

Abstract
Background:
Chronic exposure to noise induces changes on the central nervous system of exposed animals. Those changes affect not only the auditory system but also other structures indirectly related to audition. The hippocampus of young animals represents a potential target for these effects because of its essential role in individuals’ adaptation to environmental challenges.
Objective:
The aim of the present study was to evaluate hippocampus vulnerability, assessing astrocytic morphology in an experimental model of environmental noise (EN) applied to rats in pre-pubescent stage.
Materials and Methods:
Weaned Wistar male rats were subjected to EN adapted to the rats’ audiogram for 15 days, 24 h daily. Once completed, plasmatic corticosterone (CORT) concentration was quantified, and immunohistochemistry for glial fibrillary acidic protein was taken in hippocampal DG, CA3, and CA1 subareas. Immunopositive cells and astrocyte arborizations were counted and compared between groups.
Results:
The rats subjected to noise exhibited enlarged length of astrocytes arborizations in all hippocampal subareas. Those changes were accompanied by a marked rise in serum CORT levels.
Conclusions:
These findings confirm hippocampal vulnerability to EN and suggest that glial cells may play an important role in the adaptation of developing the participants to noise exposure.
Keywords: Dentate gyrus, environmental noise, GFAP, glia, stress
Introduction
Noise is an unwanted product of urbanization that is becoming increasingly dangerous for humans and animals.[1,2,3,4] It may affect patient’s health both, at auditory and non-auditory levels. Noise has been largely proved to be a common cause of hearing loss on one hand, and one of the most common causes of stress, anxiety, and sleep disturbances on the other hand.[5,6,7] Duality in auditory and non-auditory effects of noise may obey to the fact that displeasing sounds when perceived, follows two paths in central nervous system: the main path sending information to auditory structures where sounds are processed, and the other path carrying sounds to deep brain structures where they are emotionally interpreted.[8,9] Activation of these non-auditory structures should be responsible for most of the extra-auditory effects of noise.[10,11]
Between extra-auditory structures involved in noise processing, the hippocampus calls attention by being one of the most important regulators of the body’s response to aversive/stressing conditions,[12] and by being part of the extended neural network for affective sound processing.[9] As a part of these systems, the hippocampus becomes vulnerable to environmental stressors, and if conditions exceed the individuals’ adaptive capabilities, it suffers reduction on dendrite branches, alterations in neurotransmitter levels, impairment on cognitive associated processes, and damage to neurogenic niches.[13,14]
If so, most of the stress-related alterations on hippocampus have been reported on neurons, glial cells have emerged as novel targets for stressing conditions (for review, refer Jauregui-Huerta et al. [15]. These cells represent the most common and versatile cells residing in the hippocampus. Astrocytes for example, are the main physiologic support for neurons, one of the most important modulators of synaptic activity, and one of the most active regulators of brain homeostasis.[16,17,18] Every astrocyte possesses many fiber-like processes that envelope several hundred dendrites from many surrounding neurons and serves as an active sensor for any physiological or pathological change at any brain microenvironment.[19] Then, under compromised conditions, astrocytes change their morphology and the pattern of expression of many genes including the glial fibrillary acidic protein (GFAP), the main marker of astrocytes.[20] So in the view of the fact that astrocytes also express receptors for most of the stress related hormones,[15,18,21,22] they should be explored in the contexts of physiological compromise as those generated by environmental stressors.
Because urban noise may become a potent stressor[11] and the hippocampus is an important target for stress molecules, it follows that noise exposure may affect hippocampal integrity not only in the classic neuron-associated way (i.e., diminishing its dendrite branches, reducing its proliferative rates, changing its synaptic capabilities, and so on[23,24,25], but also in a non-classic astrocyte-associated way (modifying astrocyte numbers and/or morphology). Previous studies conducted for us and other laboratories confirmed that noise exposition in early stages produces long-term cognitive impairment and damage to hippocampal neurons.[26,27,28,29] However, it is still unknown as which are the effects on cell populations different from neurons. The present study was then designed to determine the effects of noise on the young hippocampus by assessing the local astrocyte response.
Materials and Methods
Experimental animals
We used 20 Swiss Wistar male rats (51.8–82 g body weight) from 21 to 36 postnatal days (PND). They preceded from an in-house breeding facility at the Centro de Investigación Biomédica de Occidente, Guadalajara, México. These rats were randomly divided into two groups of 10 animals each: Control rats (without any manipulation) were kept in vivarium in polycarbonate cages with an average background noise level of 60 dB, whereas animals in the experimental group were exposed to noise. Both groups were maintained in a 12:12 light-darkness cycle (lights on at 07:00), at 22 ± 2°C temperature, 70% humidity with ad libitum access to tap water and Ralston-rations balanced food. General procedure is shown in Figure 1.
Figure 1.
