

Abstract
The production of lignocellulosic-derived biofuels is a highly promising source of alternative energy, but it has been constrained by the lack of a microbial platform capable to efficiently degrade this recalcitrant material and cope with by-products that can be toxic to cells. Species that naturally grow in environments where carbon is mainly available as lignin are promising for finding new ways of removing the lignin that protects cellulose for improved conversion of lignin to fuel precursors. Enterobacter lignolyticus SCF1 is a facultative anaerobic Gammaproteobacteria isolated from tropical rain forest soil collected in El Yunque forest, Puerto Rico under anoxic growth conditions with lignin as sole carbon source. Whole transcriptome analysis of SCF1 during E.lignolyticus SCF1 lignin degradation was conducted on cells grown in the presence (0.1%, w/w) and the absence of lignin, where samples were taken at three different times during growth, beginning of exponential phase, mid-exponential phase and beginning of stationary phase. Lignin-amended cultures achieved twice the cell biomass as unamended cultures over three days, and in this time degraded 60% of lignin. Transcripts in early exponential phase reflected this accelerated growth. A complement of laccases, aryl-alcohol dehydrogenases, and peroxidases were most up-regulated in lignin amended conditions in mid-exponential and early stationary phases compared to unamended growth. The association of hydrogen production by way of the formate hydrogenlyase complex with lignin degradation suggests a possible value added to lignin degradation in the future.
Introduction
One of the key challenges that environmental sciences currently face is the search for renewable energy sources that are capable to satisfy future energy demands. Among several choices, biofuels derived from biomass have emerged as one of the most promising for decreasing the harmful effects produced by the uncontrolled use of fossil fuels [1, 2]. Lignocellulosic biomass represents close to the 90% of the dry weight of the total biomass material [3, 4]. Lignocellulose is composed of cellulosic and hemicellulosic polysaccharides, pectic polymers, and lignin. Lignin is a phenylpropanoid-derived heteropolymer that comprises 10–30% of the lignocellulosic biomass. The stable and complex physical structure of lignin together with its chemical inhibitory nature confers rigidity to plant tissue and also serving as a protection from pathogens [5, 6]. Due to its aromatic nature and highly branched polymer network, lignin is resistant to biological degradation and it is the primary material responsible for biomass recalcitrance [7]. Efforts aimed to advance lignin bioengineering in combination with novel lignin separation and biodegradation technologies are required to make lignocellulosic biofuels competitive with fossil fuels.
Microbial lignin degradation was thought to be a negligible bioprocess in the environment until 1928 when experiments carried out by Bavendam and collaborators showed that lignin could be biologically depolymerized [8]. These initial studies were supported by several studies carried out with white-rot or brown-rot fungi [9, 10]. In general, those microorganisms utilize extracellular peroxidases and laccases-mediated mechanisms during the aerobic degradation of lignin [5]. More recent studies have shown that lignin monomers such as vanillin, vanillic acid, ferulic acid, catechol and cinnamic acid can be metabolized under both aerobic and anaerobic conditions [11–14]. For instance, the phototrophic Rhodopseudomonas palustris [15], and the denitrifying Thauera aromatica [16] are well know because of their capability of anaerobic degradation of aromatic compounds. More recent experiments demonstrated that lignin can be degraded in the absence of oxygen [11, 17]. Facultative anaerobes have been shown to be capable of using lignin as the sole carbon source under anaerobic conditions, and we have isolated numerous lignin-degrading bacteria from El Yunque experimental forest, Puerto Rico based on their ability to grow on lignin as sole C source: Klebsiella sp. strain BRL6-2 [18], Tolumonas lignolytica sp. nov [19], and Enterobacter lignolyticus SCF1 [20].
Enterobacter lignolyticus SCF1 is a facultative anaerobic Gammaproteobacteria. Its close relative Enterobacter soli strain LF7, was also isolated from tropical forest soils in Peru using lignin as sole C source [21]. Although, these soils are characterized by periods of extremely low availability of oxygen and extended periods of low redox [22], tropical forest soils have been shown to have among the fastest rates of plant litter decomposition worldwide [23–25]. E. lignolyticus SCF1 sparked initial attention due to its ability to grow on lignin as a sole carbon source in the absence of oxygen, as well as its high tolerance to 1-ethyl-3-methylimidazolium chloride, a toxic compound utilized as a lignocellulosic biomass pretreatment in the production of biofuels [26]. These studies employed RNA-seq, a sensitive tool used to better understand the physiological status of bacteria by quantifying genome-wide gene expression [27]. Growth of E. lignolyticus SCF1 in lignin revealed increased 4-hydroxyphenylacetate degradation pathway, catalase/peroxidase enzymes, and the glutathione biosynthesis and glutathione S-transferase (GST) proteins, which have been previously implicated in anoxic lignin degradation. Growth on lignin was also associated with increased production of NADH-quinone oxidoreductase, other electron transport chain proteins, and ATP synthase and ATP-binding cassette (ABC) transporters, suggesting that lignin-amended conditions include extra reductive capacity in addition to observed oxidative and hydrolytic lignin-reducing enzymes [20].
RNA-seq studies are often conducted as static sampling experiments [28], based on the analysis of transcriptomes of microorganisms facing conditions at a specific time-point during growth. However, microorganisms respond to environmental changes in a dynamic fashion, which was the motivation for examining lignin degradation across three time points corresponding to different stages of growth. Our current experiment also differs from previous experiments due to the different substrate utilized. In former experiments, xylose, a pentose sugar commonly present in hydrolysates from lignocellulosic biomass was utilized as main C source. Those conditions may mimic slow-growing conditions that may be often found in soil environments. However, we observed that SCF-1 grew poorly during those experiments (achieving OD600 ~0.03) suggesting that this condition may have generated an additional stress that could obscure the global transcriptional response found when SCF-1 was incubated in the presence of lignin. Indeed, under those conditions, only the 6% (273/4599) of the genes were found to be differentially expressed when lignin was present [20]. In contrast, our current experiments were based on SCF-1 growing in glucose, which was chosen as a fast-growing condition, mimicking environmental conditions encountered in nutrient-rich ecosystems, such as those found by gastrointestinal microbiota. Both conditions differ not only in the affinity of their carbon source, but also in the quantity of energy (ATP) and reducing equivalents that the main pathways are capable to produce (S1 Fig). To improve our understanding of SCF1 lignin degradation over time [20], an experiment was designed to capture the gene expression profile of lignin degradation during growth (early, mid-exponential and beginning of stationary phase of growth) and in the presence and absence of lignin.
Materials and methods
Strains and culturing
The facultative anaerobe E. lignolyticus SCF1 was obtained from our laboratory culture collection. Cells were routinely grown in anaerobic modified LS4D minimal media as previously described [20], with the exception that PIPES (30 mM) was added instead of Tris-Cl (2 mM) as buffer and glucose (60 mM), instead of xylose, was added as sole carbon source. Since these conditions induced higher cell growth of E. lignolyticus SCF1 than was observed in previous experiments [20], we also duplicated the concentration of lignin supplied at the beginning of growth (0.1% w/w). The lignin source utilized for these experiments was alkali lignin (Sigma 45–471003) that according to the supplier’s specifications, it contained 4% of sulfur impurities. Cell growth was monitored by measuring the optical density at 600 nm (OD600) (Genesys 2, Spectronic Instruments, Rochester, NY). Because the lignin-amended media has a spectroscopic signature, lignin-amended cultures are reported with a blank subtraction using lignin-amended media, while unamended cultures are reported with a blank subtraction using unamended media. Lignin concentration was measured by removing 1 ml of culture from anaerobic septum bottles, diluting 1:10 in distilled water, filtering out cells, then measuring the absorbance at 310 nm and compared to a standard curve of known concentrations of lignin, as perfomed before [20, 29].
RNA-seq-based transcriptome analysis was conducted comparing cells grown in glucose (unamended control) to cells grown on both glucose and lignin (lignin-amended treatment). For this analysis, triplicates from the initial nine biological replicates of cells grown in both conditions were destructively sampled anaerobically for transcriptomics analysis at three time points. Samples for the first-time point were collected 3 hours after inoculation in both treatments, in early exponential phase (EE). Mid-exponential phase samples (ME) were collected 7 and 22 hours after inoculation in unamended control and lignin-amended cultures, respectively. The last time point was in early stationary phase (ES), for which samples were taken 22 and 47 hours after inoculation in unamended and lignin-amended cultures.
Metabolites analysis
High-pressure liquid chromatography (HPLC) to measure glucose, lactate, ethanol, formate, acetate, succinate in culture supernatants were performed on Shimadzu LC-20AD liquid chromatograph with a DGU-20A5 degasser and SIL-20 ACHT autosampler. The HPLC was equipped with an Aminex HPX-87H column, Biorad Microguard Cation H guard column and a RID-10A refractive index detector. The column was kept at 30°C with a flow rate of 0.6mL/min using 5 mM H2SO4 as the mobile phase. The coefficient of regression (R2) of all standards utilized were all over 0.99. All samples were run in triplicate. Lignin measurements were performed as previously described [19, 20]. Briefly, the absorbance of previously filtered (0.22 um) culture supernatants was measured at 310 nm and compared to a standard curve of known concentrations of lignin.
Hydrogen gas production was tested in separate growth conditions compared to cultures for transcriptomic analysis. Resting cell suspensions were prepared by harvesting cells at late exponential phase with an optical density of 0.18–0.2 by centrifugation for 12 minutes at 5000 RPM, and washed twice with washing buffer containing LS4D minimal media with no vitamins, trace elements, and C sources (glucose, lignin) added. Afterwards, cells were added into the washing buffer at a concentration of OD600 equal to 0.15, formate (150 mM) and lignin (0.05%) were added as C source and incubated anaerobically at 30°C. Controls were prepared with no addition of C source. Samples (200 μl) of headspace from each bottle at each time point were utilized to measure the amount of hydrogen gas using a gas chromatograph (Shimadzu GC-8A) equipped with a 60/80 Carboxen 1000 column (Supelco), and using Ar as the carrier gas.
mRNA extraction
Total RNA was extracted from harvested cells using Invitrogen TRIzol® Reagent (cat#15596018), followed by genomic DNA removal and cleaning using Qiagen RNase-Free DNase Set kit (cat#79254) and Qiagen Mini RNeasy™ kit (cat#74104). Agilent 2100 Bioanalyzer was used to assess the integrity of the RNA samples. Only RNA samples having RNA Integrity Number between 8–10 were used.
