

Abstract
In the face of an “epidemic” increase in myopia over the last decades and myopia prevalence predicted to reach 2.5 billion people by the end of this decade, there is an urgent need to develop effective and safe therapeutic interventions to slow down this “myopia booming” and prevent myopia-related complications and vision loss. Dopamine (DA) is an important neurotransmitter in the retina and mediates diverse functions including development, visual signaling, and refractive development. Inspired by the convergence of epidemiological and animal studies in support of the inverse relationship between outdoor activity and risk of developing myopia and by the close biological relationship between light exposure and dopamine release/signaling, we felt it is timely and important to critically review the role of DA in myopia development. This review will revisit several key points of evidence for and against DA mediating light control of myopia: 1) the causal role of extracellular retinal DA levels, 2) the mechanism and action of dopamine D1 and D2 receptors and 3) the roles of cellular/circuit retinal pathways. We examine the experiments that show causation by altering DA, DA receptors and visual pathways using pharmacological, transgenic, or visual environment approaches. Furthermore, we critically evaluate the safety issues of a DA-based treatment strategy and some approaches to address these issues. The review identifies the key questions and challenges in translating basic knowledge on DA signaling and myopia from animal studies into effective pharmacological treatments for myopia in children.
Keywords: myopia, dopamine, D2 receptor, D1 receptor, apomorphine
1. Introduction
Over the last decades, we are witnessing an “epidemic” increase in myopia: 90%–95% of teenagers and young adults are myopic in China and Seoul; and about half of the young adults in the United States and Europe are also affected by myopia. It is projected that myopia will affect one-third of the world’s population — 2.5 billion people by the end of this decade (Dolgin, 2015). Thus, there is an urgent need to develop effective and safe therapeutic interventions to slow down this “myopia booming”.
As reported at the 15th International Myopia Conference (IMC 2015, Wenzhou, China) and in the literature, there is a convergence of epidemiological and animal studies in support of the inverse relationship between bright light exposure and risk of developing myopia (for example, (Jones et al., 2007; Rose et al., 2008)). There is a critical need to understand the biological mechanism for this relationship in order to recommend the most effective interventions. Dopamine (DA) is an important neurotransmitter in the retina that mediates diverse functions including development, visual signaling, and refractive development (Witkovsky, 2004).
Since the first publication for a link between DA and the control of eye growth in 1989 (Stone et al., 1989), a growing body of work (with total ~100 publications so far) has led to the DA hypothesis (i.e. the release of DA in the retina to antagonize myopia development) as the leading hypothesis of myopia control (Stone et al., 1989). Furthermore, the light hypothesis, i.e. light-stimulated DA antagonizes myopia development (Rose et al., 2008) has been supported by a number of studies that show outdoor activity and/or bright light inhibits myopia, potentially through DA-mediated mechanisms (Ashby et al., 2009; Chen et al., 2017; Cohen et al., 2011; Cohen et al., 2012; Li et al., 2014; Morgan et al., 2012; Smith et al., 2012; Wang et al., 2015; Wu et al., 2013). The evidence from pharmacological and genetic studies has provided more details regarding how light or visual input controls retinal DA signaling and contributes to refractive development (for a recent review, see (Feldkaemper and Schaeffel, 2013)).
There is increasing recognition of the urgent need for effective pharmacological treatments that are deeply rooted in mechanistic understanding of the biological effect of DA and other neuromodulators. Extensive studies have investigated DA pharmacological effects on myopia from mammals and non-mammal models of myopia, supporting the critical role of DA in mediating control of myopia development. However, DA pharmocology studies of myopia development are limited by their partial specificity that likely contribute to some discrepancies of pharmacology in these various species used in myopia research. Furthermore, since DA receptors are expressed in various cell types and different vision pathways, DA receptors in these various sites may produce distinct/different effects on myopia development. In this review we seek to identify several key conceptual questions and challenges, with particular focus and insights from mouse model of myopia and related genetic and other molecular approaches. Though the disadvantages of the mouse model is also conspicuous because of its small eye size and the differences in ocular anatomy when compared to human, for example, the poor visual acuity and the lack of accommodation and fovea in mouse eye, this focus allows us to overcome these intrinsic limitations of DA pharmacology with the availability of different types of transgenic mice with targeted manipulation of vision pathways or DA signaling proteins. After reviewing the multiple lines of the evidence for the role of DA in mediating light control of myopia and the associated cellular and receptor mechanisms, we review the experiments in which DA, DA receptors and vision pathways have been altered using pharmacological, genetic, or visual environment approaches to show causation. Furthermore, we critically evaluate the safety issues of a DA-based treatment strategy and some approaches to address these issues. By revisiting several key conceptual questions with the evidence for and against these key points, we identify the key challenges ahead in order to create ground-breaking knowledge on myopia development and treatment.
2. Effects of altered DA levels on myopic eye growth
2.1 DA synthesis, release and metabolism
Retinal DA is synthesized in and released by subtypes of amacrine and interplexiform cells (e.g., (Dowling and Ehinger, 1978a, b; Frederick et al., 1982)). Dopamine is synthesized from the amino acid tyrosine in two steps: (1) L-tyrosine is converted to 3,4-dihydroxy-L-phenylalanine (L-DOPA) by tyrosine hydroxylase (TH); and (2) conversion of L-DOPA to DA by DOPA decarboxylase. Tyrosine hydroxylation is the rate-limiting and highly-regulated step in the synthesis of DA. Newly synthesized DA is transported into synaptic vesicles by the vesicular monoamine transporter VMAT2. Light stimulates the synthesis and release of DA, with a concomitant activation of TH that maintains steady-state stores of DA under conditions of enhanced neuronal activity (Iuvone et al., 1978; Kramer, 1971). When released, DA acts on postsynaptic and presynaptic receptors. DA also diffuses away from the synapse to act on extrasynaptic DA receptors by a process referred to as volume transmission (Bjelke et al., 1996). DA released at synapses is transported back into the DA neurons by the specific dopamine transporter (DAT) (Mitsuma et al., 1998). Inside the cell it may be repackaged in synaptic vesicles or metabolized by monoamine oxidase to form 3,4-dihydroxyphenylacetic acid (DOPAC), the primary metabolite of DA in the retina (Cohen et al., 1983). Under most conditions, changes in the level of DOPAC in the retina and vitreous are thought to reflect DA release and turnover (Megaw et al., 2001; Ohngemach et al., 1997). DOPAC may be further metabolized extra-neuronally to homovanillic acid (HVA) by catechol-O-methyltransferase (Meiser et al., 2013).
2.2 Studies implicating a role for DA in myopia
Data from several experiments across different species suggest that DA acts as a “stop” signal in refractive eye growth (Feldkaemper and Schaeffel, 2013). This is based on HPLC results of retinal or vitreal DA and/or DOPAC that show reduced levels in response to form deprivation in primates, chickens, and guinea pigs, (Dong et al., 2011b; Iuvone et al., 1989; McBrien and Gentle, 2001; Papastergiou et al., 1998; Stone et al., 1989) or negative lens defocus in chickens (Guo et al., 1995; Ohngemach et al., 1997). With partial form deprivation, retinal DOPAC is decreased locally only in the half of the retina that has been deprived (Ohngemach et al., 1997; Stone et al., 2006) and returns to normal after removal of the diffusers in chickens (Pendrak et al., 1997). Retinal DA levels after form deprivation myopia (FDM) are apparently not altered in wild-type mice (Chakraborty et al., 2015b; Park et al., 2014; Wu et al., 2015), indicating a unique emmetropization process or subtle change of DA in mice. Nonetheless, most studies from multiple species support an association between retinal DA alteration and myopia.
2.3 Effects of altered DA levels on myopic eye growth
If DA levels and/or signaling are decreased during myopic eye growth, then increasing DA level or DA receptor activity would be predicted to prevent myopia. The first step in proving this hypothesis is to increase the overall availability of DA by injecting DA directly into the eye or using L-DOPA to increase DA synthesis. Such experiments have shown that increasing DA levels prevents FDM in guinea pigs (Mao et al., 2010), rabbits (Gao et al., 2006) and mice (Landis et al., 2016). Another approach is to increase DA signaling with a non-selective DA receptor agonists, such as apomorphine (APO) and 2-amino-6,7-dihydroxy-1,2,3,4-tetrahydronaphthalene hydrobromide (ADTN). A large number of studies show that treating with APO prevents FDM in several species, including chicken (Rohrer et al., 1993; Schmid and Wildsoet, 2004; Stone et al., 1989), guinea pig (Dong et al., 2011b), monkey (Iuvone et al., 1991) and mice (Yan et al., 2015b). Similarly, ADTN inhibits FDM in chicken (Ashby et al., 2007; McCarthy et al., 2007). Notably, the inhibitory effect of APO on refraction is robust and consistent for FDM, but less consistent for negative lens-induced myopia (Dong et al., 2011b; Schmid and Wildsoet, 2004). Furthermore, treating mice with daily injections of APO inhibits FD, while continuous infusion of APO with a mini-pump produced no effect (Yan et al., 2015b). This may be due to differences in drug concentrations and their affinities for the different DA receptors, or downregulation of DA receptors resulting from continuous occupation with minipump infusion. Additionally, APO and exogenous L-DOPA have no effect on refractive development when visual input is normal (Dong et al., 2011a; Mao et al., 2010; Rohrer et al., 1993; Yan et al., 2015a). Together these studies suggest that DA receptor activation is needed for normal refractive eye growth under challenging/abnormal visual conditions (FD or lens defocus) and that increasing DA levels in the eye can prevent myopic growth signals.
If DA receptor activation is needed for normal visually-driven eye growth, then decreased levels of DA would be predicted to cause myopia in eyes with normal visual input and potentially increase the susceptibility to FD or lens defocus. Pharmacologically, DA can be reduced using 6-hydroxydopamine (6-OHDA) which is toxic to dopaminergic neurons or reserpine which depletes catecholamines from sympathetic nerve endings. However, pharmacological depletion of DA has produced less consistent results, likely due to the complex actions of these pharmacological actions. Contradictory to the hypothesis, treatment with 6-OHDA and reserpine prevents (rather than facilitates) development of FDM in chickens by reducing axial eye growth (Li et al., 1992; Schaeffel et al., 1995). Interestingly, 6-OHDA has no effect in lens-induced myopia in chickens (Schaeffel et al., 1994) or fish (Kroger et al., 1999), suggesting potentially different roles for DA with these different types of visual disruptions (Schaeffel et al., 1995). This apparently paradoxical effect might be due to the non-selective depletion of catecholamines with 6-OHDA, including DA as well as norepinephrine (Jiang et al., 2014a). There are also examples of decreased DA due to genetic defects. For example, in albino guinea pigs with defective tyrosine metabolism (Hansson et al., 1980), serum DOPA levels are markedly reduced, which may lead to spontaneous myopia and the altered response to FDM (Jiang et al., 2011; Jiang et al., 2014b). Additionally, administration of 6-OHDA in wild-type mice results in relative myopic refractive errors under normal visual conditions and enhanced FDM response (Wu et al., 2016). A recent report shows that retinal-specific TH knockout mice show spontaneous myopia with shorter axial lengths and a normal response to FD (Bergen et al., 2016), even though retinal DA levels are unaltered with FD (Bergen et al., 2016). These results suggest that modulation of visually-driven eye growth is not simply due to the level of retinal DA.
