

Abstract
Background
Both hydrogen sulfide (H2S) and hydrogen peroxide (H2O2) are separately regarded as a highly reactive molecule involved in root morphogenesis. In this report, corresponding causal link governing lateral root formation was investigated.
Methods
By using pharmacological, anatomic, and molecular approaches, evidence presented here revealed the molecular mechanism underlying tomato lateral root development triggered by H2S.
Results
A H2S donor sodium hydrosulfide (NaHS) triggered the accumulation of H2O2, the up-regulation of RBOH1 transcript, and thereafter tomato lateral root formation. Above responses were sensitive to the H2O2 scavenger (dimethylthiourea; DMTU) and the inhibitor of NADPH oxidase (diphenylene idonium; DPI), showing that the accumulations of H2O2 and increased RBOH1 transcript were respectively prevented. Lateral root primordial and lateral root formation were also impaired. Further molecular evidence revealed that H2S-modulated gene expression of cell cycle regulatory genes, including up-regulation of SlCYCA2;1, SlCYCA3;1, and SlCDKA1, and the down-regulation of SlKRP2, were prevented by the co-treatment with DMTU or DPI. Above mentioned inducing phenotypes were consistent with the changes of lateral root formation-related microRNA transcripts: up-regulation of miR390a and miR160, and with the opposite tendencies of their target genes (encoding auxin response factors). Contrasting tendencies were observed when DMTU or DPI was added together. The occurrence of H2S-mediated S-sulfhydration during above responses was preliminarily discovered.
Conclusions
Overall, these results suggested an important role of RBOH1-mediated H2O2 in H2S-elicited tomato lateral root development, and corresponding H2S-target proteins regulated at transcriptional and post-translational levels.
Electronic supplementary material
The online version of this article (10.1186/s12870-017-1110-7) contains supplementary material, which is available to authorized users.
Keywords: Hydrogen sulfide (H2S), Hydrogen peroxide (H2O2), Solanum lycopersicum, Lateral root formation, miRNA, S-sulfhydration
Background
Lateral root (LR) formation, which entirely originated from pericycle founder cells, is of critical importance for the plant root architecture [1]. Normally, LR formation depends on both genetic determinants and postembryonic developmental processes that are mainly under the influence of plant hormone (usually auxin) and environmental factors, including water and nutrient availability [1, 2]. Genetic and molecular evidence revealed that auxin regulates LR formation by modulating the transcripts of cell cycle regulatory genes, such as cyclins and Cyclin Dependent Kinases (CDK) in the pericycle cells [3–6]. Previous results showed that nitric oxide (NO) mediated the activation of auxin-dependent cell cycle regulatory genes encoding CYCA2;1, CYCA3;1, CDKA1, and the cell cycle inhibitor Kip-Related Protein KRP2 in tomato seedlings at the beginning of LR primordia formation [6]. On the other hand, auxin response factors (ARFs) appeared to play an essential role in auxin-regulated gene expression during plant development, including LR formation, etc. [7–9]. A decade ago, a class of small, non-coding RNAs, called microRNAs (miRNAs), was identified to regulate gene expression [10, 11]. Several miRNAs related to ARFs have been detected via computational approaches [12], such as miR390 targeting ARF2, ARF3 and ARF4 [13], while miR160 targeting ARF10, ARF16 and ARF17 [14].
After NO and carbon monoxide (CO) [15], hydrogen sulfide (H2S) is proposed as the third gaseous messenger to be involved in guard cell signaling [16], root organogenesis [17], and the alleviation of seed germination inhibition caused by heavy metal exposure [18]. In mammalian cells, H2S can be endogenously generated from four enzymes, such as cystathionine-γ-lyase (CSE), cystathionine-β-synthase (CBS), cysteine aminotransferase, and 3-mercaptopyruvate sulfurtransferase (3-MST) [19, 20]. In plants, H2S synthesis is partially catalyzed by L-cysteine desulfhydrase (DES; homolog with CSE in animals) [21, 22]. Related experiments discovered that H2S might be involved in auxin-induced LR formation in tomato seedlings [23]. Importantly, the discovered mechanism of physiological effects achieved by H2S in animals and recently in plants is S-sulfhydration: a posttranslational modification of protein cysteine residues (persulfide R-SSH formation) [24–26]. Above modification manner is opposed to S-nitrosylation, another posttranslational modification of protein cysteine residues by NO with the formation of S-nitrosocysteine residues (R-SNO) [27]. However, whether protein S-sulfhydration was involved in H2S-mediated LR formation, is still unknown.
It was well-known that hydrogen peroxide (H2O2) plays various vital roles in signal transduction beside its toxic effects. In fact, H2O2 is an important product of NADPH oxidase, polyamine oxidases (PAO), and diamine oxidases (DAO), etc. [28, 29]. Subsequent results showed that H2O2 mediates plant responses against adversity stresses and takes part in plant development processes, including stomatal closure [30], root gravitropism [31], and cell elongation [32]. Specially, H2O2 is also involved in auxin signaling [31, 33, 34], adventitious rooting [34, 35], and LR formation [36–39].
Although H2S and H2O2 were respectively suggested to be required for root architecture [17, 32], the cross-talk between H2S and H2O2 in tomato LR development, has not been fully elucidated. In this report, the analysis of H2S-regulated mechanisms leading to LR promotion is expanded. By using pharmacological, anatomic, and molecular approaches, evidence presented here supported the role of RBOH1-mediated H2O2 in the regulation of tomato LR development achieved by H2S. Potential mechanisms, including LR-related ARFs gene expression via miRNAs, are preliminarily elucidated. Additionally, downstream signaling events modulated by H2S might occur in both transcriptional and posttranslational levels (protein S-sulfhydration, etc.). Above results thus provide insights into H2S signaling in plant development.
Results
Increases of endogenous H2O2 contents and LR formation elicited by NaHS
Compared with NaHS alone, the decreased H2S production (determined by spectrophotography) and thereafter the impaired LR formation were previously observed when hypotaurine (HT; a H2S scavenger) was added together with NaHS [23]. To further confirm whether above NaHS response was H2S-dependent, a commercial specific fluorescent probe AzMC for H2S was applied. As expected, when together with HT or DL-propargylglycine (PAG; a synthetic inhibitor of H2S), AzMC-related florescent density and LR formation achieved by NaHS were impaired as well (Fig. 1a–e). Above results clearly confirmed that the response of NaHS in the induction of LR formation was H2S-dependent.
Fig. 1.
