

Abstract
High altitude pulmonary edema (HAPE) is a paradigm of pulmonary edema. Mutations in regulator of telomere elongation helicase1 (RTEL1) represent an important contributor to risk for pulmonary fibrosis. However, little information is found about the association between RTEL1 and HAPE risk. The present study was undertaken to tentatively explore the potential relation between single-nucleotide polymorphisms (SNPs) in RTEL1 and HAPE risk in Chinese Han population. A total of 265 HAPE patients and 303 healthy controls were included in our case-control study. Four SNPs in RTEL1 were selected and genotyped using the Sequenom MassARRAY method. Odds ratios (ORs) and 95% confidence intervals (95% CIs) were calculated by unconditional logistic regression with adjustment for gender and age. All P values were Bonferroni corrected, and statistical significance was set at P < .0025 (.05/20). In allelic model analysis, we found that the allele “G” of rs6089953 and rs6010621 and the allele “A” of rs2297441 were associated with decreased risk of HAPE. In the genetic model analysis, we found that rs6010621, rs6089953, and rs2297441 were relevant to decreased HAPE risk under dominant model (rs6010621: OR = 0.55; 95% CI = 0.39–0.78; P = .001; rs6089953: OR = 0.68; 95% CI = 0.48–0.96; P = .027; rs2297441: OR = 0.63; 95% CI = 0.45–0.89; P = .008, respectively) and additive model (rs6010621: OR = 0.51; 95% CI = 0.46–0.81; P < .001; rs6089953: OR = 0.72; 95% CI = 0.55–0.95; P = .022; rs2297441: OR = 0.73; 95% CI = 0.57–0.95; P = .019, respectively). SNPs rs6010621 remained significant after Bonferroni correction (P < .0025). In addition, haplotype “GG, GT, AT” of rs6089953-rs6010621 were detected significantly associated with HAPE risk (P < .05), haplotype “GG” remained significant after Bonferroni correction (P < .0025). Our findings provide new evidence for the association between SNPs in RTEL1 and a decreased risk HAPE in the Chinese population. The results need further confirmation.
Keywords: case-control study, genomic stability, HAPE, RTEL1, single-nucleotide polymorphism (SNP), telomere length
1. Introduction
High altitude pulmonary edema (HAPE) is a paradigm of pulmonary edema experienced by some people under hypobaric hypoxic condition of high altitude after rapidly ascending above 2500 m from sea level. The pathogenesis of HAPE is still debated. It is characterized by intense smooth muscle proliferation and exaggerated hypoxic vasoconstriction at decreased levels of alveolar oxygen, leading to sustained excessive pulmonary pressure and vascular leakage through over perfusion, resulting in abnormal buildup of fluid in the lung.[1,2] Many people will be at risk of high altitude sickness shortly to a moderately hypoxic environment, but not all affected individuals will develop HAPE, suggesting an innate contribution to HAPE susceptibility. Although the impact of altitude attained and rate of ascent on the etiology of HAPE is well known, there are clues to the contribution of a genetic background that influence the efficacy of altitude acclimatization.[3]
Regulator of telomere elongation helicase1 (RTEL1) is a DNA helicase crucial for the regulation of telomere length that plays a role in a variety of fundamental cellular mechanisms such as DNA replication, genome stability, DNA repair, and telomere maintenance.[4,5] In the absence of RTEL1, shortened telomere length, chromosome breaks, and translocations were observed, indicating that RTEL1 protein contribute to genomic stability.[6] Previous studies have found that genetic polymorphisms of RTEL1 were associated with glioma risk.[7–11] A recent study found that mutations in RTEL1 represent an important contributor to risk for pulmonary fibrosis.[12] Defective telomere maintenance leading to telomere shortening is a cause of pulmonary fibrosis.[13–15] So we hypothesized the variants of RTEL1 may have a potential relevance with lung-related disease.
A previous study reported that telomere length was significantly longer at moderate attitude than at sea-level or at simulated high attitude.[16] However, to date, little research directly focused on the association between RTEL1 polymorphisms and HAPE risk. The present study was undertaken to tentatively explore the potential relation between common single-nucleotide polymorphisms (SNPs) in RTEL1 and HAPE risk in Chinese Han population, aiming to shed a new light on the association between mutations in RTEL1 and HAPE susceptibility.