2.4 Altering DA signaling with light stimulation
Light stimulation is known to increase the release of dopamine in a linear fashion over four log units of light intensity (Cohen et al., 2012; Proll et al., 1982). Inspired by recent epidemiological studies showing the inverse relationship between the outdoor activity and development of myopia, an area of intense interest is the protective effect of bright light on myopia development, possibly via dopamine signaling (Karouta and Ashby, 2015). Children that spend more time outside have delayed myopia onset (Guggenheim et al., 2012; Rose et al., 2008; Sherwin et al., 2012). This is due to bright light, not activity or other factors (Read et al., 2014; Rose et al., 2008). Further evidence of the protective effects of bright light have been shown in a number of experiments with animal models. Using chickens (Ashby et al., 2009; Ashby and Schaeffel, 2010; Ashby et al., 2014; Backhouse et al., 2013; Lan et al., 2014), primates (Smith et al., 2012), and tree shrews (Norton and Siegwart, 2013), bright light has been reported to decrease the myopic shift induced by FD or lens defocus (Ashby and Schaeffel, 2010). These protective effects have been attributed to increased retinal DA levels and DA receptor activation as the DA D2-like receptor antagonist, spiperone blocks the protective effects of bright light (Ashby and Schaeffel, 2010). The release of DA is regulated by light intensity and image contrast, both of which are detected by photoreceptors and have been implicated in myopia development. Thus, exposure to bright light may be an environmental intervention to control myopia development by increasing DA levels in the retina. Meanwhile, APO and D2R agonist quinpirole inhibited ocular growth induced by negative lens with transient increases in choroid thickness which is similar to the effect of daily brief periods of unrestricted vision (Nickla et al., 2010). These results indicate that DA may mediate the myopia inhibition effect of visual stimulus like bright light or unrestricted vision. The retinal DA may induce choroidal thickening and ocular growth inhibition by triggerring the release of other transmitters like nitric oxide (NO) from either the retina or choroid (Nickla et al., 2009; Nickla and Wildsoet, 2004; Sekaran et al., 2005).
Importantly, two randomized, control trials reported that extending outdoor exposure time led to significant declines from 23% to 50% in the onset of myopia incidence. However, these studies also revealed modest inhibition of myopia progression from 0.06 to 0.13 D/year in elementary school children by extending outdoor expsoure time (He et al., 2015; Wu et al., 2013). Another meta-analysis included 7 cross-sectional studies reported a 2% reduced odds of myopia per additional hour of time spent outdoors per week (Sherwin et al., 2012). Furthermore, the animal studies showing that bright light reduced myopia were done under standard laboratory conditions with non-fluctuating luminance and chromaticity that does not mimic the real-world, natural environment as the luminance in the natural environment varied considerably based on time of day and cloud cover, as does chromaticity, contributing to variable effects of outdoor exposure of light on the development of FDM in chicks (Ashby et al., 2009; Stone et al., 2016). These observations suggest that a pharmacological approach to the treatment of myopia may be more efficacious than an environmental intervention.
3. Role of D1-like and D2-like receptors in myopia development
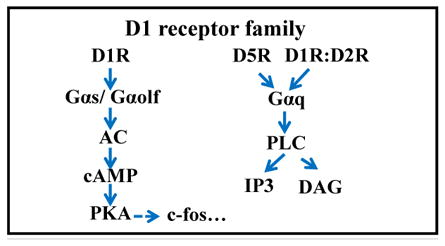
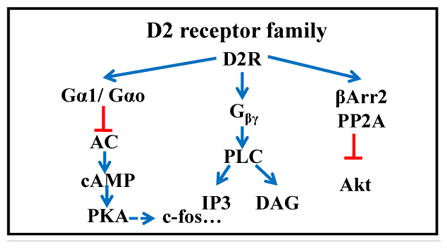
3.1 DA receptors
DA receptors are G-protein coupled receptors (Table 1). Consistent with volume transmission of DA, DA receptors are found on almost all neuronal classes within the retina. The retina expresses 4 of the 5 DA receptor subtypes: D1R, D2R, D4R, and D5R (Table 1; (Jackson et al., 2009)). D1 and D5 receptors stimulate the synthesis of the intracellular second messenger cyclic AMP, while D2 and D4 receptors inhibit its formation (Beaulieu and Gainetdinov, 2011). DA receptors have been associated with other intracellular signaling pathways as well. The next step in evaluating the involvement of DA signaling in refractive eye growth is to determine which DA receptors are involved. Using pharmacological manipulations, the influence of the activation or inhibition of specific DA receptors have been tested on refractive development under normal, FD or lens defocused conditions. One caveat of these pharmacological approaches is that they don’t target specific DA receptor subtypes, but instead they act on the families of DA receptors (D1-like and D2-like), which makes it difficult to make definitive conclusions about how DA signals visually-driven eye growth.
Table 1. Dopamine receptor biology in retina: signaling, distribution and pharmacology.
The two main families of dopamine receptors are listed with their receptor subtypes and various agonists and antagonists. RP: photoreceptor; BC: bipolar cell; HC: horizontal cell; AC: amacrine cell; RGC: retinal ganglion cell; RPE: retinal pigment epithelium.
|
|
|||||
|---|---|---|---|---|---|---|
|
|
|
|||||
| D1 | D5 | D2 | D3 | D4 | ||
|
| ||||||
| Retinal distribution | Type-specific BCs, HCs, a set of ACs, RGCs | RPE | PR Dopaminergic ACs | none | PR (mouse) | |
| Agonist Ki value (nM) | Dopamine | 0.9–2340 | <0.9–261 | 2.8–474 | 4–27 | 28–450 |
| Apomorphine | 0.7–680 | 122–168 | 0.7–24 | 20–32 | 4 | |
| ADTN | 2.9–>10000 | n/a | 1–1370 | n/a | 393 | |
| SKF-38393 | 1–150 | 0.5–100 | 150-9560 | 5000 | 1000–1300 | |
| PD-168077 | >10000 | n/a | 2820-3740 | 2810 | 8.7–25 | |
| Antagonist Ki value (nM) | Haloperidol | 27–203 | 33-151 | 0.6–1.2 | 2.74-7.8 | 2.3–5.1 |
| Spiperone | 99–350 | 135-4500 | 0.06–0.37 | 0.32–0.71 | 0.05–4 | |
| SCH-23390 | 0.11–0.35 | 0.11-0.54 | 270–1100 | 314–800 | 3000–3560 | |
| Sulpiride | 20400–45000 | 11000–77270 | 2.5–7.1 | 8–206 | 21–1000 | |
| L-745870 | n/a | n/a | 960 | 2300 | 0.43 | |
| Methylergonovine | n/a | n/a | n/a | n/a | n/a | |
3.2 Effects of D2-like receptor agonists and antagonists on myopia
Of the studies reported in the literature, most have focused on: (1) examining the effects of D1-like and D2-like receptor agonists on development of myopia (e.g., (Stone et al., 1989)); (2) use of antagonists to manipulate the effects of a DA agonist on development of myopia (APO; (Rohrer et al., 1993; Stone et al., 1989)); or the ability of DA antagonists to prevent the effect brief periods of normal vision, which inhibit development of FDM (McCarthy et al., 2007; Nickla and Totonelly, 2011; Schmid et al., 2013). In general, these studies show that activation of D2-like receptors by subconjunctival or intravitreal agonist injection mimics the protective effect of periods of unobstructed vision on FDM (McCarthy et al., 2007; Nickla and Totonelly, 2011; Schmid et al., 2013), while intravitreal injection of D2-like antagonists inhibits the protective effect of periods of unobstructed vision on FDM (Rohrer et al., 1993; Stone et al., 1989). Furthermore, co-administration of APO and a D2R antagonist abolishes the inhibitory effect of APO on myopia (Rohrer et al., 1993). It is interesting to note that D2R antagonists are not sufficient to induce myopia (Huang et al., 2014) but they facilitate myopia development induced by FDM (Schaeffel et al., 1995; Stone et al., 1989). On the other hand, systemic injection of D2 antagonists produced different effects than intravitreal injection of the drugs, indicating possible systemic effects or different DA concentrations that may contribute to this discrepancy (Huang et al., 2014). Further supporting this possibility that DA concentrations are important, D2R agonists have been found to inhibit FDM at low doses and enhance FDM at high doses (Zhou, unpublished data; Table 2). Also, like systemic injection of D2R antagonists, global genetic deletion of D2Rs in mice inhibits FDM development (Huang et al., 2014), indicating a possible developmental compensation effect of D2Rs (Table 2). It will be important to determine in future studies if D2R knockout mice have altered DA levels and/or altered spatial frequency resolution that would influence their response to FD.
Table 2.
Modulation of myopia development by genetic and pharmacological manipulation of dopamine receptor signaling: focus on mouse models.
| Normal Eye | FDM | Reference | |
|---|---|---|---|
| D1R KO | Not available | Not available | |
| D2R KO | No effects | Inhibition | Huang et al., 2014 |
| APO (daily injection) | No effects | Inhibition | Yan et al., 2015 |
| APO (continuous infusion) | No effects | No effects | Yan et al., 2015 |
| D1R agonist (SKF-38393) | No effects | Inhibition | Zhou et al., ARVO 2014 |
| D1R antagonist (SCH-23390) | No effects | Enhancement | Zhou et al., ARVO, 2014 |
| D2R agonist (Quinpirole) | No effects | Inhibiting (low doses), Enhancement (high doses) | Zhou, unpublished data |
| D2R antagonist (sulpiride) | No effects | Inhibition | Huang et al., 2014 |
| D4R agonist (PD-168077) | No effects | Enhancement | Zhou, unpublished data |
| D4R antagonist (L-745870) | Hyperopic trend | Inhibition | Zhou, unpublished data |
| 6-OHDA | Myopic shift | Enhancement | Wu et al., 2016 |
D4R is a D2-like receptor with a highly enriched expression pattern in retina. D4R agonists systemically injected into C57BL/6 mice for 4 weeks had no effect on normal refractive development, but increased the response to form deprivation (Zhou, unpublished data). In contrast, D4R antagonists produced a hyperopic trend in C57BL/6 mice under normal visual conditions and inhibited FDM (Zhou, unpublished data). In future work, it is important to determine whether the D4R recapitulate/contribute to the D2R-like drug effects.
3.3 Contribution of D1-like receptors on myopia
The role of D1R in development of myopia is less clear. Effects of D1-like antagonists have been contradictory. Some studies show D1-like antagonists to either have no effect on the protective effects of normal vision (McCarthy et al., 2007; Nickla and Totonelly, 2011) or on the protective effects of APO on FDM (Rohrer et al., 1993). Other studies have shown D1-like antagonists to abolish the ameliorating effects of periods of unobstructed vision on lens-induced myopia (Nickla et al., 2010) and to block the development of FDM in chickens (Schaeffel et al., 1995). Preliminary studies in C57BL/6 mice indicate that D1R-like agonists inhibit FDM while D1R-like antagonists enhance FDM (Zhou et al., 2014). Further studies are needed to address these apparent paradoxical protective effects of D1R agonists and antagonists and determine the exact role of D1R signaling cascades in control of visual growth and myopia development.
3.4 Working hypothesis for DA receptor contributions on myopia development
Collectively, these results suggest that D2-like (D2 and D4) receptors may play a larger role in DA signaling control of refractive eye growth than D1-like receptors in chicks. Based on the results from mouse models, a working hypothesis of homeostatic control of myopia has been proposed in which activation of D1-like receptors leads to hyperopia and activation of D2-like receptors leads to myopia (Figure 1).
