

Abstract
Posterior projections of the ophthalmic division of the trigeminal nerve (the ophthalmic nerve) are distributed in the tentorium cerebelli as recurrent meningeal branches. We investigated the morphological tentorial distribution of the ophthalmic nerve. Fifty‐two sides of the tentorium cerebelli and adjacent dura mater obtained from 29 human specimens were stained using Sihler's method to examine the nerve fibres in the dural sheets. The innervation patterns of the tentorium cerebelli were classified into the following four types according to their distributions: Type 1, where nerve fibres projected to both the straight and transverse sinuses; Type 2, where nerve fibres projected only to the transverse sinus and lateral convexity; Type 3, where nerve fibres projected medially only to the straight sinus and the posterior part of the falx cerebri; and Type 4, where the nerve fibres terminated within the tentorium cerebelli. Images of the tentorium cerebelli were superimposed to identify areas of dense innervation. The incidence rates of Types 1–4 were 71.2% (n = 37), 21.2% (n = 11), 3.8% (n = 2) and 3.8% (n = 2), respectively. More branches of nerve fibres traversed towards the transverse sinus posterolaterally than towards the straight sinus medially. The space between the anterior half of the straight sinus and the medial tentorial notch can be considered a safe surgical area where innervation is scarce. The posterior part of the falx cerebri was innervated by the ophthalmic nerve that traversed to the straight sinus. The parietal branches of the middle meningeal artery in the lateral convexity that were projected orthogonally by the ophthalmic nerve traversed the transverse sinus, implicating their vulnerability and possible sensitivity under physiological or neurosurgical conditions. This study has revealed the macroscopic tentorial innervation of the dura mater in humans, which could be useful information for both neurosurgeons and neurologists.
Keywords: nervus tentorii, ophthalmic division of the trigeminal nerve, Sihler's stain, straight sinus, tentorium cerebelli, transverse sinus
Introduction
The cranial dura mater consists of pericranial parts surrounding the neurocranium and floating parts including the falx cerebri and the tentorium cerebelli. The tentorium cerebelli is a horizontal partition that divides the cranial cavity into the cerebral fossa and posterior cranial fossa housing the cerebellum, and it is attached medially to the falx cerebri and posteriorly to the occipital bone along to the transverse sinus (Drake et al. 2009).
The cranial dura mater not only provides mechanical support to the brain but also exhibits significant metabolic and nervous activity (Cavallotti et al. 1998; May & Goadsby, 1999; Bartsch & Goadsby, 2002). Previous studies have shown that the trigeminal nerve supplies the cranial dura mater both in humans and in animal models (Lv et al. 2014; Dach et al. 2015). Two separate trigeminal nociceptive systems in the cranial dura mater have been distinguished morphologically based on the orientations of their distributions (O'Connor & van der Kooy, 1986; Andres et al. 1987; Messlinger et al. 1993). The recurrent meningeal branches from the mandibular and maxillary divisions of the trigeminal nerve run parallel to the proximal part of the middle meningeal artery in the dura mater covering the middle cranial fossa (Schueler et al. 2013, 2014), and another branch from the ophthalmic division of the trigeminal nerve projects to the transverse sinus with an orthogonal orientation in rodents (Strassman et al. 2004). However, there have been few morphological investigations of the cranial dura mater in humans that have followed the trajectory of recurrent meningeal branches of the trigeminal nerve over long distances. Posterior projections to the tentorium cerebelli of the recurrent meningeal branches of the ophthalmic nerve are designated as the nervus tentorii (Bauer et al. 2005), but their exact course and possible variations in humans are not yet fully understood due to the nerve fibres being difficult to visualize either on or in the dural sheets.
The present study investigated the anatomical basis and variability of the ophthalmic nerve in the tentorium cerebelli (the nervus tentorii) and adjacent dura mater. The supratentorial dura mater was examined using whole‐mount staining with Sihler's method to identify the global patterns of dural innervation in humans. The aim of this study was to provide more detailed information about the course of this nerve in the majority of the supratentorial dura mater, encompassing the tentorium cerebelli, falx cerebri and lateral convexity.
Methods
The supratentorial part of the cranial dura mater was obtained from 29 formalin‐embalmed human cadavers that had been donated to Konkuk University, Ulsan University and Hanyang University in Korea. The donors comprised 18 males and 11 females aged 72.6 ± 10.8 years (mean ± SD) at death. Before they died, the donors signed documents agreeing to their participation in the body donation program of the medical school and the use of their body for clinical studies.