RNA-sequencing
The Applied Biosystems SOLiD™ Total RNA-Seq kit (catalog number 4445374) was used to generate the cDNA template library. The SOLiD™ EZ Bead system was used to perform emulsion clonal bead amplification to generate bead templates for SOLiD™ platform sequencing. Samples were sequenced on the SOLiD™ 4 platform. The 50-base short read sequences produced by the 5500xl SOLiD™ sequencer were mapped in color space using SOLiD™ LifeScope™ software version 2.5 using the default settings to map the short reads onto Enterobacter lignolyticus SCF1 (NC_014618.1) reference genome and both the fasta and the GFF files can be obtained from NCBI genome database (http://www.ncbi.nlm.nih.gov/genome). Sequence reads have been submitted to the NCBI databases under accession no GSE96828.
Statistical analysis
RNA-seq samples from the SOLiD sequencing platform were normalized and processed for differential expression using the edgeR package in Bioconductor [30]. Data was retained only from genes manifesting at least three reads per million in at least three individual replicates. Time points with glucose plus lignin were compared with matched time points with glucose alone. Genes with adjusted p-values <0.05 and absolute value of log2 fold change >1 were deemed differentially expressed for these comparisons. Venn diagrams were constructing using an online tool courtesy of the Bioinformatics and Research Computing group at the Whitehead Institute for Biomedical Research (http://jura.wi.mit.edu/bioc/tools/venn3way/), with the caveat that the sizes of the circles may not precisely be to scale. Histograms were generated with selected genes using the R programming language, with heatmaps made using the gplots package. Hierarchical clustering was performed using hclust function with the “complete” method setting and euclidian distance measures.
Results and discussion
To gain insight into physiological differences during lignin degradation, the gene transcript abundances in Enterobacter lignolyticus SCF1 were compared in cultures grown on glucose minimal media to the same media amended with lignin. Our experimental design was intended to characterize temporal dynamics during the degradation of lignin, as a way of illuminating the series of progressive biological mechanisms involved in lignin degradation. Populations were sampled for transcriptomics analysis at early exponential phase (EE), mid-exponential phase (ME) and early stationary phase (ES).
Global view of the response to lignin
SCF-1 achieved twice the cell density when lignin was added in the medium compared to unamended growth (Fig 1A, S2 and S3 Figs). This difference is associated with the degradation of lignin, since in the lignin-amended cultures there was a 40% lower consumption of glucose (~8 mM) than in the unamended cultures (Fig 1B and 1C). It has been previously shown that SCF-1 can utilize lignin as the sole carbon source during anaerobic growth in agar [20, 23]. Although we have been unable to detect growth of SCF-1 as sole C source in liquid medium, it has been previously reported that SCF-1 is capable to degrade lignin up to 56% within 48 hours in the presence of xylose [20]. When growing on lignin-amended LS4D liquid medium in the presence of glucose, there was a first drop in the content of lignin of about 30% (0–30 hours) at the beginning of the growth curve, that resulted in SCF-1 achieving a slightly higher cell concentration than the control. After 30 hours, the concentration of lignin was constantly decreasing until SCF-1 ultimately degraded 60% of the lignin amended in the medium.
Fig 1.
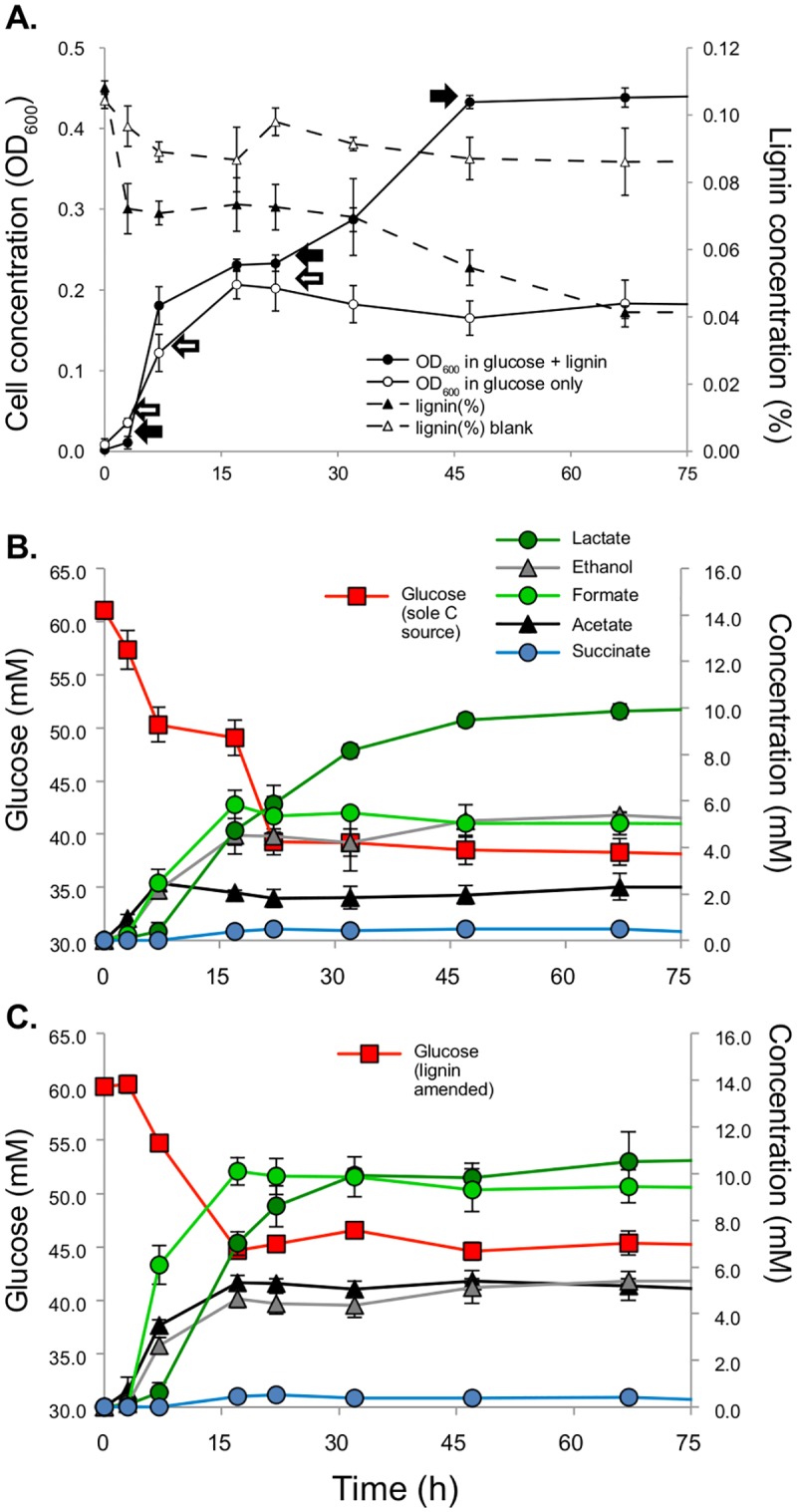
(A) Growth curve of Enterobacter lignolyticus SCF-1 in glucose medium (open circles) and glucose medium amended with lignin (closed circles). Arrows show the times samples were harvested for transcriptomic analysis (open arrows, glucose medium and closed arrows, glucose medium amended with lignin). The concentration of lignin of amended cultures is shown in closed triangles and control (medium mas lignin, no cells) is shown in open triangles. Metabolites concentrations along growth of unamended cultures (B) and lignin amended cultures (C). Glucose is shown in red squares, ethanol, in gray triangles, acetate, in black triangles, lactate, in darker green circles, succinate in light blue and formate, lighter green circles.
One third (34%) of the 4445 predicted protein-encoding ORFs in the genome of SCF1 were differentially expressed at any time point during growth on glucose amended with lignin (mean 1533 genes +/- 275 standard error, Table 1). The highest number of differentially expressed genes was detected in mid-exponential phase (ME, 2465). Genes that were over expressed in more than a single time point represented ~17% of the total pool of over expressed genes, suggesting that although the degradation lignin is a dynamic process, there is a significant proportion of genes for whose differential abundance levels were sustained over time (Fig 2A and 2B).
Table 1. Total genes up and down regulated in each of the three points monitored during growth of SCF1 in lignin-amended cultures.
The number in parenthesis indicates the percentage of the genes that were differentially expressed under each time.
| Expression level | Time point | Total | ||
|---|---|---|---|---|
| Early Exponential (EE) | Mid-Exponential (ME) | Early Stationary (ES) | ||
| Up regulated | 733 | 1308 | 442 | 2483 |
| Down regulated | 511 | 1157 | 448 | 2116 |
| Total | 1244 (27.9%) | 2465 (55.4%) | 890 (20%) | 4599 |
Fig 2.

Venn diagrams of differentially expressed genes, where (A) green circles indicate number of total genes are up-regulated each time point, and (B) red circles indicate number of total genes were down-regulated each time point. EE indicates cells analyzed in early exponential phase, ME, mid-exponential phase and ES, early stationary phase, where actual times of sampling each cell population is indicated in the methods as well as in Fig 1A. (C) Changes in the gene profile as a result of lignin exposure in early exponential phase, mid-exponential and early stationary phase of growth. In the right side of each chart (blue), number of genes with increased relative abundance, and in the left side (red), number of genes with lower relative abundance. The genes are grouped according to functional class as defined by COG annotation.
