

Abstract
It is debated whether people can actively search for more than one object or whether this results in switch costs. Using a gaze-contingent eye-tracking paradigm, we revealed a crucial role for cognitive control in multiple-target search. We instructed participants to simultaneously search for two target objects presented among distractors. In one condition, both targets were available, which gave the observer free choice of what to search for and allowed for proactive control. In the other condition, only one of the two targets was available, so that the choice was imposed, and a reactive mechanism would be required. No switch costs emerged when target choice was free, but switch costs emerged reliably when targets were imposed. Bridging contradictory findings, the results are consistent with models of visual selection in which only one attentional template actively drives selection and in which the efficiency of switching targets depends on the type of cognitive control allowed for by the environment.
Keywords: visual search, visual attention, eye movements, attentional template, cognitive control, open data, open materials
Can people search for more than one object at a time? Visual search is thought to be guided by a representation of the current search target, activated through visual working memory (VWM; e.g., Carlisle, Arita, Pardo, & Woodman, 2011; Desimone & Duncan, 1995; Olivers & Eimer, 2011). Despite VWM being able to maintain multiple items (Cowan, 2001), performance is often impaired when observers try to search for multiple objects simultaneously (e.g., Houtkamp & Roelfsema, 2009; Menneer, Cave, & Donnelly, 2009; Stroud, Menneer, Cave, Donnelly, & Rayner, 2011; Wolfe, 2012; Wolfe, Butcher, Lee, & Hyle, 2003). Olivers, Peters, Houtkamp, and Roelfsema (2011) proposed a multiple-state model of VWM (after Oberauer, 2002), in which being stored in VWM is in itself not sufficient for an item to guide attention. Rather, the capacity to actively bias visual input for selection is reserved for a single target representation (the “template”). As a consequence, multiple-target search requires costly switching between templates.
Recently, however, Beck, Hollingworth, and Luck (2012) reported an eye-tracking study suggesting that attention can be biased toward multiple objects simultaneously. In the crucial condition, participants were instructed to concurrently search for two colors within arrays of multiple colored objects, as the target would always be one of those two colors. Indeed, fixations frequently alternated between items drawn in the two relevant colors. Notably, fixation dwell time was not prolonged prior to such color switches, compared with repeated selections of the same color. Thus, Beck et al. concluded that attention can be simultaneously biased toward multiple target colors.
Why does multiple-target search result in costs in some conditions but not in others? We hypothesized that an important factor is the extent to which observers can control target selection. Specifically, two forms of control appear relevant here, referred to by Braver (2012) as proactive and reactive control. Proactive control entails activation of task-relevant information in preparation for an anticipated event, thus improving performance (Geng, 2014; Locke & Braver, 2008). Conversely, reactive control occurs when task goals have to be updated in response to conflicting stimulus demands, such as those that arise during imposed unanticipated events. Notably, since reactive mechanisms occur in response to the event, reactive control results in additional processing costs. So far, little is known about the role of these control mechanisms in visual search. Interestingly, in Beck et al.’s (2012) task, observers could freely choose which template to activate, because matching objects were always available in the search display. This way, observers could proactively control selection by preparing for either target color without having to prepare a bias for both simultaneously. Since observers could start preparing for these switches at any point during the search, switch costs would not necessarily be reflected in the fixation dwell times measured during selection.
A more stringent test of whether observers can actively search for multiple objects without costs will require that observers are not able to freely choose the target category in the next selection—because the targets are being imposed. If observers really prepare multiple attention-guiding templates simultaneously, this should not matter: Any target will match a currently active template. However, if observers can prepare a perceptual bias for only a single target category at a time, unanticipated targets will require a reactive mechanism. Indeed, many studies reporting switch costs were designed so that it was unpredictable which of the target categories would be present (e.g., Houtkamp & Roelfsema, 2009; Wolfe et al., 2003). We thus hypothesized that switch costs remain latent under conditions of proactive control, while they are revealed under conditions requiring reactive mechanisms.
In the three experiments reported here, we tested whether the presence of switch costs in multiple-target search depends on the control mechanisms allowed for by the environment. We adopted the gaze-contingent design illustrated in Figure 1. Observers were first cued as to which two targets to search for. Subsequently, a series of search displays appeared, each containing either one or two targets among distractors. Observers were instructed to rapidly fixate a target, and after they correctly fixated the target, the next display was presented. Displays consisted of either colors (Experiments 1 and 3) or iconic objects (Experiment 2). The crucial factor was target availability in the displays. When both targets were available, participants could freely choose between them and thus efficiently exert proactive control by preparing a top-down bias for either target. When only one target was available, target changes were imposed, which required reactive mechanisms—unless both targets could be prepared for simultaneously. We reasoned that if observers can prepare for both targets simultaneously, the results should be the same regardless of target availability. However, if observers can prepare for only one target at a time, we expected to observe costs in the one-target condition (in which observers would often need to deal with unprepared-for targets) but not in the both-targets condition (in which there would always be a prepared-for target present).
Fig. 1.

Example sequence for a block of Experiment 1. Each block began with a cue indicating the two target colors for the subsequent sequence of search displays. Depending on the condition, each of the 40 search displays contained either one target color (hypothesized to require reactive control on a significant portion of trials) or both target colors (which allowed for efficient proactive control throughout the block). Participants were required to fixate one of the targets (indicated here by an arrow); this triggered the next display, which appeared in an imaginary circle surrounding the location of the previously fixated target.
Experiment 1
In Experiment 1, we used color displays to provide a first test of the impact of target availability on switch costs.