Figure 1. A working hypothesis: Homeostatic control of myopia by opposing effects of D1-like and D2-like receptors in mouse eye.

Based on genetic and pharmacological results from mouse models of myopia, we postulate that D1-like and D2-like receptor activation in distinct cell types exert homeostatic control of the emmetropization process. The balance of D1-like and D2-like receptor activation in the eye modulates refractive eye development, such that over activation of D1-like receptors leads to hyperopia while over activation of D2-like receptors leads to myopia.
Dopamine is involved in both FDM and LIM, as both suppress the metabolism and release of DA in chicken (Guo et al., 1995; Papastergiou et al., 1998; Stone et al., 1989), and the axial elongation in FDM and LIM can both be blocked by DA agonists (Feldkaemper and Schaeffel, 2013). However, the DA drugs have also been shown to modulate refractive eye growth differently depending on the type of myopia that is being induced. For example, In guinea pigs, FD (but not negative len wearing) decreased DA, DOPAC and DOPAC/DA ratios with myopia development (Dong et al., 2011b). This difference may be attributed to the different mechanism or to the unmatched blurriness or the temporal course between FD and negative defocus. Pharmacological studies further support differential mechanisms between FDM and LIM: APO inhibits FD, but not hyperopic defocus in guinea pigs (Dong et al., 2011b). 6-OHDA blocks FDM in chickens, but has no effect on LIM (Schaeffel et al., 1995). To further support this notion, Nickla & Totonelly (Nickla and Totonelly, 2011) have shown that D2-like antagonists inhibit the protective effects of brief periods of unrestricted vision on FDM, but not on negative lens-induced myopia. These results suggest that DA may have a greater contribution to inhibition of myopia induced by FD versus lens defocus.
Notably, some discrepancies on the DA pharmacological effects on myopia have been reported. For examples, both DA activation (the non-specific DA receptor agonist apomorphine) and DA depletion with 6-OHDA and reserpine) have been shown to suppress myopia development in chickens (Stone, R. A et al., 1989; Schaeffel et al. 1995; Diether and Schaeffel, 1999). Also, D1 receptor agonists can have no effect in chickens or suppressive effects in guinea pigs ( McCarthy, C. S et al., 2007;Nickla et al, 2010; Jiang et al 2012;). Similarly, D2 receptor antagonists can reverse apomorphine-induced suppression of myopia (Cottrial et al, 2001; Rohrer et al, 1993) or have no effect on myopia development (Arumugam and McBrien, 2010; McCarthy et al., 2007, Cottriall et al., 2001). These mixed results may reflect the complexity and intrinsic limitation of DA pharmacology. However, even if difference exist in the mechanisms by which DA inhibits myopia, the fact that DA impacts refractive development in both mammalian and non-mammalian species emphasizes the evolutionary conservation of this neuromodulator’s role in refractive development.
In addition to the species differences, there are several possible explanations for these inconsistencies: including i) the difficulty in assessing the exact drug concentrations in retina and sclera after intra-vitreal injection versus systemic administration of dopamine agonists and antagonists; ii) lack of the comparative studies on the cellular distribution of D1 and D2 receptor in retina (versus sclera) among various species during ontogeny and myopia development. iii) based in part on recent finding from mouse model of myopia, we suggest that differential activation of dopamine receptors in distinct vision pathways (despite similar expression levels) and neural circuits (Chen et al., 2017), may also contribute to different functional outcomes. Coupled with cellular biomarkers and molecular approaches, insights from mouse genetic manipulation would provide complementary and fresh view for some of the long-held views on myopia development.
4. DA signaling and visual pathway interactions in myopia
4.1 Signaling of dopaminergic neurons in the retina
While DA levels and DA receptors play key roles in modulating visual function and refractive development, the action site of dopamine signaling in retina to control myopia is largely undefined. DA signaling is regulated by visual input and several retinal circuits provide input into the dopaminergic amacrine cells and interplexiform cells that synthesize and release DA (Dowling and Ehinger, 1978a, b; Frederick et al., 1982). These retinal dopaminergic amacrines have cell bodies at the border of the inner nuclear layer (INL) and inner plexiform layer (IPL), and long, ramifying dendrites and axons in sublamina 1 of the IPL. DA neurons in species like fish and new world monkeys have processes that ascend through the INL to the outer plexiform layer; hence the term ‘interplexiform cell’ (Ehinger and Falck, 1969). Processes of dopaminergic amacrines in the IPL intercalate with dendrites of intrinsically photosensitive retinal ganglion cells (ipRGCs) (Prigge et al., 2016b) and form rings around AII amacrine cells (Debertin et al., 2015). The retinal DA neurons receive excitatory input via ectopic synapses with ON bipolar cells in the OFF sublamina of the IPL (Dumitrescu et al., 2009a). They also receive inhibitory input from GABAergic and glycinergic amacrine cells (Qiao et al., 2016). Dopamine signaling may interact with the visual pathways by two possible mechanisms to modulate visually-driven eye growth: a) dopamine may act on the DA receptors in specific visual pathways and influence the pathways’ functions or b) visual pathway activity may influence myopia development by altering DA release and signaling. By acting on DA receptors expressed in different visual pathways, dopamine may modulate ON pathways (Farshi et al., 2016; Smith et al., 2015), OFF pathways (Maguire and Werblin, 1994; Yang et al., 2013), rod pathways (Herrmann et al., 2011) and retinal gap junctions (Bu et al., 2014; Kothmann et al., 2009; Zhang et al., 2011a), contributing to control of myopia development (Chakraborty et al., 2014; Chakraborty et al., 2015c; Park et al., 2014). Alternatively, visual inputs such as bright light and flickering light, stimulate ON pathway and ipRGC and alter dopamine synthesis and release (Contini et al., 2010; Dumitrescu et al., 2009b; Prigge et al., 2016a).
4.2 Primary or secondary effects of DA pharmacological treatments on myopia
The rate-limiting enzyme for DA synthesis, TH, is detected in dopaminergic amacrine cells, but D1-like and D2-like DA receptors are widely distributed at various cell types (including photoreceptors and retinal pigment epithelium cells for D2-like receptors, and bipolar, horizontal, amacrine and ganglion cells for D1-like receptor) (Witkovsky, 2004). It is not clear which visual pathways are specifically affected by DA pharmacological treatment. Consequently, it is unknown if the pharmacological treatments described above directly alter refractive eye growth mechanisms or if they produce secondary effects by altering visual function and/or retinal pathways that are important for myopic eye growth. Recent evidence shows that DA receptors are involved in specific aspects of visual function: D1Rs are important for spatial frequency thresholds and D4Rs are specific to contrast sensitivity thresholds and light adaptation (Jackson et al., 2012). Future studies are critically needed to define the specific visual input whereby DA signaling exerts its actions to control myopia development. For instance, experiments using retina- and cell-type-specific knockout mice or models with over-expression of D1Rs and D2Rs are needed to verify whether the effect of DA is local or systemic, and to define the cellular basis of DA action.
4.3 Retinal pathway defects alter myopia development and DA levels
It has been well-established that photic activation of retinal neurons and/or retinal circuits leads to changes in retinal dopamine release. Activation of rods, cones and ipRGCs excite dopaminergic amacrine cells, which in turn increases retinal dopamine content and turnover (Qiao et al., 2016; Zhang et al., 2008). DA is also released in response to ON bipolar cell stimulation (Boatright et al., 1994; Boelen et al., 1998; Dumitrescu et al., 2009b). Genetic-targeting of these retinal pathways in mouse models results in changes in DA and DOPAC levels, as well as refractive development and the response to FD. For instance, mice with ON pathway mutations have reduced retinal DA and DOPAC levels, developed greater myopic refractions in normal visual environment, and became more susceptible to FDM (Chakraborty et al., 2015b; Pardue et al., 2008). The exaggerated response to FDM may be due to the lower DA and DOPAC or the loss of ON pathway stimulation of another signaling pathway necessary for the response to FD. In addition, mice with mutations that cause photoreceptor degeneration have reduced DA and DOPAC and normal to hyperopic refractions during normal development with increased susceptibility to FD (Park et al., 2013). In contrast, mice with inactive rod photoreceptors also have reduced retinal DOPAC levels and DOPAC/DA ratios, but they do not have a normal pattern of refractive development across age compared to wild-type controls and do not respond to FD (Park et al., 2014). ipRGCs are a subclass of retinal ganglion cells (RGCs) that function as classical RGCs with input from the photoreceptor-driven bipolar cells, or by directly responding to light (Schmidt and Kofuji, 2011). ipRGCs control the pupillary light response and suprachiasmatic nucleus (SCN), where they entrain the biological circadian clock (Schmidt and Kofuji, 2011). In addition, ipRGCs influence retinal processes (Barnard et al., 2006) through retrograde signaling, in part through dopamine signaling (Prigge et al., 2016a; Zhang et al., 2008); but see (Cameron et al., 2009). Recent evidence also suggests that ipRGCs are involved in refractive development and FDM. Mice with targeted disruption of the Opn4 gene, which encodes melanopsin (the photopigment protein in ipRGCs), have altered refractive development and increased susceptibility to FD, implicating the involvement of ipRCSs in development of myopia (Chakraborty et al., 2015a). These findings indicate that visual stimulation through multiple pathways may alter DA signaling and DA receptors to modify refractive development. In fact, a comparison of the DA turnover in several different mouse models with retinal pathway defects shows that lower DOPAC/DA ratios at the age of FD induction results in greater susceptibility to FDM (Figure 2; (Chakraborty et al., 2015b)). These results suggest that reducing DA levels through altered visual signaling may not result in spontaneous myopia, but can alter the susceptibility to myopia. This raises an important question: do children at risk for myopia development have low DA turnover that doesn’t alter retinal function, but affects refractive development?
Figure 2. Retinal pathway defects alter dopamine signaling and susceptibility to FDM.

Several mouse models were tested that had mutations targeting retinal neurons or pathways. Comparing the myopic shift after two weeks of FD with the DOPAC/DA ratio, an indication of DA turnover, showed a strong correlation (R2 = 0.92, p<0.001). These results suggest that the level of DA turnover at the start of FD induction may alter the susceptibility to myopia, with lower DA activity predicting greater myopic shifts. (Modified from (Chakraborty and Pardue, 2015)).
5. DA signaling and myopia: therapeutic strategies and clinical implications
Given the critical need for effective and safe pharmacological treatments to prevent myopia progression, one of the key questions is whether there is sufficient preclinical and clinical data on the efficacy and safety of DA-based strategies for prevention and treatment of myopia. As described above, increasing preclinical data supports the ability of DA agonists (particularly APO and D2R agonists) to inhibit FDM. It is important to note DA agonists (such as APO) can induce selective inhibition of FDM without affecting normal visual growth in mice (Yan et al., 2015b). Additionally, the effects of L-DOPA on amblyopic vision in three clinical trials were reported (Leguire et al., 1992; Leguire et al., 1993a; Leguire et al., 1993b). Some early studies (Leguire et al., 1993a; Leguire et al., 1993b), but not others (Leguire et al., 1998), showed that L-DOPA can enhance the effect of dominant eye occlusion in treatment of amblyopia. However, a recent study with well-designed controls by the Pediatric Eye Disease Investigator Group supported by the National Eye Institute found no enhancement by L-DOPA of the effect of dominant eye occlusion therapy (Pediatric Eye Disease Investigator et al., 2015). Additional studies with L-DOPA in large numbers of children with myopia are required to demonstrate its effectiveness for prevention and treatment of myopia.
