

Abstract
Mechanical cues and substrate interaction affect the manner in which cells adhere, spread, migrate and form tissues. With increased interest in tissue-on-a-chip and co-culture systems utilizing porous membranes, it is important to understand the role of disrupted surfaces on cellular behavior. Using a transparent glass membrane with defined pore geometries, we investigated endothelial fibronectin fibrillogenesis and formation of focal adhesions as well as development of intercellular junctions. Cells formed fewer focal adhesions and had shorter fibronectin fibrils on porous membranes compared to non-porous controls, which was similar to cell behavior on continuous soft substrates with Young’s moduli seven orders of magnitude lower than glass. Additionally, porous membranes promoted enhanced cell-cell interactions as evidenced by earlier formation of tight junctions. These findings suggest that porous membranes with discontinuous surfaces promote reduced cell-matrix interactions similarly to soft substrates and may enhance tissue and barrier formation.
Keywords: membrane, focal adhesion, fibronectin, substrate stiffness
Graphical abstract
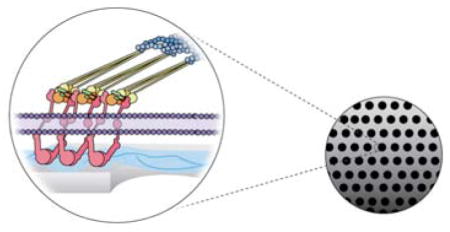
There is rapidly growing interest in developing organ-on-a-chip, membrane-supported barrier models and co-culture systems. Porous membranes have been used to support the development of in vitro models of the gut,1 lung,2 placenta,3 blood-brain-barrier4,5 and other tissues.6 Additionally, porous membranes are being used to support cell-mediated stem cell differentiation, where a second cell type induces differentiation through physical contact or paracrine signaling.7,8 Research laboratories are also using porous membranes to support a variety of multi-compartment microenvironments that recapitulate complex physiological systems. Advances in these areas are likely to dramatically reduce drug development costs and replace animal models with human cell-based assays.9
At the same time, many laboratories are researching more physiologically relevant cell culture substrates and cellular scaffolds.10 One major area of focus is optimizing substrate stiffness in order to mimic the mechanical properties of tissue.11,12 There are two commonly used approaches: tunable stiffness hydrogels13 and engineered deformable microposts.14 Studies using microposts have typically been designed with varying micropost length to control the effective substrate stiffness felt by the cells. Additionally, deflection of the posts can be used to calculate cellular contractile forces. Hydrogels have also been used to investigate the relationship between traction forces and substrate stiffness by tracking displacement of embedded beads.15,16 Like microposts, hydrogels of varying stiffness have been shown to affect a variety of cell processes including stem cell differentiation.17 While both approaches have been successful in recapitulating physical aspects of tissue, neither directly represents porous membranes used in the burgeoning organ-on-a-chip field. In fact, an array of microposts is the geometric inverse of a porous membrane with respect to cell contact area.
One of the most well studied cell-substrate interactions is the anchoring of cells via focal adhesions (FAs).18 These interactions are not just important in cellular sensing of the microenvironment during migration, but also in the development and maintenance of barriers such as the vascular wall.19 Distinct FAs are widely apparent when cells are cultured on stiff surfaces, particularly tissue culture polystyrene (TCPS) and glass.20 On the other hand, cells growing in three-dimensional tissues and on relatively soft substrates display fewer and smaller FAs.21 Research on microposts and hydrogels has shown that cellular traction forces are strongly correlated with formation of FAs.22–24
Extracellular matrix fibrillogenesis has also been shown to be affected by substrate stiffness.25 Fibronectin (FN) fibrillogenesis is associated with a cell’s ability to generate high traction forces, which in turn requires a relatively stiff supporting matrix or substrate. On the other hand, cells cultured on soft substrates display significantly shorter FN fibrils that appear less organized.26,27 In fact, the orientation of the fibrils is known to be directed by the cell’s actin stress fibers.25
In this study, we investigated the effect of membrane pore spacing on cell-substrate interactions and corresponding cell-cell interactions. We used transparent silicon dioxide (SiO2; glass) membranes with well-defined micron and sub-micron pore sizes and spacing that enabled high-resolution imaging of FAs, stress fibers and FN fibrillogenesis. We found that the disrupted surface geometry of these porous membranes negatively affected FA formation and FN fibrillogenesis. While holding total membrane contact area constant, but decreasing pore size and pore spacing, we observed cellular behavior similar to cells cultured on continuous soft substrates.