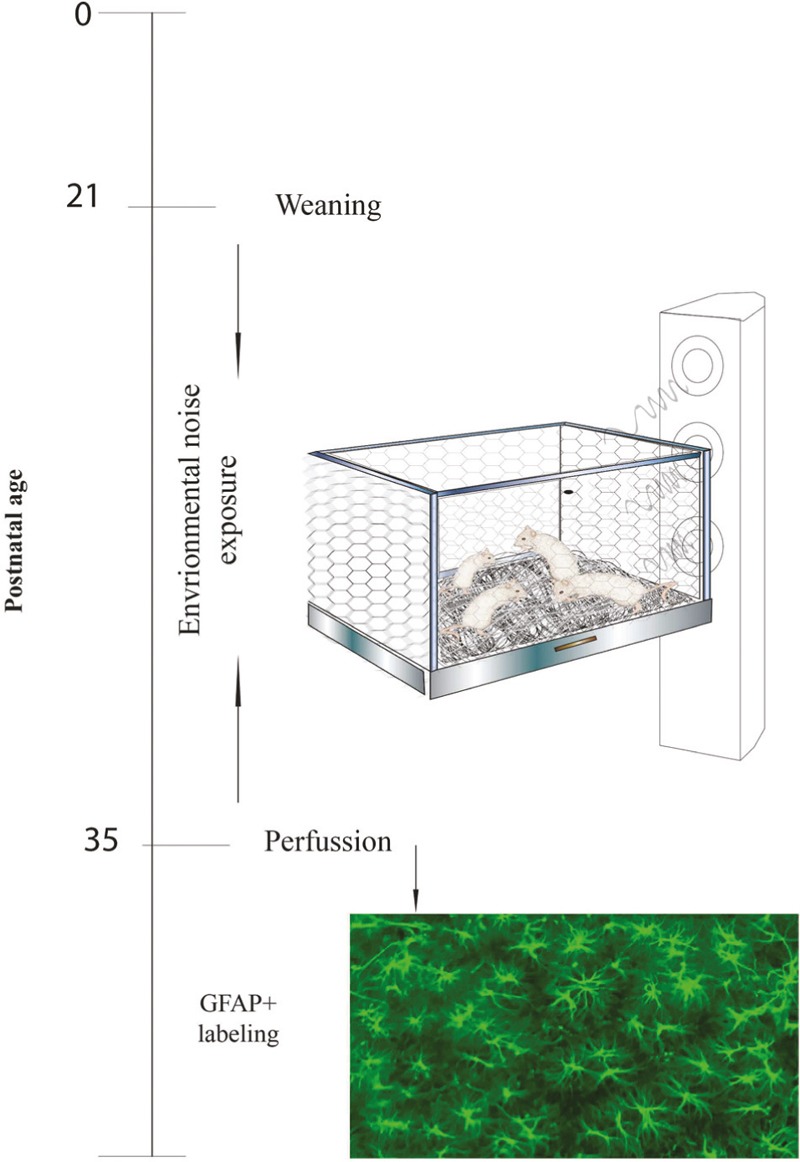
General procedure: Illustrates the general procedure followed in our experiment. Experimental procedures are chronologically depicted at the left part of the line. Procedures are illustrated on the right part of the figure
Noise exposure
From PND 21 to PND 36, the noise-exposed rats were housed in metal grid cages and translated to a soundproof room, provided with professional tweeters (Motorola) suspended 1 m above the cages. The tweeters were connected to amplifier equipment (Mackie M1400; freq. 20 Hz to 70 kHz; 300 W to 8 Ω) with mixer software that transmitted the acoustic signal at levels ranging from 70 dB to 85–103 dB. Noise intensity was measured with a sound-level meter (Radio Shack). To make noise relevant for the auditive capacity of the animals, environmental noise (EN) was adapted to fit the rats’ audiogram (made with software that translates all human noise frequencies to those of the rat). Audio files, provided by Dr. A Rabat,[30] contained unpredictable noise events with a duration from 18 to 39 s and spaced by silent intervals ranging from 20 to 165 s; these sounds were randomly played to the rats during 24 h throughout 15 days; that is, until PND 36.
Corticosterone assay
After noise exposure (PND 36), five rats of each group were decapitated and the trunk blood was collected into chilled heparinized tubes. Plasma was separated by centrifugation and stored at −20°C. Corticosterone (CORT) concentration was quantified using an EIA kit (Oxford Biomedical Research). Absorbance readings were taken at 450 nm using a microplate reader.