5.1 Potential negative side effects of dopaminergic therapies
From the perspective of developing DA based therapeutic strategies, what are the potential toxicological ramifications of treating children with dopaminergic agents? DA is the major neurotransmitter in the brain and retina and is also an important physiological regulator of cardiovascular, endocrine and immunological functions. In the developing brain, over-activation of the DA system has been shown to produce schizophrenia-like behavioral changes (Angrist, 1994). Consistent with these preclinical studies, some mild side effects were noted in a L-DOPA clinical trial in a small number of myopic children, including nausea, headache, fatigue, mood changes, emesis, dizziness, dry mouth, decreased appetite, and night mares; the use of carbidopa, a peripheral dopa decarboxylase inhibitor, in combination with L-DOPA reduced the side effects (Angrist, 1994; Repka et al., 2010). Moreover, in developing children treated with stimulants for attention deficit disorder (ADD)/attention deficit hyperactivity disorders (ADHD) that work by increasing extracellular dopamine levels in the brain (including Ritalin, Adderall, and Dexedrine), common side effects include feeling restless and jittery, difficulty sleeping, loss of appetite, headaches, upset stomach, irritability, mood swings, depression, dizziness, racing heartbeat, and tics (Jeffers et al., 2013; Salardini et al., 2016). The American Heart Association recommends that all individuals, including children, have a cardiac evaluation prior to starting a stimulant (Thomas et al., 2011). Moreover, the long-term impact of ADD/ADHD medication on the youthful, developing brain is not yet known. These studies indicate that more research and potentially novel pharmacological targeting is needed to avoid the negative side effects of systemic activation of DA signaling.
5.2 DA receptor agonists as a treatment option for myopia
A DA receptor agonist may provide a therapeutic option with less side effects. DA receptor agonists are currently FDA-approved to treat various diseases, including Parkinson’s disease, restless leg syndrome and diabetes mellitus. However, the use of DA agonists during critical periods of development may be detrimental. DA receptors have profound effects on glucose metabolism (Lopez Vicchi et al., 2016). Recent studies show that the dopamine D2R agonist bromocriptine, and the D1R agonist SKF38393 produce improvements in obesity, hyperglycemia (improved glucose metabolism) and hyperinsulinemia in obese (ob/ob) mice (Cincotta et al., 1997).eye growth may be associated with the energy consumption and vitreal glucose level (Feldkaemper et al 2000, IOVS ). D2R knockout mice develop features of Parkinson disease (Tinsley et al., 2009), prolactinoma (Cristina et al., 2006), chronic pituitary hyperplasia (Cristina et al., 2006) and a moderate decrease in MSH content (Kelly et al., 1997). Thus, extensive studies on possible side effects of DA on visual and systemic functions are necessary before DA agonists could be considered as an anti-myopic drug in children.
5.3 Local delivery of DA therapies to the eye
From a therapeutic perspective, local administration of APO and DA by eye drops may achieve therapeutic effects on myopia, with reduced systemic side effects. Using animal models, we need to study the effect of local application of DA agonists (with a wide range of doses) on refractive errors and retinal functions. Furthermore, given the wide-spread effects of DA in eye, brain and throughout the body, it is not sufficient to demonstrate only an anti-myopia effect of DA treatment. We also need to determine any potential effects on growth (endocrinology), psychiatric behaviors, blood pressure, etc. To our knowledge, only one study has examined the effects of APO eye drops; this study showed that local application reduced the development of deprivation-induced myopia in infant rhesus monkeys (Iuvone et al., 1991). While only a single dose was tested, the approach showed promise and there were no indications of systemic toxicity, or alterations in weight gain, general health, or development. Further studies are also needed to identify myopia-specific DA signaling for therapeutic intervention by characterizing DA signaling systems during FDM (receptor numbers, affinity, DA autoreceptor, sensitivity to DA drugs, cell-types, interaction with other neuromodulators, and the developmental time window).
6. Challenges in understanding the role of DA signaling in myopia
DA has been proposed as a stop signal for myopia eye growth for over 25 years, and recently emerged as a likely target for myopia therapy. It is clear from many studies that DA has an anti-myopigenic effect. However, the exact mechanism of action and the potential interaction with retinal pathways has remained elusive. Several key questions in the role of DA signaling in development of myopia remain to be addressed:
6.1 Does DA signaling represent a “stop” signal for control of axial eye growth or an adaptive or compensatory response as a result of refractive changes?
A related question is whether DA signaling in the retina affects initiation versus progression of myopia development. Recent epidemiological studies indicate that outdoor activity (with increasing light exposure) affects mainly the onset of myopia but not the progression of myopia development in children, indicating a possible effect of DA signaling on initiation of myopia (Guo et al., 2013; He et al., 2015; Jin et al., 2015). Data from mouse models with retinal pathway defects that result in decreased dopamine turnover and increased myopia susceptibility would also support the hypothesis that DA release alters myopia induction (Figure 2). Evidence in support of DA as an adaptive response includes the relatively slow response of DA to changes in visual input. In chicken retina, visual deprivation did not reduce vitreal DOPAC level immediately (within 30 minute) but reduced sustained DA release during the light phase (Megaw et al., 1997). Thus, DA release may be an adaptive response to low temporal and spatial contrast. Clarification of this question with direct measurement of the extracellular DA concentration is critical to our understanding of the role of DA in myopia development.
6.2 Is visually-driven myopia a fundamentally different process from the normal (developmental) growth of eye/vision?
In the retinal-specific TH knockout mice with selective depletion of DA in the retina from birth, animals develop spontaneous myopia but do not show an altered response to FD (Bergen et al., 2016). Furthermore, APO inhibits deprivation myopia in mice, but had no effect on axial growth of eyes with normal vision (Yan et al., 2015b). Thus, myopia that arises with normal visual input may be different than that induced with FD. Myopia development with excessive growth may differ from normal developmental growth due to upregulation of DA receptor numbers or an increase in DA receptor affinity for exogenous DA. Further studies are needed to better characterize DA signaling systems during FDM to identify “myopia-specific” DA signaling for therapeutic intervention.
6.3 Does DA exert homeostatic regulation of visual growth by distinct effects of D1-like and D2-like receptors?
The homeostatic regulation of visual growth by DA receptor subtypes is not clear. Non-specific DA receptor agonists (L-DOPA and APO) produce more consistent and effective inhibition of myopia development than D1-like and D2-like DA agonists, suggesting that the interaction between DA D1-like and D2-like receptors might be important. Furthermore, bi-phasic effects have been reported for APO and the D2-like agonist quinpirole on control of myopia: high doses of APO inhibit FDM, while low doses promote FDM in guinea pigs (Jiang et al., 2014b); in contrast, quinpirole inhibits myopia at low doses and promotes myopia at high doses in C57 mice (Zhou, unpublished data); while both effects are abolished in D2R KO mice (Zhou, unpublished data). This suggests that two distinct elements of DA receptor actions may exist with opposite effects under varying DA concentrations (Figure 1). Perhaps the DA receptors located on different retinal pathways serve as an auto-control mechanism to prevent the overactivation of specific DA receptor (D2-like?) from promoting over-growth of the eye.
6.4 How does retinal pathway transmission interact with DA signaling to control myopia development?
While studies with mutant mouse models have uncovered the critical role of ON, OFF, rod photoreceptor and ipRGC pathways on modulation of myopia development (Chakraborty et al., 2015a; Chakraborty et al., 2014; Pardue et al., 2008; Park et al., 2014), it is critical to determine whether specific retinal pathways control myopic development by a direct effect on ocular growth or by altering DA signaling and indirectly modifying visually-driven eye development. Pharmacological studies have also established the role of DA drugs (mainly APO and D2R agonists) in control of myopia development. However, the retinal pathway basis (i.e. the ON, OFF and ipRGC pathways and their down-stream signaling) of DA actions are largely undefined. New methods or techniques such as cell-type specific retinal gene knockdown or chemical genetics (such as DREADD) and optogenetics, coupled with DA pharmacology, are needed to dissect out the retinal pathway mechanism of DA actions in control of myopia.
7. Tackling key challenges in myopia studies with new technologies
To unravel these key questions in the role of DA signaling in development of myopia, new technologies and experimental approaches are critically needed.
7.1 Two-photon image and caging
The application of two-photon photorelease of caged dopamine with two-photon calcium imaging in defined cells of the living retina (Newkirk et al., 2013, 2015) may allow us to evaluate the differential effects of DA in different retinal neurons, and provide novel insight into the role of DA signaling in retinal adaptation, and thereby myopia development.
7.2 Cell-specific and retina-specific knockout mice
Transgenic methods to knock out specific types of retinal neurons or visual signaling pathways have already uncovered potential links between retinal signaling and DA in myopia development (Chakraborty and Pardue, 2015). However, more studies are needed to fully elucidate the contributions of DA signaling to visually-driven eye growth and myopia in these mutant mice. In addition, using techniques like Cre-lox recombination, transgenic mice targeting DA related proteins only in the retina have been developed (Bergen et al., 2016) which avoids the toxicity and other side effects of systemic knock-out models (Morgan et al., 2015). Further mouse models using inducible cre promoters could provide a method to induce loss of key DA proteins during the critical period of refractive development without deleting the gene from early retinal and visual development.
7.3 Genetic labeling of D1R- and D2R-containing cells
Transgenic mice harboring D1 (such as BAC-Drd1a-tdTomato) and D2 (such as BAC-Drd2a-EGFP) receptors linked with fluorescence protein allows for the genetic labeling and visualization of D1R and D2R receptors in the retina. The release of DA and excitability of dopaminergic amacrine cells can be evaluated by patch clamp recordings after induction of myopia or in response to different myopia-related visual stimuli. By coupling with c-Fos immunohistochemistry, this approach provides an unique resource to identify the activation of specific subpopulations of retinal cells containing D1Rs or D2Rs in response to light exposure (see Figure 3).
Figure 3. Identification of light-triggered D1R activation in specific subpopulations of D1R-containing cells in the retina using the BAC-Drd1a-tdTomato transgenic mice.

The BAC-Drd1a-tdTomato transgenic mice (genetic structure shown in A, available from the Jackson Laboratory, Bar Harbor, USA) coupled with c-fos immunohistochemistry allows for the identification of specific retinal subpopulations of D1R-containing cells in response to normal and bright light exposure. Bright light significantly increased c-fos expression in specific subpopulations of D1R-containing cells including horizontal and bipolar cells in retina. B left: the c-fos expression in D1R + retinal neurons in normal light (100–200 lux); B right: the c-fos expression in bright light (2500–5000 lux). White arrows indicate the D1R+ neurons; green arrows indicated c-fos expression; yellow arrows indicated the activated D1R+ neurons (marked by c-fos).