Use of ultrathin glass SiO2 membranes enabled visualization of subcellular features and nascent extracellular matrix structures. These membranes were fabricated 300 nm thick with hexagonally patterned 0.5 μm or 3.0 μm diameter pores (Figure 1). Both porous membranes had approximately 23% porosity and 77% contact area (Figure 1A and B). Fabrication details have been reported previously28 and can be found in Supporting Information. Non-porous SiO2 membranes were fabricated in the same manner and used as control substrates. We chose these pore sizes because of their ability to prevent (0.5 μm) or permit (3.0 μm) cellular migration across the barrier in co-culture and leukocyte-endothelial transmigration studies. It is challenging to culture confluent monolayers on membranes with pore sizes > 3 μm due to the tendency for endothelial and epithelial transmigration. While polymeric track-etched (TE) membranes with similar pore sizes are widely available, TE membranes have varying pore densities and inter-pore spacing (Figure 1C and D) that likely affects cell-substrate interactions. We chose to use patterned SiO2 membranes for controlled pore spacing and excellent imaging properties.
Figure 1.

Scanning electron microscopy (SEM) images of 300 nm thick SiO2 membranes with (A) 3.0 μm and (B) 0.5 μm diameter pores; Greiner Bio-One Thincert® track etched high-porosity membranes with (C) 3.0 μm and d) 0.4 μm diameter pores. Dotted white outline shows the size of a typical cell and its nucleus. Illustrations (E) and (F) show potential focal adhesion formations on 0.5 μm and 3.0 μm diameter pore SiO2 membranes.
In addition to investigating cellular behavior on stiff porous substrates, we also studied cell-substrate interactions on more compliant polydimethylsiloxane (PDMS). We used common Sylgard® 184, which has Young’s modulus of 2 MPa as well as softer Sylgard 527®, which has a Young’s modulus of just 5 kPa.29 This is seven orders of magnitude lower than the modulus of SiO2 thin films30 and comparable to tissue. For PDMS experiments, we cast silicone substrates that were >500 μm thick to minimize the influence of the underlying TCPS support. Complete cell culture and immunofluorescence methods are available in Supporting Information.
We investigated the behavior of human umbilical vein endothelial cells (HUVECs) on porous membranes because of their use in vascular barrier models. We suspected that HUVECs may not adhere and spread as well on porous membranes due to the disrupted surface geometry. Cell spread area was measured using image analysis of actin staining after 24 hours. There were only minor differences in cell spreading across porous and non-porous substrates (Figure S1). Median spread area on soft PDMS was the lowest. Actin stress fibers were most pronounced on TCPS and non-porous glass membranes, but were visible on all substrates (Figure 2, top). Nearly all cells on TCPS and non-porous glass had distinct FAs with approximately 50 per cell, which is consistent with values in the literature.31 Similar to previous studies, there were fewer FAs per cell on soft compared to stiff continuous substrates.32 Two-thirds of cells on 3.0 μm pore membranes displayed FAs. Those cells averaged 33 FAs, significantly fewer than TCPS and non-porous glass. Less than one-third of cells on 0.5 μm pore membranes displayed FAs with an average of just 18 per cell, significantly fewer than even soft PDMS (Figure 2, bottom).
Figure 2.