2. Materials and methods
2.1. Subjects
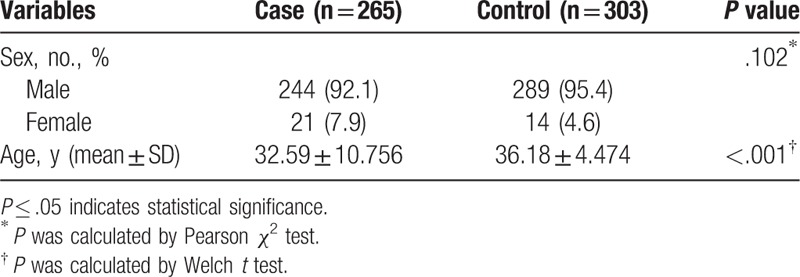
A number of 265 HAPE cases were recruited from the Affiliated Hospital of Xizang Minzu University. We recruited a total of 303 healthy unrelated samples from the outpatient departments at the hospital. The individuals in the HAPE and control groups were all unrelated, and information of all the subjects has been critically reviewed. The cases (244 males, 21 females) and controls (289 males, 14 females) did not show a significantly different sex distribution (P = .102), while they showed a significantly different age distribution (P < .001). All of the subjects were Han Chinese and residents living in Northwest China. The diagnosis of HAPE patients was dependent on standard criteria,[17] including patient interviews like cough, dyspnea, cyanosis at rest, imageological examination like X-ray radiograph, computed tomography (CT) of the patient chest or magnetic resonance imaging.[18] All the HAPE patients eventually show that chest radiographic troves of infiltrates consistent with pulmonary edema. None of the participants who were included in the study had any kind of disorder, including diabetes, renal disease, hypertension, or cardiopulmonary or coronary disorders, which could have affected the study. We collected blood samples from all the participants, and the use of the samples was approved by the Human Research Committee of the Affiliated Hospital of Xizang Minzu University. The study protocol was reviewed and approved by the Ethics Committee of Xizang Minzu University.
2.2. Gene polymorphisms analyses
In this study, 4 SNPs in RTEL1 were selected from the previous study for analysis.[19,20] The lower frequency alleles were coded as the minor allele. Minor allele frequencies of all SNPs were > 5% in the HapMap of the Chinese Han Beijing population. Genomic DNA was isolated from whole blood samples using the GoldMag-Mini Purification Kit (GoldMagCo Ltd, Xian city, China), and DNA concentrations were measured using the NanoDrop2000 (Thermo Scientific, Waltham, MA). SequenomMassarrayAssay Design 3.0 software was used to design a multiplexed SNP Mass EXTENDED Assay.[21–23] Genotyping was performed using the SequenomMassARRAY RS1000 and analyzed using SequenomTyper 4.0 Software.[24]
2.3. Statistical analyses
Fisher exact test was used to assess the variation in each SNP frequency from the Hardy–Weinberg equilibrium (HWD) in the control subjects. Differences in SNP genotype and allele distribution between cases and controls were compared using logistic regression. ORs and 95% CIs were determined using unconditional logistic regression analysis with adjustments for age and sex.[25]. All the statistical analyses above were performed with Microsoft Excel and SPSS 20.0 (SPSS Inc, Chicago, IL). Finally, the patterns of linkage disequilibrium (LD) and haplotype construction were evaluated by Haploview software package (version 4.2).[26] All P values were Bonferroni corrected, and statistical significance was set at P < .0025 (.05/20).
3. Results
A number of 265 cases and 303 controls were enrolled for genetic association analyses in our study (Table 1). Our sample size was relatively small compared with the previous GWAS study. However, we calculate the expected sample size used the online calculation software (http://sampsize.sourceforge.net/iface/s3.html#ccp). If we wish to conduct a case-control study to assess whether HAPE risk may be associated with RTEL1 polymorphisms and we wish to detect an odds-ratio of 2 with power 90%, the number of cases and controls were 188 versus 188. So the present sample size was appropriate.
Table 1.
Characteristics of cases and controls in this study.
Four SNPs in RTEL1 gene were analyzed (Table 2). We compared the differences in frequency distributions of alleles between cases and controls by χ2 test. We found that the allele “G” of rs6089953 and rs6010621 and the allele “A” of rs2297441 were associated with decreased risk of HAPE (rs6089953: OR = 0.70; 95% CI = 0.54–0.92; P = .009; rs6010621: OR = 0.60; 95% CI = 0.46–0.79; P < 0.001; rs2297441: OR = 0.74; 95% CI = 0.58–0.96; P = .021). SNPs rs6010621 remained significant after Bonferroni correction (P < .0025).