Genes with differential expression in the presence of lignin were classified under 20 categories according to their annotation function in the genome (S1 Data). Genes encoding hypothetical protein and proteins with unknown function (667) had the highest number of genes with greater abundance under lignin-amended growth, followed by genes encoding proteins with transporter activity (436). Genes involved in transcription and RNA metabolism (206), biosynthesis (201) and protein synthesis (180) were also represented in abundance in the differential expression data (Fig 2C).
Transcripts reflect the progression of activities associated with enhanced batch culture growth in the presence of lignin (Fig 3). Transcript production in response to lignin during early exponential phase growth was characterized by genes associated with biosynthesis and cell division. Catalase/peroxidase transcripts were over-expressed in lignin-amended compared to unamended conditions in every growth phase, but originating from different genes. These results suggest that lignin degradation is a very dynamic process that includes a wide set of genes involved in many cellular functions and takes place during the beginning of growth (early exponential) and the beginning of stationary phase, when the uptake of other substrates has stopped. By early stationary phase, waste products begin to accumulate as 60% of lignin has been degraded, and genes associated with continued expression of catalase/peroxidase enzymes and propanediol utilization related genes were found to be upregulated. The following sections describe in more detail the transcripts characterized by each growth phase.
Fig 3. Clustering of transcripts shows response to lignin in genes of interest in Enterobacter lignolyticus SCF1 grown in glucose with and without lignin.

Fold change indicates change in relative abundance of transcript in lignin-amended compared to unamended growth. Colors to the left of the heatmap indicate functional annotations as indicated. Each row represents gene expression of cells grown in lignin and glucose, compared to cells grown in glucose alone at the same time point.
Early exponential phase growth in lignin is characterized by accelerated growth and signaling
During early exponential phase, many genes controlling growth and biosynthesis of new cell material were up-regulated in the presence of lignin (Fig 2C, S2 Table). The large and small subunits ribosomal RNAs, the rod shape-determining operon, the membrane-bound lytic murein transglycosylase, genes involved in the bacterial outer membrane biogenesis, chromosome partitioning and cell division processes were all significantly upregulated when lignin was present compared to lignin unamended conditions.
Lignin degradation rates are constrained by the complexity of the substrate, and most bacterial strains regulate their metabolic pathways for utilization of carbon sources by carbon catabolite repression [31]. This process allows mixed sugars to be utilized only sequentially, and gene regulation attributed to this process was observed in SCF1 growth in the presence of lignin. Many genes related to transporters were found to be down-regulated when glucose was available in the medium, but they were switched on after glucose was no longer metabolized (S3 Table). Genes belonging to the phosphoenolpyruvate phosphotransferase system (PTS) related to the uptake of mannose (Entcl_3690, Entcl_3691, Entcl_3692, Entcl_3811 and Entcl_3812), xylose (Entcl_0175 and Entcl_0176) and ribose (Entcl_0174, Entcl_1204, Entcl_1206, Entcl_4081) were down-regulated in early exponential phase. However, after glucose was made unavailable due to concentrations dipping below its half-saturation constant for uptake, they were transcribed in higher abundance.
The initial down regulation of genes related to the uptake of other sugars was also observed in the case of transcripts associated with degradation of cellobiose (Entcl_2546 and Entcl_3764), chitobiose (Entcl_2547 and Entcl_2548), maltose (Entcl_3261) and mannose (Entcl_4032, Entcl_4033, Entcl_4034 and Entcl_4036). Furthermore, many genes involved in the uptake of other sugars beside glucose were up-regulated after glucose uptake was stopped (Fig 2C) (Entcl_0166, Entcl_0167, Entcl_1205, Entcl_1207, Entcl_3382, Entcl_3383, Entcl_4082, Entcl_4174, and Entcl_4403, S4 Table). This was presumably a response to the lack of availability of glucose, an effect that has been observed in other fermentative anaerobes [32].
Enterobacteriaceae is a family of Gram negative bacteria originally described as part of the gut microbiota. As being isolated from Puerto Rico tropical forest soils [23], SCF-1 may harbor metabolic capabilities toward survival in more oligotrophic environments, including environment-sensing mechanisms coupled to extra- and intra-cellular signal transduction pathways. For instance, SCF-1 has an overrepresentation of genes related to two-component regulatory system compared to those members of the Enterobacter genus that resides in the gut or plant microbiota (Table 2). Among those, transcripts of the three genes encoding the UhpABC system (encoded by the Entcl_0034-Entcl_0037 operon), a two-component response regulator involved in the transport of hexoses as well as the transcripts of hexose phosphate transporter, UhpT (encoded by Entcl_0038), were more abundant in early exponential growth with lignin [33]. Other components of the two-component regulatory system were also up-regulated, including the sensory histidine kinase QseC (encoded by Entcl_0059), the osmolarity sensory histidine kinase, EnvZ (encoded by Entcl_0331), the sensory histidine kinase, QseC (encoded by Entcl_0729), the hybrid sensory histidine kinase, EvgA (encoded by Entcl_1061), the sensory histidine kinase, AtoS (encoded by Entcl_1498), and the Osmosensitive K+ channel histidine kinase, KdpD (encoded by Entcl_3122). Most of those genes highlight the challenge that SCF-1 faces when is surrounded with environments where lignin is present, even at small concentrations.
Table 2. Summary of genome attributes of Enterobacter lignolyticus SCF1 and three close relatives, E. aerogenes, E. cloacae ATCC13047, and Enterobacter sp. 638.
| Enterobacter aerogenes | Enterobacter cloacae ATCC13047 | Enterobacter sp 638 | Enterobacter lignolyticus SCF1 | |
|---|---|---|---|---|
| Class | Gamma-proteobacteria | Gamma-proteobacteria | Gamma-proteobacteria | Gamma-proteobacteria |
| Mode of life | gut microbiota /opportunistic bacteria | gut microbiota /opportunistic bacteria | Endophytic bacteria | Tropical soil bacterium |
| Type of metabolism | facultative anaerobic | facultative anaerobic | aerobic | facultative aerobe; grows well under completely oxic and anoxic conditions |
| Size of the genome (Mb) | 5.27 | 5.59 | 4.67 | 4.81 |
| Genes related to two-component regulatory system | 21 | 20 | 27 | 33 |
| Genes related to secretion systems | 27 | 58 | 13 | 26 |
| References | [71] | [71] | [72] | [23] |
Many genes encoding proteins involved in resistance to environmental stresses were up-regulated in the presence of lignin, including the starvation sensing protein RspA (Entcl_1608), the starvation lipoprotein Slp paralog (Entcl_1986) and the universal stress protein family 3 (Entcl_2311). Carbon starvation protein A paralog (Entcl_3779) was also up-regulated in lignin-amended cultures after 7 hours, in agreement with former studies which found cstA to be upregulated at the protein and mRNA levels [20]. CstA, is a protein involved in escaping from starvation during nutrient scavenging and transition to stationary-phase growth [34] and has been shown to be regulated by the cyclic AMP (cAMP)-cAMP receptor protein complex, a global regulator involved in sugar metabolism [35]. Although the specific role of CstA in the metabolism of aromatic compounds remains to be elucidated, it has been demonstrated that carbon starvation proteins are induced by different aromatic pollutants [36], suggesting its involvement as a mechanism of adaptive response to the presence of a more recalcitrant carbon source [37]. Similar to cstA, the gene encoding the competence protein F (Entcl_0322) was found to be upregulated in lignin-amended cultures after 7 hours. This protein has also been shown to be part of the physiological response towards nutrient depravation, conferring cells the ability of uptake exogenous DNA and use it as a source of carbon and energy, providing an advantage during competition in nutrient limiting environments [38].
Within their wide set of tools to survive to dynamic environments, bacteria have evolved a way to respond to chemical or physical changes by modulating their behavior through gene expression. In one of those mechanisms, coined as "quorum sensing", bacteria respond to the concentration of a chemical molecule that reflects the bacterial population density allowing multicellular communication within a population [39]. One autoinducer, called AI-2, has been shown to be produced and recognized by several species allowing both intra and inter-cellular coordination [40]. We found that many genes related to the lsr operon (Entcl_0612, Entcl_0614, Entcl_0618 and Entcl_0619), the operon responsible for the uptake and processing of AI-2, were up-regulated in the presence of lignin.
Mid-exponential samples capture switch towards degradation of lignin
The degradation of lignin is a dynamic process, which was evident during mid-exponential phase of growth where lignin amended culture transcripts reflected a decrease in growth rate likely caused by a switch in metabolic growth from only consuming glucose to depolymerization of lignin (Fig 1). During this period, many genes encoding for the large subunits of ribosomal RNAs were in lower abundance compared to the high abundance detected in early exponential phase (S5 Table), likely due to the switch towards lignin degradation.
Lignin degrading bacteria must overcome three main challenges towards efficient utilization of lignin as substrate. First, the mechanism of degradation should be oxidative [41] Secondly, since lignin is mainly composed by large highly insoluble molecules, lignin depolymerization enzymatic machinery has to be extracellular, or having an extracellular mediator. Third, the systems must provide a pool of enzymes with low level of specificity since lignin stereochemistry is highly variable [41]. For the sake of simplification, bacterial lignin degradation could be divided in two stages. Initially, depolymerization releases lignin monomers from the complex lignin structure mainly by the cleavage of ether linkages [9, 35]. After monomers are released, enzymes in peripheral pathways proceed with the breakdown of these monomers into common intermediates that are further processed by the central metabolism [42].