Representative images of endothelial focal adhesions after 24 hours on TCPS, SiO2 membranes, Stiff (2 MPa) PDMS and Soft (5 kPA) PDMS substrates. Cells were stained for nuclei (DAPI, blue), F-actin (phalloidin, green), and focal adhesions (anti-vinculin, red). Bottom: Quantification of focal adhesion formation after 24 hours of culture. Formation of distinct focal adhesions was quantified for all substrates - percent of cells with distinct focal adhesions and number of focal adhesions per cell (n > 20 for each substrate; mean ± standard deviation; one-way ANOVA with a Tukey post hoc analysis).
We also investigated FN fibrillogenesis after 24 hours. The relationships between substrate stiffness, cellular traction forces and extracellular matrix (ECM) formation has been well studied.33 Several groups have shown that substrate stiffness is directly related to ECM fibrillogenesis, with endothelial and epithelial cells on stiffer substrates producing longer and more robust FN fibrils.26,27 This may be due to the greater internal traction forces that can be generated on stiff compared to more compliant substrates, which showed relatively shorter and fewer fibrils. On the other hand, Scott et. al. reported that FN assembly by fibroblasts on microposts was roughly constant across multiple stiffnesses.34 In our experiments we saw that HUVECs on stiff continuous non-porous SiO2 membranes produced qualitatively longer and brighter fibrils compared to soft PDMS (Figure 3A). FN fibrils were similar on TCPS and non-porous SiO2 (Figure S2 A). Interestingly, cells on 0.5 μm pore membranes displayed shorter fibrils similar to soft PDMS. We were able to document fibrillogenesis at the border between non-porous and patterned 3.0 μm pores. Fibrils appeared longer over the non-porous region, but significantly shorter over the porous region more than 20 μm from the border confirming the effect of a disrupted surface on FN fibrillogenesis. FN density on SiO2 substrates was quantified by measuring fluorescence intensity within 100×100 μm regions occupied by cells. FN intensity on both porous SiO2 membranes was significantly less than non-porous SiO2 (Figure 3B). The number of FN fibrils greater than 5 and 10 μm in length was also quantified using a custom automated imaging processing algorithm (see Supporting Information and Figure S3). Consistent with the qualitative assessment, 0.5 μm pore membranes displayed the fewest long fibrils (Figure 3C and D). Due to the heterogeneity of PDMS background fluorescence, we were unable to quantify FN characteristics on those substrates.
Figure 3.

(A) Representative images of endothelial fibronectin fibrillogenesis after 24 hours on non-porous, 3.0 μm and 0.5 μm pore diameter SiO2 membranes, 2 MPa (Stiff) PDMS and 5 kPa (Soft) PDMS substrates. The image of the 3.0 μm pore diameter SiO2 membrane shows the boundary between the porous and non-porous regions. (B) Fibronectin density on each SiO2 membrane substrate was approximated by measuring the mean fluorescence intensity of anti-fibronectin antibody within 100×100 μm regions of interest that were occupied by cells (n > 15 for each substrate). (C) and (D) Fibronectin fibril lengths were also measured using an automated image processing algorithm (comparisons were made using one-way ANOVA with a Tukey post hoc analysis).
Fewer FAs and less FN fibrillogenesis on porous membranes suggest reduced cell-substrate interactions similar to continuous soft substrates. It was hypothesized by Guo et. al.32 that substrate rigidity regulates the formation of tissues. Specifically, weak cell-substrate interactions may promote cell-cell interactions leading to tissue and barrier formation. To determine if the disrupted surface of a porous membrane would similarly promote cell-cell interactions, we investigated the presence of a tight junctional protein, ZO-1, which regulates barrier formation.35 We fixed and stained cells at 96 hours as they first reached confluence. ZO-1 staining was punctate on non-porous SiO2 and TCPS (Figure S2 B) but robust at cell borders on soft PDMS and 0.5 μm pore membranes (Figure 4). Endothelial monolayers on 3.0 μm pore membranes and stiff PDMS displayed an intermediate degree of ZO-1 staining. These data are consistent with the idea that weak cell-substrate interactions lead to strong cell-cell interactions and earlier barrier formation on porous membranes similarly to soft substrates. After 7 days, ZO-1 labeling did increase on all substrates, suggesting that cell-cell interactions may ultimately dominate for endothelial cells regardless of initial cell-substrate interactions. While this is consistent with historical success in studying endothelial behavior on continuous and impermeable culture surfaces such as TCPS, discontinuous or porous substrates may promote faster barrier formation. Future studies are necessary to determine if functional barrier properties can be improved or optimized based on reduced cell-substrate interactions.