Method
Participants
Twelve participants (age: 19–30 years, M = 23.3; 8 females, 4 males) were recruited from the Vrije Universiteit Amsterdam and participated for course credit or money. All of them reported normal or corrected-to-normal visual acuity and color vision and were naive to the purpose of the experiment. The number of participants was determined on the basis of the research of Beck et al. (2012), as well as from pilot work. The study was approved by the ethics board of the Faculty of Behavioural and Movement Sciences at Vrije Universiteit Amsterdam.
Apparatus
The experiment was designed and presented using the OpenSesame software package (Version 2.9.1; Mathôt, Schreij, & Theeuwes, 2012) in combination with PyGaze (Version 0.4), an eye-tracking toolbox (Dalmaijer, Mathôt, & Van der Stigchel, 2014). Stimuli were presented on a 22-in. Samsung Syncmaster 2233RZ with a resolution of 1,680 × 1,050 pixels and refresh rate of 60 Hz at a view-ing distance of 75 cm. Eye movements were recorded with the EyeLink 1000 eye-tracking system (SR Research, Mississauga, Ontario, Canada) at a sampling rate of 1000 Hz and a spatial resolution of 0.01° of visual angle. The experiment took place in a dimly lit, sound-attenuated room. The experimenter received real-time feedback on system accuracy on a second monitor located in an adjacent room. After every block, eye-tracker accuracy was assessed and improved as needed by applying a 9-point calibration and validation procedure.
Stimuli, procedure, and design
The stimulus set consisted of five colored disks extending over a visual angle of 1.3°. The RGB values of these colors were as follows—blue: 0, 130, 150; red: 240, 0, 0; green: 70, 135, 0; brown: 175, 100, 75; and purple: 180, 80, 170. The background color was a uniform gray (RGB value = 197, 197, 197).
A block began with a fixation cross for 500 ms, followed by a cue for 2,500 ms and another fixation cross for 500 ms (see Fig. 1). The cue consisted of two colored disks 1.06° to the left and right of fixation and indicated the target colors for the upcoming sequence of 40 search displays. The search displays each consisted of five colored disks. Participants were instructed to make an eye movement toward a disk that matched either one of the target colors. After target fixation, the stimuli disappeared from the display, and the fixated target was replaced by a black ring to provide participants with a fixation point for the next search display, which appeared after 880 to 1,080 ms (randomly jittered). Because the coordinates of the previously fixated target served as the starting point for the next display, the search moved across the screen throughout a block, similar to natural eye movements during visual search when all items are present simultaneously.
In each search display, the items were placed on an imaginary circle around fixation with a radius randomly drawn from values between 3.8° and 4.6° around the starting point and an angular distance between each of the stimuli of at least 45°. However, if no such positions could be found (e.g., if the stimuli were located at an edge or in the corner of the screen), this restriction was loosened. Fixations had to land within a radius of 1.5 times the size of the target to be counted as valid. If participants fixated one of the distractors, they received auditory feedback and were required to make a corrective eye movement toward a target. The search was aborted if no target was fixated within 3,000 ms, and a new search display appeared.
There were two main factors. First, at the block level, target availability was manipulated. In the both-targets condition, both cued target colors appeared in the search display, while in the one-target condition, only one of those colors was present. The second factor was trial type—whether target color selection switched or repeated from one trial to the next. Note that this latter factor was determined by either the observer (both-targets condition) or by a random sampling procedure, in which a sequence of switch trials and repeat trials was randomly drawn (with replacement) from a pool of potential sequences (one-target condition). In order to match switch rate and the number of consecutive repeat trials, between those conditions, we used sequences that were obtained during both-targets blocks to constitute the pool of replay sequences for one-target blocks. Only the sequence of switch and repeat trials was replayed, not the specific colors or positions of the search items. The pool of replay sequences to draw from would grow as the experiment progressed.
Because we did not yet have any sequences to fill the pool with at the outset of the experiment, we initialized the pool with four prespecified random sequences of switch and repeat trials (one each for five, six, seven, and eight switches per block). Having a small proportion of fully random sequences also further prevented participants from recognizing the order of switches and repeats in the sequences, while still closely matching switch rates between conditions. Indeed, as a paired-samples t test confirmed, switch rates did not differ significantly between conditions, t(11) = 1.78, p = .10, Cohen’s d = 0.36. The average switch rate was 6.3 for both-targets blocks and 6.5 for one-target blocks, so that approximately 16% of all trials were switches. Finally, we also asked participants after the experiment whether they were aware of this replay manipulation in the one-target blocks, and none of them were.
Note that we did not manipulate target availability (one target vs. both targets) by simply putting one target in the one-target color displays and both targets in the both-targets color displays, because in that case the conditions would differ not only in terms of target color availability, but also in the mere number of targets in the display. Rather, the one-target condition included search displays containing two targets, but both of the same color, so that still only one target color was available. In addition, we included displays in which two distractors would have the same color, so that participants could not identify targets on the basis of color repeats. Similarly, the both-colors condition contained displays in which two out of three targets would have the same color and displays in which the distractor color was repeated. Effectively, each target-availability condition contained search displays in which either the color of one target or the color of one distractor was duplicated. This way, neither the number of targets nor the number of colors in the display was predictive of target-color availability. See Table S1 in the Supplemental Material available online for illustrations of the display types and their associated saccade latencies.
Nevertheless, to make sure that these different types of displays did not by themselves affect target selection, we initially ran a three-way repeated measures analysis of variance (ANOVA) with target availability (both targets vs. one target), trial type (switch trials vs. repeat trials), and display type (repeated target color vs. repeated distractor color) as factors on saccade latency. As display type did not have any significant influence on saccade latency (the p values for the two-way interaction with trial type, two-way interaction with target availability, and three-way interaction were all above .05), the data we present are collapsed across display types. The results of the full ANOVA including this factor can be found in the Supplemental Material.