7.4 Optogenetics and DREADD
The recent development of optogenetics and DREAD (Designer Receptor Exclusively Activated by Designer Drugs) (Armbruster et al., 2007; Urban and Roth, 2015) offers unparalleled spatiotemporal resolution by allowing the manipulation of defined neural circuits in a physiologically relevant (milliseconds in optogenetics to second to minutes in DREADD) timescale in a reversible manner (Boyden et al., 2005; Cardin et al., 2010; Yizhar et al., 2011; Zhang et al., 2007). By selectively introducing adeno-associated virus (AAV) targeted expression of light-gated ion channels or pumps (Zhang et al., 2011b) or modified cholinergic hM3Dq (M3, excitatory) or hM4Di (M4, inhibitory) receptors into genetically defined populations of neurons (such as specific retinal neurons), light stimulation (optogenetics) or injection of clozapine-N-oxide (CNO) can selectively stimulate or inhibit cells in defined cell types and visual circuits. Using optogenetics or DREADD, we can further establish the critical role of specific retinal cells and retinal pathways (as identified in retinal pathway knockout studies) in controlling normal visually-driven eye growth and modulating myopic development.
8. Future Directions
The cause of the growing prevalence of myopia is unclear, but it now affects nearly a quarter of the world population (Holden et al., 2016). If current trends continue, it is estimated that by 2050 myopia will affect ~50% of the world population, 4.76 billion people, with almost a billion people suffering from high myopia (Holden et al., 2016). It is a critical time to develop effective therapy (such as DA-based strategies) for managing and preventing myopia-related complications and vision loss. It is also an exciting time in myopia research as new insights into the mechanisms of visually-driven eye growth and myopia may be revealed with use of new mammalian models of myopia that can be genetically manipulated, the creation of several new genetic tools to target DA related proteins (such as cell-specific and retina-specific DA receptor knockouts mice), and the recent development of optogenetic and chemical genetic approaches to manipulate defined retinal pathways in intact animals. To develop effective DA-based pharmacological treatments to prevent progressive myopia, future studies are critically needed to address key mechanistic questions that will deepen our biological understanding of the role of DA signaling in myopia development. Furthermore, as myopia is a complex disorder involving genetic and environmental factors, future studies are also warranted to address the role of DA signaling in genetic (myopia-related genes and locus) and environmental (light) interactions in myopia devellopment. Importantly, further studies on possible side effects of DA on visual and systemic functions are essential to clarify the possible unwanted side effects of DA agonists as an anti-myopic drug in children. Lastly, future studies to better characterize DA signaling and pharmacology in humans, particularly in developing children, are essential to translate basic knowledge on DA signaling and myopia into effective pharmacological treatments for myopia prevention in children.
Article highlights.
Retinal DA, released in response to light, is a stop signal for homeostatic control of myopic eye growth
DA exerts homeostatic control of eye growth by distinct actions of DA receptors
DA signaling modulates specific retinal pathways to modify visual functions
Studies on the unwanted effects of DA agents on visual and systemic functions are needed
Acknowledgments
Research in the authors’ laboratories is supported in part by the National Natural Science Foundation of China 81470659, 81271039(JQ), 81371047, 81422007, 81670886 (XZ); National Institutes of Health R01 EY016435 (MTP), R01 EY004864 and P30 EY006360 (PMI), R01 EY022342 (PMI/MTP); the Department of Veterans Affairs Research Career Scientist Award (MTP), Research to Prevent Blindness (Emory Eye Center).
Footnotes
Percentage of work contributed by each author in the production of the manuscript is as follows: XZ: 30%, MTP: 25%, PMI: 15%, JQ: 30%.
References
- Angrist B. Amphetamine and its analogues. Academic; San Diego: 1994. Amphetamine psychosis: clinical variations of the syndrome; pp. 387–414. [Google Scholar]
- Armbruster BN, Li X, Pausch MH, Herlitze S, Roth BL. Evolving the lock to fit the key to create a family of G protein-coupled receptors potently activated by an inert ligand. Proc Natl Acad Sci U S A. 2007;104:5163–5168. doi: 10.1073/pnas.0700293104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ashby R, McCarthy CS, Maleszka R, Megaw P, Morgan IG. A muscarinic cholinergic antagonist and a dopamine agonist rapidly increase ZENK mRNA expression in the form-deprived chicken retina. Exp Eye Res. 2007;85:15–22. doi: 10.1016/j.exer.2007.02.019. [DOI] [PubMed] [Google Scholar]
- Ashby R, Ohlendorf A, Schaeffel F. The effect of ambient illuminance on the development of deprivation myopia in chicks. Invest Ophthalmol Vis Sci. 2009;50:5348–5354. doi: 10.1167/iovs.09-3419. [DOI] [PubMed] [Google Scholar]
- Ashby RS, Schaeffel F. The effect of bright light on lens compensation in chicks. Invest Ophthalmol Vis Sci. 2010;51:5247–5253. doi: 10.1167/iovs.09-4689. [DOI] [PubMed] [Google Scholar]
- Ashby RS, Zeng G, Leotta AJ, Tse DY, McFadden SA. Egr-1 mRNA expression is a marker for the direction of mammalian ocular growth. Invest Ophthalmol Vis Sci. 2014;55:5911–5921. doi: 10.1167/iovs.13-11708. [DOI] [PubMed] [Google Scholar]
- Backhouse S, Collins AV, Phillips JR. Influence of periodic vs continuous daily bright light exposure on development of experimental myopia in the chick. Ophthalmic Physiol Opt. 2013;33:563–572. doi: 10.1111/opo.12069. [DOI] [PubMed] [Google Scholar]
- Barnard AR, Hattar S, Hankins MW, Lucas RJ. Melanopsin regulates visual processing in the mouse retina. Current biology : CB. 2006;16:389–395. doi: 10.1016/j.cub.2005.12.045. [DOI] [PubMed] [Google Scholar]
- Beaulieu JM, Gainetdinov RR. The physiology, signaling, and pharmacology of dopamine receptors. Pharmacol Rev. 2011;63:182–217. doi: 10.1124/pr.110.002642. [DOI] [PubMed] [Google Scholar]
- Bergen MA, Park HN, Chakraborty R, Landis EG, Sidhu C, He L, Iuvone PM, Pardue MT. Altered Refractive Development in Mice With Reduced Levels of Retinal Dopamine. Invest Ophthalmol Vis Sci. 2016;57:4412–4419. doi: 10.1167/iovs.15-17784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bjelke B, Goldstein M, Tinner B, Andersson C, Sesack SR, Steinbusch HW, Lew JY, He X, Watson S, Tengroth B, Fuxe K. Dopaminergic transmission in the rat retina: evidence for volume transmission. J Chem Neuroanat. 1996;12:37–50. doi: 10.1016/s0891-0618(96)00176-7. [DOI] [PubMed] [Google Scholar]
- Boatright JH, Rubim NM, Iuvone PM. Regulation of endogenous dopamine release in amphibian retina by melatonin: the role of GABA. Vis Neurosci. 1994;11:1013–1018. doi: 10.1017/s0952523800003941. [DOI] [PubMed] [Google Scholar]
- Boelen MK, Boelen MG, Marshak DW. Light-stimulated release of dopamine from the primate retina is blocked by 1–2-amino-4-phosphonobutyric acid (APB) Vis Neurosci. 1998;15:97–103. doi: 10.1017/s0952523898151040. [DOI] [PubMed] [Google Scholar]
- Boyden ES, Zhang F, Bamberg E, Nagel G, Deisseroth K. Millisecond-timescale, genetically targeted optical control of neural activity. Nature neuroscience. 2005;8:1263–1268. doi: 10.1038/nn1525. [DOI] [PubMed] [Google Scholar]
- Bu JY, Li H, Gong HQ, Liang PJ, Zhang PM. Gap junction permeability modulated by dopamine exerts effects on spatial and temporal correlation of retinal ganglion cells’ firing activities. Journal of computational neuroscience. 2014;36:67–79. doi: 10.1007/s10827-013-0469-1. [DOI] [PubMed] [Google Scholar]
- Cameron MA, Pozdeyev N, Vugler AA, Cooper H, Iuvone PM, Lucas RJ. Light regulation of retinal dopamine that is independent of melanopsin phototransduction. Eur J Neurosci. 2009;29:761–767. doi: 10.1111/j.1460-9568.2009.06631.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cardin JA, Carlen M, Meletis K, Knoblich U, Zhang F, Deisseroth K, Tsai LH, Moore CI. Targeted optogenetic stimulation and recording of neurons in vivo using cell-type-specific expression of Channelrhodopsin-2. Nature protocols. 2010;5:247–254. doi: 10.1038/nprot.2009.228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chakraborty R, Lee DC, Landis ER, Bergen MA, Park HN, Sidhu C, Hattar S, Iuvone PM, Stone RA, Pardue MT. Melanopsin knock-out mice have abnormal refractive development and increased susceptibility to form-deprivation myopia. Invest Ophthalmol Vis Sci. 2015a;56 E-Abstract 5843. [Google Scholar]
- Chakraborty R, Pardue MT. Molecular and Biochemical Aspects of the Retina on Refraction. Progress in molecular biology and translational science. 2015;134:249–267. doi: 10.1016/bs.pmbts.2015.06.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chakraborty R, Park H, Aung MH, Tan CC, Sidhu CS, Iuvone PM, Pardue MT. Comparison of refractive development and retinal dopamine in OFF pathway mutant and C57BL/6J wild-type mice. Mol Vis. 2014;20:1318–1327. [PMC free article] [PubMed] [Google Scholar]
- Chakraborty R, Park HN, Hanif AM, Sidhu CS, Iuvone PM, Pardue MT. ON pathway mutations increase susceptibility to form-deprivation myopia. Exp Eye Res. 2015b;137:79–83. doi: 10.1016/j.exer.2015.06.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chakraborty R, Park HN, Hanif AM, Sidhu CS, Iuvone PM, Pardue MT. ON pathway mutations increase susceptibility to form-deprivation myopia. Exp Eye Res. 2015c;137:79–83. doi: 10.1016/j.exer.2015.06.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen S, Zhi Z, Ruan Q, Liu Q, Li F, Wan F, Reinach PS, Chen J, Qu J, Zhou X. Bright Light Suppresses Form-Deprivation Myopia Development With Activation of Dopamine D1 Receptor Signaling in the ON Pathway in Retina. Invest Ophthalmol Vis Sci. 2017;58:2306–2316. doi: 10.1167/iovs.16-20402. [DOI] [PubMed] [Google Scholar]