Figure 4.

Representative images of tight junction protein ZO-1 after 4 days culture as the cells initially reached confluence on non-porous, 3.0 μm and 0.5 μm pore diameter SiO2 membranes, 2 MPa (Stiff) PDMS and 5 kPa (Soft) PDMS substrates. ZO-1 staining was most consistent at cell borders on 0.5 μm pore diameter SiO2 membranes and soft PDMS.
Data presented thus far suggest that a disrupted surface geometry weakens a cell’s interactions with the underlying substrate and leads instead to enhanced cell-cell interactions. Despite identical total contact area on both porous membranes, cells on 0.5 μm pore membranes displayed more pronounced differences in cell-substrate and cell-cell interactions compared to continuous glass and TCPS. This suggests that inter-pore spacing and limited contact regions are the mechanism behind the differences in behavior. We tested this hypothesis by investigating the alignment of actin stress fibers. Even though very few cells on 0.5 μm pore membranes displayed FAs, it was still possible to measure the alignment of primary stress fibers that terminated at cell-matrix adhesion sites.
The hexagonal spacing of the membrane pores resulted in what we postulated were two regions where the largest cell-substrate interactions could form (Figure 5). We investigated alignment in either the a or b directions as well as the intermediate angles by measuring the angle of the primary actin stress fibers from the a direction (Figure S4). Due to the three axes of symmetry, all data was mapped to the primary directions for the radial histogram (Figure 5). Not surprisingly, we found no preferential alignment on the non-porous substrates. Additionally, we found no preference on 3.0 μm pore membranes, suggesting that both a or b contact regions were sufficiently large to support strong cell-substrate interactions. However, cells on 0.5 μm pore membranes had stress fibers that predominately aligned in the a direction, suggesting that contact area confinement due to pore spacing was limiting. While the b region had theoretically an unlimited length, its width was significantly limited. The a region was shorter, but had greater width and circularity, which may have supported better adhesive interactions.
Figure 5.

Schematic shows pore spacing and geometries of potential cell-substrate interactions for both 0.5 μm and 3.0 μm pore size membranes. Due to hexagonal pore spacing, there are three lines of symmetry. Alignment of F-actin stress fibers was measured for non-porous SiO2, 3.0 μm and 0.5 μm pore membranes. Normalized radial histograms show the distribution of aligned cells for each substrate (n = 32–37).
This data raises the question of whether cells can generate ECM fibrils that span the pores and if this cell-generated matrix can overcome contact area confinement. We stained for vinculin at 6 days and found that cells did indeed form FAs over some pores (Figure S5), indicating that the maturing ECM spanned pores and was sufficiently stable to support cellular traction forces. Interestingly, FAs on 3.0 μm pore membranes at 6 days increased to levels comparable to non-porous substrates. Cellular FAs also increased on 0.5 μm pore membranes, but were still lower than continuous substrates. In contrast, we found no focal adhesions that spanned pores at 24 hours on 3.0 μm pore membranes (Figure S4). Due to the sub-micron pore spacing on the 0.5 μm membranes and the limits of conventional optical resolution, it was difficult to definitely state whether or not focal adhesions spanned pores on these smaller pores. The discontinuous nature of the porous membrane in combination with small pore spacing may permanently affect cell-substrate interactions. We believe the close proximity of pores results in many cell-matrix interactions occurring over open pore regions leading to more physiological sensing of ECM stiffness. On the other hand, large pore spacing may facilitate ECM-substrate interactions that result in a far stiffer response when cells exert traction forces on the matrix. These observations are consistent with the model proposed by Trichet et. al. where cellular response to substrate stiffness is large-scale and not due to individual FAs.12
In summary, we have shown that the disrupted surface of porous membranes leads to weakened cell-substrate interactions that instead promote cell-cell interactions. This behavior is similar to continuous soft PDMS substrates despite a difference in Young’s modulus of over seven orders of magnitude. Together, these data illustrate that membrane pore properties can be designed to produce a support substrate with an effective stiffness much closer to physiological levels. These results should be extendable to membranes and discontinuous substrates made of a variety of materials. However, these effects are not likely seen to the same degree on track-etched polymeric membranes due to highly variable pore density and greater pore spacing. 36,37
Supplementary Material
Acknowledgments
Funding Sources
Research reported in this publication was supported in part by NIGMS of the National Institutes of Health under award number R35GM119623 to T.G. and the Gleason Family Foundation.