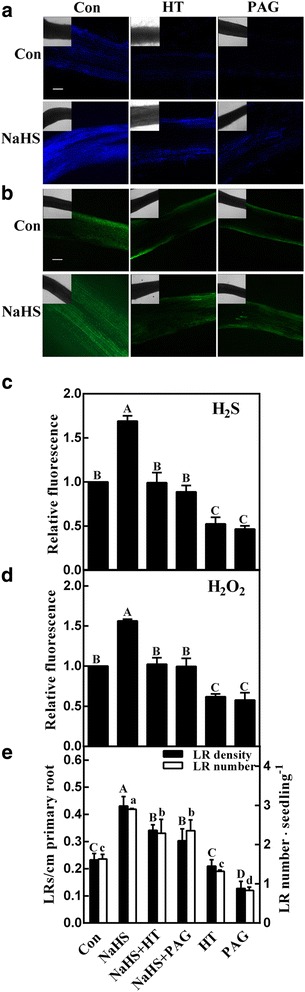
Sodium hydrosulfide (NaHS; the H2S donor) increases H2O2 accumulation and thereafter lateral root (LR) formation. Three-day-old tomato seedlings were treated with H2O (Con), 1 mM NaHS, 200 μΜ hypotaurine (HT), and 2 μΜ DL-propargylglycine (PAG), alone or their combinations. After 12 h, the confocal images of AzMC-dependent and DCF-dependent fluorescence in seedling roots were used to represent endogenous H2S (a) and H2O2 (b) contents. Scale bar = 200 μm. Meanwhile, the relative fluorescence presented as values relative to Con (c, d). Also, the emerged LR density and the number of emerged LRs (>1 mm) per seedling (e) were analyzed with plants 4 d after treatments. Mean and SE values were calculated from at least three independent experiments. Within each set of experiments, bars denoted by the same letter did not differ significantly at P < 0.05 according to Duncan’s multiple range test
Further, seedlings were loaded with reactive oxygen species (ROS)-specific fluorescent dye H2DCF-DA, and laser confocal scanning microscopy (LCSM) was used to investigate changes in ROS-induced fluorescence. Meanwhile, exogenously applied with H2O2 was regarded as a positive control. Figure 2a and b showed the images and quantified the fluorescence levels detected in H2O2-treated seedlings in the presence or absence of DMTU (a H2O2 scavenger) or DPI (an inhibitor of NADPH oxidase). Results revealed that both DMTU and DPI reduced, at least partially, the DCF-dependent fluorescence in the root tissues, consistent with the explanation that some, if not most, of the fluorescence was caused by endogenous H2O2. Thus, the fluorescence was used to report endogenous H2O2 levels throughout this study.
Fig. 2.
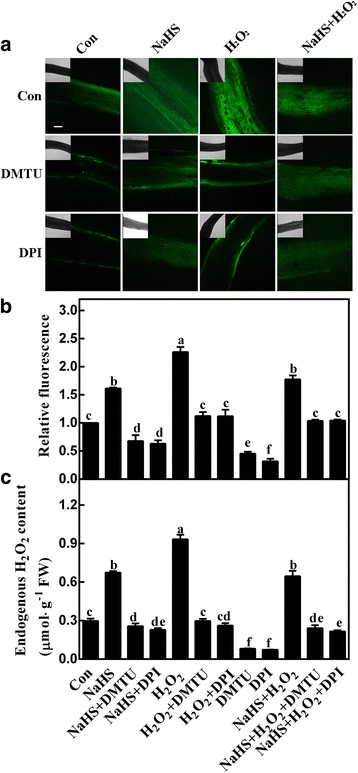
H2S-induced H2O2 accumulation is diminished by the scavenger and synthetic inhibitor of H2O2. Three-day-old tomato seedlings were treated with H2O (Con), 1 mM NaHS, 100 μΜ H2O2, 500 μΜ N,N′-dimethylthiourea (DMTU), and 0.1 μΜ diphenylene idonium (DPI), alone or their combinations for 12 h. Afterwards, corresponding confocal images of DCF-dependent fluorescence in seedling roots were provided to represent endogenous H2O2 contents (a), and the relative fluorescence were presented as values relative to Con (b). Scale bar = 200 μm. Meanwhile, the H2O2 contents were determined by spectrophotography (c). Mean and SE values were calculated from at least three independent experiments. Bars with different letters denoted significant differences at P < 0.05 according to Duncan’s multiple range test
Subsequent results revealed that endogenous H2O2 production was induced as well when NaHS was applied, since the DCF-dependent fluorescence was increased by 56%, compared to the control samples (Fig. 1b, d). By contrast, the addition of HT and PAG weaken above fluorescence induced by NaHS, suggesting that NaHS-induced H2O2 might be obviously blocked by the removal of H2S. Meanwhile, HT or PAG alone, not only decreased corresponding fluorescence, but also inhibited LR formation (Fig. 1e). Combined with the changes in LR density and its number, we thus speculated a potential interrelationship between endogenous H2S and H2O2 during lateral root formation.
H2S-induced tomato lateral rooting is sensitive to the removal of H2O2
To investigate the contribution of H2O2 during LR formation triggered by H2S, DMTU and DPI were used together with NaHS and H2O2 to evaluate tomato LR development. The results shown in Fig. 3 indicated that the addition of DMTU or DPI alone could bring about decreases in LR density (Fig. 3a and b), LR length (Fig. 3c), and LR number (Fig. 3d); while, the primary root (PR) length was increased (Fig. 3e). Further experiment revealed that both NaHS- and H2O2-induced lateral rooting were greatly reduced in the presence of DMTU and/or DPI. Microscopical analysis showed that NaHS- and H2O2-induced LR primordia (LRP) presented a similar anatomic structure, and the inducing effects achieved by NaHS and H2O2 could be apparently prevented by DMTU or DPI (Fig. 4). Above results indicated a hypothesis that endogenous H2O2 might be required for H2S-induced lateral root development. Additionally, no additive responses were found when NaHS and H2O2 were applied together.
Fig. 3.
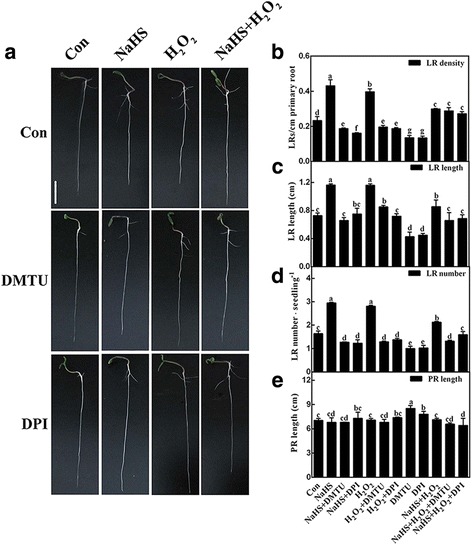
H2S-induced tomato lateral rooting is sensitive to the scavenger and synthetic inhibitor of H2O2. Three-day-old tomato seedlings were treated with H2O (Con), 1 mM NaHS, 100 μΜ H2O2, 500 μΜ N,N′-dimethylthiourea (DMTU), and 0.1 μΜ diphenylene idonium (DPI), alone or their combinations for 4 d. Corresponding photographs were taken (a). Bar = 1 cm. Meanwhile, the emerged LR density (b), LR length (c), the number of emerged LRs (>1 mm) per seedling (d), and primary root (PR) length (e) were analyzed. Mean and SE values were calculated from at least three independent experiments. Bars denoted by the same letter did not differ significantly at P < 0.05 according to Duncan’s multiple range test
Fig. 4.