- Cincotta AH, Tozzo E, Scislowski PW. Bromocriptine/SKF38393 treatment ameliorates obesity and associated metabolic dysfunctions in obese (ob/ob) mice. Life sciences. 1997;61:951–956. doi: 10.1016/s0024-3205(97)00599-7. [DOI] [PubMed] [Google Scholar]
- Cohen J, Hadjiconstantinou M, Neff NH. Activation of dopamine-containing amacrine cells of retina: light-induced increase of acidic dopamine metabolites. Brain Res. 1983;260:125–127. doi: 10.1016/0006-8993(83)90771-0. [DOI] [PubMed] [Google Scholar]
- Cohen Y, Belkin M, Yehezkel O, Solomon AS, Polat U. Dependency between light intensity and refractive development under light-dark cycles. Exp Eye Res. 2011;92:40–46. doi: 10.1016/j.exer.2010.10.012. [DOI] [PubMed] [Google Scholar]
- Cohen Y, Peleg E, Belkin M, Polat U, Solomon AS. Ambient illuminance, retinal dopamine release and refractive development in chicks. Exp Eye Res. 2012;103:33–40. doi: 10.1016/j.exer.2012.08.004. [DOI] [PubMed] [Google Scholar]
- Contini M, Lin B, Kobayashi K, Okano H, Masland RH, Raviola E. Synaptic input of ON-bipolar cells onto the dopaminergic neurons of the mouse retina. J Comp Neurol. 2010;518:2035–2050. doi: 10.1002/cne.22320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cristina C, Garcia-Tornadu I, Diaz-Torga G, Rubinstein M, Low MJ, Becu-Villalobos D. Dopaminergic D2 receptor knockout mouse: an animal model of prolactinoma. Frontiers of hormone research. 2006;35:50–63. doi: 10.1159/000094308. [DOI] [PubMed] [Google Scholar]
- Debertin G, Kantor O, Kovacs-Oller T, Balogh L, Szabo-Meleg E, Orban J, Nyitrai M, Volgyi B. Tyrosine hydroxylase positive perisomatic rings are formed around various amacrine cell types in the mammalian retina. J Neurochem. 2015;134:416–428. doi: 10.1111/jnc.13144. [DOI] [PubMed] [Google Scholar]
- Dolgin E. The myopia boom. Nature. 2015;519:276–278. doi: 10.1038/519276a. [DOI] [PubMed] [Google Scholar]
- Dong F, Zhi Z, Pan M, Xie R, Qin X, Lu R, Mao X, Chen JF, Willcox MD, Qu J, Zhou X. Inhibition of experimental myopia by a dopamine agonist: different effectiveness between form deprivation and hyperopic defocus in guinea pigs. Mol Vis. 2011a;17:2824–2834. [PMC free article] [PubMed] [Google Scholar]
- Dong F, Zhi Z, Pan M, Xie R, Qin X, Lu R, Mao X, Chen JF, Willcox MD, Qu J, Zhou X. Inhibition of experimental myopia by a dopamine agonist: different effectiveness between form deprivation and hyperopic defocus in guinea pigs. Mol Vis. 2011b;17:2824–2834. [PMC free article] [PubMed] [Google Scholar]
- Dowling JE, Ehinger B. The interplexiform cell system. I. Synapses of the dopaminergic neurons of the goldfish retina. Proceedings of the Royal Society of London Series B, Biological sciences. 1978a;201:7–26. doi: 10.1098/rspb.1978.0030. [DOI] [PubMed] [Google Scholar]
- Dowling JE, Ehinger B. Synaptic organization of the dopaminergic neurons in the rabbit retina. J Comp Neurol. 1978b;180:203–220. doi: 10.1002/cne.901800202. [DOI] [PubMed] [Google Scholar]
- Dumitrescu ON, Pucci FG, Wong KY, Berson DM. Ectopic Retinal ON Bipolar Cell Synapses in the OFF Inner Plexiform Layer: Contacts with Dopaminergic Amacrine Cells and Melanopsin Ganglion Cells. J Comp Neurol. 2009a;517:226–244. doi: 10.1002/cne.22158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dumitrescu ON, Pucci FG, Wong KY, Berson DM. Ectopic retinal ON bipolar cell synapses in the OFF inner plexiform layer: contacts with dopaminergic amacrine cells and melanopsin ganglion cells. J Comp Neurol. 2009b;517:226–244. doi: 10.1002/cne.22158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ehinger B, Falck B. Adrenergic retinal neurons of some new world monkeys. Z Zellforsch Mikrosk Anat. 1969;100:364–375. doi: 10.1007/BF00571492. [DOI] [PubMed] [Google Scholar]
- Farshi P, Fyk-Kolodziej B, Krolewski DM, Walker PD, Ichinose T. Dopamine D1 receptor expression is bipolar cell type-specific in the mouse retina. J Comp Neurol. 2016;524:2059–2079. doi: 10.1002/cne.23932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feldkaemper M, Schaeffel F. An updated view on the role of dopamine in myopia. Exp Eye Res. 2013;114:106–119. doi: 10.1016/j.exer.2013.02.007. [DOI] [PubMed] [Google Scholar]
- Frederick JM, Rayborn ME, Laties AM, Lam DM, Hollyfield JG. Dopaminergic neurons in the human retina. J Comp Neurol. 1982;210:65–79. doi: 10.1002/cne.902100108. [DOI] [PubMed] [Google Scholar]
- Gao Q, Liu Q, Ma P, Zhong X, Wu J, Ge J. Effects of direct intravitreal dopamine injections on the development of lid-suture induced myopia in rabbits. Graefes Arch Clin Exp Ophthalmol. 2006;244:1329–1335. doi: 10.1007/s00417-006-0254-1. [DOI] [PubMed] [Google Scholar]
- Guggenheim JA, Northstone K, McMahon G, Ness AR, Deere K, Mattocks C, Pourcain BS, Williams C. Time outdoors and physical activity as predictors of incident myopia in childhood: a prospective cohort study. Invest Ophthalmol Vis Sci. 2012;53:2856–2865. doi: 10.1167/iovs.11-9091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guo SS, Sivak JG, Callender MG, Diehl-Jones B. Retinal dopamine and lens-induced refractive errors in chicks. Curr Eye Res. 1995;14:385–389. doi: 10.3109/02713689508999936. [DOI] [PubMed] [Google Scholar]
- Guo Y, Liu LJ, Xu L, Lv YY, Tang P, Feng Y, Meng M, Jonas JB. Outdoor activity and myopia among primary students in rural and urban regions of Beijing. Ophthalmology. 2013;120:277–283. doi: 10.1016/j.ophtha.2012.07.086. [DOI] [PubMed] [Google Scholar]
- Hansson C, Agrup G, Rorsman H, Rosengren E. Dopa and 5-S-cysteinyldopa in the serum of albino, black, and red guinea pigs. Acta dermato-venereologica. 1980;60:155–156. [PubMed] [Google Scholar]
- He M, Xiang F, Zeng Y, Mai J, Chen Q, Zhang J, Smith W, Rose K, Morgan IG. Effect of Time Spent Outdoors at School on the Development of Myopia Among Children in China: A Randomized Clinical Trial. Jama. 2015;314:1142–1148. doi: 10.1001/jama.2015.10803. [DOI] [PubMed] [Google Scholar]
- Herrmann R, Heflin SJ, Hammond T, Lee B, Wang J, Gainetdinov RR, Caron MG, Eggers ED, Frishman LJ, McCall MA, Arshavsky VY. Rod vision is controlled by dopamine-dependent sensitization of rod bipolar cells by GABA. Neuron. 2011;72:101–110. doi: 10.1016/j.neuron.2011.07.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holden BA, Fricke TR, Wilson DA, Jong M, Naidoo KS, Sankaridurg P, Wong TY, Naduvilath TJ, Resnikoff S. Global Prevalence of Myopia and High Myopia and Temporal Trends from 2000 through 2050. Ophthalmology. 2016;123:1036–1042. doi: 10.1016/j.ophtha.2016.01.006. [DOI] [PubMed] [Google Scholar]
- Huang F, Yan T, Shi F, An J, Xie R, Zheng F, Li Y, Chen J, Qu J, Zhou X. Activation of dopamine d2 receptor is critical for the development of form-deprivation myopia in the C57BL/6 mouse. Invest Ophthalmol Vis Sci. 2014;55:5537–5544. doi: 10.1167/iovs.13-13211. [DOI] [PubMed] [Google Scholar]
- Iuvone PM, Galli CL, Garrison-Gund CK, Neff NH. Light stimulates tyrosine hydroxylase activity and dopamine synthesis in retinal amacrine neurons. Science (New York, N Y. 1978;202:901–902. doi: 10.1126/science.30997. [DOI] [PubMed] [Google Scholar]
- Iuvone PM, Tigges M, Fernandes A, Tigges J. Dopamine synthesis and metabolism in rhesus monkey retina: development, aging, and the effects of monocular visual deprivation. Vis Neurosci. 1989;2:465–471. doi: 10.1017/s0952523800012360. [DOI] [PubMed] [Google Scholar]
- Iuvone PM, Tigges M, Stone RA, Lambert S, Laties AM. Effects of apomorphine, a dopamine receptor agonist, on ocular refraction and axial elongation in a primate model of myopia. Invest Ophthalmol Vis Sci. 1991;32:1674–1677. [PubMed] [Google Scholar]
- Jackson CR, Chaurasia SS, Zhou H, Haque R, Storm DR, Iuvone PM. Essential roles of dopamine D4 receptors and the type 1 adenylyl cyclase in photic control of cyclic AMP in photoreceptor cells. J Neurochem. 2009;109:148–157. doi: 10.1111/j.1471-4159.2009.05920.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jackson CR, Ruan GX, Aseem F, Abey J, Gamble K, Stanwood G, Palmiter RD, Iuvone PM, McMahon DG. Retinal dopamine mediates multiple dimensions of light-adapted vision. J Neurosci. 2012;32:9359–9368. doi: 10.1523/JNEUROSCI.0711-12.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jeffers A, Benotsch EG, Koester S. Misuse of prescription stimulants for weight loss, psychosocial variables, and eating disordered behaviors. Appetite. 2013;65:8–13. doi: 10.1016/j.appet.2013.01.008. [DOI] [PubMed] [Google Scholar]
- Jiang BP, Le L, Xu LJ, Xiao PG. Minocycline inhibits ICAD degradation and the NF-kappaB activation induced by 6-OHDA in PC12 cells. Brain Res. 2014a;1586:1–11. doi: 10.1016/j.brainres.2014.08.001. [DOI] [PubMed] [Google Scholar]
- Jiang L, Long K, Schaeffel F, Zhang S, Zhou X, Lu F, Qu J. Disruption of emmetropization and high susceptibility to deprivation myopia in albino guinea pigs. Invest Ophthalmol Vis Sci. 2011;52:6124–6132. doi: 10.1167/iovs.10-7088. [DOI] [PubMed] [Google Scholar]
- Jiang L, Long K, Schaeffel F, Zhou X, Zheng Y, Ying H, Lu F, Stell WK, Qu J. Effects of dopaminergic agents on progression of naturally occurring myopia in albino guinea pigs (Cavia porcellus) Invest Ophthalmol Vis Sci. 2014b;55:7508–7519. doi: 10.1167/iovs.14-14294. [DOI] [PubMed] [Google Scholar]
- Jin JX, Hua WJ, Jiang X, Wu XY, Yang JW, Gao GP, Fang Y, Pei CL, Wang S, Zhang JZ, Tao LM, Tao FB. Effect of outdoor activity on myopia onset and progression in school-aged children in northeast China: the Sujiatun Eye Care Study. BMC ophthalmology. 2015;15:73–77. doi: 10.1186/s12886-015-0052-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones LA, Sinnott LT, Mutti DO, Mitchell GL, Moeschberger ML, Zadnik K. Parental history of myopia, sports and outdoor activities, and future myopia. Invest Ophthalmol Vis Sci. 2007;48:3524–3532. doi: 10.1167/iovs.06-1118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karouta C, Ashby RS. Correlation Between Light Levels and the Development of Deprivation Myopia. Investigative ophthalmology & visual science. 2015;56:299–309. doi: 10.1167/iovs.14-15499. [DOI] [PubMed] [Google Scholar]