We thank Mehdi Dehghani for taking the scanning electron microscopy membrane images and Joseph Olmsted for creating the cell-substrate interaction illustrations.
Footnotes
Author Contributions
The manuscript was written through contributions of all authors. All authors have given approval to the final version of the manuscript.
Notes
T.G. is a co-founder of SiMPore, a nanomaterials start-up company that fabricates ultrathin membranes. The content of this publication is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
The Supporting Information is available free of charge on the ACS Publications website.
Experimental details on membrane fabrication, cell culture and image analysis (PDF).
References
- 1.Hubatsch I, Ragnarsson EGE, Artursson P. Determination of drug permeability and prediction of drug absorption in Caco-2 monolayers. Nat Protoc. 2007;2(9):2111–2119. doi: 10.1038/nprot.2007.303. [DOI] [PubMed] [Google Scholar]
- 2.Huh D, Matthews BD, Mammoto A, Montoya-Zavala M, Hsin HY, Ingber DE. Reconstituting organ-level lung functions on a chip. Science. 2010;328(5986):1662–1668. doi: 10.1126/science.1188302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Lee JS, Romero R, Han YM, Kim HC, Kim CJ, Hong J-S, Huh D. Placenta-on-a-chip: a novel platform to study the biology of the human placenta. J Matern Fetal Neonatal Med. 2016;29(7):1046–1054. doi: 10.3109/14767058.2015.1038518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Ma SH, Lepak LA, Hussain RJ, Shain W, Shuler ML. An endothelial and astrocyte co-culture model of the blood?brain barrier utilizing an ultra-thin, nanofabricated silicon nitride membrane. Lab Chip. 2005;5(1):74–85. doi: 10.1039/b405713a. [DOI] [PubMed] [Google Scholar]
- 5.Naik P, Cucullo L. In vitro blood–brain barrier models: Current and perspective technologies. Journal of Pharmaceutical Sciences. 2012;101(4):1337–1354. doi: 10.1002/jps.23022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Huh D, Kim HJ, Fraser JP, Shea DE, Khan M, Bahinski A, Hamilton GA, Ingber DE. Microfabrication of human organs-on-chips. Nat Protoc. 2013;8(11):2135–2157. doi: 10.1038/nprot.2013.137. [DOI] [PubMed] [Google Scholar]
- 7.Mazzocchi AR, Man AJ, Desormeaux J-PS, Gaborski TR. Porous Membranes Promote Endothelial Differentiation of Adipose-Derived Stem Cells and Perivascular Interactions. Cellular and Molecular Bioengineering. 2014;7(3):369–378. doi: 10.1007/s12195-014-0354-7. [DOI] [Google Scholar]
- 8.Ryu S, Yoo J, Jang Y, Han J, Yu SJ, Park J, Jung SY, Ahn KH, Im SG, Char K, et al. Nanothin Coculture Membranes with Tunable Pore Architecture and Thermoresponsive Functionality for Transfer-Printable Stem Cell-Derived Cardiac Sheets. ACS Nano. 2015;9(10):10186–10202. doi: 10.1021/acsnano.5b03823. [DOI] [PubMed] [Google Scholar]
- 9.Esch EW, Bahinski A, Huh D. Organs-on-chips at the frontiers of drug discovery. Nat Rev Drug Discov. 2015;14(4):248–260. doi: 10.1038/nrd4539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Hickman GJ, Boocock DJ, Pockley AG, Perry CC. The Importance and Clinical Relevance of Surfaces in Tissue Culture. ACS Biomater Sci Eng. 2016;2(2):152–164. doi: 10.1021/acsbiomaterials.5b00403. [DOI] [PubMed] [Google Scholar]