The anterior part of the falx cerebri was elevated from the crista galli and frontal crest in the anterior cranial fossa, and then the lateral margin of the tentorium cerebelli on the superior border of the petrous body (the superior petrosal sinus) was incised to extract the whole brain (Fig. 1). Although the cadavers had been embalmed using formalin prior to dissection, the harvested cranial dura was also immersed in 10% neutralized formalin for several weeks to remove phenol in the fixation solution and to ensure complete fixation of the specimens. Each fixed specimen was washed under running tap water for about 1 h and then immersed in 3% aqueous potassium hydroxide solution for maceration while being observed continuously. The duration of maceration varied depending on the thickness of membranous structure and blood congestion in the venous sinus in order to prevent lacerating the dura mater and the loss of nerve fibres. After decalcification in Sihler's solution I (comprising 1 vol glacial acetic acid, 1 vol glycerine and 6 vol 1% w/v aqueous chloral hydrate), each specimen was processed according to the routine procedure of Sihler's method and then photographed (Mu & Sanders, 2010). Specimens that had been stained excessively due to the presence of severe fibrosis or widespread calcification of the dura mater were excluded. This protocol reduced the original 62 specimens of 31 cadavers to 52 sides of 29 cadavers. We ultimately obtained both sides of the tentorium cerebelli from 23 cadavers and one side from six cadavers.
Figure 1.
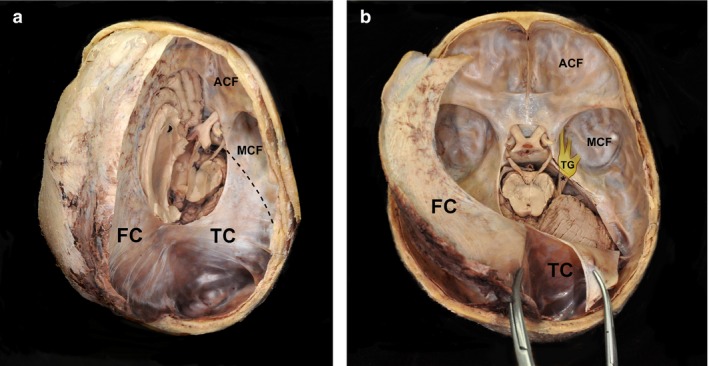
Exposed cranial dura mater from a human cadaver. (a) Removing the dura mater of the right hemisphere reveals the tentorium cerebelli covering the cerebellum and attached posteriorly to the occipital bone. (b) Elevation and reflection of the anterior portion of the falx cerebri from the frontal crest and the crista galli. The lateral margin of the tentorium cerebelli based on the superior petrosal sinus is incised and reflected. ACF, anterior cranial fossa; FC, falx cerebri; MCF, middle cranial fossa; TC, tentorium cerebelli; asterisk, trigeminal nerve from the pons; dashed line, superior border of the petrous body; TG, trigeminal ganglion under the dura mater.
The boundaries of each tentorium cerebelli (the tentorial notch, the superior petrosal sinus, the straight sinus and the transverse sinus) were outlined, and the courses of the nerve fibres were manually traced using a black line in standard image‐processing software (Adobe photoshop CS6, Adobe Systems, San Jose, CA, USA). Images of 52 sides with various outlines of the tentorium cerebelli were superimposed to identify the significant area of nerve distributions using computer software (sqirlz morph 2.0; this software can be downloaded for free from http://sqirlz-morph.en.softonic.com). Mean and SD values of the number of branches reaching the venous sinuses were calculated, and statistical comparisons were performed using standard software (version 18.0, spss for Windows, SPSS, Chicago, IL, USA).
Results
The ophthalmic nerve give rise to a tentorial branch (the nervus tentorii) at the point of the anterior edge of the tentorial notch between the diaphragma sellae and the superior petrosal sinus (Fig. 2). Applying Sihler's method resulted in the nerve fibres being stained dark purple by haematoxylin, and so they contrasted with the connective tissue of the dura mater that had been destained rapidly using the post‐treatment solution. Stained nerve fibres of the nervus tentorii projected to posterior regions in all specimens (Fig. 3). The distribution pattern of the nervus tentorii was asymmetrical on both sides of the tentorium cerebelli, with either many nerve fibres (Fig. 3b,c) or only a few fibres (Fig. 3d,e) being evident. The nervus tentorii generally projected linearly, but in 36.5% of specimens (19/52) it curved acutely in the medial or lateral direction at the middle or posterior portion of the tentorium cerebelli (Fig. 3, arrows). The various types of nervus tentorii innervation were classified based on these morphological observations.
Figure 2.
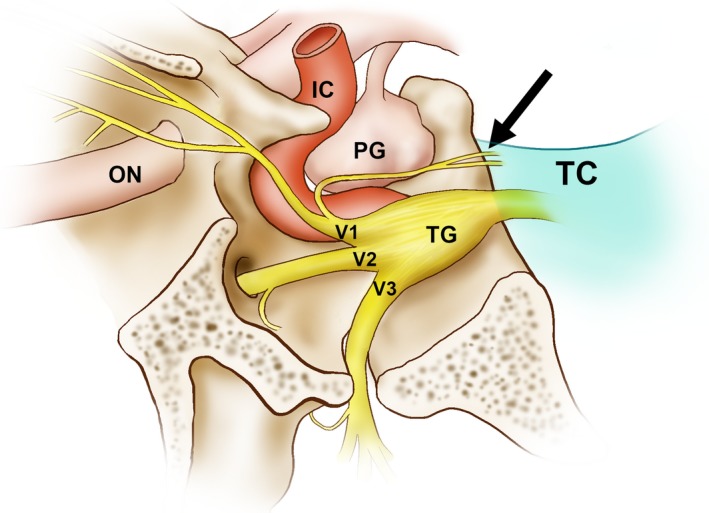
Illustration of a tentorial branch arising from the ophthalmic division (the nervus tentorii; arrow) of the trigeminal ganglion. IC, internal carotid artery; ON, optic nerve; PG, pituitary gland; TC, tentorium cerebelli; TG, trigeminal ganglion; V1, ophthalmic division; V2, maxillary division; V3, mandibular division.