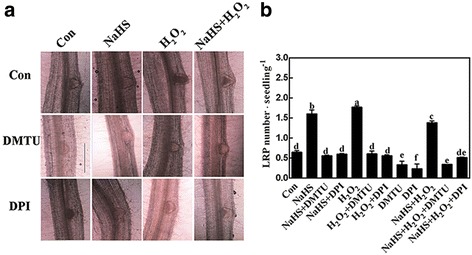
H2S-induced lateral root primordial (LRP) formation is sensitive to the removal of H2O2. Three-day-old tomato seedlings were treated with H2O (Con), 1 mM NaHS, 100 μΜ H2O2, 500 μΜ N,N′-dimethylthiourea (DMTU), and 0.1 μΜ diphenylene idonium (DPI), alone or their combinations. After various treatments for 3 d, photographs showing the representative morphology of LRP (about 75% of LRP at the shown stages), were taken (a). Bar = 0.25 mm. Meanwhile, the number of emerged LRP was also analyzed (b). Mean and SE values were calculated from at least three independent experiments. Bars with different letters denoted significant differences at P < 0.05 according to Duncan’s multiple range test
H2O2 is required for lateral root formation triggered by H2S
The role of H2O2 in H2S-induced lateral root development was further examined by monitoring H2O2 synthesis in response to applied NaHS. As expected, a significant increase in H2O2-related fluorescence was observed in NaHS-treated tomato seedling roots compared with control sample (P < 0.05), suggesting H2S-mediated H2O2 production (Fig. 2a and b). This deduction was confirmed by the co-treatment with DMTU and DPI. We also noticed that when NaHS was together with H2O2, there is no additive response in the fluorescence. The changes of endogenous H2O2 detected with spectrophotography showed the similar tendencies (Fig. 2C).
In order to assess the possible source(s) of H2O2, we thus evaluated the expression of RBOH1, the key gene responsible for H2O2 synthesis in tomato seedling roots [40]. As expected, a significant increase of RBOH1 expression was observed when tomato seedlings were incubated with NaHS, and the up-regulation of RBOH1 transcript was reversed by DMTU or DPI (Fig. 5). Meanwhile, a significant but weaker induction in RBOH1 transcript was observed in response to the addition of H2O2 with or without NaHS. Above results indicated that H2O2 might be required for LR formation elicited by H2S.
Fig. 5.
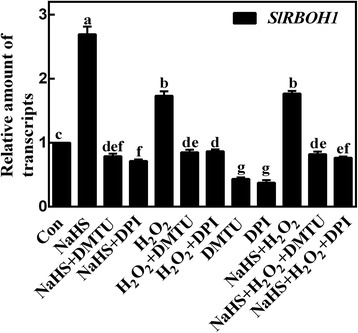
H2S modulates the expression of SlRBOH1. Three-day-old tomato seedlings were treated with H2O (Con), 1 mM NaHS, 100 μΜ H2O2, 500 μΜ N,N′-dimethylthiourea (DMTU), and 0.1 μΜ diphenylene idonium (DPI), alone or their combinations, for 6 h. Afterwards, the amount of transcript were analyzed by qPCR, and presented relative to the Con. Mean and SE values were calculated from at least three independent experiments. Bars with different letters denoted significant differences at P < 0.05 according to Duncan’s multiple range test
H2O2 modulates the expression of cell cycle regulatory genes in H2S-induced LR formation
To further study the potential relationship between H2O2 and H2S in the induction of LR formation, the influence of NaHS, H2O2, DMTU, and DPI applied alone or their combination on the expression of cell cycle regulatory genes, was analyzed by qPCR. Similar to the inducible effects triggered by H2O2, NaHS resulted in the up-regulation of SlCYCA2;1, SlCYCA3;1, and SlCDKA1 transcripts, together with simultaneous down-regulation of SlKRP2 transcripts (Fig. 6). However, DMTU or DPI significantly blocked above mentioned modulation in these transcripts triggered by treatments with NaHS and/or H2O2. These results indicated that H2S-triggered LR formation was likely to be achieved by up-regulation of H2O2-mediated cycle regulatory genes.
Fig. 6.
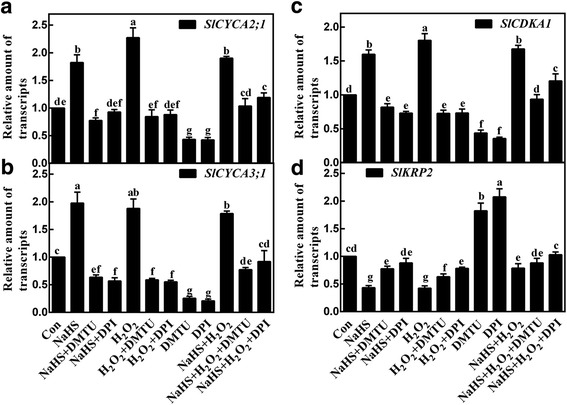
H2S affects the expression of SlCYCA2;1, SlCYCA3;1, SlCDKA1, and SlKRP2. Three-day-old tomato seedlings were treated with H2O (Con), 1 mM NaHS, 100 μΜ H2O2, 500 μΜ N,N′-dimethylthiourea (DMTU), and 0.1 μΜ diphenylene idonium (DPI), alone or their combinations for 12 h. Afterwards, SlCYCA2;1 (a), SlCYCA3;1 (b), SlCDKA1 (c), and SlKRP2 (d) transcript levels were analyzed by qPCR, and presented relative to the Con. Mean and SE values were calculated from at least three independent experiments. Bars with different letters denoted significant differences at P < 0.05 according to Duncan’s multiple range test
Expression of miRNAs and their target genes
In the subsequent experiments, several LR formation-related miRNAs and their target genes were investigated to check whether they were involved in H2S-triggered LR development. Results shown in Fig. 7 revealed that both NaHS and H2O2 up-regulated miR390a and miR160 transcripts; while, their corresponding target genes, including SlARF4 and SlARF16, were significantly reduced. Contrasting changes were observed when NaHS or H2O2 was added together with DMTU or DPI. Above results confirmed the opposite effects between changes in miRNAs and their target genes.
Fig. 7.
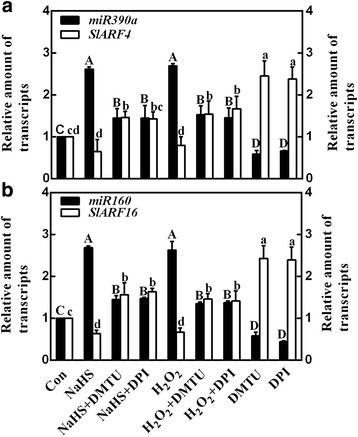
H2S affects the expression of microRNAs and their target genes. Three-day-old tomato seedlings were treated with H2O (Con), 1 mM NaHS, 100 μΜ H2O2, 500 μΜ N,N′-dimethylthiourea (DMTU), and 0.1 μΜ diphenylene idonium (DPI), alone or their combinations for 12 h. Meanwhile, miR390a (a; black), SlARF4 (a; white), miR160 (b; black), and SlARF16 (b; white) transcript levels were analyzed by qPCR, and presented relative to the Con. Mean and SE values were calculated from at least three independent experiments. Within each set of experiments, bars with different letters denoted significant differences at P < 0.05 according to Duncan’s multiple range test
Detection of S-sulfhydrated proteins in H2S-treated tomato
To further analyze the molecular mechanism underlying H2S signaling in LR formation, the pattern of S-sulfhydrated proteins in tomato roots was analyzed by using the modified biotin switch method. The results shown in Fig. 8a illustrated that treatment of tomato root extraction with Na2S (another H2S donor; [26]) enhanced S-sulfhydration, which was alleviated by DTT (a sulfhydration inhibitor; [24]). Consistently, tomato seedlings were treated with NaHS, HT, and PAG, alone or their combinations, then root extracts were used to analysis S-sulfhydrated profiles (Fig. 8B). Similarly, NaHS increased the level of S-sulfhydrated proteins, which was partially blocked by HT or PAG. Additionally, in compared with the control samples, HT or PAG alone slightly decreased sulfhydration.