Glial fibrillary acidic protein immunohistochemistry
After noise exposure (PND 36), five rats of each group received an intraperitoneal injection of a sub lethal sodium pentobarbital dose (60 mg/kg) and were then perfused through the left cardiac ventricle with 150 mL of saline solution, followed by 200 mL of 4% paraformaldehyde in 0.1 M phosphate buffer saline (PBS), pH 7.4. After perfusion, brains were removed and post-fixed in the same fixative solution for 24 h at 4°C. Coronal slices (35 μm) containing CA1, CA3 and dentate gyrus (DG) of hippocampus were obtained using a vibratome (Leica VT1000E; Leica Microsystems, Wetzlar, Germany). The free floating sections were rinsed with 0.1 M PBS three times for 10 min and blocked for 40 min in 10% normal goat serum diluted in PBS, then incubated overnight with polyclonal rabbit anti-GFAP protein (DakoCytomation) diluted at 1:500 in PBS 0.1 M containing 10% normal goat serum. The sections were washed three times, for 10 min each time in PBS at room temperature, and then incubated for two h with a 1:1000 dilution of anti-rabbit IgG Alexa Fluor 594 (red) in 10% normal goat serum diluted in PBS. After incubation of the secondary antibody, the sections were rinsed in PBS, mounted, dried, and covered with an anti-photobleaching (vectashield).
Glial fibrillary acidic protein count
Series of systematically selected brain sections representing the entire hippocampus (35 μm-thick every 140-μm starting on bregma −4.5 and ending on bregma −2.1) were manually counted using a X400 magnification. From each section (16 sections per animal, four animals per group), we counted the number of GFAP immunopositive cells in 10 microscopic fields (640 μm2) as follows: three fields belonging to the CA1 subarea, two fields belonging to the CA3 subarea, and five fields belonging to the DG subarea.
Astrocytes processes length measurement
To measure astrocytes processes length, we recorded the center of each analyzed microscopic field on photographs measuring 130 μm × 180 μm. The length of the astrocyte prolongations on each picture was measured with the automated Image-pro free software (Fiji version, NIH, USA) over the digital images obtained with a Leica DFC320 camera.[31] We calculate the signal intensity converting the frames into grayscale with homogeneously adjusted threshold. The program reported histograms indicating the number of pixels occupied by the GFAP immuno-labeled processes within the recorded area. Then, we reported the average number of pixels occupied by the GFAP marked processes as an objective measure of astrocytes processes length.
Statistical analysis
Statistical analysis of data was performed by using the IBM Statistical Package for the Social Sciences version 20.0 software (IBM Inc. SPSS, Chicago IL, United States), and graphs were created with adobe illustrator CS6. All data are expressed as mean ± standard error of the mean. For CORT concentrations, we used the unpaired Student t test to compare the two groups. Independent unpaired t tests were also applied to analyze the differences in the number and length of astrocytes processes on each counted region (CA1, CA3, and DG). In all cases, a value of P ≤ 0.05 was chosen to establish significant differences.
Results
Corticosterone concentration
To assess the intensity of stress in experimental rats exposed to noise, the serum CORT concentration was measured at completion of the 15 days period. Figure 2 illustrates CORT measures. As expected, stressed rats (n = 5) exhibited higher serum CORT concentrations than control rats (n = 5) (0.12 ± 0.10, * P < 0.05).
Figure 2.
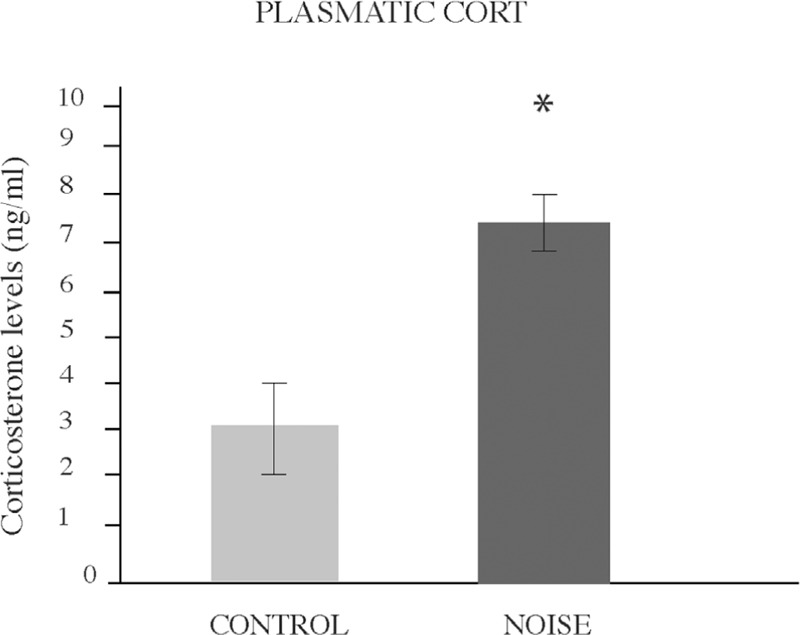
Stress-like effects of noise exposure. Mean ± SEM plasma levels of corticosterone obtained after exposure to environmental noise. Increased CORT levels were found immediately after exposure to noise (*P < 0.05). Bars represent the mean ± SEM
Glial fibrillary acidic protein immunohistochemistry
Immunocytochemical analysis of astrocytes in the hippocampus of control and experimental rats was performed to determine whether the noise exposure was capable of eliciting reactive gliosis in stressed rats. To do so, coronal brain slices were examined through bregma −2.1 to −4.5 (hippocampus territory).[32] In the tissues of stressed rats a trend towards an increase of population of GFAP positives astrocytes was distinguished in CA3, followed by DG and the CA1 areas. However, the differences were not statistically significant for this parameter. Figure 3 illustrates astrocyte numbers.