The genome of SCF1 contains several genes encoding for enzymes that have been previously implicated in the degradation of lignin [20, 23]. Glycoside hydrolases are common carbohydrate-active enzymes involved in plant biomass degradation, capable of hydrolyzing the glycosidic bonds between carbohydrates or between carbohydrates and lignin [43]. The genome of SCF-1 has four homologs encoding glycoside hydrolase family 1 β-glucosidases. Three of them (Entcl_0991, Entcl_1274 and Entcl_3004) were expressed in higher abundance during mid-exponential growth of lignin-amended cultures (Table 3). The 6-phospho-beta-glucosidase (Entcl_0991) is located in the same operon as the cellobiose-specific form of the phosphotransferase system component (Entcl_0992), suggesting the existence of an operon devoted to the degradation of components of plant material. Previous studies have shown glycoside hydrolases were abundant in a soil-derived consortia adapted to recalcitrant carbon sources, such as complex plant polysaccharides and lignin, and also highly expressed in degradation of cellulose and hemicellulose by hindgut microbiota associated to termites [44, 45].
Table 3. Genes differentially regulated during growth related to lignin degradation.
Differential expression was defined as transcripts with adjusted p-values <0.05 and absolute value of log2 fold change >1 for these comparisons.
| Gene ID | Annotation | Gene name | Fold change in transcripts | ||
|---|---|---|---|---|---|
| EE | ME | ES | |||
| Entcl_1274 | 6-phospho-beta-glucosidase (EC 3.2.1.86) | 0.041 | 0.991 | -1.131 | |
| Entcl_3004 | 6-phospho-beta-glucosidase (EC 3.2.1.86) | 0.437 | 2.680 | -0.150 | |
| Entcl_0991 | 6-phospho-beta-glucosidase (EC 3.2.1.86) | ascB | -0.915 | 1.496 | -0.362 |
| Entcl_0992 | PTS system, arbutin-, cellobiose-, and salicin-specific IIBC component (EC 2.7.1.69) | -0.653 | 2.291 | 0.463 | |
| Entcl_0735 | putative laccase (EC 1.10.3.2) | 0.523 | -0.921 | 1.354 | |
| Entcl_0736 | Probable Fe-S oxidoreductase family 2 | 1.516 | 1.037 | 1.106 | |
| Entcl_0748 | Predicted oxidoreductases (related to aryl-alcohol dehydrogenases) | -0.626 | 1.684 | 0.883 | |
| Entcl_4301 | Catalase (EC 1.11.1.6) / Peroxidase (EC 1.11.1.7) | 0.560 | 3.932 | 0.146 | |
| Entcl_2769 | Flavoprotein wrbA | -1.502 | 3.638 | 0.700 | |
| Entcl_3180 | Alkyl hydroperoxide reductase protein F (EC 1.6.4.-) | 0.186 | 3.513 | 1.719 | |
| Entcl_3181 | Alkyl hydroperoxide reductase protein C (EC 1.6.4.-) | -0.010 | 1.468 | 1.126 | |
| Entcl_3797 | 4-hydroxyphenylacetate degradation bifunctional isomerase/decarboxylase, HpaG2 subunit | 0.111 | 0.525 | 1.889 | |
| Entcl_3800 | 5-carboxymethyl-2-hydroxymuconate delta-isomerase (EC 5.3.3.10) | 0.384 | 3.639 | 1.939 | |
| Entcl_3804 | Transcriptional activator of 4-hydroxyphenylacetate 3-monooxygenase operon, XylS/AraC family | 1.314 | 2.640 | -0.058 | |
| Entcl_3805 | 4-hydroxyphenylacetate 3-monooxygenase (EC 1.14.13.3) | 0.000 | 1.706 | -1.529 | |
| Entcl_2233 | Succinate-semialdehyde dehydrogenase [NADP+] | 0.340 | 1.543 | -0.248 | |
| Entcl_0876 | Succinate-semialdehyde dehydrogenase [NADP+] (EC 1.2.1.16) | -1.962 | 0.231 | 0.408 | |
| Entcl_2810 | Gamma-glutamyl-aminobutyraldehyde dehydrogenase (EC 1.2.1.-) | -1.292 | 0.011 | 0.519 | |
| Entcl_2233 | Succinate-semialdehyde dehydrogenase [NADP+] | 0.340 | 1.543 | -0.248 | |
During the degradation of lignin, many phenolic compounds are release into the medium. Those metabolites including acids (ferulic acid, vanillic acids and 4-hydroxybenzoic acid), alcohols (guaiacol and catechol, etc) and aldehydes (vanillin, 4-hydroxylbenzaldehyde, etc) have been reported to alter the permeability of cellular membranes at even small concentrations [46]. One of the strategies of microbes towards to cope with those stressful conditions is through the breakdown of phenols to nontoxic compounds. For instance, laccases (phenoloxidases, EC 1.10.3.2) are multicopper-containing enzymes capable of breaking phenols, aromatic and aliphatic C–C bonds by oxidation of phenolic units using molecular oxygen as the final electron acceptor [47, 48]. Although fungal laccases have been shown to play a key role during lignin degradation in the white-rot fungi [49–51], little is known about their function in their bacterial counterparts. Recent research has focused on the involvement of laccases during decomposition of lignin by strains of the soil-inhabiting microorganisms Streptomyces, where the deletion of the gene encoding a laccase partially impaired lignin degradation [52]. Previous evidence has suggested that the genome of SCF1 contains two putative laccases homolgs [20]. One of those genes, encoding a multicopper oxidase type 3 (Entcl _0735), was found to be up-regulated in the presence of lignin. Its adjacent gene (Entcl_0736) that seems to be co-transcribed with, correspond to an uncharacterized enzyme that contains Radical SAM domain protein using cobalamine (Vitamine B-12) as a cofactor. Several genes involved in the synthesis of cobalamine (Entcl_1738, Entcl_1762, Entcl_1763, Entcl_1764, Entcl_1765, Entcl_1766, Entcl_1767, Entcl_1768, and Entcl_1771) were found to be up-regulated in cultures were lignin was added. Furthermore, both laccases are predicted to be extracellular and periplasmic, respectively by PSORT [53] suggesting that they may be secreted. Under anaerobic growth conditions, the role laccases during lignin degradation remains questionable, due to the fact that those enzymes employ dioxygen as electron acceptor to carry out single electron oxidations of organic compounds. To this date, it is unclear how laccases could perform oxidation with an alternative electron acceptor. Recent evidence carried out with soil bacterium Stenotrophomonas maltophilia AAP56 found that laccase activity was highly dependent on the environment and it was greatly induced by anoxic conditions. More importantly, laccase activity is often induced by addition of CuSO4 [54]. In this example, laccase activity was greatly induced by the addition of 100 μM of CuSO4 in the γ-Proteobacterium JB, an alkali-tolerant soil isolate [55]. Therefore, as the trace element solution utilized in the growth medium has some Cu, and our lignin contains some sulfur impurities, we cannot rule out the possibility that laccases were induced by CuSO4, and not by lignin.
Aryl-alcohol dehydrogenases are enzymes capable of cleaving the β-aryl ether bonds, and have been shown to play a key role during depolymerization of lignin by white-rot fungi [56, 57]. As a part of this potential mechanism in SCF-1, we found that the transcripts of an aryl-alcohol dehydrogenases (Entcl_0748) were expressed in higher abundance in lignin-amended cultures (Table 3). Aryl-hydrogenases provide the hydrogen peroxide needed by ligninolytic high redox-potential peroxidases during fungal degradation of lignin [57]. Peroxidases aid in lignin degradation by removing one electron from the non-phenolic units of lignin [58]. Indeed, lignin peroxidases (EC 1.11.1.14) utilize hydrogen peroxide (H2O2) as the co-substrate in addition to a mediator, veratryl alcohol, to degrade lignin and other phenolic compounds. H2O2 is reduced to H2O by gaining an electron from the then oxidized LiP and then LiP release this electron to vetraryl alcohol that becomes aldehyde and gets reduced to its alcohol by gaining electrons from lignin [35]. The utilization of peroxidases in lignin degradation was previously hypothesized because SCF1 was isolated from the Luquillo LTER soils, where peroxidase activities were detected across a large area [20]. One peroxidase (encoded by Entcl_4301) and a flavoprotein disulfide reductase (encoded by Entcl_2769), an enzyme capable of sensing the local concentrations of hydrogen peroxide, were found to be up-regulated in the presence of lignin during mid-exponential phase, the period in which 60% of the lignin amended in the medium was utilized. Also, the genes encoding alkyl hydroperoxide reductases (encoded by Entcl_3180 and Entcl_3181), one of the main scavenger of H2O2 inside the cells, were found to be expressed in higher abundance in the presence of lignin, during the same sampling time. Although lignin peroxidases may be important enzymes for lignin degradation in SCF-1, the way in which hydrogen peroxide is provided at a constant rate under anaerobic conditions and its role during the anaerobic degradation of lignin remains unknown.
Another proposed mechanism for the degradation of lignin is via 4-hydroxyphenylacetate catabolic pathway. This pathway is composed by nine enzymes encoded by genes within a single genomic region (HpaRGEDFHIXABC) previously described as an aromatic degrading gene cluster. This cluster has been shown to be involved in the transformation of 4-hydroxyphenylacetate, mainly coming from the degradation of aromatic compounds, to intermediates of the central metabolism [23]. We found the gene encoding for the first gene of this pathway (Entcl_3797) to be up-regulated in the presence of lignin, suggesting that the pathway was turn on. Furthermore, three genes associated to this cluster (Entcl_3800, Entcl_3804 and Entcl_3805) were expressed in higher abundance when lignin was present. This pathway takes 4-hydroxyphenylacetate, a resulting compound from the fermentation of short-chain peptides and amino acids, and degrades it to pyruvate and succinate-semialdehyde [59, 60]. Because of its toxicity, succinate-semialdehyde has to be quickly converted to succinate via NADP+-dependent succinate semialdehyde dehydrogenase previous entering into the tricarboxylic acid cycle [61]. The genome of SCF-1 contains four homologs encoding for this enzyme (Entcl_2233, Entcl_2291, Entcl_0876 and Entcl_2810) and one of them (Entcl_2233) was expressed in higher abundance in lignin-amended cultures.
