

ABSTRACT
Ca2+-activated Cl− currents have been implicated in many cellular processes in different cells, but for many years, their molecular identity remained unknown. Particularly intriguing are Ca2+-activated Cl− currents in olfactory transduction, first described in the early 90s. Well characterized electrophysiologically, they carry most of the odorant-induced receptor current in the cilia of olfactory sensory neurons (OSNs). After many attempts to determine their molecular identity, TMEM16B was found to be abundantly expressed in the cilia of OSNs in 2009 and having biophysical properties like those of the native olfactory channel. A TMEM16B knockout mouse confirmed that TMEM16B was indeed the olfactory Cl− channel but also suggested a limited role in olfactory physiology and behavior.
The question then arises of what the precise role of TMEM16b in olfaction is. Here we review the long story of this channel and its possible roles.
KEYWORDS: calcium activated chloride channels, electrophysiology, olfactory behavior, olfactory transduction, TMEM16B
Introduction
In 1991, a revolution began in the olfactory field with the discovery of olfactory receptors (ORs) by Linda Buck and Richard Axel.1 This Nobel Prize discovery fueled rapid progress in our understanding how ORs are singly expressed in a given olfactory sensory neuron (OSN), how olfactory axons target to the brain and what the logic of olfactory coding might be.
Coincidentally, at the same time, Kleene and Gesteland discovered,2 in an effort to understand the role of Ca2+ in olfactory transduction, a Ca2+-activated current in frog olfactory cilia that was almost entirely carried by Cl−. Later, Ca2+-activated Cl− currents were described in olfactory cilia of all vertebrates tested, including fish, amphibians, and rodents.3-6 It might seem much easier to determine the role of a single ion channel compared with 1000 ORs expressed in the mouse. But the quest to understand the role of the Ca2+-activated Cl− channel (CaCC) turned into a longer - at times convoluted - tale about olfactory signal transduction and the CaCC. This tale - that we will try to keep short - is still being, literally, written 25 y later.
By the mid 90s, the molecular identity of the olfactory transduction components needed to activate an odorant response had been completed, bar one. The identity of the CaCC participating in transduction remained elusive, hampering efforts to understand and characterize the role of this channel. Historically, the description, cloning, and characterization of Cl− channels, in general, have always lagged behind that of Na+, Ca2+ and K+ channels. Although Cl− is the most abundant anion in the extracellular fluids and plasma, Cl− currents have long been considered a sort of background conductance with a lack of substantial interest by neuroscientists. Additionally, the lack of knowledge about natural toxins that target the Cl− conductance certainly has not helped.
But – as it happens - Cl− channels have received increasing attentions in the last few years. This interest has been partly driven by the appreciation that Cl− can be seen as a “free agent:” it is unbound by, generally speaking, rather fixed ionic gradients across cell membranes like those for Na+ and K+, and thus can be used as an inhibitory or an excitatory ion. The increased interest in Cl− channels led to a rapid increase in our knowledge of their molecular identity and function. Indeed, in 2008 a new family of CaCCs was discovered,7-9 and it was found that one member of this family, TMEM16B (also known as Anoctamin 2) is expressed in OSNs and is the long sought after CaCC, the main topic of our review.
We will guide the reader through the numerous twists of CaCCs, from Cl− current characterization in native olfactory cilia, up to their cloning and possible role in olfaction.
Odorant transduction and its components
The very first step of odor detection takes place in the olfactory epithelium (OE) in which the bipolar OSNs are embedded. OSNs are the first functional unit of the olfactory system; their axons leave the epithelium to reach the olfactory bulb (OB) in the brain, and they extend their single dendrite to the apical surface of the OE ending with a knob which bears around 10–20 cilia. The components of the olfactory transduction machinery are expressed almost exclusively in the cilia (Fig. 1) and they convert the chemical information of the odorant-OR interaction into an electrical signal ultimately triggering action potentials (APs) that are sent to the brain.
Figure 1.
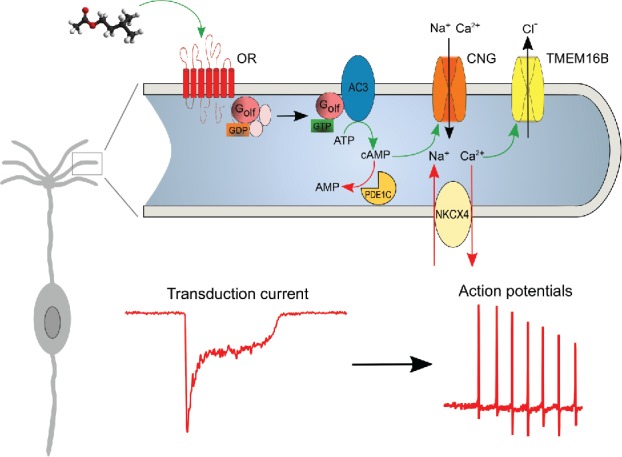
Olfactory signal transduction. The transduction current is generated upon activation of the olfactory receptor (OR) by an odorant in the cilia of an olfactory sensory neuron (OSN) that triggers a series of events (indicated by green arrows) that lead to action potential firing that is sent to the olfactory bulb (OB) in the brain. A series of negative feedbacks (indicated by red arrows) brings the situation back to resting levels. Golf, olfactory G protein; CNG, cyclic nucleotide-gated; NCKX4, Na+/Ca2+/K+ exchanger 4; PDE1C, phosphodiesterase 1C; AC3, adenylyl cyclase 3.
ORs are 7 transmembrane G protein-coupled receptors and a single OR can respond to several different odorants and may be broadly tuned or more narrowly selective.10 When an odorant molecule binds to an OR, the ensuing conformational change triggers the activation of the G protein Golf,11 which in turn favors the enzymatic activity of the adenylyl cyclase 3 (AC3),12 thus leading to an increase in intra-ciliary levels of cAMP. cAMP opens cyclic nucleotide-gated (CNG) channels,13,14 allowing Na+ and Ca2+ to enter the ciliary lumen. This first series of events is necessary and sufficient to depolarize the OSN that will fire APs to be sent to the OB.
Ca2+ increase inside the cilia is responsible for various mechanisms: among those, the activation of CaCCs that leads to Cl− exiting the cilia, further depolarizing the cell. The presence of a Na+-K+-2Cl− cotransporter, NKCC1, ensures that the Cl− concentration inside the cilia is kept high, in the same range of that present in the mucus outside the cilia, supporting the depolarizing effect of CaCCs activation.15-17 The unusually high Cl− concentration inside the cilia is the interesting feature that determines the depolarizing role of the Cl− current. Although its exact value may be species-specific the measured intracellular Cl− concentration varies from 55 mM up to 69 mM depending also on the techniques used to measure it;15,16 those values are close to the Cl− concentration measured in the mucus surrounding the knob and cilia of the OSNs. Energy dispersive X-ray micro analysis on cryosections of rat OE estimated a Cl− concentration of 69 mM inside the cells and 55 mM in the mucus, with a calculated equilibrium potential for Cl− of +6 mV.16 In another study, Cl− concentration was estimated from the intact OE of mice and rats using 2-photon fluorescence lifetime imaging of the Cl− sensitive dye MQAE (Fig. 2A & B). A maximal concentration reaching up to 70 mM was found within the dendritic knob, decreasing to 55 mM along the dendrite in the presence of a high extracellular Cl− concentration.15 For a lower extracellular Cl− concentration of 50 mM the gradient disappeared (Fig. 2B), suggesting the presence of an apical Cl− uptake system.15 This data support the view that a Cl− accumulation process charges the resting cilia prompting an excitatory Cl− efflux in olfactory transduction.17-19
Figure 2.