Figure 3.
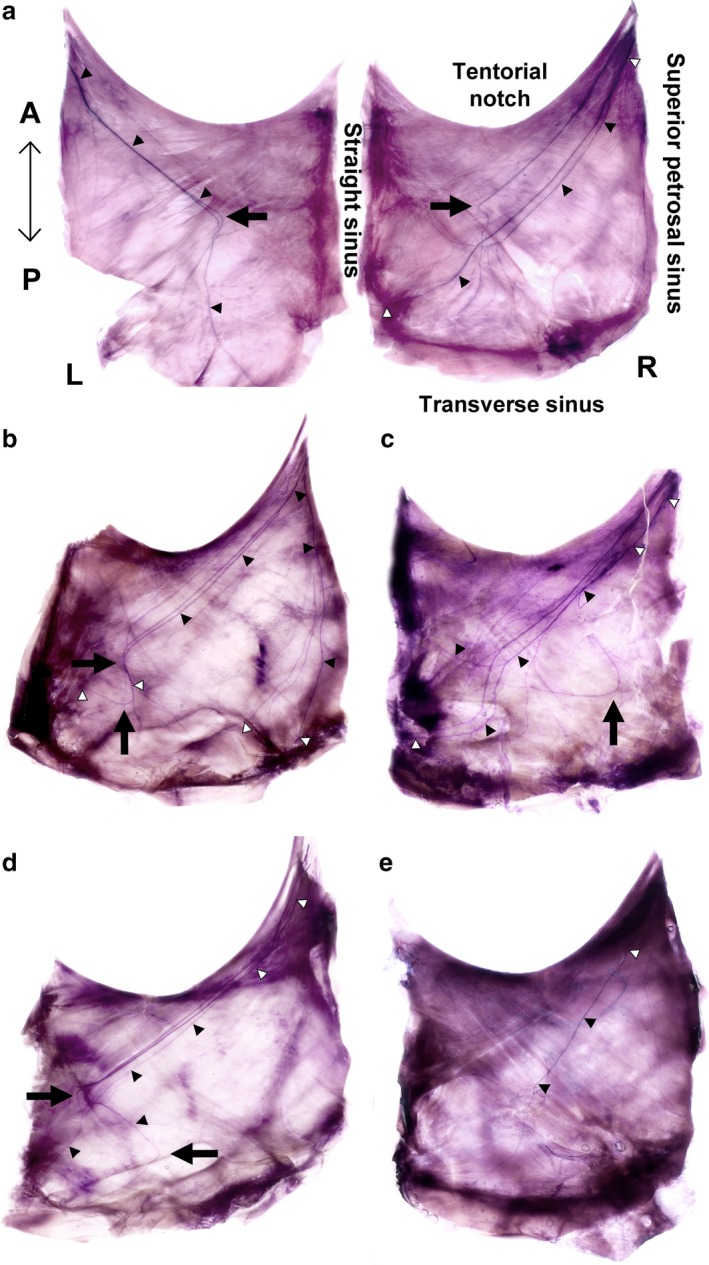
Innervation of the tentorium cerebelli identified using Sihler's stain. (a) Linear and curved projections of the nervus tentorii with an asymmetrical innervation pattern in the same individual. (b) Separate courses of independent groups of nerve fibres to both the straight and transverse sinuses. (c) Nerve fibres project to both the straight and transverse sinuses as a consequence of significant pruning. (d) Nerve fibres projecting to the straight sinus changed their directions to the transverse sinus with arborization to both the straight and transverse sinuses. (e) There were few nerve fibres within the territory of the tentorium cerebelli. A, anterior; L, left; P, posterior; R, right; black arrows, locations of nerve fibres with acutely curved projections; black and white triangles, nerve fibres in general.
Some nerve fibres from the nervus tentorii traversed the straight sinus and terminated at the posterior part of the falx cerebri (Fig. 4a,a'). Occasionally the nervus tentorii traversed the superior wall of the transverse sinus and was distributed to the lateral convexity. These nerve fibres crossed the distal part of the parietal branches of the middle meningeal artery in the dura mater of the lateral convexity in an orthogonal direction (Fig. 4b,c). The innervation of the nervus tentorii on the middle meningeal artery exhibited diverse patterns: none, a single projection or multiple fibres.
Figure 4.