- 11.Discher DE, Janmey P, Wang Y-L. Tissue cells feel and respond to the stiffness of their substrate. Science. 2005;310(5751):1139–1143. doi: 10.1126/science.1116995. [DOI] [PubMed] [Google Scholar]
- 12.Trichet L, Le Digabel J, Hawkins RJ, Vedula SRK, Gupta M, Ribrault C, Hersen P, Voituriez R, Ladoux B. Evidence of a large-scale mechanosensing mechanism for cellular adaptation to substrate stiffness. Proc Natl Acad Sci USA. 2012;109(18):6933–6938. doi: 10.1073/pnas.1117810109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Tse JR, Engler AJ. Preparation of Hydrogel Substrates with Tunable Mechanical Properties. John Wiley & Sons, Inc; Hoboken, NJ, USA: 2001. [DOI] [PubMed] [Google Scholar]
- 14.Fu J, Wang Y-K, Yang MT, Desai RA, Yu X, Liu Z, Chen CS. Mechanical regulation of cell function with geometrically modulated elastomeric substrates. Nat Meth. 2010;7(9):733–736. doi: 10.1038/nmeth.1487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Munevar S, Wang Y, Dembo M. Traction force microscopy of migrating normal and H-ras transformed 3T3 fibroblasts. Biophys J. 2001;80(4):1744–1757. doi: 10.1016/s0006-3495(01)76145-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Califano JP, Reinhart-King CA. Substrate Stiffness and Cell Area Predict Cellular Traction Stresses in Single Cells and Cells in Contact. Cellular and Molecular Bioengineering. 2010;3(1):68–75. doi: 10.1007/s12195-010-0102-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Engler AJ, Sen S, Sweeney HL, Discher DE. Matrix Elasticity Directs Stem Cell Lineage Specification. Cell. 2006;126(4):677–689. doi: 10.1016/j.cell.2006.06.044. [DOI] [PubMed] [Google Scholar]
- 18.Geiger B, Spatz JP, Bershadsky AD. Environmental sensing through focal adhesions. Nature Reviews Molecular Cell Biology. 2009;10(1):21–33. doi: 10.1038/nrm2593. [DOI] [PubMed] [Google Scholar]
- 19.Romer LH. Focal Adhesions: Paradigm for a Signaling Nexus. Circulation Research. 2006;98(5):606–616. doi: 10.1161/01.RES.0000207408.31270.db. [DOI] [PubMed] [Google Scholar]
- 20.Wehrle-Haller B. Structure and function of focal adhesions. Current Opinion in Cell Biology. 2012;24(1):116–124. doi: 10.1016/j.ceb.2011.11.001. [DOI] [PubMed] [Google Scholar]
- 21.Harunaga JS, Yamada KM. Cell-matrix adhesions in 3D. Matrix Biology. 2011;30(7–8):363–368. doi: 10.1016/j.matbio.2011.06.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Tan JL, Tien J, Pirone DM, Gray DS, Bhadriraju K, Chen CS. Cells lying on a bed of microneedles: An approach to isolate mechanical force. Proc Natl Acad Sci USA. 2003;100(4):1484–1489. doi: 10.1073/pnas.0235407100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Han SJ, Bielawski KS, Ting LH, Rodriguez ML. Decoupling substrate stiffness, spread area, and micropost density: a close spatial relationship between traction forces and focal adhesions. Biophys J. 2012;103:640–648. doi: 10.1016/j.bpj.2012.07.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Beningo KA, Wang Y-L. Flexible substrata for the detection of cellular traction forces. Trends in Cell Biology. 2002;12(2):79–84. doi: 10.1016/S0962-8924(01)02205-X. [DOI] [PubMed] [Google Scholar]