Figure 3.
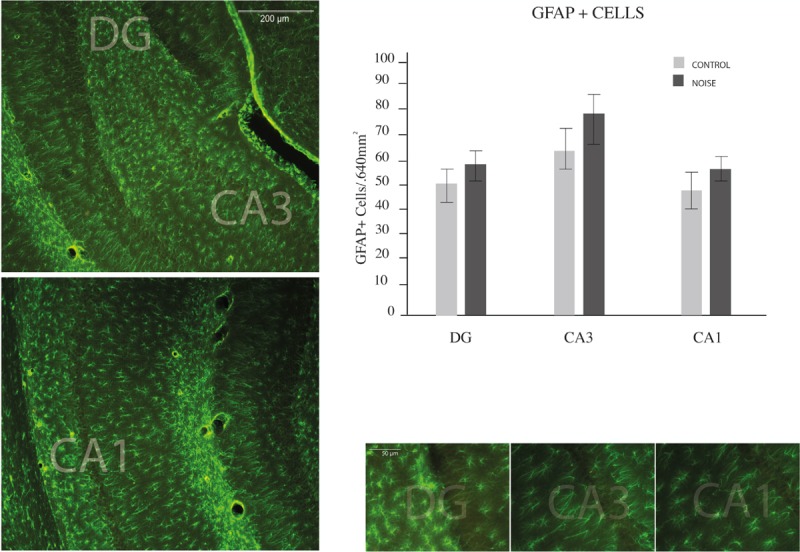
Hippocampal astrocyte numbers. Counting of GFAP (green star-like cells showed in micrographs) performed in the hippocampus of young rats. Regions of interest are displayed on a series of slices depicted in the left part of the figure. Examples of every counted region are showed on right-bottom figures. A trend to greater but non-significant population of GFAP-positive astrocytes was distinguished in CA3, followed by DG and the CA1 areas
Length measurement of astrocytic extensions revealed hypertrophic astrocytes in the hippocampus of stressed rats as compared to controls. Figure 4 shows the comparison of astrocytic arborizations in the different hippocampal regions. All analyzed areas showed statistical differences between groups: Dentate Gyrus (control 109.47 ± 6.60 vs. noise 127.28 ± 4.76, *P < 0.02); CA3 (control 80.22 ± 3.2 vs. noise 108.97 ± 6.9, * P < 0.01); and CA1 (control 90.24 ± 3.6 vs. 117.13 ± 7.4, *P < 0.02). Figure 4 also shows representative immunofluorescence images of each region.
Figure 4.
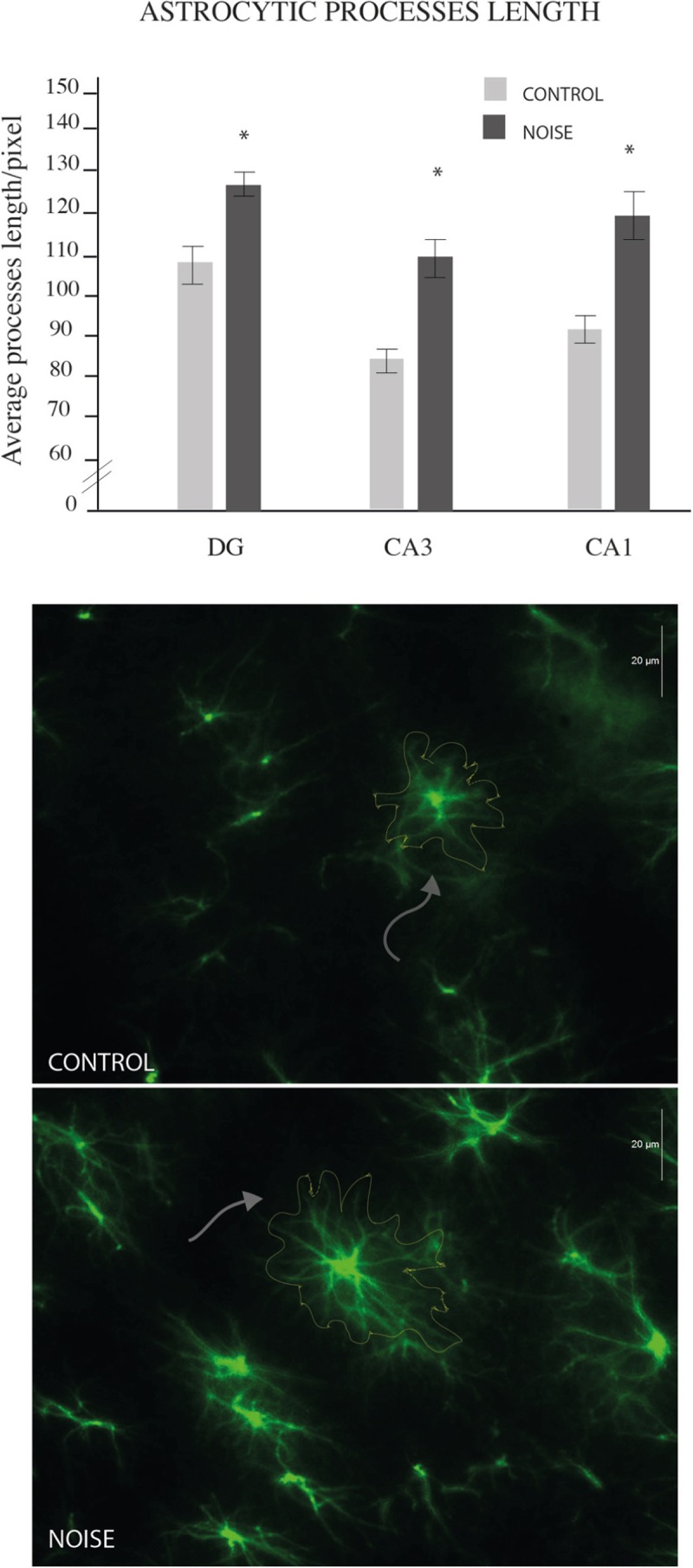
Hippocampal astrocyte lengths. Counting of astrocyte processes length performed in the hippocampus of young rats. White arrows indicate control (upper microphotograph) and noise (lower microphotograph) representative lengths. All analyzed areas showed statistical differences between groups: GD (*P < 0.02); CA3 (*P < 0.01); and CA1 (*P < 0.02)
Discussion
In the present study, we were able to investigate the cerebral effects of noise with minimal intervening variables, and how the astrocytic population in the hippocampus of young male rats responded against acoustic stress. Findings showed that exposure of these rats to EN caused raising of serum CORT and that regardless the anatomical area, astrocytes increased their processes length and tended to increase their total numbers.