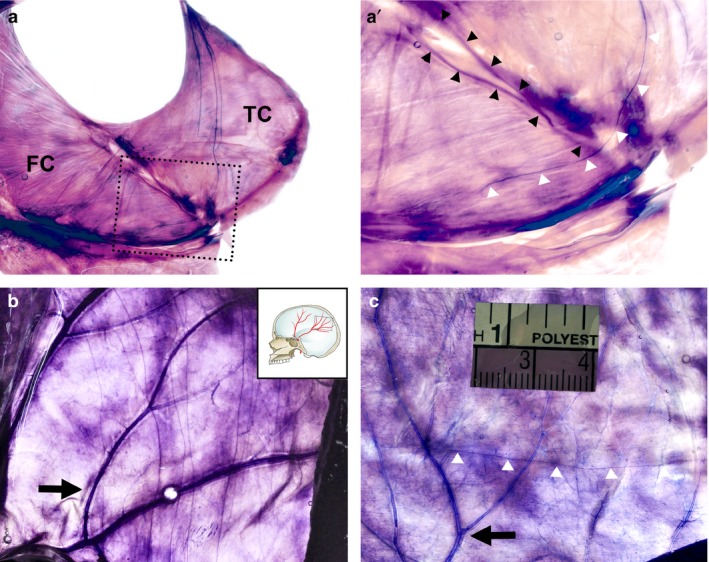
Innervation of the adjacent dura mater revealed using Sihler's stain. (a) Innervation of the posterior part of the falx cerebri elongated from the tentorium cerebelli. (a') Magnification of the dotted box in (a). White triangles indicate a nerve fibre of the nervus tentorii traversing to the straight sinus. (b) The parietal branches of the middle meningeal artery (MMA) in the lateral convexity that projects via multiple nerve fibres elongated from the tentorium cerebelli. (c) A single nerve fibre (white triangles) orthogonal to the parietal branches of the MMA in another specimen. FC, falx cerebri; TC, tentorium cerebelli; black arrows, parietal branches of the middle meningeal artery; black triangles, straight sinus; white triangles, nerve fibres.
The projection patterns of the nervus tentorii on the 52 sides of the tentorium cerebelli was classified into the following four types according to the direction to the venous sinuses: Type 1, where nerve fibres projected to both the transverse and straight sinuses with or without arborization; Type 2, where nerve fibres projected only to the transverse sinus and lateral convexity; Type 3, where nerve fibres projected medially only to the straight sinus and the posterior part of the falx cerebri; and Type 4, where nerve fibres remained within the tentorium cerebelli and did not reach the peripheral venous sinuses (Fig. 5). The incidence rates of Types 1–4 were 71.2% (n = 37), 21.2% (n = 11), 3.8% (n = 2) and 3.8% (n = 2), respectively. In Type 1, the number of nerve fibres projecting to the transverse sinus was remarkably higher than the number projecting to the straight sinus (Table 1). There was no sex difference in the projection pattern in the nerve fibre.
Figure 5.
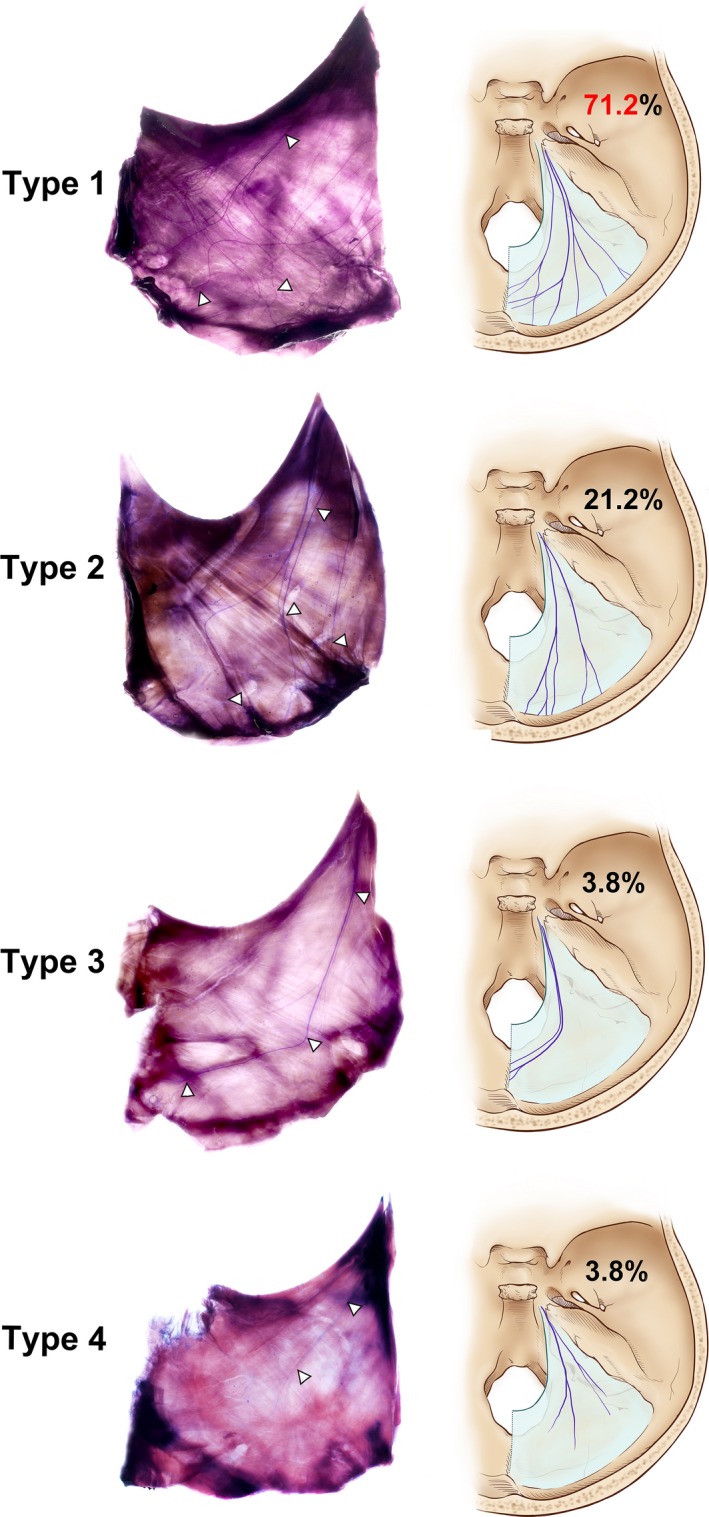
Distribution types of the tentorium cerebelli innervation. White triangles (left section) and purple lines (right section) indicate a nerve fibre of the nervus tentorii. Type 1: The nervus tentorii projecting to both the straight and transverse sinuses. Type 2: The nervus tentorii projecting to the transverse sinus. Type 3: The nervus tentorii projecting to the straight sinus. Type 4: Termination of the nervus tentorii within the tentorium cerebelli.