Fig. 8.
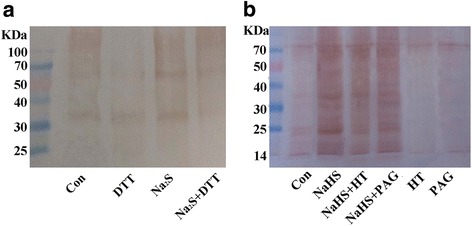
Detection of S-sulfhydrated proteins. a Protein extracts from 0.25 g of tomato roots were exogenously treated with H2O (Con), 2 mM Na2S (for 1 h; another H2S donor), and 2 mM DTT (for 30 min; a sulfhydration inhibitor), alone or their combinations (treatment with Na2S followed by DTT), and subjected to the modified biotin switch method (BSM). Finally, the labeled proteins were detected using protein blot analysis with antibodies against biotin. b Three-day-old tomato seedlings were treated with H2O (Con), 1 mM NaHS, 200 μΜ hypotaurine (HT), and 2 μΜ DL-propargylglycine (PAG), alone or their combinations for 4 d. Afterwards, protein extracts from 0.25 g of seedling roots were subjected to the BSM, and the labeled proteins were detected using protein blot analysis with antibodies against biotin. Representative pictures were provided
Discussion
H2S is proposed as the third gas messenger after NO and CO to fulfill many important roles in plants, including the inducement of LR formation [23, 41, 42]. The important function of H2O2 in the auxin-induced LR formation was also illustrated [39]. Although H2O2 involved in H2S-induced salt tolerance pathway of the Arabidopsis root was discovered [43], the relationship between H2S and H2O2 in LR formation is largely unclear. Here, we provided evidence for a previously unknown role for H2O2 in H2S-triggered LR formation in tomato seedlings.
Firstly, our results showed that an increase in the concentration of endogenous H2O2 determined by spectrophotography and LSCM, is one of the earliest responses involved in the signaling pathway governing LR formation triggered by H2S (Figs. 1 and 2). These results are in agreement with those obtained in Arabidopsis subjected to salinity stress [43], showing that NaHS induced a gradual elevation of H2O2 in NaCl-stressed seedling roots. This is an important point, since H2O2 is regarded as one of the ubiquitous components of the signaling transduction pathway [29], including responsible for the induction of LR formation [38, 39, 44] and adventitious rooting [34, 35].
Further pharmacological and microscopical evidence revealed the requirement of endogenous H2O2 in the induction of tomato LR formation triggered by H2S. This conclusion is based on several pieces of evidence: (i) the removal of endogenous H2O2 by its membrane-permeable scavenger DMTU impaired the induction of LR formation elicited by H2S (Figs. 2 and 3); (ii) the similar inhibiting responses triggered by DPI, an inhibitor of NADPH oxidase, in H2S-induced H2O2 production (Fig. 2) and thereafter LRP formation and lateral rooting (Figs. 3 and 4) were significant, implying the involvement of tomato RBOH1, at least partially. Changes in SlRBOH1 transcripts confirmed this deduction (Fig. 5). Certainly, other candidate(s) for H2O2 synthesis (such as PAO and DAO; [36]) could not be easily ruled out in this process. Although we can not exclude the possibility that above mentioned chemicals may not specifically target H2O2, above results clearly indicated that H2O2 might be the downstream messenger of H2S signaling responsible for LR formation. This deduction was consistent with the recent genetic results [45], showing that RBOH-mediated ROS production facilitated LR emergence in Arabidopsis.
Strong evidence proved that the expression of cell cycle regulatory genes plays important roles in the early LR initiation in the presence of auxin and NO [3, 4, 6]. Similar to the previous results [23], our further molecular evidence revealed that H2S could modulate four cell cycle regulatory genes, including SlCYCA2;1, SlCYCA3;1 SlCDKA1 and SlKRP2, mimicking the actions of H2O2 (Fig. 6). By contrast, the blocking effects were observed when DMTU or DPI was respectively supplemented together with H2S and/or H2O2. Combined with the changes in phenotypes (Fig. 3), we further speculated that H2S-triggered H2O2 was important in the early LR initiation by targeting cell cycle regulatory genes.
It is well-known that plant miRNAs play an important role in leaf morphogenesis [46], leaf polarity [47, 48], flowering time [49, 50], and flower development [51]. Some studies also focused on miRNAs related to plant root organogenesis [52]. For example, Marin et al. [13] and Yoon et al. [53] revealed that miR390 and AUXIN RESPONSE FACTORS (ARFs) formed an auxin-responsive regulatory network (miR390-TAS3-ARF2/ARF3/ARF4) controlling lateral root development. Another miRNA, miR160, was confirmed to have a positive role in the induction of LR formation via targeting ARF16 in Arabidopsis [54]. Thus, several representative miRNAs correlated with ARFs and LR formation [52], including miR390a for SlARF4 [55], and miR160 for SlARF16 [54], were chosen. In this study, the results of qPCR revealed that miR390a and miR160 transcripts were increased by both H2S and H2O2, and contrasting changes were observed in their target genes, including SlARF4 and SlARF16 (Fig. 7). Above mentioned changes were obviously prevented by the removal of endogenous H2O2 when DMTU or DPI was added together. These results were consistent with the changes in endogenous H2O2 levels (Fig. 2) and thereafter LR formation (Fig. 3). Thus, we deduced that auxin signaling mediated by H2O2-elicited miRNAs expression might be the important mechanism responsible for LR formation triggered by H2S. Certainly, corresponding genetic evidence should be investigated in the near future.
Recently, H2S-dependent S-sulfhydration, the conversion of cysteine -SH residues to persulfide (−SSH) which could be detected by using a modified biotin switch method, has been described to play a vital role in mammalians and plants [26, 27]. Nevertheless, whether S-sulfhydration was involved in plant LR formation is still unknown. In our experimental conditions, the S-sulfhydration conditions were strengthened when protein extracts from tomato seedling roots were treated with Na2S (another H2S donor; [26]), and the addition of DTT (a sulfhydration inhibitor; [24]) impaired above effect (Fig. 8a). Since DTT could reduce disulfide bonds, our results suggested that the modification is covalent and involves a sulfhydryl group. Similar results were obtained when tomato seedlings were subjected to the chemicals related to the alternation of endogenous H2S homeostasis (Fig. 8b). Thus, combine with the corresponding phenotypes in LR formation (Fig. 1), we provided a preliminary finding, that S-sulfhydration might be involved in H2S-promoted LR formation, although the specific S-sulfhydrated protein(s) had not been purified and elucidated. In fact, Aroca et al. [26] identified a total of 106 S-sulfhydrated proteins in Arabidopsis, and some of the proteins (ascorbate peroxidase and catalase; etc) identified were related to reactive oxygen species (ROS) metabolism. Since it was shown that ROS acted downstream of auxin action in the development of LR emergence [45], and ascorbate peroxidase (APX; a scavenging enzyme of H2O2) was previously confirmed to be S-sulfhydrated [26], the genetic and biochemical (in vitro and in vivo tests) approaches combined with proteomic and transcriptomic analyses should be applied to check whether APX acts as the S-sulfhydrated target of H2S signaling related to LR formation.