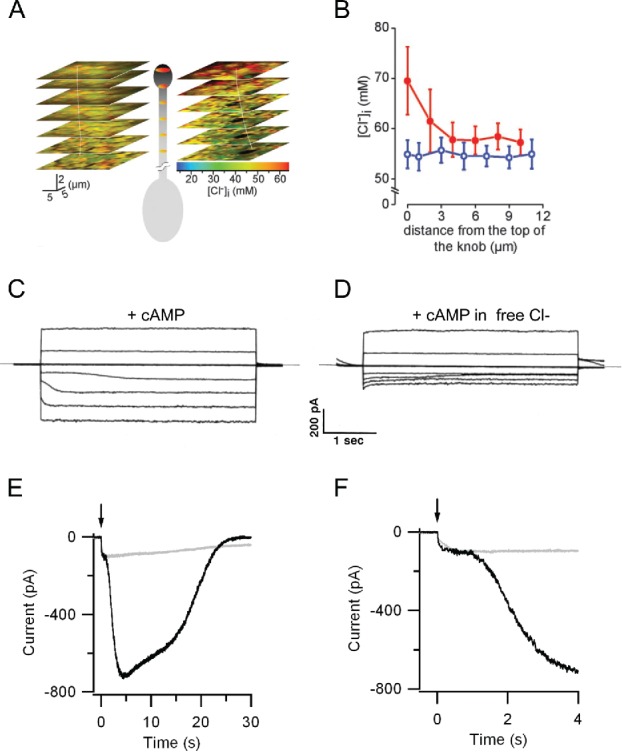
Cl− gradients in olfactory sensory neurons and activation of Ca2+-activated Cl− currents in olfactory cilia. The depolarizing action of the Ca2+-activated Cl− is possible because the intraciliary Cl− concentration is around 60 mM in the knob/cilia region of an OSN (A and B). Modified from Kaneko et al. 2004 with permission.15 A series of voltage steps in an excised frog cilium showed the existence of cAMP-activated current followed by a secondary current (C) that was absent when Cl− was removed from the pseudointracellular bath (D) Modified from Kleene 1993 with permission.61 This secondary current was also present in mouse OSNs following cAMP un-caging (E and F). It constitutes up to the 90% of the transduction current and was blocked by the Cl− channel blocker niflumic acid (gray traces). © American Physiological Society. Reproduced by permission of American Physiological Society. Permission to reuse must be obtained from the rightsholder.19
Different mechanisms are responsible for the termination of the response. cAMP is hydrolyzed by 2 phosphodiesterases (PDEs), PDE1C in the cilia and PDE4A in the other part of OSNs. Although little is known about the fate of ciliary cAMP, recent knockout studies in mice revealed that hydrolysis of ciliary cAMP is not a rate-limiting factor for termination kinetics.20,21 Indeed, it is most likely that the diffusional escape of cAMP from the ciliary lumen is sufficient to allow a rapid response termination.22,23
However, not only the CNG channels have to close, but also the CaCCs. Their closure kinetics are dictated by the rate of ciliary Ca2+ removal, which is mainly due to the activity of Na+/Ca2+ exchanger. The role of a Na+/Ca2+ exchanger in OSNs has been suggested since the mid-1990s. Preventing Ca2+ extrusion by reducing extracellular Na+, and thus abolishing the driving gradient that fuels the exchange, prolongs the odorant-induced receptor current by seconds, generating a prolonged Ca2+-activated Cl− current.24-26
More recently it has been shown that the potassium-dependent Na+/Ca2+ exchanger 4 (NCKX4) is expressed in the cilia of OSNs and plays the major role in Ca2+ extrusion and therefore, in controlling the termination of the odorant response.27-29 A further contribution to Ca2+ dynamics inside the cilia of OSNs is given by mitochondria in the dendritic knob. Not only do they contribute to the low resting Ca2+ but also they are responsible for its clearance during odorant stimulation. Noteworthy is that odorant responses are also affected since pharmacological alteration of the inner mitochondrial membrane potential shifts the dynamic range of the OSN stimulus-response function.30
cAMP-activated currents
In OSN cilia, cAMP activates a cationic current through CNG channels. First described by Nakamura and Gold (1987),13 the cAMP-activated current has been extensively characterized and proven to be required for OSN response to most odorants.14 The native olfactory CNG channels are composed of 3 different subunits encoded by CNGA2, CNGA4 and CNGB1b forming a tetramer with the stoichiometry of 2 CNGA2, 1 CNGA4 and 1 CNGB1b.31-35 They are expressed in the cilia of all canonical OSNs.34,36,37
The olfactory CNG channels are slightly more permeable to Na+ than to K+,38 and the current-voltage relation shows slight outward rectification in divalent free conditions.34,38,39 The relation between the concentration of cAMP and CNG current is well fitted by a Hill function with a Hill coefficient ranging from 1.3 to 2.3, indicating that the binding of at least 2 cAMP molecules is necessary to open the channel. It was shown that binding of the second cAMP molecule brings the channel almost to its maximum open probability,40,41 which is just 0.7–0.8.5,39,42,43 Once activated, the current does not spontaneously inactivate in the presence of cAMP.39,44,45 Ca2+ has a complex effect on CNG channel activity. Under physiologic conditions, much of the inward current is carried by Ca2+,46 but extracellular Ca2+ and Mg2+ greatly reduce inward CNG current by open-channel block. Moreover, elevated cytoplasmic Ca2+, together with one or more Ca2+-binding factors such as Ca2+–calmodulin, reduces the sensitivity of the channels to cAMP, a mechanism relevant for adaptation.47-52
cAMP activates cationic CNG channels in the ciliary membrane and knockout mice missing the CNG channel subunit CNGA2 lacked electro-olfactogram (EOG) responses (which represent the recorded summated generator potential from the surface of the OE)53,54 to most odorants tested.55 The absence of EOG responses indicated that CNGA2 KO mice are anosmic. CNGA2 KO mice struggle to survive because they are not even able to feed during nursing and weaning. Since that initial report, residual responses to some odorants have been demonstrated in mice lacking CNGA2.56,57
Ca2+-activated Cl− currents in olfactory cilia
The odorant-induced current is made of 2 components: a cationic one activated by cAMP and an anionic one activated by Ca2+. This could be demonstrated by excising olfactory cilia and exposing them to cAMP (Fig. 2C). Only at negative holding potentials, which favors Ca2+ influx, was a bi-phasic current observed with the second component being absent in Cl−-free solution (Fig. 2D). The latter current, a Cl− current, is gated directly by the Ca2+ that entered through the CNG channels. Taking advantage of flash photolysis Boccaccio and Menini showed that upon cAMP uncaging in intact OSNs a clearly biphasic current developed at a holding potential of −50 mV (Fig. 2E & F).19,58 The first and fastest step reached a plateau that in some cases lasted several ms and was then followed by a larger component that was absent at +50 mV when, again only little Ca2+ entered through the CNG channel not sufficient to activate the Cl− current. Moreover, the second component was strongly reduced by lowering the intracellular Cl− concentration and it was blocked by niflumic acid (Fig. 2E and F, gray traces), a commonly used Cl− channel blocker. Single cell electrophysiology of OSNs indicated that the Ca2+-activated Cl− currents constitute up to 90% of the receptor current in rodents. The relative magnitude of the Cl− compared with that of the CNG current is probably due to a higher density of CaCC in the olfactory cilia,5 even though this phenomenon and the estimated density of CaCC could be species-specific. Despite lacking its molecular counterpart at the time, the biophysical characteristics of the current have been extensively studied (see below).
Although the high input resistance of OSNs could allow even a small depolarization - stemming only from CNG channels - to fire APs, the larger contribution of the Cl− current to the transduction current serves as a non-linear amplifier of the signal. Lowe and Gold showed in rat OSNs that the Ca2+-activated Cl− current boosts the cooperativity of the odorant-induced current.18 While in excised patch experiments the apparent cooperativity of the cAMP-induced current is about 1.5,59 in intact OSNs, with the presence of Ca2+-activated Cl− current, the cooperativity reaches much higher values, between 3.5 and 5.4,18,21 clearly indicating that the presence of a secondary Ca2+-activated Cl− current increases the nonlinear amplification of the receptor current. Such nonlinear activation of the Cl− current depends on the Ca2+ cooperativity of the conductance. Excised patch experiments from OSN cilia/knob show that the dose–response for Ca2+ was well fitted by the Hill equation with half-maximal activation between 2.2 and 4.7 μM and Hill coefficient between 2.0 and 2.8,5 sufficiently steep to support the high odorant-induced cooperativity observed in OSNs.
Moreover, Ca2+-activated Cl− currents show a Ca2+-dependent inactivation, which is reversible after removal of Ca2+, but also an irreversible run-down when recorded from excised patches, indicating that some modulatory components of the channel may be lost after the excision of the membrane.5,17 The olfactory Ca2+-activated Cl− current is apparently not directly affected by Ca2+–calmodulin5 and, at present, no modulators of channel activity are known. Another recently described property of the native channel is that upon flash photolysis of caged Ca2+, the reversal potential for some external large anions changes with time,60 a phenomenon which requires further mechanistic and physiologic studies.
Before the molecular identification of the Cl− channel, the only genetic approach available to alter or investigate the excitatory role of the Ca2+-activated Cl− currents was by using a knockout mouse for the Na+-K+-2Cl− cotransporter NKCC1.17 As mentioned earlier, NKCC1 provides for active accumulation of the intracellular ciliary Cl− that reaches a concentration close to that of the embedding mucus.16 Thus, by knocking out NKCC1, Cl− could not accumulate in the cilia, changing the Cl− reversal potential, and the Ca2+-activated Cl− component of the odorant response was largely reduced, leaving only 20% of the total odorant response. The Cl− channel, though, was still present when excised patch experiments were performed. In summary, although still elusive in its identity, the native CaCC was fully biophysically characterized and hypotheses about its functionality were possible.
Hunting for the Ca2+-activated Cl− channel
The elusive molecular nature of Ca2+-activated Cl− currents was an issue not only for olfaction but for other fields as well. Different Ca2+-activated Cl− currents have been characterized biophysically in various tissues and cell types without establishing their molecular identity.62-66
In the OE, the first molecular candidate to be proposed was ClCa1 cloned from the rat olfactory mucosa in 2003 ,67 which was then followed by another candidate, Bestrophin2 (Best2).59
Best2, which belongs to the family of Bestrophin channels, was at the time a promising candidate because of several properties: it was cloned from mouse OE, was expressed in the ciliary layer of the OE and co-localized with the CNGA2 channel. Best2's biophysical properties were very similar to those of the native channel (anion selectivity, unitary conductance, voltage dependences, sensitivity to blockers).59 But there was one obvious difference: Best2 is about 12 times more sensitive to Ca2+ than the native olfactory CaCC. Although such differences could simply be due to the absence of additional subunits or modulators in the expression system, it called for caution in concluding that Best2 was the native olfactory channel. Overall, it was reasonable to propose that Best2 might be the olfactory CaCC—or at least a part of it—but later it was shown that mice lacking Best2 still had the Ca2+-activated Cl− currents and displayed normal olfactory behavior.68 In conclusion, these results eliminated a promising candidate, but sent the search for the molecular identity of the olfactory CaCC back to the beginning.