Table 2.
Allele frequency of SNPs in cases and controls and association with risk of HAPE.

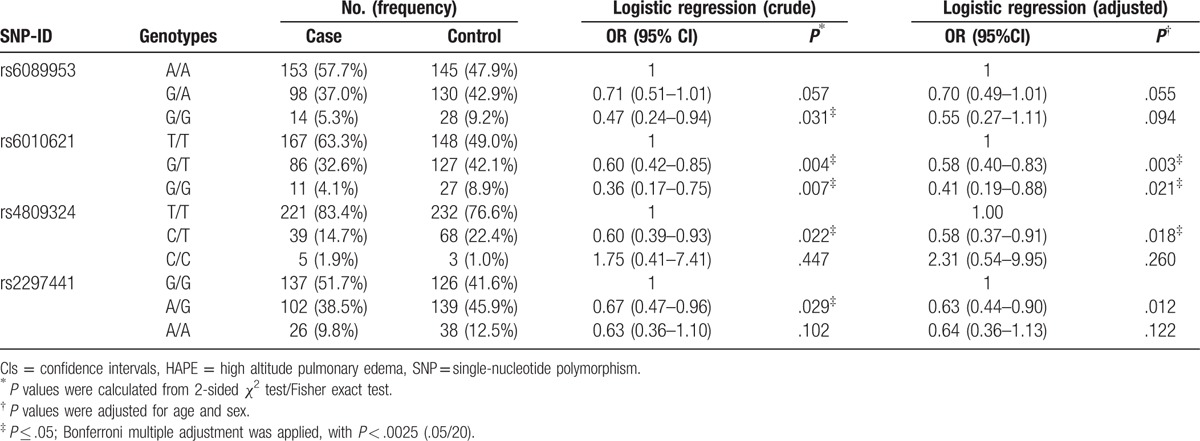
The genotype frequencies of the RTEL1 polymorphisms were shown in Table 3. We identified 2 SNPs associated with the decreased HAPE risk after adjusted for age and gender. Compared with the TT genotype, the “GG” and “GT” frequencies of rs6010621 polymorphism among cases were different from the controls (GG vs TT: OR = 0.41; 95% CI = 0.19–0.88; P = .021; GT vs TT: OR = 0.58, 95% CI = 0.40–0.83; P = .003; respectively), which suggested that the rs6010621 polymorphism had a decreased effect on HAPE risk. Additionally, compared with individuals with the rs4809324 TT genotype, individuals with CT genotype had a decreased HAPE risk (CT vs TT: OR = 0.58; 95% CI = 0.37–0.91; P = .018). However, neither of them was significant after Bonferroni correction (P < .0025).
Table 3.
Distributions of genotypes of SNPs and their associations with risk of HAPE.
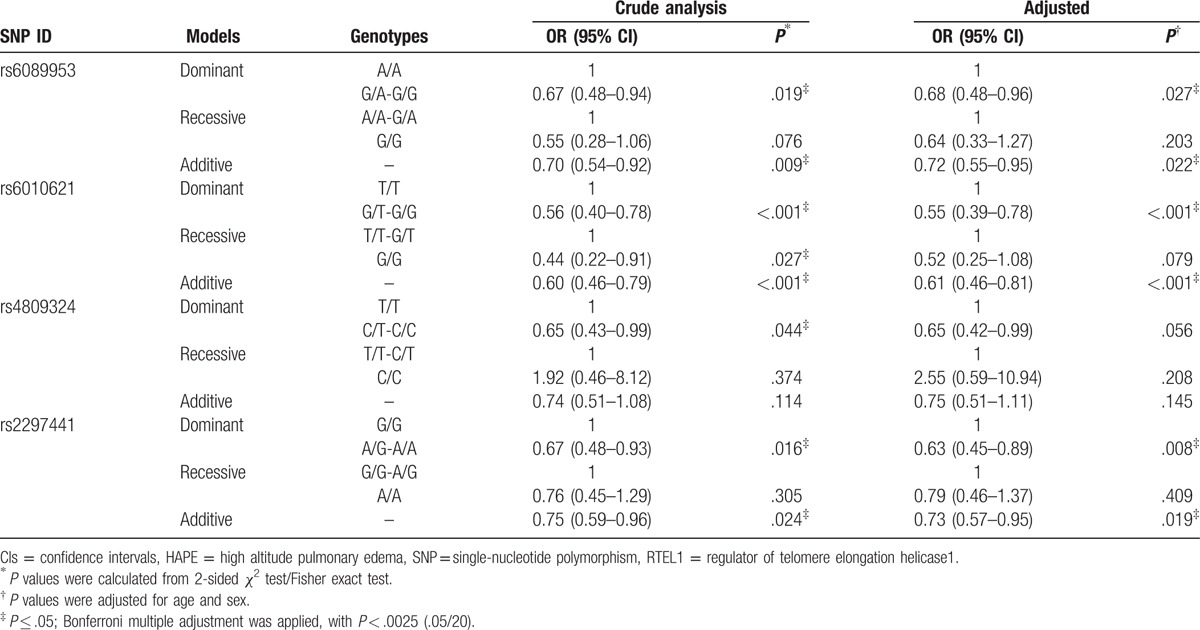
We further analyzed the association of SNPs and HAPE risk using logistic regression including dominant model, recessive model, and additive model. We assumed the minor allele of each SNP as a risk factor compared with the wild-type allele (Table 4). We found that rs6010621, rs6089953, and