Table 1.
Numbers of nerve fibres projecting to the straight and transverse sinuses for each type of innervation pattern of the tentorium cerebelli
| Type 1 (n = 37) | Type 2 (n = 11) | Type 3 (n = 2) | Type 4 (n = 2) | |
|---|---|---|---|---|
| S | 2.6 ± 1.3 | 4.0 ± 2.0 | ||
| T | 4.2 ± 2.3a | 3.2 ± 0.6 |
Data are mean ± SD values.
S, number of nerve fibres projecting to the straight sinus; T, number of nerve fibres projecting to the transverse sinus.
P < 0.05.
Images of the 52 sides of the tentorium cerebelli and their innervation were merged with the aid of various outlines of the tentorium cerebelli. Superimposing images of the nervus tentorii revealed dense innervation in the anterior and lateral parts of the tentorial notch, the entire region of the transverse sinus and the posterior half of the straight sinus (Fig. 6a). In contrast, the innervation was scarce in the space between the anterior half of the straight sinus and the medial tentorial notch exhibit. The nervus tentorii that traversed the transverse sinus crossed the parietal branches of the middle meningeal artery orthogonally and projected to the lateral convexity (Fig. 6b). The medially curved nervus tentorii innervated the posterior part of the falx cerebri, passing through the straight sinus (Fig. 6c). Nerve fibres stained using Sihler's method were not observed in anterior and middle parts of the falx cerebri or the superior sagittal sinus.
Figure 6.
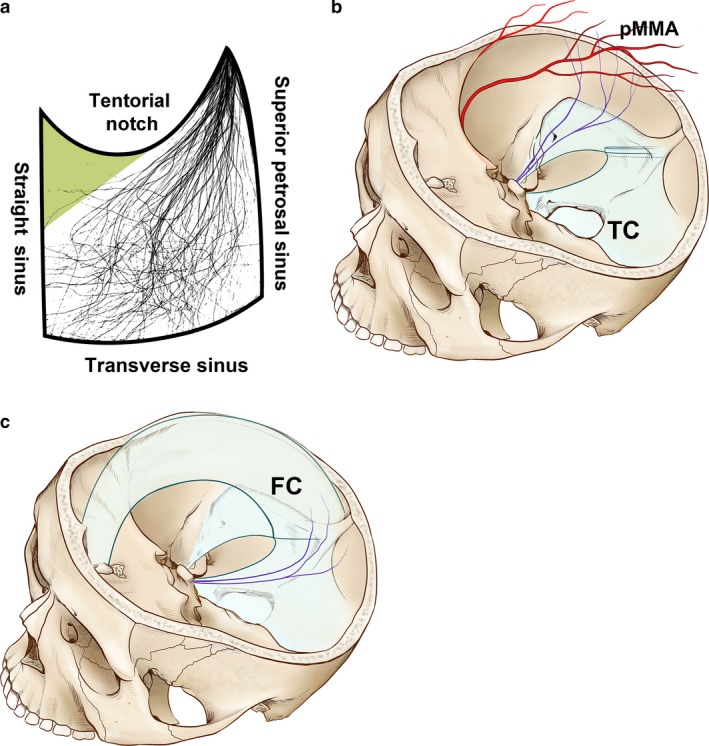
(a) Images of nerve fibres in the tentorium cerebelli superimposed to reveal the pattern of innervation density. The entire area of the transverse sinus and the posterior half of the straight sinus show significant innervation, whereas innervation is scarce in the tentorial notch near the anterior part of the falx cerebri (green triangle). (b,c) Schematics of the nerve distribution in the tentorium cerebelli and adjacent dura mater. Nerve fibres (purple lines) traversing the transverse sinus are distributed to the parietal branches of the MMA in the lateral convexity in an orthogonal direction. The posterior part of the falx cerebri receives nerve fibres elongated from the tentorium cerebelli via the straight sinus. FC, falx cerebri; pMMA, parietal branches of the middle meningeal artery; TC, tentorium cerebelli.