Since this kind of noise is unpredictable, inescapable and physically perceptible, we concluded that it caused stress in exposed animals. Previous studies are consistent with this finding and allow us to presume that changes on glial citoarchitecture may − at least in part − be related to stress/hormone deregulation.[29,33] Studies showing that glucocorticoid administration increase astrocyte numbers in the hippocampus support this suggestion.[34] Moreover, our findings agree with other reports of astrogliosis in the hippocampus as a consequence of stress exposure.[35,36] To our knowledge, this is the first study providing evidence for astrocyte reactions under EN exposure.
The increased synthesis of GFAP, the principal component of intermediate filaments (IF) of astrocytic cytoskeleton, is one of the best known manifestations of the early reaction of astrocytes against various classes of stimuli.[37] On one hand, the physiological conditions such as increased cell motility, migration, proliferation, vesicle trafficking, or changes on astrocyte-neuron interactions lead to glial reactions characterized by high-level expression of GFAP.[38] On the other hand, the pathological conditions such as trauma, ischemia, and neurodegeneration are also accompanied by hypertrophic changes on IF, a characteristic response identified as reactive gliosis.[20] Results obtained in our experiment confirmed that hippocampal GFAP is sensible to environmental stimuli, at least during the early stages of life. However, despite the fact that GFAP counts exhibited a clear enhancement, it is not clear whether these changes correspond to pathological gliosis or could it be a part of a more adaptive response. We believe on the basis of previous studies exploring long-term benefits of early-life stress[39] that changes reported here represent a more adaptive/plastic response. In any case, our results evidenced a convincing response of hippocampal astrocytes to chronic noise that clearly reflected a compromised microenvironment. Then, astrocyte reactions could be very helpful in assessing vulnerabilities of hippocampus and other non-auditory structures to noise.