Conclusions
In summary, the results of this investigation indicated that an increase in H2O2 production might be an early response of H2S that contributes to the induction of LR formation by (i) modulating the expression of cell cycle regulatory genes; (ii) regulating auxin signaling mediated by miRNAs expression; and (iii) at least partially involving S-sulfhydrated proteins (Fig. 9). Additionally, our results provide indications of transcriptional and post-translational regulatory mechanism that contributed to the development of LR formation elicited by H2S.
Fig. 9.
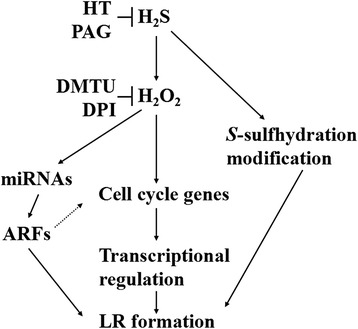
Schematic representation of the proposed model involving H2O2 homeostasis during H2S-triggered LR formation. The above pathway might be mediated by the expression of cell cycle genes in tomato seedlings. The involvement of miRNAs expression and S-sulfhydration modification were also suggested by solid lines. The possibility was suggested by dashed lines. T bars, inhibition
Methods
Chemicals
All chemicals were purchased from Sigma (St Louis, MO, USA) unless stated otherwise. Sodium hydrosulfide (NaHS) was used at 1 mM as a H2S donor. 200 μM hypotaurine (HT; an H2S scavenger) and 2 μM DL-propargylglycine (PAG; a synthetic inhibitor of H2S) were also used. Hydrogen peroxide (H2O2) was applied at 100 μM. N,N′-dimethylthiourea (DMTU), a scavenger of H2O2, was used at 500 μM. 0.1 μM diphenylene idonium (DPI) was regarded as an inhibitor of H2O2 synthetic enzyme (NADPH oxidase). A H2S fluorescent probe 7-azido-4-methylcoumarin (AzMC) and a reactive oxygen species (ROS) fluorescent probe 2′,7′-dichlorofluorescein diacetate (H2DCF-DA) were both used at a final concentration of 20 μM. According to our pilot experiments, the concentration of above chemicals exhibiting the effective responses was chosen.
Plant material and growth conditions
Tomato (Solanum lycopersicum L.) seeds “baiguoqiangfeng” were surface-sterilized in 2% sodium hypochlorite for 6 min, rinsed extensively and germinated in distilled water at 25 ± 1 °C in the dark for 3 days. Afterwards, the selected identical seedlings with radicles 2–3 mm were transferred to 4 ml treatment solutions containing the indicated chemicals and grown in an illuminating incubator (25 ± 1 °C) with a light intensity of 200 μmol m−2 s−1 at 14/10 h (light/dark) photoperiod.
After treatments for 4 d or the indicated time points, photographs were taken. Meanwhile, according to the previous methods [23, 39], the number of emerged lateral roots (LRs; >1 mm) per seedling, the length of primary root (PR), the length of LR and the emerged LR density (the number of LR per cm primary root; LRs/cm) were determined by using Image J software. Additionally, LR primordial (LRP) per seedling were observed after 3 d of treatments by root squash preparations and quantified with a light microscope. Unless stated otherwise, only the lateral root-inducible segments were used for the subsequent biochemical and molecular analyses. Thus, the root apical meristems were cut off, and the shoots of seedlings were removed by cutting below the root-shoot junction.
Laser scanning confocal microscopy (LSCM)
According to the previous methods with minor modification [56, 57], endogenous H2O2 and H2S production were determined by a laser scanning confocal microscope (LSCM) using the ROS fluorescent probe H2DCF-DA and a H2S fluorescent probe AzMC. After treatments, roots were incubated in 20 mM HEPES-NaOH buffer (pH 7.5) containing 20 μM probe for 30 min in dark (25 °C). Afterwards, the roots were washed three times (15 min each time) with fresh HEPES buffer, and observed by using Zeiss LSM 710 confocal microscope (Carl Zeiss, Oberkochen, Germany) with the same exposure time.
All manipulations were performed at 25 °C. Each photograph were taken at the eyepiece 5 × magnification based on 20 overlapping confocal planes of 15 μm each using ZEN software (300 μm sections along Z stack). For each picture, the overall fluorescence of maturation zone of the primary root (about an area of 500,000 μm2), where cells become differentiated, and at a later stage lateral roots emerge, was quantified [9]. The bright-field (BF) images corresponding to the fluorescent images were also shown at the top left corners of the photograph. Representative photographs with similar results were obtained after the analysis of at least fifteen samples for each experiment. Afterwards, the average intensities of 15 photographs (1 photograph per sample) for each treatment were calculated. The relative fluorescence was presented as values relative to control sample.
Measurement of H2O2 content
The content of H2O2 was analyzed by the FOX1 method [58, 59]. Samples were extracted with 200 mM perchloric acid (HClO4). After centrifugation at 4 °C, 10,000 g for 15 min, 500 μL supernatant was transferred to 500 μL assay solution containing 500 μM ammonium ferrous sulfate, 50 mM H2SO4, 200 μM xylenol orange, and 200 mM sorbitol, for 45 min in dark (25 °C). Afterwards, the absorbance values were detected at 560 nm. The specificity for H2O2 was tested by eliminating H2O2 in the reaction mixture with catalase (CAT). Standard curves of H2O2 were obtained for each independent experiment by adding variable amounts of H2O2.
Quantitative real-time RT-PCR (qPCR) analysis
qPCR was used to analyze the expression of cell cycle regulatory genes, ARFs genes, and miRNA. After various treatments, total RNA from about 100 mg (fresh weight) samples was isolated by using Trizol reagent (Invitrogen, Gaithersburg, MD, USA). Afterwards, the RNA samples were reverse-transcribed using an oligo d(T) primer and M-MLV reverse transcriptase (BioTeke, Beijing, China). Quantitative RT-PCR reactions were performed using a Mastercycler® ep realplex real-time PCR system (Eppendorf, Hamburg, Germany) with SYBR® Premix Ex Taq™ (TransGen Biotech, Beijing, China) according to the manufacturer’s instructions. The accession numbers (GenBank/miRBase) and oligonucleotide primers were shown in Additional file 1: Table S1. Three biological and three technological repeats were performed in qPCR. Relative expression levels of corresponding genes were calculated by using the 2−ΔΔC T method [60, 61], and were presented as values relative to that of corresponding control samples at the indicated times, after normalization with Actin and GAPDH transcript levels.
A One Step PrimeScript miRNA cDNA synthesis kit (TaKaRa Bio Inc., Dalian, China) was used to synthesize cDNA for analyzing miRNA expression by qPCR. The specific 5′ primers were listed in Additional file 1: Table S1. The 3′ primer was supplied in the kit. U6 snRNA was used as internal control. The rest steps were the same as the approaches described previously [62].
Modified biotin switch method
The modified biotin switch method was carried out as previously described protocol with minor modification [26, 63]. Total proteins extracted from samples were homogenized in HEN buffer containing 250 mM Hepes-NaOH (pH 7.7), 1 mM EDTA and 0.1 mM neocuproine, and centrifuged at 10000 g for 15 min at 4 °C. The supernatant was transferred to fresh tubes, and added with three volumes of blocking buffer (HEN buffer supplemented with 2.5% SDS and 20 mM methyl methanethiosulfonate (MMTS)). Then, the solution was incubated at 4 °C for 12 h to block free sulfhydryl groups. The MMTS was then removed, and ice-cold acetone was used to precipitate the proteins at −20 °C for 20 min. After the removal of acetone, the proteins were resuspended in HENS buffer (HEN buffer supplemented with 1% SDS). Afterwards, the S-sulfhydrated proteins were labeled using 4 mM N-[6-(biotinamido)hexyl]-3′-(2′-pyridyldithio)propionamide (Biotin-HPDP) for 3 h at 25 °C in the dark.