rs2297441 were relevant to decreased HAPE risk under dominant model (rs6010621: OR = 0.55; 95% CI = 0.39–0.78; P = .001; rs6089953: OR = 0.68; 95% CI = 0.48–0.96; P = .027; rs2297441: OR = 0.63; 95% CI = 0.45–0.89; P = .008, respectively) and additive model (rs6010621: OR = 0.51; 95% CI = 0.46–0.81; P < .001; rs6089953: OR = 0.72; 95% CI = 0.55–0.95; P = .022; rs2297441: OR = 0.73; 95% CI = 0.57–0.95; P = .019, respectively). SNPs rs6010621 remained significant after Bonferroni correction (P < .0025).
Table 4.
Analysis of association between genotypic model of RTEL1 gene polymorphisms and the risk of HAPE.
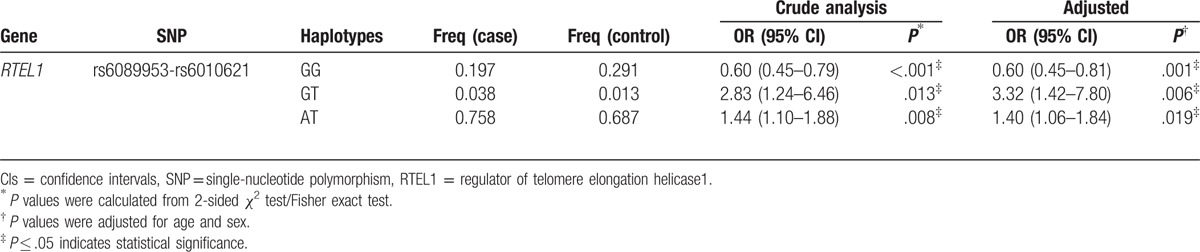
Furthermore, the linkage D analysis for the 4 SNPs was performed, 1 block rs6089953-rs6010621 in RTEL1 gene showed strong linkage (Fig. 1). On the basis of the LD pattern of RTEL1 gene, we conducted haplotype-based analysis. Haplotype “GG” of rs6089953-rs6010621 was detected associated with a decreased risk of HAPE (OR = 0.60; 95% CI = 0.45–0.81; P = .001), while the haplotypes “GT” and “AT” were associated with increased risk of HAPE (GT: OR = 3.32; 95% CI = 1.42–7.80; P = .006; AT: OR = 1.40; 95% CI = 1.06–1.84; P = .019) (Table 5). However, only haplotype “GG” remained significant after Bonferroni correction (P < .0025).
Figure 1.