Discussion
Previous descriptions of the supratentorial innervation have been crude and inconsistent, including the medical illustrations that appear in both textbooks and journal articles (Drake et al. 2009; Gilroy et al. 2009; Moore et al. 2009; Kemp et al. 2012; Bauer et al. 2005). Accurate knowledge of the anatomy of the venous sinuses in the dura mater is important during neurosurgery involving the arteriovenous fistula or when resecting meningiomas and other types of brain tumours (Tubbs et al. 2007; Tatarli et al. 2013; Xu et al. 2013; Tanaka, 2015; Amato et al. 2016; Shibao et al. 2016), and so the present study investigated the straight and transverse sinuses adjacent to the tentorium cerebelli in order to elucidate the course of the nervus tentorii.
We found that innervation was scarce in the space between the medial tentorial notch and the anterior half of the straight sinus, whereas the entire region of the transverse sinus and the posterior half of the straight sinus were densely innervated by the nervus tentorii. The nervus tentorii distributed to a broad area of the human dura mater in a variable manner, including the entire region of the tentorium cerebelli, the posterior part of the falx cerebri and the distal middle meningeal artery on the lateral convexity. These morphological findings are consistent with neurosurgical recommendations that traction or contacting the posterior part of the falx cerebri and the tentorium cerebelli should be minimized to avoid eliciting the haemodynamic fluctuations (Kemp et al. 2012; Bauer et al. 2005). Manipulation or cauterization of the cranial dura mater during neurosurgery can result in patients undergoing sudden onset of hypotension, bradycardia, arrhythmia, asystole or apnoea caused by the sensory nerve endings of the trigeminal nerve sending neural signals via the trigeminal ganglion to the sensory nucleus of the trigeminal nerve in the brainstem (May & Goadsby, 1999; Gharabaghi et al. 2006; Acioly et al. 2011; Meuwly et al. 2015). Minimizing traction over the trigeminal nerve or the related zone of the dura mater is strongly emphasized in the literature on neurosurgery to prevent these trigeminocardiac reflexes (Schaller et al. 2009; Etezadi et al. 2013; Bauer et al. 2005; Chowdhury et al. 2015).
Approximately one‐third of the specimens in the present study showed acutely curved projections of the nervus tentorii in the middle or posterior portion of the tentorium cerebelli, in contrast to the general branching or distribution pattern of nerve fibres (Fig. 3, arrows). Diverse patterns of supratentorial innervation might result from variations in the dynamics of the cranial dura mater and trigeminal ganglion during the early stage of embryo development (O'Rahilly & Müller, 2007; Davidson et al. 2012; Amato et al. 2016). The Carnegie Collections indicate that at stage 13 of embryo development (when the embryo has a size of 4–6 mm), the ophthalmic division is the clearest of the tripartite divisions of the trigeminal ganglion, and both mesenchymal and neural crest‐derived cells appear to be involved in the formation of the primary meninges (O'Rahilly & Müller, 2007; Tanaka, 2015). As the brain hemispheres increase in size, the dural mesenchymal tissue of floating parts such as the tentorium cerebelli and falx cerebri become fused and compressed at an earlier stage compared with the dural membrane of the bony structure (Tanaka, 2015; Amato et al. 2016). Furthermore, the density of dural innervation decreases due to axon traction, apoptosis of the trigeminal ganglion cells, and changes in neuropeptide expression in trigeminal ganglion cells, mostly during post‐natal days 5–55 (Davidson et al. 2012). Individual differences in the mode of fusion of the falx cerebri, physiological compression of the tentorium cerebelli during the early stage and extensive remodelling of trigeminal axons after birth might result in disarrangement of the nervus tentorii projections along with significant variations between individuals. Further studies therefore should investigate the acute angulation of the nerve fibre in the tentorium cerebelli, along with the underlying developmental mechanisms.
The nervus tentorii traversed the superior wall of the transverse sinus, occasionally innervating the dura mater covering the lateral aspect of the occipital lobe. These fibres elongated from the nervus tentorii contacting the distal part of the parietal branch of the middle meningeal artery orthogonally, with variability among individuals (Fig. 4b). Activation of the meningeal sensory fibres is thought to be involved in some types of headaches, including migraine (Bartsch & Goadsby, 2003; Strassman et al. 2004; Calhoun et al. 2010; Xu et al. 2013). Vasodilation of the middle meningeal artery and neurogenic inflammation of the recurrent meningeal branches from the maxillary and mandibular divisions of the trigeminal nerve in the skull base have been suggested as causes of vascular headache (Messlinger et al. 1993; Schaller et al. 2009). The interesting finding of the present study that the middle meningeal artery in the dura mater covering the occipitoparietal lobe is innervated by multiple sensory nerve fibres far from the middle cranial fossa, would implicate another course of sensitization also under physiological conditions – that is, not only neurosurgical conditions.