In total, there were 40 blocks consisting of 40 search displays each. The five potential target colors were combined into 10 unique two-color cue combinations. For each target-availability condition, each of these combinations was used twice as the pair of target colors for a block. Before the experiment started, observers practiced two both-targets and two one-target blocks.
Data preprocessing
In the critical analysis, we compared the saccade latencies of eye movements (dwell time before a saccade toward a target was executed) for repeat trials (current target category the same as the previous one) with those for switch trials (current target category different from the previous one) when either one or two targets colors were available. We took the first saccade after search-display onset with an amplitude threshold of 1° of visual angle around initial fixation, provided that saccade was directed toward the selected target (i.e., it was directed more to the target than to any of the other items in the display, and its direction was no more than 30 angular degrees away from the direction of the target). This resulted in an average of 23% of trials being removed.
Next, a saccade latency filter was applied, in which saccades quicker than 100 ms and slower than 3 standard deviations above the block mean for that participant were excluded (4.4% of all search displays). Also, search displays were excluded in which the boundary regions to determine which stimulus was being fixated turned out to have a slight overlap (12.3% of all search displays). This could happen when the sequence of fixations led into a corner of the screen. If no target was being fixated, as could have happened when the eye-tracker calibration deteriorated, both the current as well as the next search display were also excluded because neither could be meaningfully labeled as a switch or repeat (2.6% of all search displays). For the same reason, we excluded the first search display of each block (2.5% of all search displays). In total, 30% of all search displays were removed during preprocessing.
Results
Figure 2 shows mean saccade latency prior to switch trials and repeat trials as a function of target availability. We found switch costs in one-target blocks but not in both-targets blocks. A two-way repeated measures ANOVA on mean saccade latency with target availability and trial type as factors revealed a significant main effect of trial type, F(1, 11) = 18.52, p = .001, η2 = .63, and target availability, F(1, 11) = 21.2, p < .001, η2 = .66, as well as a two-way interaction between target availability and trial type, F(1, 11) = 11.2, p = .006, η2 = .51. A Bayes factor (BF) analysis confirmed these results by suggesting that the full model with both main effects and the interaction effect was supported most by the data (BF = 8.12 × 104) and was 6.34 times more likely than the next best-fitting model (only main effects and no interaction effect). In both-targets blocks, saccade latencies were smaller than in one-target blocks, but did not differ significantly for switch trials (M = 234 ms) and repeat trials (M = 232 ms), t(11) = 0.25, p > .250, Cohen’s d = 0.09. In one-target blocks, however, saccade latencies for switch trials (M = 305 ms) were significantly higher than for repeat trials (M = 258 ms), t(11) = 4.16, p = .002, Cohen’s d = 1.05, which indicates that switch costs were incurred. Table S1 in the Supplemental Material shows the saccade latencies associated with each specific display type.
Fig. 2.
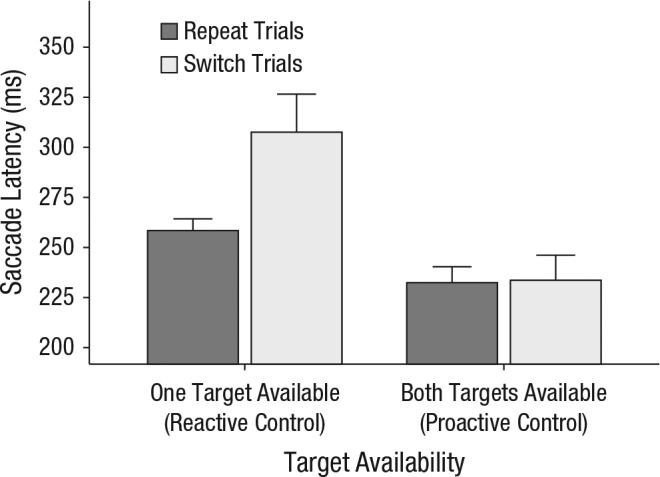
Results of Experiment 1: mean saccade latency prior to switch trials and repeat trials for each level of target availability (one target vs. both targets). For clarity, we also added the control mechanisms that apply in those conditions: Proactive control gives the observer free choice of what to search for, while reactive control limits observer choice to a single target category. Error bars represent the upper limit of within-subjects 95% confidence intervals (Morey, 2008).
Table 1 shows participants’ fixation accuracy. The pattern confirms the findings for the saccade latencies, which precludes an interpretation of the results in terms of speed/accuracy trade-offs. A repeated measures ANOVA with target availability and trial type as factors yielded significant main effects of target availability, F(1, 11) = 55.3, p < .001, η2 = .83, and trial type, F(1, 11) = 51.5, p < .001, η2 = .82, as well as a significant interaction between them, F(1, 11) = 14.2, p = .003, η2 > .56. Overall, performance was worse in the one-target than in the both-targets condition, but especially after a switch.
Table 1.
Percentage of Correctly Fixated Targets on Switch and Repeat Trials
| Experiment and number of available targets | Switch trials | Repeat trials |
|---|---|---|
| Experiment 1 | ||
| Both targets | 94.7 [93.7, 95.7] | 96.6 [96.0, 97.2] |
| One target | 85.9 [83.9, 87.9] | 94.6 [93.7, 95.5] |
| Experiment 2 | ||
| Both targets | 92.6 [92.0, 93.2] | 93.8 [92.8, 94.8] |
| One target | 86.3 [85.2, 87.4] | 89.5 [88.7, 90.3] |
| Experiment 3 | ||
| Both targets | 94.8 [93.7, 95.9] | 95.6 [94.3, 96.9] |
| One target | 75.3 [73.4, 77.2] | 84.2 [83.1, 85.3] |
Note: Within-subjects 95% confidence intervals (Morey, 2008) are given in brackets.