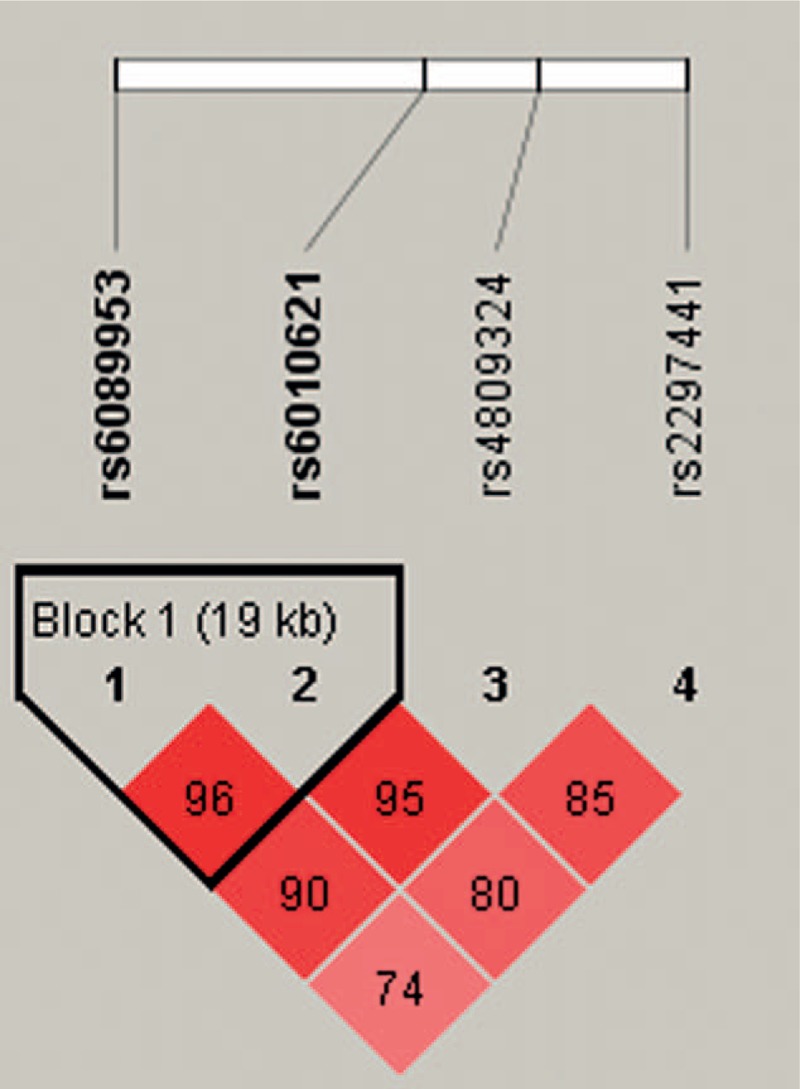
Haplotype block map for the SNPs in RTEL1 gene. A standard color scheme was used to display linkage disequilibrium (LD), with bright red corresponding to strong LD (LOD = 2, D’ = 1), white corresponding to no LD (LOD < 2, D’ < 1), and pink/red (LOD = 2, D’ < 1) and blue (LOD < 2, D’ = 1) corresponding to intermediate LD. RTEL1 = regulator of telomere elongation helicase1, SNPs = single-nucleotide polymorphisms.
Table 5.
Haplotype frequencies of SNPs in the RTEL1 gene and the association with HAPE risk in case and control subjects.
4. Discussion
In this case-control study, we estimated the relationship between RTEL1 and HAPE risk, confirmed by multivariate analysis adjusted for age and gender, the “G” of rs6089953, the“ G” of rs6010621, the “A” of rs2297441 in RTEL were associated with a decreased risk of HAPE, rs6010621 remained significant after Bonferroni correction. We also observed that the GG, GT, and AT haplotypes of rs6089953-rs6010621 in RTEL1 were significantly associated with decreased susceptibility to HAPE compared with the wild type, haplotype “GG” remained significant after Bonferroni correction. These results provided the first evidence that telomere length maintaining gene RTEL1 may play a potential part in the risk of HAPE in Chinese Han population.
RTEL1 is located in 20q13.3, regulates genomic stability and telomere maintenance. Inactivation of RTEL1 caused chromosome breaks, fusions, and telomere loss in mice.[27] The human RTEL1 is an ortholog of the mouse RTEL1, and their protein products likely play similar roles. It is more likely to happen gene replacement and sister chromatid exchange when RTEL1 is deficient in cells.[28]RTEL1-deficient stem cells were susceptible to localized chromosome breakage, and the proliferative ability was significantly decreased in cell differentiation.[29] Additionally, RTEL1-deficient cell was more sensitive to several DNA damaging agents during the embryonal developmental stages. However, in the present study, carriers with these variants in RTEL1 showed a significantly decreased susceptibility to HAPE. Although our data offer clues for the association between RTEL1 and HAPE risk, comprehensive biochemical and functional analysis of RTEL1 polymorphisms is needed to confirm the results.