The above biotin-labeled proteins were separated using non-reducing SDS-PAGE on 12% polyacrylamide gels. Then, the proteins were transferred to polyvinylidene fluoride membranes (Roche, Basel, Switzerland) according to the manufacturer’s instructions. Anti-biotin antibody (HRP) (Abcam antibodies, Cambridge, UK) was diluted 1:10,000. Meanwhile, Coomassie Brilliant Blue-stained gels were used to confirm the equal amounts of proteins loaded (data not shown).
Statistical analysis
All results were shown as the mean values ± SE of at least three independent experiments with at least three biological replicates for each. By using SPSS 17.0 software, data was analyzed by one-way analysis of variance (ANOVA) followed by Duncan’s multiple range test, and P values <0.05 were considered statistically significant.
Acknowledgements
None.
Funding
This work was supported by the National Natural Science Foundation of China (31772292), the Natural Science Foundation of Jiangsu Province of China (BK20141361), and the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD).
Availability of data and materials
All relevant data are within this article and its supporting information files.
Abbreviations
- 3-MST
3-Mercaptopyruvate sulfurtransferase
- APX
Ascorbate peroxidase
- ARFs
Auxin response factors
- AzMC
7-Azido-4-methylcoumarin
- Biotin-HPDP
N-[6-(Biotinamido)hexyl]-3′-(2′-pyridyldithio)propionamide
- CAT
Catalase
- CBS
Cystathionine-β-synthase
- CDK
Cyclin Dependent Kinases
- CO
Carbon monoxide
- CSE
Cystathionine-γ-lyase
- DAO
Diamine oxidases
- DMTU
Dimethylthiourea
- DPI
Diphenylene idonium
- H2DCF-DA
2′,7′-Dchlorofluorescein diacetate
- H2O2
Hydrogen peroxide
- H2S
-
Hydrogen sulfide
HRP
Anti-biotin antibody
- HT
Hypotaurine
- LR
Lateral root
- LRP
Lateral root primordial
- LSCM
Laser scanning confocal microscopy
- MiRNAs
MicroRNAs;
- MMTS
Methyl methanethiosulfonate
- NaHS
Sodium hydrosulfide
- NO
Nitric oxide
- PAG
DL-Propargylglycine
- PAO
Polyamine oxidases
- PR
Primary root
- ROS
Reactive oxygen species
- qPCR
Quantitative real-time RT-PCR
Additional file
The accession numbers and primer sequences of real-time RT-PCR (qPCR). (DOC 45 kb)
Authors’ contributions
Conception and design of the study: LH. Acquisition of data for the study: YM, HC, WS1, and WS2. Analysis of data for the work: YM and WS1. Interpretation of data for the work: YM, HC, WS1, WS2, and LH. All authors read and approved the final manuscript.
Ethics approval and consent to participate
Tomato (Solanum lycopersicum L.) seeds “baiguoqiangfeng” were purchased from Jiangsu Academy of Agricultural Sciences. The Tomato seeds have obtained the permission of Ministry of Agriculture of the People’s Republic of China, which the phytosanitary certificate number is 620,900,200,000,857.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Footnotes
Electronic supplementary material
The online version of this article (10.1186/s12870-017-1110-7) contains supplementary material, which is available to authorized users.
Contributor Information
Yudong Mei, Email: 2015116112@njau.edu.cn.
Haotian Chen, Email: 1689554589@qq.com.
Wenbiao Shen, Email: wbshenh@njau.edu.cn.
Wei Shen, Email: swgdj@njau.edu.cn.
Liqin Huang, Phone: +86 25 8439 6524, Email: lqhuangs@njau.edu.cn.
References
- 1.Casimiro I, Marchant A, Bhalerao RP, Beeckman T, Dhooge S, Swarup R, et al. Auxin transport promotes Arabidopsis lateral root initiation. Plant Cell. 2001;13:843–852. doi: 10.1105/tpc.13.4.843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Malamy JE. Intrinsic and environmental response pathways that regulate root system architecture. Plant Cell Environ. 2005;28:67–77. doi: 10.1111/j.1365-3040.2005.01306.x. [DOI] [PubMed] [Google Scholar]
- 3.Himanen K, Boucheron E, Vanneste S, de Almeida EJ, Inzé D, Beeckman T. Auxin-mediated cell cycle activation during early lateral root initiation. Plant Cell. 2002;14:2339–2351. doi: 10.1105/tpc.004960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Himanen K, Vuylsteke M, Vanneste S, Vercruysse S, Boucheron E, Alard P, et al. Transcript profiling of early lateral root initiation. Proc Natl Acad Sci U S A. 2004;101:5146–5151. doi: 10.1073/pnas.0308702101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Casimiro I, Beeckman T, Graham N, Bhalerao R, Zhang H, Casero P, et al. Dissecting Arabidopsis lateral root development. Trends Plant Sci. 2003;8:165–171. doi: 10.1016/S1360-1385(03)00051-7. [DOI] [PubMed] [Google Scholar]
- 6.Correa-Aragunde N, Graziano M, Chevalier C, Lamattina L. Nitric oxide modulates the expression of cell cycle regulatory genes during lateral root formation in tomato. J Exp Bot. 2006;57:581–588. doi: 10.1093/jxb/erj045. [DOI] [PubMed] [Google Scholar]
- 7.Guilfoyle TJ, Ulmasov T, Hagen G. The ARF family of transcription factors and their role in plant hormone-responsive transcription. Cel Mol Life Sci. 1998;54:619–627. doi: 10.1007/s000180050190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Lee HW, Kim NY, Lee DJ, Kim J. LBD18/ASL20 regulates lateral root formation in combination with LBD16/ASL18 downstream of ARF7 and ARF19 in Arabidopsis. Plant Physiol. 2009;151:1377–1389. doi: 10.1104/pp.109.143685. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Zhu D, Mei Y, Shi Y, Hu D, Ren Y, Gu Q, et al. Involvement of glutathione in β-cyclodextrin-hemin complex-induced lateral root formation in tomato seedlings. J Plant Physiol. 2016;204:92–100. doi: 10.1016/j.jplph.2016.07.015. [DOI] [PubMed] [Google Scholar]
- 10.He L, Hannon GJ. MicroRNAs: small RNAs with a big role in gene regulation. Nat Rev Genet. 2004;5:522–531. doi: 10.1038/nrg1379. [DOI] [PubMed] [Google Scholar]
- 11.Megraw M, Baev V, Rusinov V, Jensen ST, Kalantidis K, Hatzigeorgiou AG. MicroRNA promoter element discovery in Arabidopsis. RNA. 2006;12:1612–1619. doi: 10.1261/rna.130506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Zhang B, Pan X, Wang Q, Cobb GP, Anderson TA. Computational identification of microRNAs and their targets. Comput Biol Chem. 2006;30:395–407. doi: 10.1016/j.compbiolchem.2006.08.006. [DOI] [PubMed] [Google Scholar]