TMEM16B is the cilial Ca2+-activated Cl− channel
Help came from an unexpected side in 2008, when 3 groups using different approaches found that members of the TMEM16 family, a group of proteins with largely unclear functions, encoded for CaCCs. It was proposed to change the name of TMEM16 to Anoctamin because of the anion permeability and bioinformatics analysis predicted that they have 8 transmembrane domains (octamins). The appropriateness of the name has been actively discussed, in particular, because it is possible that only 2 members of this family are Cl− channels and they might actually have 10, not 8, transmembrane domains.69-72
But was one of the newly discovered TMEM16 family members the native olfactory channel? The answer came soon after when anoctamin2/TMEM16B was biophysically characterized in axolotl oocytes and HEK 293 cells.8,60,73,74
Not only was it a CaCC but also most of its properties resembled those of the native olfactory channel.8,60,73-75 In addition, TMEM16B was found to be a prominent protein in the olfactory ciliary proteome with a relative abundance of TMEM16B to CNGA2,75,76 in agreement with electrophysiological data showing that CaCCs have a greater density than CNG channels. Anecdotally, in 2005, while trying to identify markers for different populations of cell types in the OE, it was shown that TMEM16B transcripts were abundantly localized in mature OSNs; unfortunately, at the time nobody could tell that TMEM16B was a CaCC.77 When, via adenoviral infections, GFP-tagged TMEM16B was expressed in OSNs, it trafficked to the ciliary membrane,75 and immunohistochemical experiments showed that it was localized in the ciliary layer of the OE (Fig. 3A).60,78,79
Figure 3.
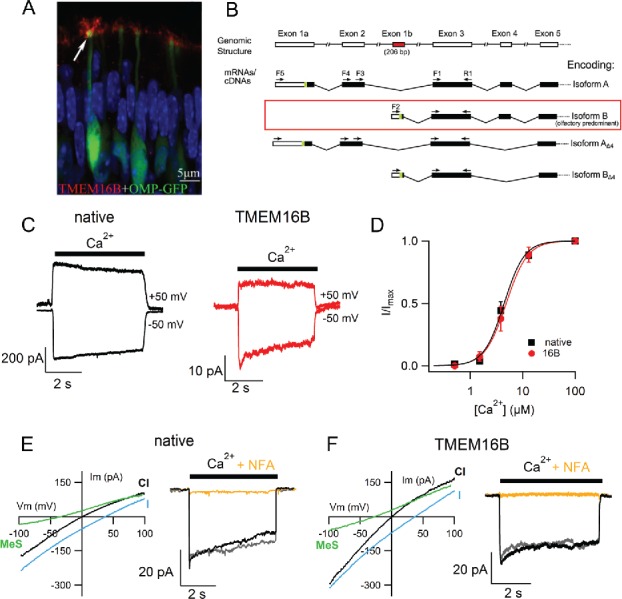
TMEM16B is expressed in the OE and a side by side comparison with the olfactory native channel revealed surprisingly similar features. Immunofluorescence experiments revealed that TMEM16B is expressed in the cilia of OSNs (A). Modified from Maurya and Menini 2014 with permission.82 Its gene has 4 splice variants. (B) Diagram summarizes the 5’ TMEM16B exon splicing structure. The green sections indicate the most 5’ AUG translation start codons, and the subsequent black bars indicate predicted protein-coding sequence. The variants containing exons 1a and 2 are less abundant in the olfactory epithelium than the variants containing the previously unknown exon 1b. From Saidu et al. 2013 originally published in the Journal of General Physiology.85 (C) Currents activated by 100 μM Ca2+ at −50 or +50 mV. (D) Normalized currents measured at −50 mV were plotted versus Ca2+ concentrations and fitted with the Hill equation. (E and F) Current–voltage relations from a ramp protocol activated by 100 μM Ca2+. Bath solutions contained 140 mM NaCl, or the sodium salt of methanesulfonate (MeS, green trace) or of the more permeable ion Iodide (I, cyan trace), as indicated, showing a similar shift in the reversal potential. Intracellular blockage by 300 μM niflumic acid (NFA, orange trace) of the current activated by 100 μM Ca2+ at −50 mV. Modified from Pifferi et al. 2006b,59 copyright (2006) National Academy of Sciences, USA, and from Pifferi et al. 2009a.73
Several other members of the TMEM16 family (16A, 16F, 16H, 16K) were found in the OE.60,79-81 Only—TMEM16A and TMEM16F—were further studied by immunofluorescence and only the latter was shown to be expressed in the cilia of the OSNs,80 while TMEM16A was expressed in some supporting cells of the OE.82,83 Whether TMEM16F is part of the native channel requires further studies.80,84
Four different splice variants of TMEM16B are expressed in the OE (Fig. 3B). Two variants (isoform B, the most abundant, and isoform BΔ4) have the transcription start site in a newly identified exon (exon 1b) therefore encoding for a shorter N-terminal region than TMEM16B in the retina, whereas the other 2, low-abundance variants (isoform A and AΔ4) have the canonical start site. Interestingly, in 2 isoforms (AΔ4 and BΔ4) exon 4 is spliced out and they are not able to form functional channels if expressed alone in a heterologous system.85
A side-by-side comparison of the functional properties measured in excised inside-out patches from the native olfactory Cl− channel and the TMEM16B-induced current in HEK 293 cells showed remarkable similarities (Fig. 3C - F). Both channels showed Ca2+ activated currents (Fig. 3C) with dose–response relations having half-maximal concentration of Ca2+ of 4.9 μM for TMEM16B very similar to 4.7 μM for native channels at −50 mV (Fig. 3D). Moreover, both channels have the same anion selectivity: they are more permeable to anions larger than Cl− and have little permeability for methanesulfonate (Fig. 3E–F). In addition, both TMEM16B and native CaCCs are similarly blocked by niflumic acid (Fig. 3G–I). Moreover, TMEM16B displays a Ca2+-dependent inactivation and an irreversible run-down similar to native olfactory CaCCs.5,73,75,85 Co-expression of the 2 splice variants B and BΔ4 gives rise to a channel that reproduces the native voltage-dependent inactivation at positive holding potentials better than isoform B alone. Thus, TMEM16B is an important and required component of the olfactory Cl− channel that has been confirmed by several laboratories independently.86
Finally, the TMEM16B knockout
Since the identification of TMEM16B expression in the OE, the knockout (KO) mouse model was in the air: it took 3 y to have the first mouse model where the TMEM16B gene was knocked out. In 2011 Billig et al.81 published the TMEM16B KO mouse model in a paper with an unexpected title: “Ca2+-activated Cl− currents are dispensable for olfaction.” It was shown that the Ca2+-activated Cl− current was entirely absent in OSNs from TMEM16B KO mice, confirming that TMEM16B is a mandatory part of the Cl− channel. However, KO mice performed indistinguishably from WT mice in an olfactory-guided Go/No Go task, suggesting that Ca2+-activated Cl− currents are not relevant for near normal olfactory behavior. This result was truly puzzling, considering that the Cl− channel is expressed at high levels at a significant metabolic cost and evolutionarily conserved across many species. Would this channel only have a merely minor accessory role or none at all?
Using the same animal model (kindly provided by Prof. Jentsch), Pietra et al. (2016)87 re-examined the question from a slightly different perspective, starting with a naïve behavioral task, the “finding the buried food test” which, unlike the Go/NoGo task, does not involve any animal training and pre-learning. Indeed, from the perspective of the mouse, the Go/NoGo task is an artificial operant conditioning experiment for which the mouse has to be extensively trained and be exposed to the same odorant repeatedly. Our approach, instead, involved odor-guided food-seeking where mice must use their olfactory ability to locate a food item buried under the bedding chips in the cage. This task did not require any learning sessions, making it a very simple and naïve task that lets the animals rely more on their innate olfactory abilities. TMEM16B KO mice were accomplishing the task significantly slower than wild type mice. Interestingly, when mice were tested once a day for several days, KO mice progressively caught up with WT mice and, after 2–3 d of testing, they performed equally well. This suggests that once KO mice “learned” the identity of the odor, they could perform equally well (Fig. 4A and B). If we challenged the KO with yet another novel odor, TMEM16B KO again required longer times to locate the food compared with WT mice, whereas for odors already known to the mice, e.g., retesting with the same food or using their standard chow, their performance was similar to WT (Fig. 4C). Thus our different behavioral tests gave different results compared with Billig et al.,81 although their tests aimed at detecting different behavioral aspects. Given that we found that KO mice required longer times to identify new odorants, it would be interesting to investigate how, in a Go/No Go paradigm, mice trade off accuracy for odorant-sampling duration (meaning the KO mice might sample for a longer period to achieve the same accuracy), as WT mice do.81
Figure 4.