Beyond the actual meaning of these morphological changes, our study also added evidence for mechanisms linking EN to high order non-auditory structures. Recent studies have argued that high-order cognitive abilities such as learning and memory become permanently affected under exposure to severe noise.[29,40] Since the hippocampus has been established as the main regulator of memory processes,[41,42] studies like ours could be relevant to better understand the effects of noise on cognition. Apart from us, a growing group of researchers have focused this question.[26,27,33,40,43,44,45,46] Liu et al.,[46] for example, found that noise was able to produce long-term deficit in spatial learning, and that this deficit was correlated with the degree of hearing loss and the decrease of hippocampal neurogenesis. Manikandan et al. [26] reported that exposure to chronic noise caused impairment in spatial memory, and that this impairment was associated with reduced dendrite branches in the hippocampus. Furthermore, using the same urban noise adaptation, Dr. Rabat and coworkers demonstrated that intermittent and randomly spaced noisy events produced long-term memory deficits in vulnerable rats;[27] a result confirmed in our laboratory that also evidenced long-term impairment in hippocampal neurogenesis.[29] So taken together, experiments above referred strongly suggest that the hippocampus is in fact a key structure in mediating the effects of noise on high-order cognitive processes. We extended these results by providing evidence that astrocytes also serve as mediators of these effects.
To conclude, our study provided evidence of mechanisms linking noise to non-classical auditory structures. We demonstrated that the hippocampus of young male rats is sensible to the effects EN. Astrocytes residing in the hippocampus of these animals may play an important role in the physiologic/pathologic response to EN. Since CORT levels were increased after noise exposure, we believe that these non-auditory effects were − at least in part − mediated by stress regulatory systems.
Financial support and sponsorship
This work was supported by research grants, IMSS R-2007-1305-7 to JGE, FOMIX-JAL 2008-05-99060 to SL, CONACyT 238313 to YRD, CONACyT 221092 to FJH and PROMEP-NPTC 236855 to YRD.
Conflicts of interest
There are no conflicts of interest.
Acknowledgements
We thank Dr. Arnoud Rabat for the donation of noise recordings. We also thank Dr. Horacio Rivera and Pastor Eric Andrae for critical review of the manuscript.
REFERENCES
- 1.Mochizuki K. [Effects of noise pollution] Hokenfu Zasshi. 1974;30:174–5. [PubMed] [Google Scholar]
- 2.Stansfeld S, Haines M, Brown B. Noise and health in the urban environment. Rev Environ Health. 2000;15:43–82. doi: 10.1515/reveh.2000.15.1-2.43. [DOI] [PubMed] [Google Scholar]
- 3.Barber JR, Crooks KR, Fristrup KM. The costs of chronic noise exposure for terrestrial organisms. Trends Ecol Evol. 2010;25:180–9. doi: 10.1016/j.tree.2009.08.002. [DOI] [PubMed] [Google Scholar]
- 4.Seidman M, Standring R. Noise and quality of life. Int J Environ Res Public Health. 2010;7:3730–8. doi: 10.3390/ijerph7103730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Kryter KD. Non-auditory effects of environmental noise. Am J Public Health. 1972;62:389–98. doi: 10.2105/ajph.62.3.389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Bogden JD. Detrimental health effects of noise pollution. J Med Soc N J. 1974;71:847–51. [PubMed] [Google Scholar]
- 7.Kight CR, Swaddle JP. How and why environmental noise impacts animals: An integrative, mechanistic review. Ecol Lett. 2011;14:1052–61. doi: 10.1111/j.1461-0248.2011.01664.x. [DOI] [PubMed] [Google Scholar]
- 8.Moore DR. Anatomy and physiology of binaural hearing. Audiology. 1991;30:125–34. doi: 10.3109/00206099109072878. [DOI] [PubMed] [Google Scholar]
- 9.Fruhholz S, Trost W, Kotz SA. The sound of emotions − Towards a unifying neural network perspective of affective sound processing. Neurosci Biobehav Rev. 2016;68:96–110. doi: 10.1016/j.neubiorev.2016.05.002. [DOI] [PubMed] [Google Scholar]
- 10.Schiff M. Nonauditory of effects of noise. Trans Am Acad Ophthalmol Otolaryngol. 1973;77:L384–398. [PubMed] [Google Scholar]