The Bonferroni correction is the most commonly used method when making multiple statistical tests. We determined that SNP rs6010621 remained significant after Bonferroni correction, while rs6089953 and rs2297441 were not significant. This may result from the strict SNP filtering criteria and small sample size. Additionally, the Bonferroni correction has its inherent defects,[30] we adjusted the P values based on 4 SNPs and 5 genetic models, some significant differences may be deemed insignificant effects, and false-positive results may be generated.
Some possible limitations in this case- control analysis should be addressed. First, our sample size was relatively small, we did not do further subgroup analyses based on age or gender. Second, HAPE is a heterogeneous disease with many other risk factor, we did not investigate the gene–gene and gene–environment interactions because of the limited data. Third, all the samples were collected from hospital, type false positive error for association study may be generated. Based on the limitations of the present study listed above, detailed studies are warranted to confirm our findings. Moreover, more gene–gene and gene–environment interactions should also be considered in future analysis.
5. Conclusions
Our findings provide new evidence for the association between SNPs in RTEL1 and HAPE risk in the Chinese population for the first time. Because RTEL1 is required for genomic stability, we predict that changes in telomere stability may underlie the increased susceptibility to HAPE. Further functional studies are required to test this hypothesis.
Footnotes
Abbreviations: CIs = confidence intervals, HAPE = high altitude pulmonary edema, HWE = Hardy–Weinberg equilibrium, LD = linkage disequilibrium, MAF = minor allele frequency, OR = odds ratios, RTEL1 = regulator of telomere elongation helicase1, SNPs = single nucleotide polymorphisms.
HR and XH contributed equally to this work.
This work is supported by the Innovation Support Program for College Young Teachers Projects of Xizang (Tibet) Autonomous Region(QCZ2016-38), and Natural Science Foundation of Xizang (Tibet) Autonomous Region (2016ZR-MY-02).
The authors have no conflicts of interest to disclose.
References
- [1].Bartsch P, Mairbaurl H, Maggiorini M, et al. Physiological aspects of high-altitude pulmonary edema. J Appl Physiol (1985) 2005;98:1101–10. [DOI] [PubMed] [Google Scholar]
- [2].Dehnert C, Risse F, Ley S, et al. Magnetic resonance imaging of uneven pulmonary perfusion in hypoxia in humans. Am J Respir Crit Care Med 2006;174:1132–8. [DOI] [PubMed] [Google Scholar]
- [3].MacInnis MJ, Wang P, Koehle MS, et al. The genetics of altitude tolerance: the evidence for inherited susceptibility to acute mountain sickness. J Occup Environ Med 2011;53:159–68. [DOI] [PubMed] [Google Scholar]
- [4].Ding H, Schertzer M, Wu X, et al. Regulation of murine telomere length by Rtel: an essential gene encoding a helicase-like protein. Cell 2004;117:873–86. [DOI] [PubMed] [Google Scholar]
- [5].Uringa EJ, Youds JL, Lisaingo K, et al. RTEL1: an essential helicase for telomere maintenance and the regulation of homologous recombination. Nucleic Acids Res 2011;39:1647–55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [6].Uringa EJ, Lisaingo K, Pickett HA, et al. RTEL1 contributes to DNA replication and repair and telomere maintenance. Mol Biol Cell 2012;23:2782–92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [7].Jin TB, Zhang JY, Li G, et al. RTEL1 and TERT polymorphisms are associated with astrocytoma risk in the Chinese Han population. Tumor Biol 2013;34:3659–66. [DOI] [PubMed] [Google Scholar]
- [8].Li G, Jin T, Liang H, et al. RTEL1 tagging SNPs and haplotypes were associated with glioma development. Diagn Pathol 2013;8:83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [9].Du SL, Geng TT, Feng T, et al. The RTEL1 rs6010620 polymorphism and glioma risk: a meta-analysis based on 12 case-control studies. Asian Pacific J Cancer Prevent APJCP 2014;15:10175. [DOI] [PubMed] [Google Scholar]