Behavioral impairment in the TMEM16B KO mice. In “find the buried cookie test” (A) a piece of Oreo cookie was buried under the cage bedding and the mouse when introduced into the cage was free to explore to find the cookie. TMEM16B KO mice were slower initially compared with WT but over the following days they improved and accomplished the task as fast as the WT (B, compare day 1 with day 4). When a new food item was buried the TMEM16B KO mice were again slower than the WT (B and C). Modified from Pietra et al. 2016 originally published in the Journal of General Physiology with permission.87
Clearly, though, in comparison to CNGA2 KO or AC3 KO mice, which are anosmic, TMEM16B KO are not anosmic. But CaCCs are not simply dispensable for all olfactory tasks, they may be dispensable for the kind of tasks which require extensive learning and training. Therefore, a broader battery of behavioral tests is required to delineate those differences. Also, as afore mentioned TMEM16B is expressed in other cell types 62-66 and in particular in brain structures that may be linked to the olfactory systems.88-90
Thus, a future approach to better dissect out the exact contribute of TMEM16B to the olfactory system may require conditional KO model where this channel is deleted in a population of mature OSNs.
TMEM16B involvement in odorant-induced action potential firing
The transduction current drives a generator potential that quickly reaches the threshold for AP firing to be conveyed to the OB. Since in OSNs, relatively small neurons with a high input resistance,91 CNG currents alone are sufficient to generate APs, what is the role of the CaCC? If in OSNs CNG currents alone are sufficient to generate APs,18,91 what is then the role of the Ca2+-activated Cl− currents?
Usually, APs in OSNs are generated at the very onset of an odorant response and by the end of the rising phase of the transduction current, their spike amplitude has collapsed into the noise of the recording. This is most likely due to the progressive and ultimately complete inactivation of voltage-gated Na+ channels during strong and sustained depolarization.92 In addition, the presence of a rapidly inactivating K+ conductance that recovers only slowly from inactivation has been suggested to contribute to the generation of short bursts of spike firing.93
While the transduction current saturates far above the odorant threshold, the number of APs fired increases monotonically with stimulus concentration only up to half maximal odorant concentrations, then showing a decreasing number of AP (typically 1–2) at higher concentrations.24,94-96
In other words, the number of APs saturates well before that the transduction current saturates.
In response to IBMX (a phosphodiesterase blocker used as surrogate for odorant stimulation) or to an odorant, OSNs fire a train of few APs (Fig. 5A and C for WT) with a relatively short duration (Fig. 5F, black). However, when TMEM16B is absent the number of APs in the train increased and so did its duration (Fig. 5B and D for KO). The number of APs in the KO increased about 10-fold and the AP train duration about 3-fold (Fig. 5E and F). In conclusion, the lack of TMEM16B, and thus of any Cl− component, affects the firing behavior by prolonging the AP train duration and increasing the number of APs in response to a stimulus.
Figure 5.
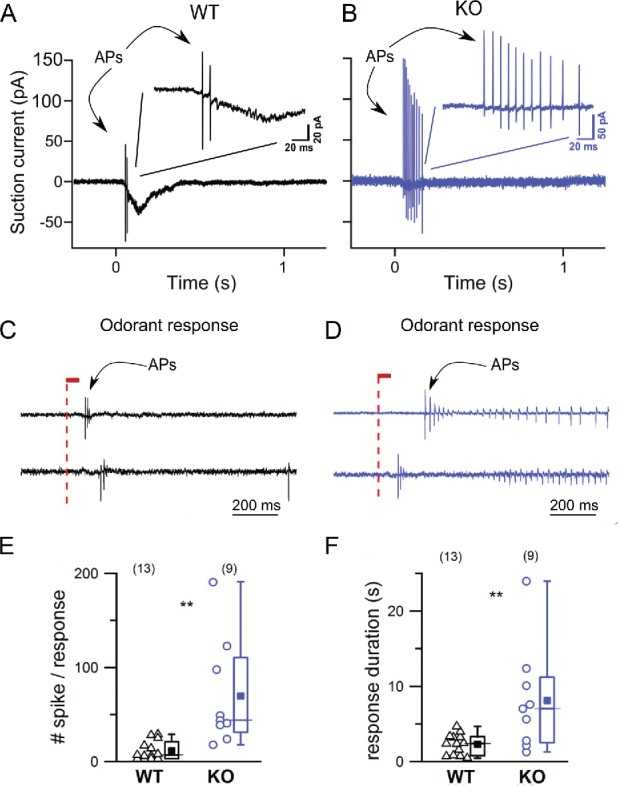
TMEM16 role in activity dependent firing. (A-B) IBMX responses recorded by using the suction electrode technique from isolated OSNs.98 OSNs from WT mice typically fired 2–3 APs in response to 1 mM IBMX (1 s stimulation) while the KO fired a more prolonged AP train. This finding was confirmed using a different approach: (C-D) loose patch recordings from the dendritic knob of OSNs in the intact epithelium (expressing the I7 OR stimulated with its ligand heptanal). OSNs from KO mice not only fired more APs (E) but also the AP train duration was prolonged (F). Modified from Pietra et al. 2016 originally published in the Journal of General Physiology with permission.87
Therefore, the depolarizing action of CaCCs, although somewhat counterintuitive, is that of shortening AP firing by tuning it with the odorant strength, driving in this way the time course of the response that causes quick inactivation of voltage dependent Na+ channels. Both the number of APs and AP train duration might be required for the appropriate flow of information that ultimately is sent to the brain to correctly signal for the right odorant concentration and stimulus duration.94,97
TMEM16B involvement in spontaneous action potential firing
Beginning with the early electrophysiological characterization of OSNs, spontaneous electrical activity in the form of basal spiking was described. OSNs showed basal membrane voltage fluctuations which are able to reach the threshold for spontaneous APs firing. This “noise” is caused by cAMP fluctuations generated by the spontaneous activity of an OR which in turn drives G protein and AC3 activation to open the CNG channel and then TMEM16B.99 Interestingly, different ORs have different constitutive activities that drive cAMP fluctuations, therefore spontaneous firing depends on the OR that an OSN expresses.99,100
An interesting case is that of the I7 OR that has been extensively investigated for its pharmacological profile;101,102 it has a very high basal activity resulting in a higher spontaneous firing rate when compared with much quieter ORs (i.e. mOR-Eg). When we investigated the spontaneous firing rate of I7 OSNs lacking the TMEM16B channels, their spontaneous firing dramatically decreased (Fig. 6). These results along with the evidence that spontaneous APs firing is completely and reversibly blocked by applying the CaCC blocker niflumic acid demonstrate that even at very low levels of activity, the CaCC is what drives the AP output.99
Figure 6.
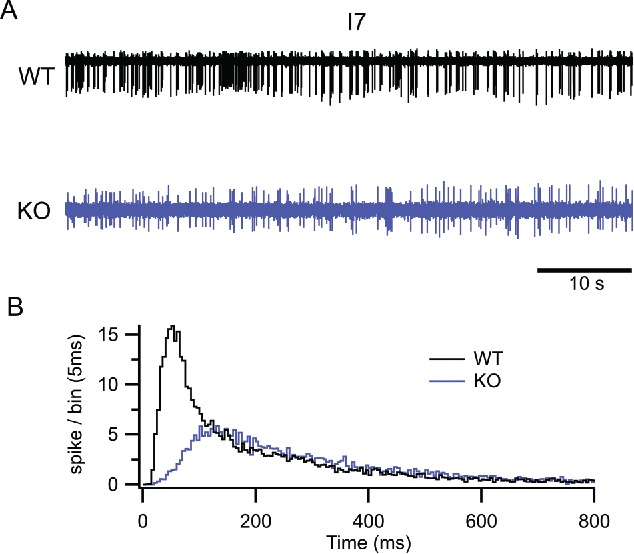
The lack of TMEM16B reduces the spontaneous firing in I7-expressing OSNs. (A) Loose-patch recordings (60-s recordings) of the spontaneous firing activity from WT (top) and KO mice (bottom) OSNs expressing the I7 OR. (B) Interspike interval (ISI) distribution histograms (bin = 5 ms) normalized to the number of recorded neurons in control (black line) and KO mice (blue line) shows that brief ISIs were missing in TMEM16B KO OSNs compared with WT and that their distribution was shifted to longer ISIs. Modified from Pietra et al. 2016 originally published in the Journal of General Physiology with permission.87
In conclusion, while for odorant responses TMEM16B is involved in controlling the spike train duration that may encode stimulus strength, for the spontaneous activity of an OSN expressing an OR with high basal activity, the CaCC provides the depolarizing current that amplifies the small CNG currents. OSNs expressing other ORs with different basal activity might be affected differently by the lack of TMEM16B, though. Forgotten for a long time, we think that spontaneous activity is an emerging and interesting aspect of the olfactory system and TMEM16B involvement.
TMEM16B-mediated electrical activity has a role in glomerular targeting in the OB
Basal activity of ORs and basal noise could be considered as an epiphenomenon of a system that expresses 1000 different G protein-coupled receptors, but recent evidence suggests that spontaneous activity could be more than just noise since, for example, spontaneously generated levels of cAMP could set the threshold above which a stimulus could give rise to a meaningful response. Additionally, basal activity has been implicated in targeting of OSN axons to the OB.