Discussion
We found no switch costs when both target items were available for selection. This replicates the main finding of Beck et al. (2012), who also used displays with both relevant colors present. Notably, when only one target color was available, clear switch costs emerged in both saccade latency and accuracy. This dissociation on the basis of target availability is not predicted when observers prepare a bias for both targets simultaneously. Instead, these findings suggest that observers switch between target representations and that the cost of switching is determined by the cognitive-control mechanism allowed for by the environment. When both targets are present, proactively preparing a bias toward one target is sufficient. This is not the case when there is a single, unprepared-for target, which then requires a reactive response.
In the one-target condition, observers may have coincidentally prepared for the correct color on switch trials or the wrong color on repeat trials, which would have resulted in an underestimation of switch costs. Nevertheless, the magnitude of switch costs here (40–50 ms) was less than previous estimates, which have been in the 100- to 300-ms range (e.g., Dombrowe, Donk, & Olivers, 2011; Wolfe, Horowitz, Kenner, Hyle, & Vasan, 2004). Several factors may have contributed to this difference, as these studies contained more distractors in the display, defined targets as feature conjunctions rather than single colors (Wolfe et al., 2004), or contained direct target-distractor swaps (Dombrowe et al., 2011).
Finally, we observed a main effect of target availability, with faster reaction times when both targets were present than when only one target was present. The possibility that a target would frequently not be present in the one-target search displays may have made participants more cautious throughout, which may have increased their reaction times even in repeat trials.
Experiment 2
Objects are typically defined by more features than color alone. Moreover, it may be argued that our results are confined to color, which is known to provide strong guidance of attention (Wolfe & Horowitz, 2004). Experiment 2 therefore investigated whether our findings generalize to object search. We used iconic objects consisting of conjunctions of lines and surfaces and presented them in black and white, which typically provides less guidance than color (e.g., Wolfe, 2007, 2012). Nevertheless, we did not intend guidance to be completely absent, because the task required rapid saccades to only targets.
Method
Participants
A new sample of 20 participants (age: 18–34 years, M = 23.3; 14 females, 6 males) was recruited. The number of participants was predetermined using the same criteria as in Experiment 1, but we increased the sample size because we expected less guidance and thus less clear effects with these types of stimuli.
Stimuli
Instead of colors, the stimulus set consisted of iconic images of real-world objects (see Fig. 3). These objects showed slight variations along multiple feature dimensions, such as luminance, shape, spatial frequency, and orientation, which makes detection based on any single feature or feature dimension unlikely. Objects spanned 3.4° of visual angle (156 × 156 pixels). To maximize the distance between them, we spread the five objects in the search display evenly on an imaginary circle of 360° around the participant’s target fixation on the previous trial, except when the sequence of search displays approached an edge of the screen, in which case the stimuli were spread across the 200° span that led search away from the edge toward the middle of the screen. As in Experiment 1, the background color was a uniform gray (RGB value = 197, 197, 197). Stimuli were created and the experiment was run using the same materials and apparatus as in Experiment 1.
Fig. 3.

Stimulus set for Experiment 2. The objects in the top row were potential targets (bottle, screwdriver, corkscrew, lighter, spray can); the ones in the bottom row were potential distractors (shoe, brush, sandwich, pen, toothpaste).
Procedure
We used the same procedure as in Experiment 1 with only minor modifications. To familiarize participants with the stimuli, we added five learning blocks in the beginning of the experiment, in which each of the five potential targets served once as target. Furthermore, a fixation had to land within 1.04° around the center of the target to be counted as valid. Note that because stimuli had irregular shapes, this region did not always cover the entire object. Therefore, participants were instructed to fixate the center of the objects. Finally, the intertrial interval between two consecutive search displays was decreased, now randomly jittered between 700 and 800 ms in order to keep the total duration of the experiment at around 60 min.
Data preprocessing
On the basis of the same exclusion criteria as in Experiment 1, we removed 34.4% of all search displays during preprocessing: 4.4% because no target was fixated, 4.7% because saccades on that trial were quicker than 100 ms and slower than 3 standard deviations above the block mean for that participant, 2.5% because the display was the first in a block, and 32.1% because the first saccade was not directed to the target.
Results
Figure 4 shows mean saccade latency prior to switch trials and repeat trials as a function of target availability. Overall, saccade latencies (M = 298 ms) were numerically higher than in Experiment 1 (M = 258 ms), although this difference was not significant (p = .12). They were also slightly less accurate, F(1, 30) = 2.25, p = .14, although accuracy was still well above chance. This pattern is consistent with an overall reduction in guidance relative to Experiment 1, though not with an absence of guidance.
Fig. 4.