- 11.Westman JC, Walters JR. Noise and stress: A comprehensive approach. Environ Health Perspect. 1981;41:291–309. doi: 10.1289/ehp.8141291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Jacobson L, Sapolsky R. The role of the hippocampus in feedback regulation of the hypothalamic-pituitary-adrenocortical axis. Endocr Rev. 1991;12:118–34. doi: 10.1210/edrv-12-2-118. [DOI] [PubMed] [Google Scholar]
- 13.Kim JJ, Diamond DM. The stressed hippocampus, synaptic plasticity and lost memories. Nat Rev Neurosci. 2002;3:453–62. doi: 10.1038/nrn849. [DOI] [PubMed] [Google Scholar]
- 14.Joels M, Karst H, Alfarez D, Heine VM, Qin Y, van Riel E, et al. Effects of chronic stress on structure and cell function in rat hippocampus and hypothalamus. Stress. 2004;7:221–31. doi: 10.1080/10253890500070005. [DOI] [PubMed] [Google Scholar]
- 15.Jauregui-Huerta F, Ruvalcaba-Delgadillo Y, Gonzalez-Castaneda R, Garcia-Estrada J, Gonzalez-Perez O, Luquin S. Responses of glial cells to stress and glucocorticoids. Curr Immunol Rev. 2010;6:195–204. doi: 10.2174/157339510791823790. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Araque A, Parpura V, Sanzgiri RP, Haydon PG. Tripartite synapses: Glia, the unacknowledged partner. Trends Neurosci. 1999;22:208–15. doi: 10.1016/s0166-2236(98)01349-6. [DOI] [PubMed] [Google Scholar]
- 17.Fields RD, Stevens-Graham B. New insights into neuron-glia communication. Science. 2002;298:556–62. doi: 10.1126/science.298.5593.556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Kurosinski P, Gotz J. Glial cells under physiologic and pathologic conditions. Arch Neurol. 2002;59:1524–8. doi: 10.1001/archneur.59.10.1524. [DOI] [PubMed] [Google Scholar]
- 19.Halassa MM, Fellin T, Takano H, Dong JH, Haydon PG. Synaptic islands defined by the territory of a single astrocyte. J Neurosci. 2007;27:6473–7. doi: 10.1523/JNEUROSCI.1419-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Pekny M, Pekna M. Astrocyte intermediate filaments in CNS pathologies and regeneration. J Pathol. 2004;204:428–37. doi: 10.1002/path.1645. [DOI] [PubMed] [Google Scholar]
- 21.O’Callaghan JP, Brinton RE, McEwen BS. Glucocorticoids regulate the synthesis of glial fibrillary acidic protein in intact and adrenalectomized rats but do not affect its expression following brain injury. J Neurochem. 1991;57:860–9. doi: 10.1111/j.1471-4159.1991.tb08230.x. [DOI] [PubMed] [Google Scholar]
- 22.Vardimon L, Ben-Dror I, Avisar N, Oren A, Shiftan L. Glucocorticoid control of glial gene expression. J Neurobiol. 1999;40:513–27. doi: 10.1002/(sici)1097-4695(19990915)40:4<513::aid-neu8>3.0.co;2-d. [DOI] [PubMed] [Google Scholar]
- 23.Gould E, Tanapat P. Stress and hippocampal neurogenesis. Biol Psychiatry. 1999;46:1472–9. doi: 10.1016/s0006-3223(99)00247-4. [DOI] [PubMed] [Google Scholar]
- 24.de Kloet ER. Hormones, brain and stress. Endocr Regul. 2003;37:51–68. [PubMed] [Google Scholar]
- 25.Kim JJ, Song EY, Kosten TA. Stress effects in the hippocampus: Synaptic plasticity and memory. Stress. 2006;9:1–11. doi: 10.1080/10253890600678004. [DOI] [PubMed] [Google Scholar]
- 26.Manikandan S, Padma MK, Srikumar R, Jeya Parthasarathy N, Muthuvel A, Sheela Devi R. Effects of chronic noise stress on spatial memory of rats in relation to neuronal dendritic alteration and free radical-imbalance in hippocampus and medial prefrontal cortex. Neurosci Lett. 2006;399:17–22. doi: 10.1016/j.neulet.2006.01.037. [DOI] [PubMed] [Google Scholar]
- 27.Rabat A, Bouyer JJ, George O, Le Moal M, Mayo W. Chronic exposure of rats to noise: Relationship between long-term memory deficits and slow wave sleep disturbances. Behav Brain Res. 2006;171:303–12. doi: 10.1016/j.bbr.2006.04.007. [DOI] [PubMed] [Google Scholar]
- 28.Jauregui-Huerta F, Ruvalcaba-Delgadillo Y, Garcia-Estrada J, Feria-Velasco A, Ramos-Zuniga R, Gonzalez-Perez O, et al. Early exposure to noise followed by predator stress in adulthood impairs the rat’s re-learning flexibility in Radial Arm Water Maze. Neuro Endocrinol Lett. 2010;31:538–48. [PubMed] [Google Scholar]
- 29.Jauregui-Huerta F, Garcia-Estrada J, Ruvalcaba-Delgadillo Y, Trujillo X, Huerta M, Feria-Velasco A, et al. Chronic exposure of juvenile rats to environmental noise impairs hippocampal cell proliferation in adulthood. Noise Health. 2011;13:286–91. doi: 10.4103/1463-1741.82961. [DOI] [PubMed] [Google Scholar]