Other genes with a potential role during aromatic metabolism that support lignin degradation were found to be differentially expressed during lignin-amended growth. For instance, genes encoding for two anthranilate synthases (Entcl_2523 and Entcl_2524), a peripheral pathway that play an important role during anaerobic degradation of heterocyclic aromatic compounds, were found to be among top-5 upregulated genes in lignin amended cultures [42]. Also, genes encoding enzymes that belong to xanthine/uracil family permeases, such as Entcl_0083, Entcl_1586, Entcl_4435, Entcl_3516, Entcl_3019 and genes encoding for enzymes that catalyzes the further metabolization of xanthine, such as Entcl_4100, Entcl_3480, Entcl_2780, Entcl_2781, Entcl_2782, and Entcl_2795, were found to be up-regulated during lignin-amended mid-exponential growth. This set of enzymes belong to a mechanism of recycling of uric acid, xanthine and other nitrogenous products that has been previously reported to be relevant in the gut microbiome of wood-feeding beetles [62].
Lignin amendment was also associated with higher abundance of transcripts for genes associated with the uptake and metabolism of sugars. For example, the gene encoding for glucokinase (Entcl_1357), an enzyme that catalyzes the first step in glycolysis (reaction #1, S1 Fig), was found to be upregulated during mid-exponential growth, The expression of genes encoding for glucose-6-phosphate isomerase (Entcl_3693 and Entcl_3694) (reaction #2, S1 Fig), 6-phosphofructokinase class II (Entcl_2086 and Entcl_4346) (reaction #3, S1 Fig) and fructose-1,6-bisphosphatase, GlpX type (Entcl_0096) (reaction #4, S1 Fig) were also found to be expressed in higher abundance in early stationary phase. Similar behavior was observed in the genes encoding for the steps downstream in glycolysis, such as fructose-bisphosphate aldolase, class II (encoded by Entcl_4084) (reaction #5, S1 Fig), and the NAD-dependent glyceraldehyde-3-phosphate dehydrogenase (encoded by Entcl_2022).
Lignin-amended cultures produce very different metabolites by early-exponential phase
E. lignolyticus SCF1, as other enterobacteria, primarily carry out heteroenzymatic fermentation using pentose and hexose sugars as carbon and energy sources, and producing acetate, ethanol, formate and lactate, as main fermentation products [23]. A high concentration level of glucose remained in the medium after SCF-1 stopped growing, 45 mM in the presence of lignin and 40 mM in the control. The inability to keep metabolizing the substrates can be explained by the increased formation of acidic metabolites, such as acetate, formate, lactate, and succinate, which destroys the cell's ability to maintain internal pH [63].
Lignin amendment resulted in a significant difference in the production of fermentation subproducts, which may be attributed to the higher quantity of C atoms that have to be funneled into common intermediates of the central metabolism (pyruvate and Acetyl-CoA). For instance, the pyruvate formate-lyase, an enzyme in charge of supplying the citric acid cycle with acetyl-CoA during anaerobic glycolysis (encoded by Entcl_0584, Entcl_2993, Entcl_4043, Entcl_4296, Entcl_4297) and also its activating enzyme (encoded by Entcl_2992, Entcl_4042, and Entcl_4288; S6 Table) were expressed in higher abundance when lignin degradation rate was at its maximum. There was about 5 mM of more formate that was released by the cells in lignin-amended cultures (Fig 1C), though most of this accumulated in the first 15 hours of growth.
Many genes belonging to metabolic pathways devoted to degrade other components of plant material were also regulated under lignin amendments. For instance, several genes encoding enzymes involved in the degradation of pectin, a structural polysaccharide that is key for the stability of plant cell walls, were found to be upregulated (S7 Table). A considerable high number of genes (48) remained upregulated across the three growth phases underlying the idea that some of those genes may represent a set of metabolic functions that have to be constitutively expressed in the presence of lignin, rather than a part of a progressive metabolic or biological mechanism towards degradation of lignin (S8 Table).
Because of the increased formate production and transcription of formate hydrogenlyase (FHL) complex transcripts with lignin, we hypothesized that SCF-1 would produce hydrogen as an alternative to regulate pH in the lignin-amended treatments. Many members of Enterobacteriaceae, such as E.coli and Salmonella enterica, contain a membrane-associated FHL complex capable to utilize formic acid as electron donor and protons as electron acceptors during the production of H2 and CO2 [64, 65]. This complex allows the accumulation of the excess of reducing equivalents produced by fermentation into hydrogen, preventing further acidification of the medium and promoting interspecies H2 transfer. The FHL complex is composed of two main parts that catalyze hydrogen production from formate in two steps. The molybdenum-dependent formate dehydrogenase (FDHH) encoded by fdhF gene (Entcl_4087) catalyzes the reversible oxidation of formate to CO2 with the release of protons and two electrons. A second step is catalyzed by the NiFe hydrogenase, Hyd3, encoded by Entcl_0986, that utilize these electrons to further reduce protons to H2 [66]. Hyd3, belongs to the Group 4 [NiFe] hydrogenases, a phylogenetically distinct family of membrane-bound, cytoplasmically oriented hydrogenases that so far has been rarely characterized, and are predominantly involved in H2 production rather than H2 oxidation [67, 68]. Hyd3 differs from other hydrogenases because it allows accumulation of significant concentrations H2, without inhibition, making SCF-1 especially well-suited to biotechnological H2 production.
Cell suspensions were capable of anaerobically producing H2 in the presence of formate (150 mM) and lignin (0.05% w/w) (S4 Fig), which is evidence suggesting that the FHL complex is functional in E. lignolyticus SCF-1. However, during hydrogen production in cell suspensions, ~30% less H2 was produced in those incubations where lignin was added compared to formate added alone. In the RNA-seq results, the two components of formate hydrogenlyase complex, the formate dehydrogenase H (Entcl_4087), the seven subunits of the formate hydrogenlyase complex (Entcl_0982-Entcl_0988), its transcriptional activator (Entcl_0976) and the neighbor NiFe hydrogenase assembly enzymes (Entcl_0977-Entcl_0982) were down-regulated during the late stages of growth. On the other hand, genes belonging to the formate dehydrogenase type N (Entcl_2347–2349), an enzyme that generally oxidizes formate taking electrons towards dissimilatory nitrate reduction, were expressed in higher abundance when lignin was present after mid-exponential growth. We also observed a higher concentration of acetate in those amended cultures (Fig 1C). This overproduction could be attributed to the higher expression of Phosphate acetyltransferase (Entcl_1432) (reaction number 21, S1 Fig) and acetate kinase (Entcl_0583 and Entcl_1433) (reaction number 22, S1 Fig) in the earlier stages of growth.
Conclusions
Several omic techniques (genomics, transcriptomics, proteomics and metabolomics) are suitable to improve our understanding of microbial communities, enzyme interactions, and how lignocellulose breakdown occurs, in both isolated cultures or microbial communities [69, 70]. Although, there is current limited understanding of how aromatic metabolism is capable to support anaerobic lignin degradation, the set of up-regulated enzymes during lignin-amended growth could conceivably suggest their role during this process. By examining the degradation of lignin in batch culture over time, we were able to capture the transcripts involved in the initial accelerated growth and transport of glucose in the presence of lignin during early exponential phase, the diverse complement of lignin-degrading enzymes expressed during mid-exponential growth, and transporters and metabolic machinery responsible for the increased formate production observed concurrent with 60% of lignin reduced. This work will contribute to our understanding of anaerobic lignin degradation as well as the cellular machinery necessary to balance metabolism and manage toxic products formed as a result of growth in lignin amended media.
Supporting information
Positive numbers indicate upregulation and negative numbers indicate downregulation. T1 means beginning of exponential phase, T2 means mid-exponential phase, and T3 means beginning of stationary phase.
(XLSX)
Red and blue boxes indicate the consumption and production of the compound, respectively. Numbers in parenthesis indicate the name of the reaction as following: (1)ATP:D-glucose 6-phosphotransferase, EC 2.7.1.2, encoded by Entcl_1357; (2) Glucose-6-phosphate isomerase, EC 5.3.1.9, encoded by Entcl_3693, Entcl_4141; (3) ATP:D-fructose-6-phosphate-1-phosphotransferase, EC2.7.1.11, encoded by Entcl_2086 and Entcl_4346; (4) D-Fructose 1,6 biphosphate-1-phosphohydrolase, EC 3.1.3.11, encoded by Entcl_3933; (5) D-fructose 1,6 biphosphate D-glyceraldehyde 3-phosphate lyase, EC 4.1.2.13, encoded by Entcl_0829; (6) D-glyceraldehide-3-phosphate ketol-isomerase, EC 5.3.1.1, encoded by Entcl_0092; (7) glyceraldehyde-3-phosphate dehydrogenase, EC 1.2.1.12, encoded by Entcl_2214, Entcl_2615, and Entcl_2022; (8) phosphoglycerate kinase, EC 2.7.2.3, encoded by Entcl_0828; (9) phosphoglyceratemutase, EC 5.4.2.11, encoded by Entcl_3728, Entcl_0131, and Entcl_3075; (10) 2-phospho-D-glycerate hydro-lyase, EC 4.2.1.11, encoded by Entcl_0935; (11) phosphoenolpyruvatecarboxykinase (ATP), EC 4.1.1.49 encoded by Entcl_0332; (12) malate dehydrogenase, EC 1.1.1.37 encoded by Entcl_0474; (13) fumaratehydratase, EC 4.2.1.2, encoded by Entcl_1727, Entcl_1688, Entcl_2213, Entcl_2234; (14) succinate dehydrogenase, EC 1.3.5.1, encoded by Entcl_3102, Entcl_3103, Entcl_3104, and Entcl_3105; (15) phosphoenolpyruvate kinase, EC 2.7.1.40, encoded by Entcl_1936 and Entcl_2127; (16) Pyruvate-formatelyase, EC 2.3.1.54, encoded by Entcl_2993, Entcl_4043, Entcl_4289, Entcl_4296; Formatehydrogenlyase complex, encoded by Entcl_0982, Entcl_0983, Entcl_0985, Entcl_0986, Entcl_0987, and Entcl_0989; (18) pyruvate ferredoxin/flavodoxinoxidoreductase, EC 1.2.7.1, encoded by Entcl_2436; (19) L-lactate dehydrogenase, EC:1.1.1.28, encoded by Entcl_1569 and Entcl_3787; (20) Acetolactate synthase, EC 2.2.1.6, encoded by Entcl_0028, Entcl_3647, Entcl_3648, Entcl_4268, and Entcl_4269; (21) Phosphate acetyltransferase, EC 2.3.1.8, encoded by Entcl_1309, and Entcl_1432; (22) Acetate kinase, EC 2.7.2.1, encoded by Entcl_0583, Entcl_1740, and Entcl_1433; (23) Acetaldehyde dehydrogenase, EC 1.2.1.10, encoded by Entcl_2072, and Entcl_1745; (24) Alcohol dehydrogenase, EC 1.1.1.1, encoded by Entcl_2072, Entcl_0739, Entcl_1314, Entcl_1745, Entcl_3458, and Entcl_4294; (25) Xylose Isomerase, EC 5.3.1.5, encoded by Entcl_0177; (26) Xylulose kinase, EC 2.7.1.17, encoded by Entcl_0178; (27) L-ribulose-5-phosphate 4-epimerase, EC 5.1.3.4, encoded by Entcl_0156, Entcl_3664, Entcl_3966; (28) Phosphogluconate dehydrogenase, EC 1.1.1.44, encoded by Entcl_1664; (29) Gluconokinase, EC 2.7.1.12, encoded by Entcl_0304, and Entcl_3682; (30) Gluconolactonase, EC 3.1.1.17, encoded by Entcl_0174; and (31) 2-dehydro-3-deoxy-phosphogluconate aldolase, EC 4.1.2.14, encoded by Entcl_1940.