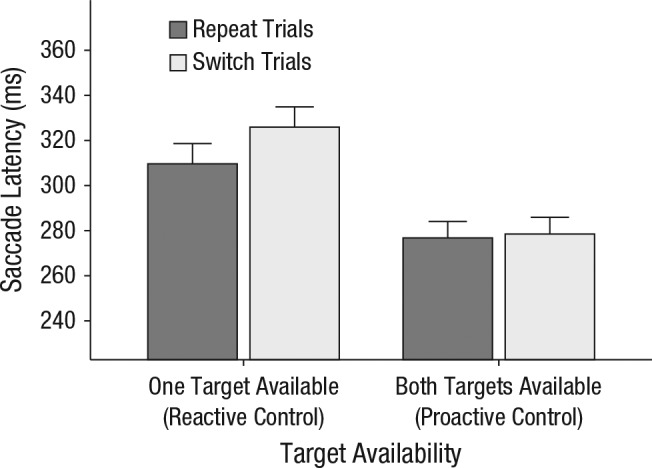
Results of Experiment 2: mean saccade latency prior to switch trials and repeat trials for each level of target availability (one target vs. both targets). For clarity, we also added the control mechanisms that apply in those conditions: Proactive control gives the observer free choice of what to search for, while reactive control limits observer choice to a single target category. Error bars represent the upper limit of within-subjects 95% confidence intervals (Morey, 2008).
We again found switch costs in one-target blocks but not in both-targets blocks. A two-way repeated measures ANOVA on mean saccade latency with target availability and trial type as factors revealed a significant main effect of trial type, F(1, 19) = 5.92, p = .025, η2 = .24, and target availability, F(1, 19) = 22.70, p < .001, η2 = .54, as well as a two-way interaction between them, F(1, 19) = 7.01, p = .016, η2 = .27. Saccade latencies were smaller in both-targets blocks than in one-target blocks. In both-targets blocks, participants switched on 34% of all displays and initiated saccades toward the target with the same latency on switch trials (M = 279 ms) and on repeat trials (M = 277 ms), t(19) = 0.99, p > .250, Cohen’s d = 0.02, whereas in one-target blocks, saccade latencies for switch trials (M = 326 ms) were significantly higher than for repeat trials (M = 310 ms), t(19) = 2.6, p = .02, Cohen’s d = 0.17, which indicates that switch costs were incurred. The pattern was not supported by a standard Bayes factor analysis: The model that contained only the main effect of target availability was supported strongest by the data (BF = 3.62 × 106), which suggests that neither the main effect of target selection nor the interaction effect had a meaningful influence.
As the Bayes factor and classical analysis contradicted each other, we ran further, more specific tests with the Bayesian approach. Following Morey and Wagenmakers (2014), we incorporated our hypothesis concerning the direction of switch costs in each of the conditions, on the basis of the direction of the interaction that was found in Experiment 1, by using order restrictions. We expected switch costs in the one-target condition but not in the both-targets condition. After adding this specific prediction, the model that included both the main effect and the interaction effect was supported most (BF = 1.02 × 107) and was 2.81 times more likely than the model including only the main effect of target availability. Even though this is still not strong evidence in favor of the interaction, it is in line with the predictions of the classical approach.
Table 1 shows participants’ fixation accuracy. The pattern is very similar to the pattern of the saccade latencies. A repeated measures ANOVA with target availability and trial type as factors yielded significant main effects of target availability, F(1, 19) = 46.9, p < .001, η2 = .71, and trial type, F(1, 19) = 16.0, p < .001, η2 = .46, and a significant interaction between them, F(1, 19) = 4.6, p = .045, η2 = .20. Overall, performance was worse in the one-target than in the both-targets condition, especially after a switch.
Discussion
Experiment 2 shows that our findings generalize to properties other than color, in this case black-and-white object icons. However, switch costs were smaller than in Experiment 1, which may be due to reduced guidance toward anticipated targets.
Experiment 3
Because the previous two experiments manipulated target availability at a block level, the results might have originated from strategic influences other than the cognitive-control mechanisms we were interested in. Experiment 3 therefore replicated the effects in a mixed design (Fig. 5). While searching for both targets, participants could proactively choose which target to fixate in the majority (80%) of search displays, because both targets were present. We expected no switch costs for these displays. On the remaining 20% of displays, only one target was present. Unless observers can simultaneously prepare a bias for both targets, unprepared-for one-target search displays should again require reactive mechanisms, which would result in switch costs.
Fig. 5.

Example sequence for a block of Experiment 3. As in Experiment 1, each block began with a cue indicating the two target colors for the subsequent sequence of 30 search displays. However, each display in Experiment 3 contained only three disks. Depending on the condition, displays contained either both target colors (80% of all trials), which allowed participants to choose freely between them, or one target color (20% of all trials), so participants had no choice, and reactive control was hypothesized to be involved on a significant portion of the trials. Only switch trials are shown in this figure.
Method
Participants
A new sample of 20 participants (age: 21–35 years, M = 26.4; 11 females, 9 males) was recruited. We kept the same sample size that was used for Experiment 2, again using a larger number of participants than in Experiment 1 because of anticipated smaller effects in a mixed compared with a block design.
Stimuli, procedure, and design
Experiment 3 used a simplified design relative to Experiment 1, as we showed that the various display types (with or without duplicate colors) had no discernable effect. The stimulus set consisted of four colored disks. Their RGB values were as follows—blue: 79, 129, 237; pink: 247, 70, 193; green: 89, 162, 1; and brown: 195, 115, 30; the background color was a uniform gray (RGB value = 197, 197, 197). Each of the 30 search displays in a block contained three colored disks. On 80% of the trials within a block, these three stimuli consisted of two targets and only a single distractor, which allowed for proactive control to be effective. In the other 20%, the display consisted of two distractors and just one target, which thus involved reactive control. Target availability was randomized per block, with the restrictions that both targets were present in each of the first five searches of a block and that between two one-target displays, there was at least one both-targets display. The color of the target in one-target search displays was set contingently on the selected target on the previous search, which imposed switch and repeat trials equally often. The target cue was shown for 3,000 ms, and the fixation screens before and after the cue lasted for 750 ms. Stimuli were created and the experiment was run using the same materials and apparatus as in Experiment 1.