- 30.Rabat A, Bouyer JJ, Aran JM, Courtiere A, Mayo W, Le Moal M. Deleterious effects of an environmental noise on sleep and contribution of its physical components in a rat model. Brain Res. 2004;1009:88–97. doi: 10.1016/j.brainres.2004.02.046. [DOI] [PubMed] [Google Scholar]
- 31.Hernandez V, Luquín S, Jáuregui-Huerta F, Corona-Morales A, Medina M, Ruíz-Velasco S, et al. Dopamine receptor dysregulation in hippocampus of aged rats underlies chronic pulsatile l-Dopa treatment induced cognitive and emotional alterations. Neuropharmacology. 2014;82:88–100. doi: 10.1016/j.neuropharm.2013.11.013. [DOI] [PubMed] [Google Scholar]
- 32.Paxinos G, Watson C. The Rat Brain in Stereotaxic Coordinates. Amsterdam, Boston: Academic Press/Elsevier; 2007. [Google Scholar]
- 33.Gonzalez-Perez O, Chavez-Casillas O, Jauregui-Huerta F, Lopez-Virgen V, Guzman-Muniz J, Moy-Lopez N, et al. Stress by noise produces differential effects on the proliferation rate of radial astrocytes and survival of neuroblasts in the adult subgranular zone. Neurosci Res. 2011;70:243–50. doi: 10.1016/j.neures.2011.03.013. [DOI] [PubMed] [Google Scholar]
- 34.Bridges N, Slais K, Sykova E. The effects of chronic corticosterone on hippocampal astrocyte numbers: A comparison of male and female Wistar rats. Acta Neurobiol Exp (Wars) 2008;68:131–8. doi: 10.55782/ane-2008-1682. [DOI] [PubMed] [Google Scholar]
- 35.Lambert KG, Gerecke KM, Quadros PS, Doudera E, Jasnow AM, Kinsley CH. Activity-stress increases density of GFAP-immunoreactive astrocytes in the rat hippocampus. Stress. 2000;3:275–84. doi: 10.3109/10253890009001133. [DOI] [PubMed] [Google Scholar]
- 36.Barros VG, Duhalde-Vega M, Caltana L, Brusco A, Antonelli MC. Astrocyte-neuron vulnerability to prenatal stress in the adult rat brain. J Neurosci Res. 2006;83:787–800. doi: 10.1002/jnr.20758. [DOI] [PubMed] [Google Scholar]
- 37.Pekny M, Nilsson M. Astrocyte activation and reactive gliosis. Glia. 2005;50:427–34. doi: 10.1002/glia.20207. [DOI] [PubMed] [Google Scholar]
- 38.Middeldorp J, Hol EM. GFAP in health and disease. Prog Neurobiol. 2011;93:421–43. doi: 10.1016/j.pneurobio.2011.01.005. [DOI] [PubMed] [Google Scholar]
- 39.Jauregui-Huerta F, Zhang L, Yanez-Delgadillo G, Hernandez-Carrillo P, Garcia-Estrada J, Luquin S. Hippocampal cytogenesis and spatial learning in senile rats exposed to chronic variable stress: Effects of previous early life exposure to mild stress. Front Aging Neurosci. 2015;7:159. doi: 10.3389/fnagi.2015.00159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Cheng L, Wang SH, Chen QC, Liao XM. Moderate noise induced cognition impairment of mice and its underlying mechanisms. Physiol Behav. 2011;104:981–8. doi: 10.1016/j.physbeh.2011.06.018. [DOI] [PubMed] [Google Scholar]
- 41.O’Keefe J, Dostrovsky J. The hippocampus as a spatial map. Preliminary evidence from unit activity in the freely-moving rat. Brain Res. 1971;34:171–5. doi: 10.1016/0006-8993(71)90358-1. [DOI] [PubMed] [Google Scholar]
- 42.Altman J, Brunner RL, Bayer SA. The hippocampus and behavioral maturation. Behav Biol. 1973;8:557–96. doi: 10.1016/s0091-6773(73)80144-0. [DOI] [PubMed] [Google Scholar]
- 43.Arnsten AF, Goldman-Rakic PS. Noise stress impairs prefrontal cortical cognitive function in monkeys: Evidence for a hyperdopaminergic mechanism. Arch Gen Psychiatry. 1998;55:362–8. doi: 10.1001/archpsyc.55.4.362. [DOI] [PubMed] [Google Scholar]
- 44.Samson J, Sheeladevi R, Ravindran R, Senthilvelan M. Stress response in rat brain after different durations of noise exposure. Neurosci Res. 2007;57:143–7. doi: 10.1016/j.neures.2006.09.019. [DOI] [PubMed] [Google Scholar]
- 45.Ruvalcaba-Delgadillo Y, Luquín S, Ramos-Zúñiga R, Feria-Velasco A, González-Castañeda RE, Pérez-Vega MI, et al. Early life exposure to noise permanently reduces mPFC astrocyte numbers and T-maze alternation/discrimination task performance in male rats. Noise Health. 2015;17:216–26. doi: 10.4103/1463-1741.160703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Liu L, Shen P, He T, Chang Y, Shi L, Tao S, et al. Noise induced hearing loss impairs spatial learning/memory and hippocampal neurogenesis in mice. Sci Rep. 2016;6:20374. doi: 10.1038/srep20374. [DOI] [PMC free article] [PubMed] [Google Scholar]