In sensory physiology, the notion that “neurons that fire together wire together” is used to describe that spiking can determine the fate of the projecting axons to higher areas of the brain. In the olfactory system, this notion has been complicated by data showing that the “biochemical intrinsic spontaneous activity” (as supposed to electrical activity) is likely to be involved in the control of the glomerular formation and targeting.103,104
The process of OR selection during OSN development is crucial and determines the fate of these neurons, not only for their response profile but also for the targeting of their axons to the bulb. Each OSN projects to a neuropil-like structure called glomerulus, which groups together axons from OSNs expressing the same OR. There are typically 2 glomeruli per OR, one on the lateral and one on the medial side of the OB. The OR identity regulates the expression levels of many molecules involved in glomerular positioning and targeting and the development of the olfactory map is largely target-independent. The guidance molecule implicated in anterior-posterior targeting was proposed to be Neuropilin1 (Nrp1) and recent evidence showed that OR spontaneous activity (and not ligand-induced activity), probably via the cAMP-PKA pathway,105-107 affects its expression level. Genetically swapping an OR of low basal activity with a high activity OR in a given OSN caused a posterior shift of the glomeruli. This model, though, has been questioned and requires further studies,108-110 but the idea of having an OR-dependent regulation of the expression of guidance molecules involved in OB targeting and glomerular formation seems to hold because of the mosaic pattern of expression of such molecules in the OB (i.e., Nrp1).94,108,110 Once the axons of OSNs expressing the same ORs converge in a general area, ligand-dependent activity determines the expression of molecules involved in axon sorting into individual glomeruli.
That the spontaneous AP firing could be important for glomerular formation is also supported by results from the TMEM16B KO mouse: additional, supernumerary glomeruli were observed for I7-expressing OSNs. The number of glomeruli in the OB onto which I7-expressing OSNs converge almost doubled in the TMEM16B KO mice.87 As the expression of I7 ORs confers to OSNs a high rate of spontaneous firing, I7-expressing OSNs are more sensitive to the reduction of the firing rate as is the case in the TMEM16B KO, while OSNs expressing other more silent ORs might not be affected.87,81 Indeed, this might be the case in the 2 other ORs, M72 and P2, for which glomerular targeting has also been studied in TMEM16B KO mice, which appeared to target normally to the OB.81 Thus, TMEM16B could play an important role to enable correct glomerular targeting, and the role of spontaneous firing in targeting needs to be carefully reconsidered. The TMEM16B KO mouse model could be a suitable tool to accomplish the task.
Conclusions and perspectives
The tale of the olfactory CaCC continues. It is like a journey, and the destination is to determine its role in OSNs and in the olfactory system. Since identifying its molecular identity, and hence being able to knock it out, TMEM16B has been shown to control the duration of OSN firing in response to a stimulus, with KO OSNs displaying a prolonged spike train and also axon mis-targeting to the OB. Alteration in AP firing alters the input to glomeruli in the OB where changes in temporal patterns could cause a change in how odor quality and intensity are encoded, hence leading to problems in performing naïve behavioral tasks and recognizing novel odors. Changes in basal and evoked activity could also alter glomerular connectivity to second order neurons in the bulb, such as interneurons and mitral/tufted cells. Thus, the Cl− channel in OSNs might have multiple and complex effects on the physiology of OSNs themselves, their connectivity to higher order neurons and how they contribute to olfactory coding of odorants. This might be only the beginning of the journey to understand this complexity.
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed
Funding
Rita Levi Montalcini Award from Italian Ministry of Education, University, and Research (DM100915_685 to Michele Dibattista), NIH (DC009613 to Johannes Reisert), Fondazione Compagnia di San Paolo, Torino (2013.0922 to Anna Boccaccio), Italian Ministry of Education, University, and Research (2010599KBR to Anna Menini)
References
- [1].Axel R., Buck L. A novel multigene family may encode odorant receptors: A molecular basis for odor recognition. Cell 1991; 65:175-87; PMID:1840504; http://dx.doi.org/ 10.1016/0092-8674(91)90418-X [DOI] [PubMed] [Google Scholar]
- [2].Kleene SJ, Gesteland RC. Calcium-activated chloride conductance in frog olfactory cilia. J Neurosci 1991; 11:3624-9; PMID:1941099 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [3].Kurahashi T, Yau KW. Co-existence of cationic and chloride components in odorant-induced current of vertebrate olfactory receptor cells. Nature 1993; 363:71-4; PMID:7683113; http://dx.doi.org/ 10.1038/363071a0 [DOI] [PubMed] [Google Scholar]
- [4].Zhainazarov AB, Ache BW. Odor-induced currents in Xenopus olfactory receptor cells measured with perforated-patch recording. J Neurophysiol 1995; 74:479-83; PMID:7472351 [DOI] [PubMed] [Google Scholar]
- [5].Reisert J, Bauer PJ, Yau KW, Frings S. The Ca-activated Cl channel and its control in rat olfactory receptor neurons. J Gen Physiol 2003; 122:349-63; PMID:12939394; http://dx.doi.org/ 10.1085/jgp.200308888 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [6].Sato K, Suzuki N. The contribution of a Ca(2+)-activated Cl(−) conductance to amino-acid-induced inward current responses of ciliated olfactory neurons of the rainbow trout. J Exp Biol 2000; 203:253-62; PMID:10607535 [DOI] [PubMed] [Google Scholar]
- [7].Caputo A, Caci E, Ferrera L, Pedemonte N, Barsanti C, Sondo E, Pfeffer U, Ravazzolo R, Zegarra-Moran O, Galietta LJ. TMEM16A, a membrane protein associated with calcium-dependent chloride channel activity. Science 2008; 322:590-4; PMID:18772398; http://dx.doi.org/ 10.1126/science.1163518 [DOI] [PubMed] [Google Scholar]
- [8].Schroeder BC, Cheng T, Jan YN, Jan LY. Expression cloning of TMEM16A as a calcium-activated chloride channel subunit. Cell 2008; 134:1019-29; PMID:18805094; http://dx.doi.org/ 10.1016/j.cell.2008.09.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [9].Yang YD, Cho H, Koo JY, Tak MH, Cho Y, Shim WS, Park SP, Lee J, Lee B, Kim BM, et al.. TMEM16A confers receptor-activated calcium-dependent chloride conductance. Nature 2008; 455:1210-5; PMID:18724360; http://dx.doi.org/ 10.1038/nature07313 [DOI] [PubMed] [Google Scholar]
- [10].Malnic B, Hirono J, Sato T, Buck LB. Combinatorial receptor codes for odors. Cell 1999; 96:713-23; PMID:10089886; http://dx.doi.org/ 10.1016/S0092-8674(00)80581-4 [DOI] [PubMed] [Google Scholar]
- [11].Jones DT, Reed RR. Golf: an olfactory neuron specific-G protein involved in odorant signal transduction. Science 1989; 244:790-5; PMID:2499043; http://dx.doi.org/ 10.1126/science.2499043 [DOI] [PubMed] [Google Scholar]
- [12].Bakalyar HA, Reed RR. Identification of a specialized adenylyl cyclase that may mediate odorant detection. Science 1990; 250:1403-6; PMID:2255909; http://dx.doi.org/ 10.1126/science.2255909 [DOI] [PubMed] [Google Scholar]
- [13].Nakamura T, Gold GH. A cyclic nucleotide-gated conductance in olfactory receptor cilia. Nature 1987; 325:442-4; PMID:3027574; http://dx.doi.org/ 10.1038/325442a0 [DOI] [PubMed] [Google Scholar]
- [14].Firestein S, Zufall F, Shepherd GM. Single odor-sensitive channels in olfactory receptor neurons are also gated by cyclic nucleotides. J Neurosci 1991; 11:3565-72; PMID:1719165 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [15].Kaneko H, Putzier I, Frings S, Kaupp UB, Gensch T. Chloride accumulation in mammalian olfactory sensory neurons. J Neurosci 2004; 24:7931-8; PMID:15356206; http://dx.doi.org/ 10.1523/JNEUROSCI.2115-04.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [16].Reuter D, Zierold K, Schröder WH, Frings S. A depolarizing chloride current contributes to chemoelectrical transduction in olfactory sensory neurons in Situ. J Neurosci 1998; 18:6623-30; PMID:9712634 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [17].Reisert J, Lai J, Yau KW, Bradley J. Mechanism of the excitatory Cl− response in mouse olfactory receptor neurons. Neuron 2005; 45:553-61; PMID:15721241; http://dx.doi.org/ 10.1016/j.neuron.2005.01.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [18].Lowe G, Gold GH. Nonlinear amplification by calcium-dependent chloride channels in olfactory receptor cells. Nature 1993; 366:283-6; PMID:8232590; http://dx.doi.org/ 10.1038/366283a0 [DOI] [PubMed] [Google Scholar]