- Kelly MA, Rubinstein M, Asa SL, Zhang G, Saez C, Bunzow JR, Allen RG, Hnasko R, Ben-Jonathan N, Grandy DK, Low MJ. Pituitary lactotroph hyperplasia and chronic hyperprolactinemia in dopamine D2 receptor-deficient mice. Neuron. 1997;19:103–113. doi: 10.1016/s0896-6273(00)80351-7. [DOI] [PubMed] [Google Scholar]
- Kothmann WW, Massey SC, O’Brien J. Dopamine-stimulated dephosphorylation of connexin 36 mediates AII amacrine cell uncoupling. J Neurosci. 2009;29:14903–14911. doi: 10.1523/JNEUROSCI.3436-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kramer SG. Dopamine: A retinal neurotransmitter. I. Retinal uptake, storage, and light-stimulated release of H3-dopamine in vivo. Invest Ophthalmol. 1971;10:438–452. [PubMed] [Google Scholar]
- Kroger RH, Hirt B, Wagner HJ. Effects of retinal dopamine depletion on the growth of the fish eye. Journal of comparative physiology. 1999;184:403–412. doi: 10.1007/s003590050339. [DOI] [PubMed] [Google Scholar]
- Lan W, Feldkaemper M, Schaeffel F. Intermittent episodes of bright light suppress myopia in the chicken more than continuous bright light. PLoS One. 2014;9:e110906. doi: 10.1371/journal.pone.0110906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Landis E, Park H, Chakraborty R, Sidhu C, Iuvone PM, Pardue MT. Ascorbic acid, and not L-DOPA, protects against form-deprivation myopia in retinal degeneration mouse models. Invest Ophthalmol Vis Sci. 2016;57 E-Absract No pagination specified. [Google Scholar]
- Leguire LE, Rogers GL, Bremer DL, Walson P, Hadjiconstantinou-Neff M. Levodopa and childhood amblyopia. Journal of pediatric ophthalmology and strabismus. 1992;29:290–298. doi: 10.3928/0191-3913-19920901-08. discussion 299. [DOI] [PubMed] [Google Scholar]
- Leguire LE, Rogers GL, Bremer DL, Walson PD, McGregor ML. Levodopa/carbidopa for childhood amblyopia. Invest Ophthalmol Vis Sci. 1993a;34:3090–3095. [PubMed] [Google Scholar]
- Leguire LE, Rogers GL, Walson PD, Bremer DL, McGregor ML. Occlusion and levodopa-carbidopa treatment for childhood amblyopia. Journal of AAPOS : the official publication of the American Association for Pediatric Ophthalmology and Strabismus/American Association for Pediatric Ophthalmology and Strabismus. 1998;2:257–264. doi: 10.1016/s1091-8531(98)90080-5. [DOI] [PubMed] [Google Scholar]
- Leguire LE, Walson PD, Rogers GL, Bremer DL, McGregor ML. Longitudinal study of levodopa/carbidopa for childhood amblyopia. Journal of pediatric ophthalmology and strabismus. 1993b;30:354–360. doi: 10.3928/0191-3913-19931101-04. [DOI] [PubMed] [Google Scholar]
- Li W, Lan W, Yang S, Liao Y, Xu Q, Lin L, Yang Z. The effect of spectral property and intensity of light on natural refractive development and compensation to negative lenses in guinea pigs. Invest Ophthalmol Vis Sci. 2014;55:6324–6332. doi: 10.1167/iovs.13-13802. [DOI] [PubMed] [Google Scholar]
- Li XX, Schaeffel F, Kohler K, Zrenner E. Dose-dependent effects of 6-hydroxy dopamine on deprivation myopia, electroretinograms, and dopaminergic amacrine cells in chickens. Vis Neurosci. 1992;9:483–492. doi: 10.1017/s0952523800011287. [DOI] [PubMed] [Google Scholar]
- Lopez Vicchi F, Luque GM, Brie B, Nogueira JP, Garcia Tornadu I, Becu-Villalobos D. Dopaminergic drugs in type 2 diabetes and glucose homeostasis. Pharmacological research. 2016;109:74–80. doi: 10.1016/j.phrs.2015.12.029. [DOI] [PubMed] [Google Scholar]
- Maguire G, Werblin F. Dopamine enhances a glutamate-gated ionic current in OFF bipolar cells of the tiger salamander retina. J Neurosci. 1994;14:6094–6101. doi: 10.1523/JNEUROSCI.14-10-06094.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mao J, Liu S, Qin W, Li F, Wu X, Tan Q. Levodopa inhibits the development of form-deprivation myopia in guinea pigs. Optom Vis Sci. 2010;87:53–60. doi: 10.1097/OPX.0b013e3181c12b3d. [DOI] [PubMed] [Google Scholar]
- McBrien NA, Gentle A. The role of visual information in the control of scleral matrix biology in myopia. Curr Eye Res. 2001;23:313–319. doi: 10.1076/ceyr.23.5.313.5440. [DOI] [PubMed] [Google Scholar]
- McCarthy CS, Megaw P, Devadas M, Morgan IG. Dopaminergic agents affect the ability of brief periods of normal vision to prevent form-deprivation myopia. Exp Eye Res. 2007;84:100–107. doi: 10.1016/j.exer.2006.09.018. [DOI] [PubMed] [Google Scholar]
- Megaw P, Morgan I, Boelen M. Vitreal dihydroxyphenylacetic acid (DOPAC) as an index of retinal dopamine release. J Neurochem. 2001;76:1636–1644. doi: 10.1046/j.1471-4159.2001.00145.x. [DOI] [PubMed] [Google Scholar]
- Megaw PL, Morgan IG, Boelen MK. Dopaminergic behaviour in chicken retina and the effect of form deprivation. Australian and New Zealand journal of ophthalmology. 1997;25(Suppl 1):S76–78. doi: 10.1111/j.1442-9071.1997.tb01764.x. [DOI] [PubMed] [Google Scholar]
- Meiser J, Weindl D, Hiller K. Complexity of dopamine metabolism. Cell communication and signaling : CCS. 2013;11:34. doi: 10.1186/1478-811X-11-34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mitsuma T, Rhue H, Hirooka Y, Kayama M, Wago T, Takagi J, Adachi K, Ping J, Ohtake M, Nogimori T, Sakai J. Distribution of Dopamine Transporter in the Rat: an Immunohistochemical Study. Endocrine regulations. 1998;32:71–75. [PubMed] [Google Scholar]
- Morgan IG, Ohno-Matsui K, Saw SM. Myopia. Lancet. 2012;379:1739–1748. doi: 10.1016/S0140-6736(12)60272-4. [DOI] [PubMed] [Google Scholar]
- Morgan RG, Gibbs JT, Melief EJ, Postupna NO, Sherfield EE, Wilson A, Keene CD, Montine TJ, Palmiter RD, Darvas M. Relative contributions of severe dopaminergic neuron ablation and dopamine depletion to cognitive impairment. Experimental neurology. 2015;271:205–214. doi: 10.1016/j.expneurol.2015.06.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Newkirk GS, Hoon M, Wong RO, Detwiler PB. Inhibitory inputs tune the light response properties of dopaminergic amacrine cells in mouse retina. Journal of neurophysiology. 2013;110:536–552. doi: 10.1152/jn.00118.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Newkirk GS, Hoon M, Wong RO, Detwiler PB. Response Properties of a Newly Identified Tristratified Narrow Field Amacrine Cell in the Mouse Retina. PLoS One. 2015;10:e0137702. doi: 10.1371/journal.pone.0137702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nickla DL, Damyanova P, Lytle G. Inhibiting the neuronal isoform of nitric oxide synthase has similar effects on the compensatory choroidal and axial responses to myopic defocus in chicks as does the non-specific inhibitor L-NAME. Exp Eye Res. 2009;88:1092–1099. doi: 10.1016/j.exer.2009.01.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nickla DL, Totonelly K. Dopamine antagonists and brief vision distinguish lens-induced- and form-deprivation-induced myopia. Exp Eye Res. 2011;93:782–785. doi: 10.1016/j.exer.2011.08.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nickla DL, Totonelly K, Dhillon B. Dopaminergic agonists that result in ocular growth inhibition also elicit transient increases in choroidal thickness in chicks. Exp Eye Res. 2010;91:715–720. doi: 10.1016/j.exer.2010.08.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nickla DL, Wildsoet CF. The effect of the nonspecific nitric oxide synthase inhibitor NG-nitro-L-arginine methyl ester on the choroidal compensatory response to myopic defocus in chickens. Optom Vis Sci. 2004;81:111–118. doi: 10.1097/00006324-200402000-00009. [DOI] [PubMed] [Google Scholar]
- Norton TT, Siegwart JT., Jr Light levels, refractive development, and myopia--a speculative review. Exp Eye Res. 2013;114:48–57. doi: 10.1016/j.exer.2013.05.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ohngemach S, Hagel G, Schaeffel F. Concentrations of biogenic amines in fundal layers in chickens with normal visual experience, deprivation, and after reserpine application. Vis Neurosci. 1997;14:493–505. doi: 10.1017/s0952523800012153. [DOI] [PubMed] [Google Scholar]
- Papastergiou GI, Schmid GF, Laties AM, Pendrak K, Lin T, Stone RA. Induction of axial eye elongation and myopic refractive shift in one-year-old chickens. Vision Res. 1998;38:1883–1888. doi: 10.1016/s0042-6989(97)00347-7. [DOI] [PubMed] [Google Scholar]
- Pardue MT, Faulkner AE, Fernandes A, Yin H, Schaeffel F, Williams RW, Pozdeyev N, Iuvone PM. High susceptibility to experimental myopia in a mouse model with a retinal on pathway defect. Invest Ophthalmol Vis Sci. 2008;49:706–712. doi: 10.1167/iovs.07-0643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park H, Jabbar SB, Tan CC, Sidhu CS, Abey J, Aseem F, Schmid G, Iuvone PM, Pardue MT. Visually-driven ocular growth in mice requires functional rod photoreceptors. Invest Ophthalmol Vis Sci. 2014;55:6272–6279. doi: 10.1167/iovs.14-14648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park H, Tan CC, Faulkner A, Jabbar SB, Schmid G, Abey J, Iuvone PM, Pardue MT. Retinal degeneration increases susceptibility to myopia in mice. Mol Vis. 2013;19:2068–2079. [PMC free article] [PubMed] [Google Scholar]
- Pediatric Eye Disease Investigator G. Repka MX, Kraker RT, Dean TW, Beck RW, Siatkowski RM, Holmes JM, Beauchamp CL, Golden RP, Miller AM, Verderber LC, Wallace DK. A randomized trial of levodopa as treatment for residual amblyopia in older children. Ophthalmology. 2015;122:874–881. doi: 10.1016/j.ophtha.2015.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pendrak K, Nguyen T, Lin T, Capehart C, Zhu X, Stone RA. Retinal dopamine in the recovery from experimental myopia. Curr Eye Res. 1997;16:152–157. doi: 10.1076/ceyr.16.2.152.5090. [DOI] [PubMed] [Google Scholar]