(DOCX)
We systematically found that the optical density of lignin-amended treatments were higher that unamended controls.
(DOCX)
(DOCX)
Cell suspensions were monitored over time for headspace accumulation of hydrogen gas (H2), which should have been produced as a result of formatehydrogenlyase activity.
(DOCX)
(DOCX)
Differential expression was defined as transcripts with adjusted p-values <0.05 and absolute value of log2 fold change >1 for these comparisons.
(DOCX)
Differential expression was defined as transcripts with adjusted p-values <0.05 and absolute value of log2 fold change >1 for these comparisons.
(DOCX)
Differential expression was defined as transcripts with adjusted p-values <0.05 and absolute value of log2 fold change >1 for these comparisons.
(DOCX)
Differential expression was defined as transcripts with adjusted p-values <0.05 and absolute value of log2 fold change >1 for these comparisons.
(DOCX)
Differential expression was defined as transcripts with adjusted p-values <0.05 and absolute value of log2 fold change >1 for these comparisons.
(DOCX)
Differential expression was defined as transcripts with adjusted p-values <0.05 and absolute value of log2 fold change >1 for these comparisons.
(DOCX)
Differential expression was defined as transcripts with adjusted p-values <0.05 and absolute value of log2 fold change >1 for these comparisons.
(DOCX)
Acknowledgments
This work was supported by the University of Massachusetts Amherst and the Joint BioEnergy Institute (http://www.jbei.org) supported by the US Department of Energy, Office of Science, Office of Biological and Environmental Research, under Contract No. DE-AC02-05CH11231. Some work was conducted at EMSL, a national scientific user facility sponsored by DOE Office of Biological and Environmental Research and located at Pacific Northwest National Laboratory. PNNL is operated by Battelle for the US DOE under contract AC06-76RLO1830. We also want to thank Begüm Topçuoğlu and Dr. Jim Holden for their assistance with the gas chromatography and hydrogen quantification.
Data Availability
RNA sequence reads have been submitted to the NCBI databases under accession number GSE96828.
Funding Statement
This work was supported by the University of Massachusetts Amherst and the Joint BioEnergy Institute (http://www.jbei.org) supported by the US Department of Energy, Office of Science, Office of Biological and Environmental Research, under Contract No. DE-AC02-05CH11231. Some work was conducted at EMSL (Environmental Molecular Sciences Laboratory), a national scientific user facility sponsored by DOE Office of Biological and Environmental Research and located at Pacific Northwest National Laboratory. PNNL is operated by Battelle for the US DOE under contract AC06-76RLO1830. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Lee SK, Chou H, Ham TS, Lee TS, Keasling JD. Metabolic engineering of microorganisms for biofuels production: from bugs to synthetic biology to fuels. Current Opinion in Biotechnology. 2008;19(6):556–63. 10.1016/j.copbio.2008.10.014. [DOI] [PubMed] [Google Scholar]
- 2.Ho DP, Ngo HH, Guo W. A mini review on renewable sources for biofuel. Bioresource Technology. 2014;169:742–9. 10.1016/j.biortech.2014.07.022. [DOI] [PubMed] [Google Scholar]
- 3.Ingram LO, Aldrich HC, Borges AC, Causey TB, Martinez A, Morales F, et al. Enteric bacterial catalysts for fuel ethanol production. Biotechnol Prog. 1999;15(5):855–66. 10.1021/bp9901062 [DOI] [PubMed] [Google Scholar]
- 4.Girio FM, Fonseca C, Carvalheiro F, Duarte LC, Marques S, Bogel-Lukasik R. Hemicelluloses for fuel ethanol: A review. Bioresour Technol. 2010;101(13):4775–800. 10.1016/j.biortech.2010.01.088 [DOI] [PubMed] [Google Scholar]
- 5.Bugg TDH, Ahmad M, Hardiman EM, Rahmanpour R. Pathways for degradation of lignin in bacteria and fungi. Natural Product Reports. 2011;28(12):1883–96. 10.1039/c1np00042j [DOI] [PubMed] [Google Scholar]
- 6.Ahmad M, Taylor CR, Pink D, Burton K, Eastwood D, Bending GD, et al. Development of novel assays for lignin degradation: comparative analysis of bacterial and fungal lignin degraders. Mol Biosyst. 2010;6(5):815–21. 10.1039/b908966g [DOI] [PubMed] [Google Scholar]
- 7.Eudes A, Liang Y, Mitra P, Loqué D. Lignin bioengineering. Current Opinion in Biotechnology. 2014;26:189–98. 10.1016/j.copbio.2014.01.002. [DOI] [PubMed] [Google Scholar]
- 8.Käärik A. The identification of the mycelia of wood-decay fungi by their oxidation reactions with phenolic compounds. Studia Forestalia Suecica. 1966:31–80. [Google Scholar]
- 9.Brown ME, Chang MCY. Exploring bacterial lignin degradation. Curr Opin Chem Biol. 2014;19:1–7. 10.1016/j.cbpa.2013.11.015 [DOI] [PubMed] [Google Scholar]
- 10.Lindeberg G. On the occurrence of polyphenol oxidases in soil-inhabiting Basidiomycetes. Physiologia Plantarum. 1948;1(2):196–205. 10.1111/j.1399-3054.1948.tb07124.x [DOI] [Google Scholar]
- 11.Young LY, Frazer AC. The fate of lignin and lignin-derived compounds in anaerobic environments. Geomicrobiology Journal. 1987;5(3–4):261–93. 10.1080/01490458709385973 [DOI] [Google Scholar]
- 12.Healy JB, Young LY, Reinhard M. Methanogenic decomposition of ferulic acid, a model lignin derivative. Appl Environ Microb. 1980;39(2):436–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Fuchs G, Boll M, Heider J. Microbial degradation of aromatic compounds—from one strategy to four. Nat Rev Micro. 2011;9(11):803–16. [DOI] [PubMed] [Google Scholar]
- 14.Kato S, Chino K, Kamimura N, Masai E, Yumoto I, Kamagata Y. Methanogenic degradation of lignin-derived monoaromatic compounds by microbial enrichments from rice paddy field soil. Scientific Reports. 2015;5:14295 10.1038/srep14295 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Dutton PL, Evans WC. The metabolism of aromatic compounds by Rhodopseudomonas palustris. A new, reductive, method of aromatic ring metabolism. The Biochemical journal. 1969;113(3):525–36. Epub 1969/07/01. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Schuhle K, Gescher J, Feil U, Paul M, Jahn M, Schagger H, et al. Benzoate-coenzyme A ligase from Thauera aromatica: an enzyme acting in anaerobic and aerobic pathways. J Bacteriol. 2003;185(16):4920–9. Epub 2003/08/05. 10.1128/JB.185.16.4920-4929.2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Akobi C, Yeo H, Hafez H, Nakhla G. Single-stage and two-stage anaerobic digestion of extruded lignocellulosic biomass. Applied Energy. 2016;184:548–59. 10.1016/j.apenergy.2016.10.039. [DOI] [Google Scholar]
- 18.Woo HL, Ballor NR, Hazen TC, Fortney JL, Simmons B, Davenport KW, et al. Complete genome sequence of the lignin-degrading bacterium Klebsiella sp. strain BRL6-2. Stand Genomic Sci. 2014;9(1):19 10.1186/1944-3277-9-19 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Billings AF, Fortney JL, Hazen TC, Simmons B, Davenport KW, Goodwin L, et al. Genome sequence and description of the anaerobic lignin-degrading bacterium Tolumonas lignolytica sp. nov. Stand Genomic Sci. 2015;10(1):106 10.1186/s40793-015-0100-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.DeAngelis KM, Sharma D, Varney R, Simmons B, Isern NG, Markilllie LM, et al. Evidence supporting dissimilatory and assimilatory lignin degradation in Enterobacter lignolyticus SCF1. Front Microbiol. 2013;4(280). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Manter DK, Hunter WJ, Vivanco JM. Enterobacter soli sp. nov.: a lignin-degrading gamma-proteobacteria isolated from soil. Curr Microbiol. 2011;62(3):1044–9. 10.1007/s00284-010-9809-9 [DOI] [PubMed] [Google Scholar]
- 22.Liptzin D, Silver WL, Detto M. Temporal dynamics in soil oxygen and greenhouse gases in two humid tropical forests. Ecosystems. 2011;14(2):171–82. 10.1007/s10021-010-9402-x [DOI] [Google Scholar]
- 23.DeAngelis KM, D'Haeseleer P, Chivian D, Fortney JL, Khudyakov J, Simmons B, et al. Complete genome sequence of "Enterobacter lignolyticus" SCF1. Stand Genomic Sci. 2011;5(1):69–85. 10.4056/sigs.2104875 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Parton W, Silver WL, Burke IC, Grassens L, Harmon ME, Currie WS, et al. Global-scale similarities in Nitrogen release patterns during long-term decomposition. Science. 2007;315(5810):361–4. 10.1126/science.1134853 [DOI] [PubMed] [Google Scholar]