Sihler's method did not reveal any nerve fibres in the anterior and middle parts of the falx cerebri and the superior sagittal sinus, despite them having been detected by gene‐related peptide‐like and substance‐P‐like immunoreactivities in animal models (Andres et al. 1987; Messlinger et al. 1993; Strassman et al. 2004). However, Sihler's method as used in the present study has some limitations that can result in underestimations of complete innervation patterns. Some investigators have reported that the haematoxylin used in this method is effective at staining myelinated nerve fibres but not unmyelinated nerve fibres (Mu & Sanders, 2010). The myelin sheath surrounding nerve fibres gradually disappears near the nerve terminals, which could have resulted in tiny nerve endings of the nervus tentorii being poorly stained in the present study.
In the Type 4 innervation pattern (Fig. 5d), the transverse sinus was not distinctly innervated by the nervus tentorii. In this pattern it is suspected that nerve fibres in the posterior cranial fossa constitute the dominant innervation in the dura mater of the occipital region. In addition to the middle cranial fossa and supratentorial parts, the dura mater covering the posterior cranial fossa has been known to be innervated by the recurrent meningeal branches of the vagus, facial and glossopharyngeal nerves, and the upper three cervical nerves (Bauer et al. 2005; Kemp et al. 2012). Our preliminary study found that only one specimen of the posterior cranial fossa showed a linear projection of unknown nerve fibres from the foramen magnum (data not shown). We are currently investigating the innervation of the posterior cranial fossa with reference to the hypoglossal canal, jugular foramen, foramen magnum and the posterior meningeal artery in order to provide a more comprehensive understanding of the innervation of the cranial dura mater.
Conclusion
This study has revealed diverse courses of the nervus tentorii in the human supratentorial dura mater. The nervus tentorii mainly projected to both the straight and transverse sinuses, with more nerve fibres traversing towards the transverse sinus. The dense innervation of the entire region of the transverse sinus and the posterior half of the straight sinus suggests that the middle meningeal artery in the lateral convexity and the posterior part of the falx cerebri could also be areas that receive significant innervation from the ophthalmic nerve; that is, in addition to the tentorium cerebelli itself. These data could represent useful information for both neurosurgeons and neurologists.
Authors' contributions
Conception and design: K.O.H. and S.O.N.G. Acquisition of data: L.E.E. and S.H.I.N. Analysis and interpretation of data: L.E.E. Drafting the article: L.E.E. Critically revising the article: K.O.H. and S.O.N.G. Reviewed submitted version of manuscript: L.E.E. and S.O.N.G. Administrative/technical/material support: S.H.I.N. and S.O.N.G. Study supervision: K.O.H. and S.O.N.G. Illustrator: L.E.E.
Ethics statement
The cadavers came from people who had given their informed consent to use their bodies.
Acknowledgements
This research was supported by the Basic Science Research Program funded by the Konkuk University (2016‐A019‐0338). The authors report no conflict of interest concerning the materials or methods used in this study or the findings specified in this paper.
References
- Acioly MA, Carvalho CH, Koerbel A, et al. (2011) The role of the trigeminocardiac reflex in postoperative hearing function in non‐vestibular schwannoma cerebellopontine angle tumors. J Clin Neurosci 18(2), 237–240. [DOI] [PubMed] [Google Scholar]
- Amato MC, Tirapelli LF, Carlotti CG, et al. (2016) Straight sinus: ultrastructural analysis aimed at surgical tumor resection. J Neurosurg 125(2), 494–507. [DOI] [PubMed] [Google Scholar]
- Andres KH, Von DM, Muszynski K, et al. (1987) Nerve fibres and their terminals of the dura mater encephali of the rat. Anat Embryol (Berl) 175(3), 289–301. [DOI] [PubMed] [Google Scholar]
- Bartsch T, Goadsby PJ (2002) Stimulation of the greater occipital nerve induces increased central excitability of dural afferent input. Brain 125(7), 1496–1509. [DOI] [PubMed] [Google Scholar]
- Bartsch T, Goadsby PJ (2003) Increased responses in trigeminocervical nociceptive neurons to cervical input after stimulation of the dura mater. Brain 126(8), 1801–1813. [DOI] [PubMed] [Google Scholar]
- Bauer DF, Youkilis A, Schenck C, et al. (2005) The falcine trigeminocardiac reflex: case report and review of the literature. Surg Neurol 63(2), 143–148. [DOI] [PubMed] [Google Scholar]
- Calhoun AH, Ford S, Millen C, et al. (2010) The prevalence of neck pain in migraine. Headache 50(8), 1273–1277. [DOI] [PubMed] [Google Scholar]
- Cavallotti D, Artico M, De Santis S, et al. (1998) Catecholaminergic innervation of the human dura mater involved in headache. Headache 38(5), 352–355. [DOI] [PubMed] [Google Scholar]