- [19].Boccaccio A, Menini A. Temporal development of cyclic nucleotide-gated and Ca2+-activated Cl− currents in isolated mouse olfactory sensory neurons. J Neurophysiol 2007; 98:153-60; PMID:17460108; http://dx.doi.org/ 10.1152/jn.00270.2007 [DOI] [PubMed] [Google Scholar]
- [20].Cygnar KD, Zhao H. Phosphodiesterase 1C is dispensable for rapid response termination of olfactory sensory neurons. Nat Neurosci 2009; 12:454-62; PMID:19305400; http://dx.doi.org/ 10.1038/nn.2289 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [21].Boccaccio A, Lagostena L, Hagen V, Menini A. Fast Adaptation in Mouse Olfactory Sensory Neurons Does Not Require the Activity of Phosphodiesterase. J Gen Physiol 2006; 128:171-84; PMID:16880265; http://dx.doi.org/ 10.1085/jgp.200609555 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [22].Flannery RJ, French DA, Kleene SJ. Clustering of cyclic-nucleotide-gated channels in olfactory cilia. Biophys J 2006; 91:179-88; PMID:16603488; http://dx.doi.org/ 10.1529/biophysj.105.079046 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [23].Chen C, Nakamura T, Koutalos Y. Cyclic AMP diffusion coefficient in frog olfactory cilia. Biophys J 1999; 76:2861-7; PMID:10233102; http://dx.doi.org/ 10.1016/S0006-3495(99)77440-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [24].Reisert J, Matthews HR. Response properties of isolated mouse olfactory receptor cells. J Physiol 2001; 530:113-22; PMID:11136863; http://dx.doi.org/ 10.1111/j.1469-7793.2001.0113m.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- [25].Antolin S, Matthews HR. The effect of external sodium concentration on sodium–calcium exchange in frog olfactory receptor cells. J Physiol 2007; 581:495-503; PMID:17379630; http://dx.doi.org/ 10.1113/jphysiol.2007.131094 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [26].Reisert J, Matthews HR. Na+-dependent Ca2+ Extrusion Governs Response Recovery in Frog Olfactory Receptor Cells. J Gen Physiol 1998; 112:529-35; PMID:9806962; http://dx.doi.org/ 10.1085/jgp.112.5.529 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [27].Pyrski M, Koo JH, Polumuri SK, Ruknudin AM, Margolis JW, Schulze DH, Margolis FL. Sodium/calcium exchanger expression in the mouse and rat olfactory systems. J Comp Neurol 2007; 501:944-58; PMID:17311327; http://dx.doi.org/ 10.1002/cne.21290 [DOI] [PubMed] [Google Scholar]
- [28].Stephan AB, Tobochnik S, Dibattista M, Wall CM, Reisert J, Zhao H. The Na(+)/Ca(2+) exchanger NCKX4 governs termination and adaptation of the mammalian olfactory response. Nat Neurosci 2012; 15:131-7; http://dx.doi.org/ 10.1038/nn.2943 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [29].Ferguson CH, Zhao H. Simultaneous Loss of NCKX4 and CNG Channel Desensitization Impairs Olfactory Sensitivity. J Neurosci Off J Soc Neurosci 2017; 37:110-9; http://dx.doi.org/ 10.1523/JNEUROSCI.2527-16.2017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [30].Fluegge D, Moeller LM, Cichy A, Gorin M, Weth A, Veitinger S, Cainarca S, Lohmer S, Corazza S, Neuhaus EM, et al.. Mitochondrial Ca(2+) mobilization is a key element in olfactory signaling. Nat Neurosci 2012; 15:754-62; PMID:22446879; http://dx.doi.org/ 10.1038/nn.3074 [DOI] [PubMed] [Google Scholar]
- [31].Bradley J, Li J, Davidson N, Lester HA, Zinn K. Heteromeric olfactory cyclic nucleotide-gated channels: a subunit that confers increased sensitivity to cAMP. Proc Natl Acad Sci U S A 1994; 91:8890-4; PMID:7522325; http://dx.doi.org/ 10.1073/pnas.91.19.8890 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [32].Liman ER, Buck LB. A second subunit of the olfactory cyclic nucleotide-gated channel confers high sensitivity to cAMP. Neuron 1994; 13:611-21; PMID:7522482; http://dx.doi.org/ 10.1016/0896-6273(94)90029-9 [DOI] [PubMed] [Google Scholar]
- [33].Sautter A, Zong X, Hofmann F, Biel M. An isoform of the rod photoreceptor cyclic nucleotide-gated channel beta subunit expressed in olfactory neurons. Proc Natl Acad Sci U S A 1998; 95:4696-701; PMID:9539801; http://dx.doi.org/ 10.1073/pnas.95.8.4696 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [34].Bönigk W, Bradley J, Müller F, Sesti F, Boekhoff I, Ronnett GV, Kaupp UB, Frings S. The native rat olfactory cyclic nucleotide-gated channel is composed of three distinct subunits. J Neurosci 1999; 19:5332-47; PMID:10377344 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [35].Zheng J, Zagotta WN. Stoichiometry and assembly of olfactory cyclic nucleotide-gated channels. Neuron 2004; 42:411-21; PMID:15134638; http://dx.doi.org/ 10.1016/S0896-6273(04)00253-3 [DOI] [PubMed] [Google Scholar]
- [36].Dhallan RS, Yau KW, Schrader KA, Reed RR. Primary structure and functional expression of a cyclic nucleotide-activated channel from olfactory neurons. Nature 1990; 347:184-7; PMID:1697649; http://dx.doi.org/ 10.1038/347184a0 [DOI] [PubMed] [Google Scholar]
- [37].Ludwig J, Margalit T, Eismann E, Lancet D, Kaupp UB. Primary structure of cAMP-gated channel from bovine olfactory epithelium. FEBS Lett 1990; 270:24-9; PMID:1699793; http://dx.doi.org/ 10.1016/0014-5793(90)81226-E [DOI] [PubMed] [Google Scholar]
- [38].Frings S, Lynch JW, Lindemann B. Properties of cyclic nucleotide-gated channels mediating olfactory transduction. Activation, selectivity, and blockage. J Gen Physiol 1992; 100:45-67; http://dx.doi.org/ 10.1085/jgp.100.1.45 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [39].Kurahashi T, Kaneko A. Gating properties of the cAMP-gated channel in toad olfactory receptor cells. J Physiol 1993; 466:287-302; PMID:8410695 [PMC free article] [PubMed] [Google Scholar]
- [40].Nache V, Schulz E, Zimmer T, Kusch J, Biskup C, Koopmann R, Hagen V, Benndorf K. Activation of olfactory-type cyclic nucleotide-gated channels is highly cooperative. J Physiol 2005; 569:91-102; PMID:16081488; http://dx.doi.org/ 10.1113/jphysiol.2005.092304 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [41].Biskup C, Kusch J, Schulz E, Nache V, Schwede F, Lehmann F, Hagen V, Benndorf K. Relating ligand binding to activation gating in CNGA2 channels. Nature 2007; 446:440-3; PMID:17322905; http://dx.doi.org/ 10.1038/nature05596 [DOI] [PubMed] [Google Scholar]
- [42].Larsson HP, Kleene SJ, Lecar H. Noise analysis of ion channels in non-space-clamped cables: estimates of channel parameters in olfactory cilia. Biophys J 1997; 72:1193-203; PMID:9138566; http://dx.doi.org/ 10.1016/S0006-3495(97)78767-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [43].Kleene SJ. High-gain, low-noise amplification in olfactory transduction. Biophys J 1997; 73:1110-7; PMID:9251827; http://dx.doi.org/ 10.1016/S0006-3495(97)78143-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [44].Kramer RH, Siegelbaum SA. Intracellular Ca2+ regulates the sensitivity of cyclic nucleotide-gated channels in olfactory receptor neurons. Neuron 1992; 9:897-906; PMID:1384576; http://dx.doi.org/ 10.1016/0896-6273(92)90242-6 [DOI] [PubMed] [Google Scholar]
- [45].Zufall F, Firestein S, Shepherd GM. Analysis of single cyclic nucleotide-gated channels in olfactory receptor cells. J Neurosci 1991; 11:3573-80; PMID:1719166 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [46].Dzeja C, Hagen V, Kaupp UB, Frings S. Ca2+ permeation in cyclic nucleotide-gated channels. EMBO J 1999; 18:131-44; PMID:9878057; http://dx.doi.org/ 10.1093/emboj/18.1.131 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [47].Pifferi S, Boccaccio A, Menini A. Cyclic nucleotide-gated ion channels in sensory transduction. FEBS Lett 2006; 580:2853-9; PMID:16631748; http://dx.doi.org/ 10.1016/j.febslet.2006.03.086 [DOI] [PubMed] [Google Scholar]
- [48].Reisert J, Zhao H. Response kinetics of olfactory receptor neurons and the implications in olfactory coding. J Gen Physiol 2011; 138:303-10; PMID:21875979; http://dx.doi.org/ 10.1085/jgp.201110645 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [49].Song Y, Cygnar KD, Sagdullaev B, Valley M, Hirsh S, Stephan A, Reisert J, Zhao H. Olfactory CNG channel desensitization by Ca2+/CaM via the B1b subunit affects response termination but not sensitivity to recurring stimulation. Neuron 2008; 58:374-86; PMID:18466748; http://dx.doi.org/ 10.1016/j.neuron.2008.02.029 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [50].Menini A. Calcium signalling and regulation in olfactory neurons. Curr Opin Neurobiol 1999; 9:419-26; PMID:10448159; http://dx.doi.org/ 10.1016/S0959-4388(99)80063-4 [DOI] [PubMed] [Google Scholar]