- 25.Silver WL, Lugo AE, Keller M. Soil oxygen availability and biogeochemistry along rainfall and topographic gradients in upland wet tropical forest soils. Biogeochemistry. 1999;44(3):301–28. 10.1007/Bf00996995 [DOI] [Google Scholar]
- 26.Khudyakov JI, D’haeseleer P, Borglin SE, DeAngelis KM, Woo H, Lindquist EA, et al. Global transcriptome response to ionic liquid by a tropical rain forest soil bacterium, Enterobacter lignolyticus. Proceedings of the National Academy of Sciences. 2012;109(32):E2173–E82. 10.1073/pnas.1112750109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Wang Z, Gerstein M, Snyder M. RNA-Seq: a revolutionary tool for transcriptomics. Nature reviews Genetics. 2009;10(1):57–63. 10.1038/nrg2484 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Oh S, Song S, Dasgupta N, Grabowski G. The analytical landscape of static and temporal dynamics in transcriptome data. Frontiers in Genetics. 2014;5:35 10.3389/fgene.2014.00035 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Taylor CR, Hardiman EM, Ahmad M, Sainsbury PD, Norris PR, Bugg TD. Isolation of bacterial strains able to metabolize lignin from screening of environmental samples. J Appl Microbiol. 2012;113(3):521–30. 10.1111/j.1365-2672.2012.05352.x [DOI] [PubMed] [Google Scholar]
- 30.Robinson MD, McCarthy DJ, Smyth GK. edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics. 2010;26(1):139–40. 10.1093/bioinformatics/btp616 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Gorke B, Stulke J. Carbon catabolite repression in bacteria: many ways to make the most out of nutrients. Nat Rev Microbiol. 2008;6(8):613–24. 10.1038/nrmicro1932 [DOI] [PubMed] [Google Scholar]
- 32.Grimmler C, Held C, Liebl W, Ehrenreich A. Transcriptional analysis of catabolite repression in Clostridium acetobutylicum growing on mixtures of d-glucose and d-xylose. Journal of Biotechnology. 2010;150(3):315–23. 10.1016/j.jbiotec.2010.09.938. [DOI] [PubMed] [Google Scholar]
- 33.Webber CA, Kadner RJ. Involvement of the amino-terminal phosphorylation module of UhpA in activation of uhpT transcription in Escherichia coli. Mol Microbiol. 1997;24(5):1039–48. [DOI] [PubMed] [Google Scholar]
- 34.Yelle DJ, Kaparaju P, Hunt CG, Hirth K, Kim H, Ralph J, et al. Two-dimensional NMR evidence for cleavage of lignin and xylan substituents in wheat straw through hydrothermal pretreatment and enzymatic hydrolysis. Bioenerg Res. 2013;6(1):211–21. 10.1007/s12155-012-9247-6 [DOI] [Google Scholar]
- 35.Reiter J, Strittmatter H, Wiemann LO, Schieder D, Sieber V. Enzymatic cleavage of lignin beta-O-4 aryl ether bonds via net internal hydrogen transfer. Green Chem. 2013;15(5):1373–81. 10.1039/C3gc40295a [DOI] [Google Scholar]
- 36.Gu F, Posoknistakul P, Shimizu S, Yokoyama T, Jin YC, Matsumoto Y. Synergistic contribution of hydrosulfide and carbonate anions to the beta-O-4 bond cleavage of lignin model compounds in a green liquor pretreatment for enzymatic hydrolysis of lignocellulosic materials. J Wood Sci. 2014;60(5):346–52. 10.1007/s10086-014-1411-5 [DOI] [Google Scholar]
- 37.Lim SH, Nahm K, Ra CS, Cho DW, Yoon UC, Latham JA, et al. Effects of Alkoxy groups on arene rings of lignin beta-O-4 model compounds on the efficiencies of single electron transfer-promoted photochemical and enzymatic C-C bond cleavage reactions. J Org Chem. 2013;78(18):9431–43. 10.1021/jo401680z [DOI] [PubMed] [Google Scholar]
- 38.Blom A, Harder W, Matin A. Unique and overlapping pollutant stress proteins of Escherichia coli. Appl Environ Microb. 1992;58(1):331–4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Miller MB, Bassler BL. Quorum Sensing in Bacteria. Annual Review of Microbiology. 2001;55(1):165–99. 10.1146/annurev.micro.55.1.165 [DOI] [PubMed] [Google Scholar]
- 40.Federle MJ. Autoinducer-2-based chemical communication in bacteria: Complexities of interspecies signaling. Contributions to microbiology. 2009;16:18–32. 10.1159/000219371 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.De Souza WR. Microbial degradation of lignocellulosic biomass 2013. [Google Scholar]
- 42.Carmona M, Zamarro MT, Blazquez B, Durante-Rodriguez G, Juarez JF, Valderrama JA, et al. Anaerobic catabolism of aromatic compounds: a genetic and genomic view. Microbiol Mol Biol R. 2009;73(1):71–+. 10.1128/Mmbr.00021-08 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Berlemont R, Martiny AC. Genomic potential for polysaccharide deconstruction in bacteria. Appl Environ Microb. 2015;81(4):1513–9. 10.1128/aem.03718-14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Jimenez DJ, de Lima Brossi MJ, Schuckel J, Kracun SK, Willats WG, van Elsas JD. Characterization of three plant biomass-degrading microbial consortia by metagenomics- and metasecretomics-based approaches. Appl Microbiol Biotechnol. 2016;100(24):10463–77. 10.1007/s00253-016-7713-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Warnecke F, Luginbuhl P, Ivanova N, Ghassemian M, Richardson TH, Stege JT, et al. Metagenomic and functional analysis of hindgut microbiota of a wood-feeding higher termite. Nature. 2007;450(7169):560–5. 10.1038/nature06269 [DOI] [PubMed] [Google Scholar]
- 46.Ibraheem O, Ndimba BK. Molecular adaptation mechanisms employed by ethanologenic bacteria in response to lignocellulose-derived inhibitory compounds. International Journal of Biological Sciences. 2013;9(6):598–612. 10.7150/ijbs.6091 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.ten Have R, Teunissen PJM. Oxidative mechanisms involved in lignin degradation by white-rot fungi. Chem Rev. 2001;101(11):3397–413. 10.1021/Cr000115l [DOI] [PubMed] [Google Scholar]
- 48.Rico A, Rencoret J, del Rio JC, Martinez AT, Gutierrez A. Pretreatment with laccase and a phenolic mediator degrades lignin and enhances saccharification of Eucalyptus feedstock. Biotechnol Biofuels. 2014;7 Artn 6 10.1186/1754-6834-7-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Mäkelä MR, Lundell T, Hatakka A, Hildén K. Effect of copper, nutrient nitrogen, and wood-supplement on the production of lignin-modifying enzymes by the white-rot fungus Phlebia radiata. Fungal Biology. 2013;117(1):62–70. 10.1016/j.funbio.2012.11.006. [DOI] [PubMed] [Google Scholar]
- 50.Parenti A, Muguerza E, Redin Iroz A, Omarini A, Conde E, Alfaro M, et al. Induction of laccase activity in the white rot fungus Pleurotus ostreatus using water polluted with wheat straw extracts. Bioresource Technology. 2013;133(0):142–9. 10.1016/j.biortech.2013.01.072. [DOI] [PubMed] [Google Scholar]
- 51.Record E, Punt PJ, Chamkha M, Labat M, van den Hondel CAMJJ, Asther M. Expression of the Pycnoporus cinnabarinus laccase gene in Aspergillus niger and characterization of the recombinant enzyme. Eur J Biochem. 2002;269(2):602–9. 10.1046/j.0014-2956.2001.02690.x [DOI] [PubMed] [Google Scholar]
- 52.Majumdar S, Lukk T, Solbiati JO, Bauer S, Nair SK, Cronan JE, et al. Roles of small laccases from Streptomyces in lignin degradation. Biochemistry-Us. 2014;53(24):4047–58. 10.1021/Bi500285t [DOI] [PubMed] [Google Scholar]
- 53.Yu NY, Wagner JR, Laird MR, Melli G, Rey S, Lo R, et al. PSORTb 3.0: improved protein subcellular localization prediction with refined localization subcategories and predictive capabilities for all prokaryotes. Bioinformatics. 2010;26(13):1608–15. 10.1093/bioinformatics/btq249 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Galai S, Touhami Y, Marzouki MN. Response surface methodology applied to laccases activities exhibited by Stenotrophomonas maltophilia AAP56 in different growth conditions 2012. [Google Scholar]
- 55.Singh G, Capalash N, Goel R, Sharma P. A pH-stable laccase from alkali-tolerant γ-proteobacterium JB: Purification, characterization and indigo carmine degradation. Enzyme and Microbial Technology. 2007;41(6):794–9. 10.1016/j.enzmictec.2007.07.001. [DOI] [Google Scholar]
- 56.Sjostrom E. Chapter 4—LIGNIN Wood Chemistry (Second Edition). San Diego: Academic Press; 1993. p. 71–89. [Google Scholar]
- 57.Ferreira P, Hernandez-Ortega A, Herguedas B, Martínez ÁT, Medina M. Aryl-alcohol oxidase involved in lignin degradation: A mechanistic study based on steady and pre-steady state kinetics and primary and solvent Isotope effects with two alcohol substrates. Journal of Biological Chemistry. 2009;284(37):24840–7. 10.1074/jbc.M109.011593 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Hatakka A. Biodegradation of Lignin Biopolymers Online: Wiley-VCH Verlag GmbH & Co. KGaA; 2005. [Google Scholar]