- Prigge CL, Yeh PT, Liou NF, Lee CC, You SF, Liu LL, McNeill DS, Chew KS, Hattar S, Chen SK, Zhang DQ. M1 ipRGCs Influence Visual Function through Retrograde Signaling in the Retina. J Neurosci. 2016a;36:7184–7197. doi: 10.1523/JNEUROSCI.3500-15.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prigge CL, Yeh PT, Liou NF, Lee CC, You SF, Liu LL, McNeill DS, Chew KS, Hattar S, Chen SK, Zhang DQ. M1 ipRGCs Influence Visual Function through Retrograde Signaling in the Retina. Journal of Neuroscience. 2016b;36:7184–7197. doi: 10.1523/JNEUROSCI.3500-15.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Proll MA, Kamp CW, Morgan WW. Use of liquid chromatography with electrochemistry to measure effects of varying intensities of white light on DOPA accumulation in rat retinas. Life sciences. 1982;30:11–19. doi: 10.1016/0024-3205(82)90630-0. [DOI] [PubMed] [Google Scholar]
- Qiao SN, Zhang ZJ, Ribelayga CP, Zhong YM, Zhang DQ. Multiple cone pathways are involved in photic regulation of retinal dopamine. Scientific reports. 2016:6. doi: 10.1038/srep28916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Read SA, Collins MJ, Vincent SJ. Light exposure and physical activity in myopic and emmetropic children. Optom Vis Sci. 2014;91:330–341. doi: 10.1097/OPX.0000000000000160. [DOI] [PubMed] [Google Scholar]
- Repka MX, Kraker RT, Beck RW, Atkinson CS, Bacal DA, Bremer DL, Davis PL, Gearinger MD, Glaser SR, Hoover DL, Laby DM, Morrison DG, Rogers DL, Sala NA, Suh DW, Wheeler MB Pediatric Eye Disease Investigator G. Pilot study of levodopa dose as treatment for residual amblyopia in children aged 8 years to younger than 18 years. Archives of ophthalmology. 2010;128:1215–1217. doi: 10.1001/archophthalmol.2010.178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rohrer B, Spira AW, Stell WK. Apomorphine blocks form-deprivation myopia in chickens by a dopamine D2-receptor mechanism acting in retina or pigmented epithelium. Vis Neurosci. 1993;10:447–453. doi: 10.1017/s0952523800004673. [DOI] [PubMed] [Google Scholar]
- Rose KA, Morgan IG, Ip J, Kifley A, Huynh S, Smith W, Mitchell P. Outdoor activity reduces the prevalence of myopia in children. Ophthalmology. 2008;115:1279–1285. doi: 10.1016/j.ophtha.2007.12.019. [DOI] [PubMed] [Google Scholar]
- Salardini E, Zeinoddini A, Kohi A, Mohammadi MR, Mohammadinejad P, Khiabany M, Shahriari M, Akhondzadeh S. Agomelatine as a Treatment for Attention-Deficit/Hyperactivity Disorder in Children and Adolescents: A Double-Blind, Randomized Clinical Trial. J Child Adol Psychop. 2016;26:513–519. doi: 10.1089/cap.2016.0024. [DOI] [PubMed] [Google Scholar]
- Schaeffel F, Bartmann M, Hagel G, Zrenner E. Studies on the role of the retinal dopamine/melatonin system in experimental refractive errors in chickens. Vision Res. 1995;35:1247–1264. doi: 10.1016/0042-6989(94)00221-7. [DOI] [PubMed] [Google Scholar]
- Schaeffel F, Hagel G, Bartmann M, Kohler K, Zrenner E. 6-Hydroxy dopamine does not affect lens-induced refractive errors but suppresses deprivation myopia. Vision Res. 1994;34:143–149. doi: 10.1016/0042-6989(94)90327-1. [DOI] [PubMed] [Google Scholar]
- Schmid KL, Strasberg G, Rayner CL, Hartfield PJ. The effects and interactions of GABAergic and dopaminergic agents in the prevention of form deprivation myopia by brief periods of normal vision. Exp Eye Res. 2013;110:88–95. doi: 10.1016/j.exer.2013.02.017. [DOI] [PubMed] [Google Scholar]
- Schmid KL, Wildsoet CF. Inhibitory effects of apomorphine and atropine and their combination on myopia in chicks. Optom Vis Sci. 2004;81:137–147. doi: 10.1097/00006324-200402000-00012. [DOI] [PubMed] [Google Scholar]
- Schmidt TM, Kofuji P. An isolated retinal preparation to record light response from genetically labeled retinal ganglion cells. Journal of visualized experiments : JoVE. 2011 doi: 10.3791/2367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sekaran S, Cunningham J, Neal MJ, Hartell NA, Djamgoz MB. Nitric oxide release is induced by dopamine during illumination of the carp retina: serial neurochemical control of light adaptation. Eur J Neurosci. 2005;21:2199–2208. doi: 10.1111/j.1460-9568.2005.04051.x. [DOI] [PubMed] [Google Scholar]
- Sherwin JC, Reacher MH, Keogh RH, Khawaja AP, Mackey DA, Foster PJ. The association between time spent outdoors and myopia in children and adolescents: a systematic review and meta-analysis. Ophthalmology. 2012;119:2141–2151. doi: 10.1016/j.ophtha.2012.04.020. [DOI] [PubMed] [Google Scholar]
- Smith BJ, Cote PD, Tremblay F. D1 dopamine receptors modulate cone ON bipolar cell Nav channels to control daily rhythms in photopic vision. Chronobiology international. 2015;32:48–58. doi: 10.3109/07420528.2014.951054. [DOI] [PubMed] [Google Scholar]
- Smith EL, 3rd, Hung LF, Huang J. Protective effects of high ambient lighting on the development of form-deprivation myopia in rhesus monkeys. Invest Ophthalmol Vis Sci. 2012;53:421–428. doi: 10.1167/iovs.11-8652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stone RA, Cohen Y, McGlinn AM, Davison S, Casavant S, Shaffer J, Khurana TS, Pardue MT, Iuvone PM. Development of Experimental Myopia in Chicks in a Natural Environment. Invest Ophthalmol Vis Sci. 2016;57:4779–4789. doi: 10.1167/iovs.16-19310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stone RA, Lin T, Laties AM, Iuvone PM. Retinal dopamine and form-deprivation myopia. Proc Natl Acad Sci U S A. 1989;86:704–706. doi: 10.1073/pnas.86.2.704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stone RA, Pendrak K, Sugimoto R, Lin T, Gill AS, Capehart C, Liu J. Local patterns of image degradation differentially affect refraction and eye shape in chick. Curr Eye Res. 2006;31:91–105. doi: 10.1080/02713680500479517. [DOI] [PubMed] [Google Scholar]
- Thomas PE, Carlo WF, Decker JA, Cannon BC, Kertesz NJ, Friedman RA, Kim JJ. Impact of the American Heart Association Scientific Statement on Screening Electrocardiograms and Stimulant Medications. Arch Pediat Adol Med. 2011;165:166–170. doi: 10.1001/archpediatrics.2010.278. [DOI] [PubMed] [Google Scholar]
- Tinsley RB, Bye CR, Parish CL, Tziotis-Vais A, George S, Culvenor JG, Li QX, Masters CL, Finkelstein DI, Horne MK. Dopamine D2 receptor knockout mice develop features of Parkinson disease. Annals of neurology. 2009;66:472–484. doi: 10.1002/ana.21716. [DOI] [PubMed] [Google Scholar]
- Urban DJ, Roth BL. DREADDs (designer receptors exclusively activated by designer drugs): chemogenetic tools with therapeutic utility. Annu Rev Pharmacol Toxicol. 2015;55:399–417. doi: 10.1146/annurev-pharmtox-010814-124803. [DOI] [PubMed] [Google Scholar]
- Wang Y, Ding H, Stell WK, Liu L, Li S, Liu H, Zhong X. Exposure to sunlight reduces the risk of myopia in rhesus monkeys. PLoS One. 2015;10:e0127863. doi: 10.1371/journal.pone.0127863. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Witkovsky P. Dopamine and retinal function. Doc Ophthalmol. 2004;108:17–40. doi: 10.1023/b:doop.0000019487.88486.0a. [DOI] [PubMed] [Google Scholar]
- Wu PC, Tsai CL, Wu HL, Yang YH, Kuo HK. Outdoor activity during class recess reduces myopia onset and progression in school children. Ophthalmology. 2013;120:1080–1085. doi: 10.1016/j.ophtha.2012.11.009. [DOI] [PubMed] [Google Scholar]
- Wu XH, Li YY, Zhang PP, Qian KW, Ding JH, Hu G, Weng SJ, Yang XL, Zhong YM. Unaltered retinal dopamine levels in a C57BL/6 mouse model of form-deprivation myopia. Invest Ophthalmol Vis Sci. 2015;56:967–977. doi: 10.1167/iovs.13-13362. [DOI] [PubMed] [Google Scholar]
- Wu XH, Qian KW, Xu GZ, Li YY, Ma YY, Huang F, Wang YQ, Zhou X, Qu J, Yang XL, Zhong YM, Weng SJ. The Role of Retinal Dopamine in C57BL/6 Mouse Refractive Development as Revealed by Intravitreal Administration of 6-Hydroxydopamine. Invest Ophthalmol Vis Sci. 2016;57:5393–5404. doi: 10.1167/iovs.16-19543. [DOI] [PubMed] [Google Scholar]
- Yan T, Xiong W, Huang F, Zheng F, Ying H, Chen J, Qu J, Zhou X. Daily Injection but not Continuous Infusion of Apomorphine Inhibits Form-Deprivation Myopia in Mice. Invest Ophthalmol Vis Sci. 2015a doi: 10.1167/iovs.13-12361. [DOI] [PubMed] [Google Scholar]
- Yan T, Xiong W, Huang F, Zheng F, Ying H, Chen JF, Qu J, Zhou X. Daily Injection But Not Continuous Infusion of Apomorphine Inhibits Form-Deprivation Myopia in Mice. Invest Ophthalmol Vis Sci. 2015b;56:2475–2485. doi: 10.1167/iovs.13-12361. [DOI] [PubMed] [Google Scholar]
- Yang J, Pahng J, Wang GY. Dopamine modulates the off pathway in light-adapted mouse retina. Journal of neuroscience research. 2013;91:138–150. doi: 10.1002/jnr.23137. [DOI] [PubMed] [Google Scholar]
- Yizhar O, Fenno LE, Davidson TJ, Mogri M, Deisseroth K. Optogenetics in neural systems. Neuron. 2011;71:9–34. doi: 10.1016/j.neuron.2011.06.004. [DOI] [PubMed] [Google Scholar]
- Zhang AJ, Jacoby R, Wu SM. Light- and dopamine-regulated receptive field plasticity in primate horizontal cells. J Comp Neurol. 2011a;519:2125–2134. doi: 10.1002/cne.22604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang DQ, Wong KY, Sollars PJ, Berson DM, Pickard GE, McMahon DG. Intraretinal signaling by ganglion cell photoreceptors to dopaminergic amacrine neurons. Proc Natl Acad Sci U S A. 2008;105:14181–14186. doi: 10.1073/pnas.0803893105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang F, Vierock J, Yizhar O, Fenno LE, Tsunoda S, Kianianmomeni A, Prigge M, Berndt A, Cushman J, Polle J, Magnuson J, Hegemann P, Deisseroth K. The microbial opsin family of optogenetic tools. Cell. 2011b;147:1446–1457. doi: 10.1016/j.cell.2011.12.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang F, Wang LP, Brauner M, Liewald JF, Kay K, Watzke N, Wood PG, Bamberg E, Nagel G, Gottschalk A, Deisseroth K. Multimodal fast optical interrogation of neural circuitry. Nature. 2007;446:633–639. doi: 10.1038/nature05744. [DOI] [PubMed] [Google Scholar]
- Zhou X, Xiong W, Huang F, Yang J, Qu J. C57BL/6 mouse eyes treated by dopamine D1 receptor agonist and antagonist during form deprivation: an opposite effect on axial length and refractive development. Invest Ophthalmol Vis Sci. 2014:55. E-Abstract 3038. [Google Scholar]