- [51].Kurahashi T, Menini A. Mechanism of odorant adaptation in the olfactory receptor cell. Nature 1997; 385:725-9; PMID:9034189; http://dx.doi.org/ 10.1038/385725a0 [DOI] [PubMed] [Google Scholar]
- [52].De Palo G, Boccaccio A, Miri A, Menini A, Altafini C. A dynamical feedback model for adaptation in the olfactory transduction pathway. Biophys J 2012; 102:2677-86; PMID:22735517; http://dx.doi.org/ 10.1016/j.bpj.2012.04.040 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [53].Cygnar KD, Stephan AB, Zhao H. Analyzing responses of mouse olfactory sensory neurons using the air-phase electroolfactogram recording. J Vis Exp JoVE 2010; (37) pii: 1850; PMID:20197755 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [54].Pinato G, Rievaj J, Pifferi S, Dibattista M, Masten L, Menini A. Electroolfactogram responses from organotypic cultures of the olfactory epithelium from postnatal mice. Chem Senses 2008; 33:397-404; PMID:18303030; http://dx.doi.org/ 10.1093/chemse/bjn007 [DOI] [PubMed] [Google Scholar]
- [55].Brunet LJ, Gold GH, Ngai J. General anosmia caused by a targeted disruption of the mouse olfactory cyclic nucleotide–gated cation channel. Neuron 1996; 17:681-93; PMID:8893025; http://dx.doi.org/ 10.1016/S0896-6273(00)80200-7 [DOI] [PubMed] [Google Scholar]
- [56].Zhao H, Reed RR. X Inactivation of the OCNC1 channel gene reveals a role for activity-dependent competition in the olfactory system. Cell 2001; 104:651-60; PMID:11257220; http://dx.doi.org/ 10.1016/S0092-8674(01)00262-8 [DOI] [PubMed] [Google Scholar]
- [57].Lin W, Arellano J, Slotnick B, Restrepo D. Odors detected by mice deficient in cyclic nucleotide-gated channel subunit A2 stimulate the main olfactory system. J Neurosci 2004; 24:3703-10; PMID:15071119; http://dx.doi.org/ 10.1523/JNEUROSCI.0188-04.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [58].Boccaccio A, Sagheddu C, Menini A. Flash photolysis of caged compounds in the cilia of olfactory sensory neurons. J Vis Exp JoVE 2011; e3195; PMID:22064384 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [59].Pifferi S, Pascarella G, Boccaccio A, Mazzatenta A, Gustincich S, Menini A, Zucchelli S. Bestrophin-2 is a candidate calcium-activated chloride channel involved in olfactory transduction. Proc Natl Acad Sci 2006; 103:12929-34; PMID:16912113; http://dx.doi.org/ 10.1073/pnas.0604505103 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [60].Sagheddu C, Boccaccio A, Dibattista M, Montani G, Tirindelli R, Menini A. Calcium concentration jumps reveal dynamic ion selectivity of calcium-activated chloride currents in mouse olfactory sensory neurons and TMEM16b-transfected HEK 293T cells. J Physiol 2010; 588:4189-204; PMID:20837642; http://dx.doi.org/ 10.1113/jphysiol.2010.194407 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [61].Kleene SJ. Origin of the chloride current in olfactory transduction. Neuron 1993; 11:123-32; PMID:8393322; http://dx.doi.org/ 10.1016/0896-6273(93)90276-W [DOI] [PubMed] [Google Scholar]
- [62].Duran C, Thompson CH, Xiao Q, Hartzell HC. Chloride channels: often enigmatic, rarely predictable. Annu Rev Physiol 2010; 72:95-121; PMID:19827947; http://dx.doi.org/ 10.1146/annurev-physiol-021909-135811 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [63].Yang C, Delay RJ. Calcium-activated chloride current amplifies the response to urine in mouse vomeronasal sensory neurons. J Gen Physiol 2010; 135:3-13; PMID:20038523; http://dx.doi.org/ 10.1085/jgp.200910265 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [64].Kim S, Ma L, Yu CR. Requirement of calcium-activated chloride channels in the activation of mouse vomeronasal neurons. Nat Commun 2011; 2:365; PMID:21694713; http://dx.doi.org/ 10.1038/ncomms1368 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [65].Dibattista M, Amjad A, Maurya DK, Sagheddu C, Montani G, Tirindelli R, Menini A. Calcium-activated chloride channels in the apical region of mouse vomeronasal sensory neurons. J Gen Physiol 2012; 140:3-15; PMID:22732308; http://dx.doi.org/ 10.1085/jgp.201210780 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [66].Amjad A, Hernandez-Clavijo A, Pifferi S, Maurya DK, Boccaccio A, Franzot J, Rock J, Menini A. Conditional knockout of TMEM16A/anoctamin1 abolishes the calcium-activated chloride current in mouse vomeronasal sensory neurons. J Gen Physiol 2015; 145:285-301; PMID:25779870; http://dx.doi.org/ 10.1085/jgp.201411348 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [67].Hartzell C, Qu Z, Wei R, Mann W, Fischmeister R. Molecular physiology of calcium-activated chloride channels. speaker abstracts. J Gen Physiol 2003; 122:1a-46a; http://dx.doi.org/ 10.1085/jgp.20031221abstracts [DOI] [Google Scholar]
- [68].Pifferi S, Dibattista M, Sagheddu C, Boccaccio A, Al Qteishat A, Ghirardi F, Tirindelli R, Menini A. Calcium-activated chloride currents in olfactory sensory neurons from mice lacking bestrophin-2. J Physiol 2009; 587:4265-79; PMID:19622610; http://dx.doi.org/ 10.1113/jphysiol.2009.176131 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [69].Yu K, Duran C, Qu Z, Cui YY, Hartzell HC. Explaining calcium-dependent gating of anoctamin-1 chloride channels requires a revised topologynovelty and significance. Circ Res 2012; 110:990-9; PMID:22394518; http://dx.doi.org/ 10.1161/CIRCRESAHA.112.264440 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [70].Brunner JD, Lim NK, Schenck S, Duerst A, Dutzler R. X-ray structure of a calcium-activated TMEM16 lipid scramblase. Nature 2014; 516:207-12; PMID:25383531; http://dx.doi.org/ 10.1038/nature13984 [DOI] [PubMed] [Google Scholar]
- [71].Pedemonte N, Galietta LJV. Structure and function of TMEM16 proteins (Anoctamins). Physiol Rev 2014; 94:419-59; PMID:24692353; http://dx.doi.org/ 10.1152/physrev.00039.2011 [DOI] [PubMed] [Google Scholar]
- [72].Picollo A, Malvezzi M, Accardi A. TMEM16 proteins: unknown structure and confusing functions. J Mol Biol 2015; 427:94-105; PMID:25451786; http://dx.doi.org/ 10.1016/j.jmb.2014.09.028 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [73].Pifferi S, Dibattista M, Menini A. TMEM16B induces chloride currents activated by calcium in mammalian cells. Pflüg Arch Eur J Physiol 2009; 458:1023-38; http://dx.doi.org/ 10.1007/s00424-009-0684-9 [DOI] [PubMed] [Google Scholar]
- [74].Stöhr H, Heisig JB, Benz PM, Schöberl S, Milenkovic VM, Strauss O, Aartsen WM, Wijnholds J, Weber BHF, Schulz HL. TMEM16B, A novel protein with calcium-dependent chloride channel activity, associates with a presynaptic protein complex in photoreceptor terminals. J Neurosci 2009; 29:6809-18; PMID:19474308; http://dx.doi.org/ 10.1523/JNEUROSCI.5546-08.2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [75].Stephan AB, Shum EY, Hirsh S, Cygnar KD, Reisert J, Zhao H. ANO2 is the cilial calcium-activated chloride channel that may mediate olfactory amplification. Proc Natl Acad Sci 2009; 106:11776-81; PMID:19561302; http://dx.doi.org/ 10.1073/pnas.0903304106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [76].Mayer U, Küller A, Daiber PC, Neudorf I, Warnken U, Schnölzer M, Frings S, Möhrlen F. The proteome of rat olfactory sensory cilia. Proteomics 2009; 9:322-34; PMID:19086097; http://dx.doi.org/ 10.1002/pmic.200800149 [DOI] [PubMed] [Google Scholar]
- [77].Yu T-T, McIntyre JC, Bose SC, Hardin D, Owen MC, McClintock TS. Differentially expressed transcripts from phenotypically identified olfactory sensory neurons. J Comp Neurol 2005; 483:251-62; PMID:15682396; http://dx.doi.org/ 10.1002/cne.20429 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [78].Hengl T, Kaneko H, Dauner K, Vocke K, Frings S, Möhrlen F. Molecular components of signal amplification in olfactory sensory cilia. Proc Natl Acad Sci 2010; 107:6052-7; PMID:20231443; http://dx.doi.org/ 10.1073/pnas.0909032107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [79].Rasche S, Toetter B, Adler J, Tschapek A, Doerner JF, Kurtenbach S, Hatt H, Meyer H, Warscheid B, Neuhaus EM. Tmem16b is Specifically Expressed in the Cilia of Olfactory Sensory Neurons. Chem Senses 2010; 35:239-45; PMID:20100788; http://dx.doi.org/ 10.1093/chemse/bjq007 [DOI] [PubMed] [Google Scholar]