Data preprocessing
On the basis of the same exclusion criteria as in the previous experiments, we removed 25.6% of all search displays during preprocessing: 4.1% because saccades on that trial were quicker than 100 ms and slower than 3 standard deviations above the block mean for that participant, 3.3% because the display was the first in a block, 0.1% because the boundary regions to determine which stimulus was being fixated slightly overlapped, and 20.7% because the first saccade was not directed to the target.
Results
Figure 6 shows mean saccade latency as a function of target availability and trial type. Switch costs did not occur when both target colors were available but did emerge when only one target color was available. A two-way repeated measures ANOVA on saccade latency with target availability and trial type as factors yielded significant main effects of target availability, F(1, 19) = 17.7, p < .001, η2 = .48, and trial type, F(1, 19) = 13.6, p = .002, η2 = .42, as well as an interaction between them, F(1, 19) = 14.95, p = .001, η2 = .44. Moreover, a Bayes factor analysis demonstrated strong evidence for the full model containing both main effects and the interaction effect, which suggests that each of the main effects as well as the interaction contributed significantly to the data (BF = 6.76 × 104). Furthermore, adding the same order restrictions as in Experiment 2 increased the Bayes factor to 5.35 × 105, which makes the full model 37 times more likely than the model including only the main effects.
Fig. 6.
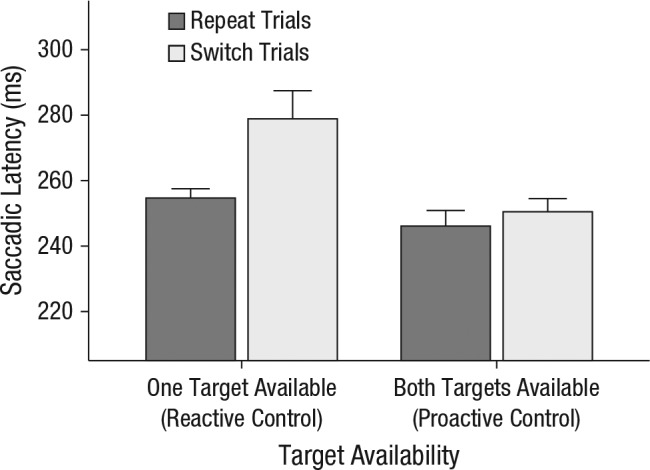
Results of Experiment 3: mean saccade latency prior to switch trials and repeat trials for each level of target availability (one target vs. both targets). For clarity, we also added the control mechanisms that apply in those conditions: Proactive control gives the observer free choice of what to search for, while reactive control limits observer choice to a single target category. Error bars represent the upper limit of within-subjects 95% confidence intervals (Morey, 2008).
When both targets were available, observers switched on 37% of all search displays, yet there was no difference in saccade latency between switch trials (M = 251 ms) and repeat trials (M = 246 ms), t(19) = 1.60, p = .13, Cohen’s d = 0.08. When only one target was available, however, latencies were longer on switch trials (M = 279 ms) than on repeat trials (M = 255 ms), t(19) = 4.05, p < .001, Cohen’s d = 0.33.
In addition, fixation accuracy confirmed the pattern of results (see Table 1). A repeated measures ANOVA with target availability and trial type as factors yielded significant main effects of target availability, F(1, 19) = 131.0, p < .001, η2 = .88, and trial type, F(1, 19) = 49.2, p < .001, η2 = .72, as well as a significant interaction between them, F(1, 19) = 64.1, p < .001, η2 > .77. Overall, performance was worse in the one-target than in the both-targets condition, which was furthermore the only condition in which switch costs were found.
Discussion
The results replicated those of Experiments 1 and 2, but within blocks. When both targets were available, there were no switch costs. When only one target was available, switch costs occurred, consistent with the notion of reactive control processes in response to unprepared-for targets.
General Discussion
The current findings support a crucial role for cognitive control in multiple-target search. When observers were asked to search for two targets, they switched between them from one fixation to the next without apparent costs, as long as both targets were available. We argue that with multiple targets available, observers can proactively prioritize any one target. This may occur during the slack of the previous eye movement or even several fixations in advance (Godijn & Theeuwes, 2003), so that any potential switch costs remain latent. In contrast, clear switch costs emerged when only one target was available for selection. This is not predicted by a multiple-template account, which assumes that observers can prepare a bias for both targets, regardless of which one is available. Instead, the results are in line with multiple-state models of working memory, which claim that only a single item in working memory has priority for the task at hand (Huang & Pashler, 2007; Oberauer, 2002; Olivers et al., 2011). When none of the items match the prioritized template, a reactive switch is required, which results in switch costs.
Recently, Beck and Hollingworth (2017; see the Supplemental Material for additional discussion) published new results that appear to contradict our findings. Using a similar gaze-contingent paradigm, they asked observers to search for one of two targets when both targets were available. Their main finding was that observers switched as often as they repeated targets from one display to the next. They argued that this is not predicted when observers can search for only one target. Indeed, we too found frequent switches (on up to 37% of trials). However, our interpretation is different: When such switches can be freely prepared for, there is no reason not to switch, as switching prior to display onset implies little cost for saccade latencies. What our results show is that when both targets are available in the display, the absence of switch costs is not diagnostic for simultaneous multiple-target search.