- 59.Smith EA, Macfarlane GT. Enumeration of human colonic bacteria producing phenolic and indolic compounds: effects of pH, carbohydrate availability and retention time on dissimilatory aromatic amino acid metabolism. J Appl Bacteriol. 1996;81(3):288–302. [DOI] [PubMed] [Google Scholar]
- 60.Prieto MA, Díaz E, García JL. Molecular characterization of the 4-hydroxyphenylacetate catabolic pathway of Escherichia coli W: engineering a mobile aromatic degradative cluster. Journal of Bacteriology. 1996;178(1):111–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Fuhrer T, Chen L, Sauer U, Vitkup D. Computational prediction and experimental verification of the gene encoding the NAD(+)/NADP(+)-dependent succinate semialdehyde dehydrogenase in Escherichia coli. Journal of Bacteriology. 2007;189(22):8073–8. 10.1128/JB.01027-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Scully ED, Geib SM, Carlson JE, Tien M, McKenna D, Hoover K. Functional genomics and microbiome profiling of the Asian longhorned beetle (Anoplophora glabripennis) reveal insights into the digestive physiology and nutritional ecology of wood feeding beetles. BMC Genomics. 2014;15(1):1096 10.1186/1471-2164-15-1096 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Bowles LK, Ellefson WL. Effects of butanol on Clostridium acetobutylicum. Appl Environ Microb. 1985;50(5):1165–70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Zbell AL, Maier RJ. Role of the Hya hydrogenase in recycling of anaerobically produced H2 in Salmonella enterica Serovar typhimurium. Appl Environ Microb. 2009;75(5):1456–9. 10.1128/aem.02064-08 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Axley MJ, Grahame DA, Stadtman TC. Escherichia coli formate-hydrogen lyase. Purification and properties of the selenium-dependent formate dehydrogenase component. Journal of Biological Chemistry. 1990;265(30):18213–8. [PubMed] [Google Scholar]
- 66.Bagramyan K, Trchounian A. Structural and functional features of formate hydrogen lyase, an enzyme of mixed-Acid fermentation from Escherichia coli. Biochemistry (Moscow). 2003;68(11):1159–70. 10.1023/B:BIRY.0000009129.18714.a4 [DOI] [PubMed] [Google Scholar]
- 67.Coppi MV. The hydrogenases of Geobacter sulfurreducens: a comparative genomic perspective. Microbiology. 2005;151(4):1239–54. 10.1099/mic.0.27535-0 [DOI] [PubMed] [Google Scholar]
- 68.McDowall JS, Murphy BJ, Haumann M, Palmer T, Armstrong FA, Sargent F. Bacterial formate hydrogenlyase complex. Proceedings of the National Academy of Sciences. 2014;111(38):E3948–E56. 10.1073/pnas.1407927111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Rosnow JJ, Anderson LN, Nair RN, Baker ES, Wright AT. Profiling microbial lignocellulose degradation and utilization by emergent omics technologies. Critical Reviews in Biotechnology. 2016:1–15. 10.1080/07388551.2016.1209158 [DOI] [PubMed] [Google Scholar]
- 70.Kameshwar AkS, Qin W. Recent developments in using advanced sequencing technologies for the genomic studies of lignin and cellulose degrading microorganisms. International Journal of Biological Sciences. 2016;12(2):156–71. 10.7150/ijbs.13537 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Davin-Regli A, Pagès J-M. Enterobacter aerogenes and Enterobacter cloacae; versatile bacterial pathogens confronting antibiotic treatment. Frontiers in Microbiology. 2015;6:392 10.3389/fmicb.2015.00392 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Taghavi S, van der Lelie D, Hoffman A, Zhang YB, Walla MD, Vangronsveld J, et al. Genome sequence of the plant growth promoting endophytic bacterium Enterobacter sp. 638. PLoS Genet. 2010;6(5):1000943. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Positive numbers indicate upregulation and negative numbers indicate downregulation. T1 means beginning of exponential phase, T2 means mid-exponential phase, and T3 means beginning of stationary phase.
(XLSX)
Red and blue boxes indicate the consumption and production of the compound, respectively. Numbers in parenthesis indicate the name of the reaction as following: (1)ATP:D-glucose 6-phosphotransferase, EC 2.7.1.2, encoded by Entcl_1357; (2) Glucose-6-phosphate isomerase, EC 5.3.1.9, encoded by Entcl_3693, Entcl_4141; (3) ATP:D-fructose-6-phosphate-1-phosphotransferase, EC2.7.1.11, encoded by Entcl_2086 and Entcl_4346; (4) D-Fructose 1,6 biphosphate-1-phosphohydrolase, EC 3.1.3.11, encoded by Entcl_3933; (5) D-fructose 1,6 biphosphate D-glyceraldehyde 3-phosphate lyase, EC 4.1.2.13, encoded by Entcl_0829; (6) D-glyceraldehide-3-phosphate ketol-isomerase, EC 5.3.1.1, encoded by Entcl_0092; (7) glyceraldehyde-3-phosphate dehydrogenase, EC 1.2.1.12, encoded by Entcl_2214, Entcl_2615, and Entcl_2022; (8) phosphoglycerate kinase, EC 2.7.2.3, encoded by Entcl_0828; (9) phosphoglyceratemutase, EC 5.4.2.11, encoded by Entcl_3728, Entcl_0131, and Entcl_3075; (10) 2-phospho-D-glycerate hydro-lyase, EC 4.2.1.11, encoded by Entcl_0935; (11) phosphoenolpyruvatecarboxykinase (ATP), EC 4.1.1.49 encoded by Entcl_0332; (12) malate dehydrogenase, EC 1.1.1.37 encoded by Entcl_0474; (13) fumaratehydratase, EC 4.2.1.2, encoded by Entcl_1727, Entcl_1688, Entcl_2213, Entcl_2234; (14) succinate dehydrogenase, EC 1.3.5.1, encoded by Entcl_3102, Entcl_3103, Entcl_3104, and Entcl_3105; (15) phosphoenolpyruvate kinase, EC 2.7.1.40, encoded by Entcl_1936 and Entcl_2127; (16) Pyruvate-formatelyase, EC 2.3.1.54, encoded by Entcl_2993, Entcl_4043, Entcl_4289, Entcl_4296; Formatehydrogenlyase complex, encoded by Entcl_0982, Entcl_0983, Entcl_0985, Entcl_0986, Entcl_0987, and Entcl_0989; (18) pyruvate ferredoxin/flavodoxinoxidoreductase, EC 1.2.7.1, encoded by Entcl_2436; (19) L-lactate dehydrogenase, EC:1.1.1.28, encoded by Entcl_1569 and Entcl_3787; (20) Acetolactate synthase, EC 2.2.1.6, encoded by Entcl_0028, Entcl_3647, Entcl_3648, Entcl_4268, and Entcl_4269; (21) Phosphate acetyltransferase, EC 2.3.1.8, encoded by Entcl_1309, and Entcl_1432; (22) Acetate kinase, EC 2.7.2.1, encoded by Entcl_0583, Entcl_1740, and Entcl_1433; (23) Acetaldehyde dehydrogenase, EC 1.2.1.10, encoded by Entcl_2072, and Entcl_1745; (24) Alcohol dehydrogenase, EC 1.1.1.1, encoded by Entcl_2072, Entcl_0739, Entcl_1314, Entcl_1745, Entcl_3458, and Entcl_4294; (25) Xylose Isomerase, EC 5.3.1.5, encoded by Entcl_0177; (26) Xylulose kinase, EC 2.7.1.17, encoded by Entcl_0178; (27) L-ribulose-5-phosphate 4-epimerase, EC 5.1.3.4, encoded by Entcl_0156, Entcl_3664, Entcl_3966; (28) Phosphogluconate dehydrogenase, EC 1.1.1.44, encoded by Entcl_1664; (29) Gluconokinase, EC 2.7.1.12, encoded by Entcl_0304, and Entcl_3682; (30) Gluconolactonase, EC 3.1.1.17, encoded by Entcl_0174; and (31) 2-dehydro-3-deoxy-phosphogluconate aldolase, EC 4.1.2.14, encoded by Entcl_1940.
(DOCX)
We systematically found that the optical density of lignin-amended treatments were higher that unamended controls.
(DOCX)
(DOCX)
Cell suspensions were monitored over time for headspace accumulation of hydrogen gas (H2), which should have been produced as a result of formatehydrogenlyase activity.
(DOCX)
(DOCX)
Differential expression was defined as transcripts with adjusted p-values <0.05 and absolute value of log2 fold change >1 for these comparisons.
(DOCX)
Differential expression was defined as transcripts with adjusted p-values <0.05 and absolute value of log2 fold change >1 for these comparisons.
(DOCX)
Differential expression was defined as transcripts with adjusted p-values <0.05 and absolute value of log2 fold change >1 for these comparisons.
(DOCX)
Differential expression was defined as transcripts with adjusted p-values <0.05 and absolute value of log2 fold change >1 for these comparisons.
(DOCX)
Differential expression was defined as transcripts with adjusted p-values <0.05 and absolute value of log2 fold change >1 for these comparisons.
(DOCX)
Differential expression was defined as transcripts with adjusted p-values <0.05 and absolute value of log2 fold change >1 for these comparisons.
(DOCX)
Differential expression was defined as transcripts with adjusted p-values <0.05 and absolute value of log2 fold change >1 for these comparisons.
(DOCX)
Data Availability Statement
RNA sequence reads have been submitted to the NCBI databases under accession number GSE96828.