- [10].Jin T, Wang Y, Li G, et al. Analysis of difference of association between polymorphisms in the XRCC5, RPA3 and RTEL1 genes and glioma, astrocytoma and glioblastoma. Am J Cancer Res 2015;5:2294. [PMC free article] [PubMed] [Google Scholar]
- [11].Yang B, Heng L, Du S, et al. Association between RTEL1, PHLDB1, and TREH polymorphisms and glioblastoma risk: a case-control study. Med Sci Monit 2015;21:1983–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [12].Kropski JA, Loyd JE. Telomeres revisited: RTEL1 variants in pulmonary fibrosis. Eur Respir J 2015;46:312–4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [13].Cogan JD, Kropski JA, Zhao M, et al. Rare variants in RTEL1 are associated with familial interstitial pneumonia. Am J Respir Crit Care Med 2015;191:646–55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [14].Kannengiesser C, Borie R, Menard C, et al. Heterozygous RTEL1 mutations are associated with familial pulmonary fibrosis. Eur Respir J 2015;46:474–85. [DOI] [PubMed] [Google Scholar]
- [15].Alder JK, Chen JJ, Lancaster L, et al. Short telomeres are a risk factor for idiopathic pulmonary fibrosis. Proc Natl Acad Sci U S A 2008;105:13051–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [16].Wang Y, Zhou WD, Yang Y, et al. Telomeres are elongated in rats exposed to moderate altitude. J Physiol Anthropol 2014;33:19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [17].Hultgren HN, Marticorena EA. High altitude pulmonary edema. Epidemiologic observations in Peru. Chest 1978;74:372–6. [DOI] [PubMed] [Google Scholar]
- [18].Pennardt A. High-altitude pulmonary edema: diagnosis, prevention, and treatment. Curr Sports Med Rep 2013;12:115–9. [DOI] [PubMed] [Google Scholar]
- [19].Jin TB, Zhang JY, Li G, et al. RTEL1 and TERT polymorphisms are associated with astrocytoma risk in the Chinese Han population. Tumour Biol 2013;34:3659–66. [DOI] [PubMed] [Google Scholar]
- [20].Song X, Zhou K, Zhao Y, et al. Fine mapping analysis of a region of 20q13.33 identified five independent susceptibility loci for glioma in a Chinese Han population. Carcinogenesis 2012;33:1065–71. [DOI] [PubMed] [Google Scholar]
- [21].Gabriel S, Ziaugra L, Tabbaa D. SNP genotyping using the Sequenom MassARRAY iPLEX platform. Curr Protoc Hum Genet 2009;Chapter 2: Unit 2.12. [DOI] [PubMed] [Google Scholar]
- [22].Kochl S, Niederstatter H, Parson W. DNA extraction and quantitation of forensic samples using the phenol-chloroform method and real-time PCR. Methods Mol Biol 2005;297:13–30. [DOI] [PubMed] [Google Scholar]
- [23].Trembizki E, Smith H, Lahra MM, et al. High-throughput informative single nucleotide polymorphism-based typing of Neisseria gonorrhoeae using the Sequenom MassARRAY iPLEX platform. J Antimicrob Chemother 2014;69:1526–32. [DOI] [PubMed] [Google Scholar]
- [24].Carlson CS, Eberle MA, Rieder MJ, et al. Selecting a maximally informative set of single-nucleotide polymorphisms for association analyses using linkage disequilibrium. Am J Hum Genet 2004;74:106–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [25].Bland JM, Altman DG. Statistics notes. The odds ratio. BMJ 2000;320:1468. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [26].Yong Y, Lin H. SHEsis, a powerful software platform for analyses of linkage disequilibrium, haplotype construction, and genetic association at polymorphism loci. Cell Res 2005;15:97–8. [DOI] [PubMed] [Google Scholar]
- [27].Youds JL, Mets DG, McIlwraith MJ, et al. RTEL-1 enforces meiotic crossover interference and homeostasis. Science (New York, NY) 2010;327:1254–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [28].Wang RC, Smogorzewska A, de Lange T. Homologous recombination generates T-loop-sized deletions at human telomeres. Cell 2004;119:355–68. [DOI] [PubMed] [Google Scholar]
- [29].Wu X, Sandhu S, Nabi Z, et al. Generation of a mouse model for studying the role of upregulated RTEL1 activity in tumorigenesis. Transgenic Res 2012;21:1109–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [30].Perneger TV. What's wrong with Bonferroni adjustments. BMJ 1998;316:1236–8. [DOI] [PMC free article] [PubMed] [Google Scholar]