- 25.Lemmon CA, Chen CS, Romer LH. Cell Traction Forces Direct Fibronectin Matrix Assembly. Biophys J. 2009;96(2):729–738. doi: 10.1016/j.bpj.2008.10.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Rachelle N, Palchesko KLLJLFAWF. In Vitro Expansion of Corneal Endothelial Cells on Biomimetic Substrates. Sci Rep. 2015;5 doi: 10.1038/srep07955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Eisenberg JL, Safi A, Wei X, Espinosa HD, Budinger GS, Takawira D, Hopkinson SB, Jones JC. Substrate stiffness regulates extracellular matrix deposition by alveolar epithelial cells. Res Rep Biol. 2011;2011(2):1–12. doi: 10.2147/RRB.S13178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Carter RN, Casillo SM, Mazzocchi AR, Desormeaux J-PS, Roussie JA, Gaborski TR. Ultrathin transparent membranes for cellular barrier and co-culture models. Biofabrication. 2017;9(1):015019. doi: 10.1088/1758-5090/aa5ba7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Palchesko RN, Zhang L, Sun Y, Feinberg AW. Development of Polydimethylsiloxane Substrates with Tunable Elastic Modulus to Study Cell Mechanobiology in Muscle and Nerve. PLoS ONE. 2012;7(12):e51499. doi: 10.1371/journal.pone.0051499.s001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Li H, Vlassak JJ. Determining the elastic modulus and hardness of an ultra-thin film on a substrate using nanoindentation. J Mater Res. 2011;24(03):1114–1126. doi: 10.1557/jmr.2009.0144. [DOI] [Google Scholar]
- 31.Kanda S, Miyata Y, Kanetake H. Role of focal adhesion formation in migration and morphogenesis of endothelial cells. Cellular Signalling. 2004;16(11):1273–1281. doi: 10.1016/j.cellsig.2004.03.010. [DOI] [PubMed] [Google Scholar]
- 32.Guo W, Frey MT, Burnham NA, Wang Y. Substrate rigidity regulates the formation and maintenance of tissues. Biophys J. 2006;90(6):2213–2220. doi: 10.1529/biophysj.105.070144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Singh P, Carraher C, Schwarzbauer JE. Assembly of Fibronectin Extracellular Matrix. Annu Rev Cell Dev Biol. 2010;26(1):397–419. doi: 10.1146/annurev-cellbio-100109-104020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Scott LE, Mair DB, Narang JD, Feleke K, Lemmon CA. Fibronectin fibrillogenesis facilitates mechano-dependent cell spreading, force generation, and nuclear size in human embryonic fibroblasts. Integr Biol (Camb) 2015;7(11):1454–1465. doi: 10.1039/c5ib00217f. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Tornavaca O, Chia M, Dufton N, Almagro LO, Conway DE, Randi AM, Schwartz MA, Matter K, Balda MS. ZO-1 controls endothelial adherens junctions, cell–cell tension, angiogenesis, and barrier formation. J Cell Biol. 2015;208(6):821–838. doi: 10.1083/jcb.201404140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Apel PY, Blonskaya IV, Dmitriev SN. Structure of polycarbonate track-etch membranes: Origin of the “paradoxical” pore shape. Journal of Membrane Science. 2006;282:393–400. doi: 10.1016/j.memsci.2006.05.045. [DOI] [Google Scholar]
- 37.Kim MY, Li DJ, Pham LK, Wong BG, Hui EE. Microfabrication of high-resolution porous membranes for cell culture. Journal of Membrane Science. 2014;452:460–469. doi: 10.1016/j.memsci.2013.11.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.