The absence of switch costs in the both-targets conditions might appear surprising considering the switch costs found in the literature on intertrial priming in visual search (e.g., Found & Müller, 1996; Maljkovic & Nakayama, 1994). However, consistent with our one-target conditions, those studies typically presented only one target per search display, which resulted in switch costs. When all targets are presented, any sensory priming is equated, but more important, the proactive nature of the task allows the observer’s currently active template to become a priming force itself, which takes precedence over selection history. In line with this interpretation, intertrial priming is not only driven by selection history, but is also sensitive to task goals (Fecteau, 2007; Olivers & Meeter, 2006; Rangelov, Müller, & Zehetleitner, 2011; though see Maljkovic & Nakayama, 1994).
Even when attention cannot be biased toward multiple potential targets simultaneously (limiting active visual search to one object at a time), observers can still remember multiple targets. Observers may even reverse the search process by conducting a serial scan through the objects in a display and use these to perform a multiple-target memory search instead. Indeed, Wolfe (2012) has shown that observers can successfully look for 100 objects—although adding more targets to memory resulted in additional (but logarithmically diminishing) costs. Thus, “look for” is too broad a term in this respect, as it can reflect multiple mechanisms, including template-based guidance, reactive memory retrieval (Nosofsky, Cox, Cao, & Shiffrin, 2014), or both (e.g., Cunningham & Wolfe, 2014, Experiment 3). Our results suggest that proactive template-based guidance is limited, while we make no claims about the mechanisms underlying memory-based search. Which of these mechanisms prevails in a certain search task is likely to also be important in real-world situations, such as baggage screening, in which a vast multitude of potential target objects invites a reactive strategy—unless one has a specific expectation (e.g., for a gun) and adopts a proactive approach instead.
The distinction between proactive and reactive control is well established in the cognitive-control literature and has been shown to recruit different brain regions (Braver, 2012). So far, though, it has seen little application in visual search, except for a role in the suppression of unwanted distractors (Geng, 2014). We show here that the level of control that can be exerted has implications for the efficiency of target selection when observers search for multiple targets.
Supplementary Material
Footnotes
Action Editor: Philippe G. Schyns served as action editor for this article.
Declaration of Conflicting Interests: The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
Funding: This work was supported by Open Research Area Grant 464-13-003 from the Netherlands Organization for Scientific Research and by European Research Council Consolidator Grant ERC-2013-CoG-615423 to C. N. L. Olivers.
Supplemental Material: Additional supporting information can be found at http://journals.sagepub.com/doi/suppl/10.1177/0956797617705667
Open Practices:


All data and materials have been made publicly available via the Open Science Framework and can be accessed at www.osf.io/qknug. The complete Open Practices Disclosure for this article can be found at http://journals.sagepub.com/doi/suppl/10.1177/0956797617705667. This article has received the badges for Open Data and Open Materials. More information about the Open Practices badges can be found at http://www.psychologicalscience.org/publications/badges.
References
- Beck V. M., Hollingworth A. (2017). Competition in saccade target selection reveals attentional guidance by simultaneously active working memory representations. Journal of Experimental Psychology: Human Perception and Performance, 43, 225–230. doi: 10.1037/xhp0000306 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beck V. M., Hollingworth A., Luck S. J. (2012). Simultaneous control of attention by multiple working memory representations. Psychological Science, 23, 887–898. doi: 10.1177/0956797612439068 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Braver T. S. (2012). The variable nature of cognitive control: A dual mechanisms framework. Trends in Cognitive Sciences, 16, 106–113. doi: 10.1016/j.tics.2011.12.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carlisle N. B., Arita J. T., Pardo D., Woodman G. F. (2011). Attentional templates in visual working memory. The Journal of Neuroscience, 31, 9315–9322. doi: 10.1523/JNEU-ROSCI.1097-11.2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cowan N. (2001). The magical number 4 in short-term memory: A reconsideration of mental storage capacity. Behavioral & Brain Sciences, 24, 87–185. doi: 10.1017/S0140525X01003922 [DOI] [PubMed] [Google Scholar]
- Cunningham C. A., Wolfe J. M. (2014). The role of object categories in hybrid visual and memory search. Journal of Experimental Psychology: General, 143, 1585–1599. doi: 10.1037/a0036313 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dalmaijer E. S., Mathôt S., Van der Stigchel S. (2014). PyGaze: An open-source, cross-platform toolbox for minimal-effort programming of eyetracking experiments. Behavior Research Methods, 46, 913–921. doi: 10.3758/s13428-013-0422-2 [DOI] [PubMed] [Google Scholar]
- Desimone R., Duncan J. (1995). Neural mechanisms of selective visual attention. Annual Review of Neuroscience, 18, 193–222. doi:0.1146/annurev.neuro.18.1.193 [DOI] [PubMed] [Google Scholar]
