal of Cancer Research. 2016;6(2):498–508. [PMC free article] [PubMed] [Google Scholar]
- 22.Molz S., Dal-Cim T., Budni J., et al. Neuroprotective effect of guanosine against glutamate-induced cell death in rat hippocampal slices is mediated by the phosphatidylinositol-3 kinase/Akt/glycogen synthase kinase 3β pathway activation and inducible nitric oxide synthase inhibition. Journal of Neuroscience Research. 2011;89(9):1400–1408. doi: 10.1002/jnr.22681. [DOI] [PubMed] [Google Scholar]
- 23.Pu H., Wang Z., Huang Q. Biatractylolide of guinea pigs in vitro atrial muscle function. Chinese Pharmacological Bulletin. 2000;16(5):60–62. [Google Scholar]
- 24.Molinuevo J. L., Lladó A., Rami L. Memantine: Targeting glutamate excitotoxicity in Alzheimer's disease and other dementias. American Journal of Alzheimer's Disease and other Dementias. 2005;20(2):77–85. doi: 10.1177/153331750502000206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Liu K., Liu P.-C., Liu R., Xing W. Dual AO/EB staining to detect apoptosis in osteosarcoma cells compared with flow cytometry. Medical Science Monitor Basic Research. 2015;21:15–20. doi: 10.12659/MSMBR.893327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Dolai N., Islam A., Haldar P. K. Antiproliferative activity and apoptosis inducing mechanism of Anthocephalus cadamba on dalton’s lymphoma ascites cells. Iranian Journal of Pharmaceutical Research. 2016;15(3):505–514. [PMC free article] [PubMed] [Google Scholar]
- 27.Yang Z., Zhao Y., Yao Y., Li J., Wang W., Wu X. Equol induces mitochondria-dependent apoptosis in human gastric cancer cells via the sustained activation of ERK1/2 pathway. Molecules and Cells. 2016;39(10):742–749. doi: 10.14348/molcells.2016.0162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Bhat A. H., Dar K. B., Anees S., et al. Oxidative stress, mitochondrial dysfunction and neurodegenerative diseases; a mechanistic insight. Biomedicine & Pharmacotherapy. 2015;74:101–110. doi: 10.1016/j.biopha.2015.07.025. [DOI] [PubMed] [Google Scholar]
- 29.Kim G. H., Kim J. E., Rhie S. J., Yoon S. The role of oxidative stress in neurodegenerative diseases. Experimental Neurobiology. 2015;24(4):325–340. doi: 10.5607/en.2015.24.4.325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Wang L.-H., Chang G.-Q., Zhang H.-J., et al. Neutrophil gelatinase-associated lipocalin regulates intracellular accumulation of Rh123 in cancer cells. Genes to Cells. 2012;17(3):205–217. doi: 10.1111/j.1365-2443.2012.01584.x. [DOI] [PubMed] [Google Scholar]
- 31.Wu Y. M., Xia X. Y., Pan L. J., et al. Evaluation of sperm mitochondrial function using Rh123/PI dual fluorescent staining. Zhong Hua Nan Ke Xue. 2006;12(9):803–806. [PubMed] [Google Scholar]
- 32.Liu F., Gong X., Zhang G., Marquis K., Reinhart P., Andree T. H. The inhibition of glycogen synthase kinase 3β by a metabotropic glutamate receptor 5 mediated pathway confers neuroprotection to Aβ peptides. Journal of Neurochemistry. 2005;95(5):1363–1372. doi: 10.1111/j.1471-4159.2005.03474.x. [DOI] [PubMed] [Google Scholar]
- 33.Shen Y., Zhang Q., Gao X., Ding F. An active fraction of achyranthes bidentata polypeptides prevents apoptosis induced by serum deprivation in SH-SY5Y cells through activation of PI3K/Akt/Gsk3β pathways. Neurochemical Research. 2011;36(11):2186–2194. doi: 10.1007/s11064-011-0543-x. [DOI] [PubMed] [Google Scholar]