- [80].Henkel B, Drose DR, Ackels T, Oberland S, Spehr M, Neuhaus EM. Co-expression of anoctamins in cilia of olfactory sensory neurons. Chem Senses 2015; 40:73-87; PMID:25500808; http://dx.doi.org/ 10.1093/chemse/bju061 [DOI] [PubMed] [Google Scholar]
- [81].Billig GM, Pál B, Fidzinski P, Jentsch TJ. Ca2+-activated Cl- currents are dispensable for olfaction. Nat Neurosci 2011; 14:763-9; PMID:21516098; http://dx.doi.org/ 10.1038/nn.2821 [DOI] [PubMed] [Google Scholar]
- [82].Maurya DK, Menini A. Developmental expression of the calcium-activated chloride channels TMEM16A and TMEM16B in the mouse olfactory epithelium. Dev Neurobiol 2014; 74:657-75; PMID:24318978; http://dx.doi.org/ 10.1002/dneu.22159 [DOI] [PubMed] [Google Scholar]
- [83].Maurya DK, Henriques T, Marini M, Pedemonte N, Galietta LJV, Rock JR, Harfe BD, Menini A. Development of the olfactory epithelium and nasal glands in TMEM16A−/− and TMEM16A+/+ mice. PloS One 2015; 10:e0129171; PMID:26067252; http://dx.doi.org/ 10.1371/journal.pone.0129171 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [84].Tien J, Lee HY, Minor DL, Jan YN, Jan LY. Identification of a dimerization domain in the TMEM16A calcium-activated chloride channel (CaCC). Proc Natl Acad Sci 2013; 110:6352-7; PMID:23576756; http://dx.doi.org/ 10.1073/pnas.1303672110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [85].Saidu SP, Stephan AB, Talaga AK, Zhao H, Reisert J. Channel properties of the splicing isoforms of the olfactory calcium-activated chloride channel Anoctamin 2. J Gen Physiol 2013; 141:691-703; PMID:23669718; http://dx.doi.org/ 10.1085/jgp.201210937 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [86].Pifferi S, Cenedese V, Menini A. Anoctamin 2/TMEM16B: a calcium-activated chloride channel in olfactory transduction. Exp Physiol 2012; 97:193-9; PMID:21890523; http://dx.doi.org/ 10.1113/expphysiol.2011.058230 [DOI] [PubMed] [Google Scholar]
- [87].Pietra G, Dibattista M, Menini A, Reisert J, Boccaccio A. The Ca2+-activated Cl− channel TMEM16B regulates action potential firing and axonal targeting in olfactory sensory neurons. J Gen Physiol 2016; 148:293-311; PMID:27619419; http://dx.doi.org/ 10.1085/jgp.201611622 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [88].Huang WC, Xiao S, Huang F, Harfe BD, Jan YN, Jan LY. Calcium-activated chloride channels (CaCCs) regulate action potential and synaptic response in hippocampal neurons. Neuron 2012; 74:179-92; PMID:22500639; http://dx.doi.org/ 10.1016/j.neuron.2012.01.033 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [89].Zhang W, Schmelzeisen S, Parthier D, Frings S, Möhrlen F. Anoctamin calcium-activated chloride channels may modulate inhibitory transmission in the cerebellar cortex. PLOS One 2015; 10:e0142160; PMID:26558388; http://dx.doi.org/ 10.1371/journal.pone.0142160 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [90].Ha GE, Lee J, Kwak H, Song K, Kwon J, Jung SY, Hong J, Chang GE, Hwang EM, Shin HS, et al.. The Ca(2+)-activated chloride channel anoctamin-2 mediates spike-frequency adaptation and regulates sensory transmission in thalamocortical neurons. Nat Commun 2016; 7:13791; PMID:27991499; http://dx.doi.org/ 10.1038/ncomms13791 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [91].Lynch JW, Barry PH. Action potentials initiated by single channels opening in a small neuron (rat olfactory receptor). Biophys J 1989; 55:755-68; PMID:2470428; http://dx.doi.org/ 10.1016/S0006-3495(89)82874-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [92].Trotier D. Intensity coding in olfactory receptor cells. Semin Cell Biol 1994; 5:47-54; PMID:7514456; http://dx.doi.org/ 10.1006/scel.1994.1007 [DOI] [PubMed] [Google Scholar]
- [93].Lynch JW, Barry PH. Inward rectification in rat olfactory receptor neurons. Proc Biol Sci 1991; 243:149-53; PMID:1676518; http://dx.doi.org/ 10.1098/rspb.1991.0024 [DOI] [PubMed] [Google Scholar]
- [94].Dibattista M, Reisert J. The odorant receptor-dependent role of olfactory marker protein in olfactory receptor neurons. J Neurosci 2016; 36:2995-3006; PMID:26961953; http://dx.doi.org/ 10.1523/JNEUROSCI.4209-15.2016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [95].Rospars JP, Lansky P, Chaput M, Duchamp-Viret P. Competitive and noncompetitive odorant interactions in the early neural coding of odorant mixtures. J Neurosci 2008; 28:2659-66; PMID:18322109; http://dx.doi.org/ 10.1523/JNEUROSCI.4670-07.2008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [96].Gesteland RC, Sigwart CD. Olfactory receptor units—a mammalian preparation. Brain Res 1977; 133:144-9; PMID:902082; http://dx.doi.org/ 10.1016/0006-8993(77)90055-5 [DOI] [PubMed] [Google Scholar]
- [97].Firestein S, Picco C, Menini A. The relation between stimulus and response in olfactory receptor cells of the tiger salamander. J Physiol 1993; 468:1-10; PMID:8254501; http://dx.doi.org/ 10.1113/jphysiol.1993.sp019756 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [98].Ponissery Saidu S, Dibattista M, Matthews HR, Reisert J. Odorant-induced responses recorded from olfactory receptor neurons using the suction pipette technique. J Vis Exp JoVE 2012; e3862; PMID:22508037 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [99].Reisert J. Origin of basal activity in mammalian olfactory receptor neurons. J Gen Physiol 2010; 136:529-40; PMID:20974772; http://dx.doi.org/ 10.1085/jgp.201010528 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [100].Connelly T, Savigner A, Ma M. Spontaneous and sensory-evoked activity in mouse olfactory sensory neurons with defined odorant receptors. J Neurophysiol 2013; 110:55-62; PMID:23596334; http://dx.doi.org/ 10.1152/jn.00910.2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [101].Araneda RC, Kini AD, Firestein S. The molecular receptive range of an odorant receptor. Nat Neurosci 2000; 3:1248-55; PMID:11100145; http://dx.doi.org/ 10.1038/81774 [DOI] [PubMed] [Google Scholar]
- [102].Kurland MD, Newcomer MB, Peterlin Z, Ryan K, Firestein S, Batista VS. Discrimination of saturated aldehydes by the rat I7 olfactory receptor. Biochemistry (Mosc) 2010; 49:6302-4; http://dx.doi.org/ 10.1021/bi100976w [DOI] [PMC free article] [PubMed] [Google Scholar]
- [103].Zou D-J, Chesler A, Firestein S. How the olfactory bulb got its glomeruli: a just so story? Nat Rev Neurosci 2009; 10:611-8; PMID:19584894; http://dx.doi.org/ 10.1038/nrn2666 [DOI] [PubMed] [Google Scholar]
- [104].Lorenzon P, Redolfi N, Podolsky MJ, Zamparo I, Franchi SA, Pietra G, Boccaccio A, Menini A, Murthy VN, Lodovichi C. Circuit formation and function in the olfactory bulb of mice with reduced spontaneous afferent activity. J Neurosci 2015; 35:146-60; PMID:25568110; http://dx.doi.org/ 10.1523/JNEUROSCI.0613-14.2015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [105].Imai T, Suzuki M, Sakano H. Odorant receptor-derived cAMP signals direct axonal targeting. Science 2006; 314:657-61; PMID:16990513; http://dx.doi.org/ 10.1126/science.1131794 [DOI] [PubMed] [Google Scholar]
- [106].Nakashima A, Takeuchi H, Imai T, Saito H, Kiyonari H, Abe T, Chen M, Weinstein LS, Yu CR, Storm DR, et al.. Agonist-independent GPCR activity regulates anterior-posterior targeting of olfactory sensory neurons. Cell 2013; 154:1314-25; PMID:24034253; http://dx.doi.org/ 10.1016/j.cell.2013.08.033 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [107].Takeuchi H, Sakano H. Neural map formation in the mouse olfactory system. Cell Mol Life Sci CMLS 2014; 71:3049-57; PMID:24638094; http://dx.doi.org/ 10.1007/s00018-014-1597-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [108].Zapiec B, Bressel OC, Khan M, Walz A, Mombaerts P. Neuropilin-1 and the positions of glomeruli in the mouse olfactory bulb. eNeuro 2016; 3; PMID:27844052; http://dx.doi.org/ 10.1523/ENEURO.0123-16.2016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [109].Movahedi K, Grosmaitre X, Feinstein P. Odorant receptors can mediate axonal identity and gene choice via cAMP-independent mechanisms. Open Biol [Internet] 2016. [cited 2016 Dec 24]; 6:pii: 160018. Available from: http://www.ncbi.nlm.nih.gov/pmc/articles/PMC4967819/; PMID:27466441; http://dx.doi.org/ 10.1098/rsob.160018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [110].Assens A, Dal Col J, Njoku A, Dietschi Q, Kan C, Feinstein P, Carleton A, Rodriguez I. Alteration of Nrp1 signaling at different stages of olfactory neuron maturation promotes glomerular shifts along distinct axes in the olfactory bulb. Dev Camb Engl 2016; 143(20):3817-25 [DOI] [PMC free article] [PubMed] [Google Scholar]