- Chowdhury T, Mendelowith D, Golanov E, et al. (2015) Trigemino‐Cardiac Reflex Examination Group. Trigeminocardiac reflex: the current clinical and physiological knowledge. J Neurosurg Anesthesiol 27(2), 136–147. [DOI] [PubMed] [Google Scholar]
- Dach F, Éckeli ÁL, Ferreira Kdos S, et al. (2015) Nerve block for the treatment of headaches and cranial neuralgias – a practical approach. Headache 55, 59–71. [DOI] [PubMed] [Google Scholar]
- Davidson JR, Mack J, Gutnikova A, et al. (2012) Developmental changes in human dural innervation. Childs Nerv Syst 28(5), 665–671. [DOI] [PubMed] [Google Scholar]
- Drake RL, Vogal A, Mitchell A (2009) Gray's Anatomy for Students, 2nd edn Churchill Livingstone: Elsevier, pp. 831. [Google Scholar]
- Etezadi F, Orandi AA, Orandi AH, et al. (2013) Trigeminocardiac reflex in neurosurgical practice: an observational prospective study. Surg Neurol Int 4, 116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gharabaghi A, Koerbel A, Samii A, et al. (2006) The impact of hypotension due to the trigeminocardiac reflex on auditory function in vestibular schwannoma surgery. J Neurosurg 104(3), 369–375. [DOI] [PubMed] [Google Scholar]
- Gilroy AM, MacPherson BR, Ross LM, et al. (2009) Atlas of Anatomy, 2nd edn Thieme Medical Publishers: Thieme, pp. 603. [Google Scholar]
- Kemp WJ 3rd, Tubbs RS, Cohen‐Gadol AA (2012) The innervation of the cranial dura mater: neurosurgical case correlates and a review of the literature. World Neuroserg 78(5), 505–510. [DOI] [PubMed] [Google Scholar]
- Lv X, Wu Z, Li Y (2014) Innervation of the cerebral dura mater. Neuroradiol J 27, 293–298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- May A, Goadsby PJ (1999) The trigeminovascular system in humans: pathophysiologic implications for primary headache syndromes of the neural influences on the cerebral circulation. J Cereb Blood Flow Metab 19(2), 115–127. [DOI] [PubMed] [Google Scholar]
- Messlinger K, Hanesch U, Baumgärtel M, et al. (1993) Innervation of the dura mater encephali of cat and rat: ultrastructure and calcitonin gene‐related peptide‐like and substance P‐like immunoreactivity. Anat Embryol 188(3), 219–237. [DOI] [PubMed] [Google Scholar]
- Meuwly C, Chowdhury T, Sandu N, et al. (2015) Anesthetic influence on occurrence and treatment of the trigemino‐cardiac reflex: a systematic literature review. Medicine (Baltimore) 94(18), e807. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moore KL, Agur AM, Dalley AF (2009) Clinically Oriented Anatomy, 6th edn Wolters Kluwer: Lippincott Williams & Wilkins, pp. 873. [Google Scholar]
- Mu L, Sanders I (2010) Sihler's whole mount nerve staining technique: a review. Biotech Histochem 85(1), 19–42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O'Connor TP, van der Kooy D (1986) Pattern of intracranial and extracranial projections of trigeminal ganglion cells. J Neurosci 6(8), 2200–2207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O'Rahilly R, Müller F (2007) The development of the neural crest in the human. J Anat 211(3), 335–351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schaller B, Cornelius JF, Prabhakar H, et al. (2009) The trigemino‐cardiac reflex: an update of the current knowledge. J Neurosurg Anesthesiol 3, 187–195. [DOI] [PubMed] [Google Scholar]
- Schueler M, Messlinger K, Dux M, et al. (2013) Extracranial projections of meningeal afferents and their impact on meningeal nociception and headache. Pain 154(9), 1622–1631. [DOI] [PubMed] [Google Scholar]
- Schueler M, Neuhuber WL, De Col R, et al. (2014) Innervation of rat and human dura mater and pericranial tissues in the parieto‐temporal region by meningeal afferents. Headache 54, 996–1009. [DOI] [PubMed] [Google Scholar]
- Shibao S, Toda M, Orii M, et al. (2016) Various patterns of the middle cerebral vein and preservation of venous drainage during the anterior transpetrosal approach. J Neurosurg 124(2), 432–439. [DOI] [PubMed] [Google Scholar]
- Strassman AM, Weissner W, Williams M, et al. (2004) Axon diameters and intradural trajectories of the dural innervation in the rat. J Comp Neurol 473, 364–376. [DOI] [PubMed] [Google Scholar]
- Tanaka M (2015) Embryological consideration of dural arteriovenous fistulas. Neurol Med Chir (Tokyo) 56(9), 544–551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tatarli N, Ceylan D, Canaz H, et al. (2013) Falcine venous plexus within the falx cerebri: anatomical and scanning electron microscopic findings and clinical significance. Acta Neurochir (Wien) 155(11), 2183–2189. [DOI] [PubMed] [Google Scholar]
- Tubbs RS, Loukas M, Louis RG Jr, et al. (2007) Anatomy of the falcine venous plexus. J Neurosurg 107(1), 155–157. [DOI] [PubMed] [Google Scholar]
- Xu Z, Su C, Xiao Y (2013) A massive calcification and ossification of the transverse sinus and the neighbouring dura mimicking meningioma. BMC Neurol 13, 143. [DOI] [PMC free article] [PubMed] [Google Scholar]