- 13.Marin E, Jouannet V, Herz A, Lokerse AS, Weijers D, Vaucheret H, et al. miR390, Arabidopsis TAS3 tasiRNAs, and their AUXIN RESPONSE FACTOR targets define an autoregulatory network quantitatively regulating lateral root growth. Plant Cell. 2010;22:1104–1117. doi: 10.1105/tpc.109.072553. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Mallory AC, Bartel DP, Bartel B. MicroRNA-directed regulation of Arabidopsis AUXIN RESPONSE FACTOR17 is essential for proper development and modulates expression of early auxin response genes. Plant Cell. 2005;17:1360–1375. doi: 10.1105/tpc.105.031716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Wang R. Two’s company, three’s a crowd: can H2S be the third endogenous gaseous transmitter? FASEB J. 2002;16:1792–1798. doi: 10.1096/fj.02-0211hyp. [DOI] [PubMed] [Google Scholar]
- 16.García-Mata C, Lamattina L. Hydrogen sulphide, a novel gasotransmitter involved in guard cell signalling. New Phytol. 2010;188:977–984. doi: 10.1111/j.1469-8137.2010.03465.x. [DOI] [PubMed] [Google Scholar]
- 17.Zhang H, Tang J, Liu XP, Wang Y, Yu W, Peng WY, et al. Hydrogen sulfide promotes root organogenesis in Ipomoea batatas, Salix matsudana, and Glycine max. J Integr Plant Biol. 2009;51:1086–1094. doi: 10.1111/j.1744-7909.2009.00885.x. [DOI] [PubMed] [Google Scholar]
- 18.Zhang H, Hu LY, Hu KD, He YD, Wang SH, Luo JP. Hydrogen sulfide promotes wheat seed germination and alleviates oxidative damage against copper stress. J Integr Plant Biol. 2008;50:1518–1529. doi: 10.1111/j.1744-7909.2008.00769.x. [DOI] [PubMed] [Google Scholar]
- 19.Olas B. Hydrogen sulfide in signaling pathways. Clin Chim Acta. 2015;439:212–218. doi: 10.1016/j.cca.2014.10.037. [DOI] [PubMed] [Google Scholar]
- 20.Guo H, Xiao T, Zhou H, Xie Y, Shen W. Hydrogen sulfide: a versatile regulator of environmental stress in plants. Acta Physiol Plant. 2016;38:1–13. doi: 10.1007/s11738-015-2023-4. [DOI] [Google Scholar]
- 21.Papenbrock J, Riemenschneider A, Kamp A, Schulz-Vogt HN, Schmidt A. Characterization of cysteine-degrading and H2S-releasing enzymes of higher plants-from the field to the test tube and back. Plant Biol (Stuttg) 2007;9:582–588. doi: 10.1055/s-2007-965424. [DOI] [PubMed] [Google Scholar]
- 22.Álvarez C, Calo L, Romero LC, García I, Gotor C. An O-acetylserine(thiol)lyase homolog with L-cysteine desulfhydrase activity regulates cysteine homeostasis in Arabidopsis. Plant Physiol. 2010;152:656–669. doi: 10.1104/pp.109.147975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Fang T, Cao Z, Li J, Shen W, Huang L. Auxin-induced hydrogen sulfide generation is involved in lateral root formation in tomato. Plant Physiol Biochem. 2014;76:44–51. doi: 10.1016/j.plaphy.2013.12.024. [DOI] [PubMed] [Google Scholar]
- 24.Mustafa AK, Gadalla MM, Sen N, Kim S, Mu W, Gazi SK, et al. H2S signals through protein S-sulfhydration. Sci Signal. 2009;2:ra72. doi: 10.1126/scisignal.2000464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Paul BD, Snyder SH. H2S signalling through protein sulfhydration and beyond. Nat Rev Mol Cell Biol. 2012;13:499–507. doi: 10.1038/nrm3391. [DOI] [PubMed] [Google Scholar]
- 26.Aroca Á, Serna A, Gotor C, Romero LC. S-sulfhydration: a cysteine posttranslational modification in plant systems. Plant Physiol. 2015;168:334–342. doi: 10.1104/pp.15.00009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Lu C, Kavalier A, Lukyanov E, Gross SS. S-sulfhydration/desulfhydration and S-nitrosylation/denitrosylation: a common paradigm for gasotransmitter signaling by H2S and NO. Methods. 2013;62:177–181. doi: 10.1016/j.ymeth.2013.05.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Bouchereau A, Aziz A, Larher F, Martin-Tanguy J. Polyamines and environmental challenges: recent developments. Plant Sci. 1999;140:103–125. doi: 10.1016/S0168-9452(98)00218-0. [DOI] [Google Scholar]
- 29.Neill S, Desikan R, Hancock J. Hydrogen peroxide signaling. Curr Opin Plant Biol. 2002;5:388–395. doi: 10.1016/S1369-5266(02)00282-0. [DOI] [PubMed] [Google Scholar]
- 30.Pei ZM, Murata Y, Benning G, Thomine S, Klüsener B, Allen GJ, et al. Calcium channels activated by hydrogen peroxide mediate abscisic acid signalling in guard cells. Nature. 2000;406:731–734. doi: 10.1038/35021067. [DOI] [PubMed] [Google Scholar]
- 31.Joo JH, Bae YS, Lee JS. Role of auxin-induced reactive oxygen species in root gravitropism. Plant Physiol. 2001;126:1055–1060. doi: 10.1104/pp.126.3.1055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Foreman J, Demidchik V, Bothwell JH, Mylona P, Miedema H, Torres MA, et al. Reactive oxygen species produced by NADPH oxidase regulate plant cell growth. Nature. 2003;422:442–446. doi: 10.1038/nature01485. [DOI] [PubMed] [Google Scholar]
- 33.Song YJ, Joo JH, Ryu HY, Lee JS, Bae YS, Nam KH. Reactive oxygen species mediate IAA-induced ethylene production in mungbean (Vigna radiata L.) hypocotyls. J Plant Biol. 2007;50:18–23. doi: 10.1007/BF03030595. [DOI] [Google Scholar]
- 34.Bai X, Todd CD, Desikan R, Yang Y, Hu X. N-3-oxo-decanoyl-L-homoserine-lactone activates auxin-induced adventitious root formation via hydrogen peroxide- and nitric oxide-dependent cyclic GMP signaling in mung bean. Plant Physiol. 2012;158:725–736. doi: 10.1104/pp.111.185769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Li S, Xue L, Xu S, Feng H, An L. Hydrogen peroxide involvement in formation and development of adventitious roots in cucumber. Plant Growth Regul. 2007;52:173–180. doi: 10.1007/s10725-007-9188-9. [DOI] [Google Scholar]
- 36.Su GX, Zhang WH, Liu YL. Involvement of hydrogen peroxide generated by polyamine oxidative degradation in the development of lateral roots in soybean. J Integr Plant Biol. 2006;48:426–432. doi: 10.1111/j.1744-7909.2006.00236.x. [DOI] [Google Scholar]
- 37.Wang P, Du Y, Li Y, Ren D, Song CP. Hydrogen peroxide-mediated activation of MAP kinase 6 modulates nitric oxide biosynthesis and signal transduction in Arabidopsis. Plant Cell. 2010;22:2981–2998. doi: 10.1105/tpc.109.072959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Chen YH, Chao YY, Hsu YY, Kao CH. Heme oxygenase is involved in H2O2-induced lateral root formation in apocynin-treated rice. Plant Cell Rep. 2013;32:219–226. doi: 10.1007/s00299-012-1356-3. [DOI] [PubMed] [Google Scholar]