- Dombrowe I., Donk M., Olivers C. N. L. (2011). The costs of switching attentional sets. Attention, Perception, & Psychophysics, 73, 2481–2488. doi: 10.3758/s13414-011-0198-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fecteau J. H. (2007). Priming of pop-out depends upon the current goals of observers. Journal of Vision, 7(6), Article 1. doi: 10.1167/7.6.1 [DOI] [PubMed] [Google Scholar]
- Found A., Müller H. J. (1996). Searching for unknown feature targets on more than one dimension: Investigating a “dimension-weighting” account. Attention, Perception, & Psychophysics, 58, 88–101. doi: 10.3758/BF03205479 [DOI] [PubMed] [Google Scholar]
- Geng J. J. (2014). Attentional mechanisms of distractor suppression. Current Directions in Psychological Science, 23, 147–153. doi: 10.1177/0963721414525780 [DOI] [Google Scholar]
- Godijn R., Theeuwes J. (2003). Parallel allocation of attention prior to the execution of saccade sequences. Journal of Experimental Psychology: Human Perception and Performance, 29, 882–896. doi: 10.1037/0096-1523.29.5.882 [DOI] [PubMed] [Google Scholar]
- Houtkamp R., Roelfsema P. R. (2009). Matching of visual input to only one item at any one time. Psychological Research, 73, 317–326. doi: 10.1007/s00426-008-0157-3 [DOI] [PubMed] [Google Scholar]
- Huang L., Pashler H. (2007). A Boolean map theory of visual attention. Psychological Review, 114, 599–631. doi: 10.1037/0033-295X.114.3.599 [DOI] [PubMed] [Google Scholar]
- Locke H. S., Braver T. S. (2008). Motivational influences on cognitive control: Behavior, brain activation, and individual differences. Cognitive, Affective, & Behavioral Neuroscience, 8, 99–112. doi: 10.3758/CABN.8.1.99 [DOI] [PubMed] [Google Scholar]
- Maljkovic V., Nakayama K. (1994). Priming of pop-out: I. Role of features. Memory & Cognition, 22, 657–672. doi: 10.3758/BF03209251 [DOI] [PubMed] [Google Scholar]
- Mathôt S., Schreij D., Theeuwes J. (2012). OpenSesame: An open-source, graphical experiment builder for the social sciences. Behavior Research Methods, 44, 314–324. doi: 10.3758/s13428-011-0168-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Menneer T., Cave K. R., Donnelly N. (2009). The cost of search for multiple targets: Effects of practice and target similarity. Journal of Experimental Psychology: Applied, 15, 125–139. doi: 10.1037/a0015331 [DOI] [PubMed] [Google Scholar]
- Morey R. D. (2008). Confidence intervals from normalized data: A correction to Cousineau (2005). Tutorials in Quantitative Methods for Psychology, 4, 61–64. doi: 10.3758/s13414-012-0291-2 [DOI] [Google Scholar]
- Morey R. D., Wagenmakers E. J. (2014). Simple relation between Bayesian order-restricted and point-null hypothesis tests. Statistics & Probability Letters, 92, 121–124. doi: 10.1016/j.spl.2014.05.010 [DOI] [Google Scholar]
- Nosofsky R. M., Cox G. E., Cao R., Shiffrin R. M. (2014). An exemplar-familiarity model predicts short-term and long-term probe recognition across diverse forms of memory search. Journal of Experimental Psychology: Learning, Memory, and Cognition, 40, 1524–1539. doi: 10.1037/xlm0000015 [DOI] [PubMed] [Google Scholar]
- Oberauer K. (2002). Access to information in working memory: Exploring the focus of attention. Journal of Experimental Psychology: Learning, Memory, and Cognition, 28, 411–421. doi: 10.1037/0278-7393.28.3.411 [DOI] [PubMed] [Google Scholar]
- Olivers C. N. L., Eimer M. (2011). On the difference between working memory and attentional set. Neuropsychologia, 49, 1553–1558. doi: 10.1016/j.neuropsychologia.2010.11.033 [DOI] [PubMed] [Google Scholar]
- Olivers C. N. L., Meeter M. (2006). On the dissociation between compound and present/absent tasks in visual search: Intertrial priming is ambiguity driven. Visual Cognition, 13, 1–28. doi: 10.1080/13506280500277488 [DOI] [Google Scholar]
- Olivers C. N. L., Peters J., Houtkamp R., Roelfsema P. R. (2011). Different states in visual working memory: When it guides attention and when it does not. Trends in Cognitive Sciences, 15, 327–334. doi: 10.1016/j.tics.2011.05.004 [DOI] [PubMed] [Google Scholar]
- Rangelov D., Müller H. J., Zehetleitner M. (2011). Dimension-specific intertrial priming effects are task-specific: Evidence for multiple weighting systems. Journal of Experimental Psychology: Human Perception and Performance, 37, 100–114. doi: 10.1037/a0020364 [DOI] [PubMed] [Google Scholar]
- Stroud M. J., Menneer T., Cave K. R., Donnelly N., Rayner K. (2011). Search for multiple targets of different colours: Misguided eye movements reveal a reduction of colour selectivity. Applied Cognitive Psychology, 25, 971–982. doi: 10.1002/acp.1790 [DOI] [Google Scholar]
- Wolfe J. M. (2007). Guided search 4.0: Current progress with a model of visual search. In Gray W. D. (Ed.), Integrated models of cognitive systems (pp. 99–119). Oxford, England: Oxford University Press. [Google Scholar]
- Wolfe J. M. (2012). Saved by a log: How do humans perform hybrid visual and memory search? Psychological Science, 23, 698–703. doi: 10.1177/0956797612443968 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wolfe J. M., Butcher S. J., Lee C., Hyle M. (2003). Changing your mind: On the contributions of top-down and bottom-up guidance in visual search for feature singletons. Journal of Experimental Psychology: Human Perception and Performance, 29, 483–502. [DOI] [PubMed] [Google Scholar]
- Wolfe J. M., Horowitz T. S. (2004). What attributes guide the deployment of visual attention and how do they do it? Nature Reviews Neuroscience, 5, 495–501. doi: 10.1038/nrn1411 [DOI] [PubMed] [Google Scholar]
- Wolfe J. M., Horowitz T. S., Kenner N., Hyle M., Vasan N. (2004). How fast can you change your mind? The speed of top-down guidance in visual search. Vision Research, 44, 1411–1426. doi: 10.1016/j.visres.2003.11.024 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.