- 39.Ma F, Wang L, Li J, Samma MK, Xie Y, Wang R, et al. Interaction between HY1 and H2O2 in auxin-induced lateral root formation in Arabidopsis. Plant Mol Biol. 2013;85:49–61. doi: 10.1007/s11103-013-0168-3. [DOI] [PubMed] [Google Scholar]
- 40.Yi C, Yao K, Cai S, Li H, Zhou J, Xia X, et al. High atmospheric carbon dioxide-dependent alleviation of salt stress is linked to RESPIRATORY BURST OXIDASE 1 (RBOH1)-dependent H2O2 production in tomato (Solanum lycopersicum) J Exp Bot. 2015;66:7391–7404. doi: 10.1093/jxb/erv435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Correa-Aragunde N, Graziano M, Lamattina L. Nitric oxide plays a central role in determining lateral root development in tomato. Planta. 2004;218:900–905. doi: 10.1007/s00425-003-1172-7. [DOI] [PubMed] [Google Scholar]
- 42.Cao ZY, Xuan W, Liu ZY, Li XN, Zhao N, Xu P, et al. Carbon monoxide promotes lateral root formation in rapeseed. J Integr Plant Biol. 2007;49:1070–1079. doi: 10.1111/j.1672-9072.2007.00482.x. [DOI] [Google Scholar]
- 43.Li J, Jia H, Wang J, Cao Q, Wen Z. Hydrogen sulfide is involved in maintaining ion homeostasis via regulating plasma membrane Na+/H+ antiporter system in the hydrogen peroxide-dependent manner in salt-stress Arabidopsis Thaliana root. Protoplasma. 2014;251:899–912. doi: 10.1007/s00709-013-0592-x. [DOI] [PubMed] [Google Scholar]
- 44.Cao Z, Fang T, Chen M, Li J, Shen W, Huang L. Involvement of haem oxygenase-1 in hydrogen peroxide-induced lateral root formation in tomato. Acta Physiol Plant. 2014;36:931–943. doi: 10.1007/s11738-013-1472-x. [DOI] [Google Scholar]
- 45.Orman-Ligeza B, Parizot B, de Rycke R, Fernandez A, Himschoot E, Van Breusegem F, et al. RBOH-mediated ROS production facilitates lateral root emergence in Arabidopsis. Development. 2016;143:3328–3339. doi: 10.1242/dev.136465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Palatnik JF, Allen E, Wu X, Schommer C, Schwab R, Carrington JC, et al. Control of leaf morphogenesis by microRNAs. Nature. 2003;425:257–263. doi: 10.1038/nature01958. [DOI] [PubMed] [Google Scholar]
- 47.Juarez MT, Kui JS, Thomas J, Heller BA, Timmermans MC. MicroRNA-mediated repression of rolled leaf1 specifies maize leaf polarity. Nature. 2004;428:84–88. doi: 10.1038/nature02363. [DOI] [PubMed] [Google Scholar]
- 48.Kidner CA, Martienssen RA. Spatially restricted microRNA directs leaf polarity through ARGONAUTE1. Nature. 2004;428:81–84. doi: 10.1038/nature02366. [DOI] [PubMed] [Google Scholar]
- 49.Chen X. A microRNA as a translational repressor of APETALA2 in Arabidopsis flower development. Science. 2004;303:2022–2025. doi: 10.1126/science.1088060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Lee H, Yoo SJ, Lee JH, Kim W, Yoo SK, Fitzgerald H, et al. Genetic framework for flowering-time regulation by ambient temperature-responsive miRNAs in Arabidopsis. Nucleic Acids Res. 2010;38:3081–3093. doi: 10.1093/nar/gkp1240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Laufs P, Peaucelle A, Morin H, Traas J. MicroRNA regulation of the CUC genes is required for boundary size control in Arabidopsis meristems. Development. 2004;131:4311–4322. doi: 10.1242/dev.01320. [DOI] [PubMed] [Google Scholar]
- 52.Yin Z, Li C, Han X, Shen F. Identification of conserved microRNAs and their target genes in tomato (Lycopersicon esculentum) Gene. 2008;414:60–66. doi: 10.1016/j.gene.2008.02.007. [DOI] [PubMed] [Google Scholar]
- 53.Yoon EK, Yang JH, Lim J, Kim SH, Kim SK, Lee WS. Auxin regulation of the microRNA390-dependent transacting small interfering RNA pathway in Arabidopsis lateral root development. Nucleic Acids Res. 2010;38:1382–1391. doi: 10.1093/nar/gkp1128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Wang JW, Wang LJ, Mao YB, Cai WJ, Xue HW, Chen XY. Control of root cap formation by microRNA-targeted auxin response factors in Arabidopsis. Plant Cell. 2005;17:2204–2216. doi: 10.1105/tpc.105.033076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Meng Y, Ma X, Chen D, Wu P, Chen M. MicroRNA-mediated signaling involved in plant root development. Biochem Bioph Res Co. 2010;393:345–349. doi: 10.1016/j.bbrc.2010.01.129. [DOI] [PubMed] [Google Scholar]
- 56.Xie Y, Mao Y, Zhang W, Lai D, Wang Q, Shen W. Reactive oxygen species-dependent nitric oxide production contributes to hydrogen-promoted stomatal closure in Arabidopsis. Plant Physiol. 2014;165:759–773. doi: 10.1104/pp.114.237925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Liu D, Xu S, Hu H, Pan J, Li P, Shen W. Endogenous hydrogen sulfide homeostasis is responsible for the alleviation of senescence of postharvest daylily flower via increasing antioxidant capacity and maintained energy status. J Agr Food Chem. 2017;65:718–726. doi: 10.1021/acs.jafc.6b04389. [DOI] [PubMed] [Google Scholar]
- 58.Jiang ZY, Woollard ACS, Wolff SP. Hydrogen peroxide production during experimental protein glycation. FEBS Lett. 1990;268:69–71. doi: 10.1016/0014-5793(90)80974-N. [DOI] [PubMed] [Google Scholar]
- 59.Bellincampi D, Dipierro N, Salvi G, Cervone F, De Lorenzo G. Extracellular H2O2 induced by oligogalacturonides is not involved in the inhibition of the auxin-regulated rolB gene expression in tobacco leaf explants. Plant Physiol. 2000;122:1379–1385. doi: 10.1104/pp.122.4.1379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods. 2001;25:402–408. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- 61.Schmittgen TD, Livak KJ. Analyzing real-time PCR data by the comparative CT method. Nat Protoc. 2008;3:1101–1108. doi: 10.1038/nprot.2008.73. [DOI] [PubMed] [Google Scholar]
- 62.Xu S, Jiang Y, Cui W, Jin Q, Zhang Y, Bu D, et al. Hydrogen enhances adaptation of rice seedlings to cold stress via the reestablishment of redox homeostasis mediated by miRNA expression. Plant Soil. 2017;414:53–67. doi: 10.1007/s11104-016-3106-8. [DOI] [Google Scholar]
- 63.Qi F, Xiang Z, Kou N, et al. Nitric oxide is involved in methane-induced adventitious root formation in cucumber. Physiol Plant. 2017;159:366–377. doi: 10.1111/ppl.12531. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All relevant data are within this article and its supporting information files.