

Abstract
Nitric oxide (NO) has a wide variety of physiological functions in the kidney. Besides the regulatory effects in intrarenal haemodynamics and glomerular microcirculation, in vivo studies reported the diuretic and natriuretic effects of NO. However, opposite results showing the stimulatory effect of NO on Na+ reabsorption in the proximal tubule led to an intense debate on its physiological roles. Animal studies have showed the biphasic effect of angiotensin II (Ang II) and the overall inhibitory effect of NO on the activity of proximal tubular Na+ transporters, the apical Na+/H+ exchanger isoform 3, basolateral Na+/K+ ATPase, and the Na+/HCO3− cotransporter. However, whether these effects could be reproduced in humans remained unclear. Notably, our recent functional analysis of isolated proximal tubules demonstrated that Ang II dose-dependently stimulated human proximal tubular Na+ transport through the NO/guanosine 3′,5′-cyclic monophosphate (cGMP) pathway, confirming the human-specific regulation of proximal tubular transport via NO and Ang II. Of particular importance for this newly identified pathway is its possibility of being a human-specific therapeutic target for hypertension. In this review, we focus on NO-mediated regulation of proximal tubular Na+ transport, with emphasis on the interaction with individual Na+ transporters and the crosstalk with Ang II signalling.
1. Introduction
The proximal tubule is a key site for Na+ homeostasis via reabsorption of most of the water and solutes filtered by the glomerulus. Angiotensin II (Ang II), a powerful vasoconstrictor peptide, affects blood pressure via maintenance of Na+ homeostasis through its effects on the machinery for proximal tubular Na+ transport, including the apical Na+/H+ exchanger isoform 3 (NHE3), basolateral Na+/K+ ATPase (NKA), and the Na+/HCO3− cotransporter (NBCe1) [1–5]. Interestingly, previous animal studies have confirmed that the effect of Ang II on proximal tubular transport is biphasic. Thus, low concentrations of Ang II increase transport activities, whereas high concentrations inhibit transport activities [6, 7]. However, whether the biphasic effect of Ang II could be reproduced in humans remained unclear.
Nitric oxide (NO) is a small gas molecule that diffuses freely through the plasma membranes of target cells and activates guanosine 3′,5′-cyclic monophosphate (cGMP) formation. Although NO was previously regarded as a toxic air pollutant, the discovery that it is identical to the previously recognised endothelium-derived relaxing factor [8, 9] has revealed its various physiological roles in the cardiovascular, neurologic, and immune systems [10–12]. Indeed, investigations have demonstrated the net natriuretic and diuretic effects and have indicated the inhibitory effect of NO on proximal tubular Na+ transport [13–15]. However, the reported effects were sometimes inconsistent or even controversial, partly because of the modification of NO effects by other regulatory and/or compensatory mechanisms occurring in a specific experimental condition. On the other hand, studies have shown that Ang II can mediate NO production via its specific cell-surface receptors (AT1 and AT2), suggesting a possible role of NO as a secondary messenger for Ang II-mediated regulation of proximal tubular transport [12, 16]. Therefore, the crosstalk between NO and Ang II signalling is an important factor to determine the effects of NO on proximal tubular transport. Here, we review NO-mediated regulation of proximal tubular Na+ transport, with emphasis on the interaction with individual Na+ transporters and the crosstalk with Ang II signalling.
2. Nitric Oxide Synthase (NOS): Isoforms and Distribution
NOS is a flavohemeprotein that catalyses the oxidation of l-arginine to l-citrulline with the production of NO, an important bioregulatory molecule in various physiological process such as neurotransmission, vasorelaxation, platelet aggregation, and immune responses [17–19]. The three known NOS isoforms are neuronal NOS (nNOS), endothelial NOS (eNOS), and inducible NOS (iNOS), and these show tissue-specific distribution and physiological functions [20–22].
nNOS is constitutively expressed throughout the central and peripheral nervous system, and it is associated with neuronal signalling. It produces low levels of NO, and its enzyme activity is strictly dependent on intracellular Ca2+ and calmodulin [23]. Similar to nNOS, eNOS generates relatively low levels of NO in the vascular endothelium and regulates vascular homeostasis in a Ca2+-dependent manner [24]. In contrast with these constitutive NOS enzymes, iNOS expression lacks cell specificity. Thus, it is not usually expressed but can be induced in almost all cell types on immune-related stimulation or gene transcription associated with bacterial lipopolysaccharides (LPSs), inflammatory cytokines, and other chemical mediators [25, 26]. Once induced, iNOS produces high levels of NO independently of the intracellular Ca2+ concentration [27], contributing to the pathophysiology of inflammatory disease and septic shock [25, 28].
3. Distribution of NOS in the Kidney
In the kidney, all three NOS isoforms are expressed at various locations along the nephron and are subjected to distinct control mechanisms [18, 29]. Immunostaining analysis has shown high expression of nNOS in the macula densa in all species, including humans [30, 31], suggesting an important role of nNOS in the juxtaglomerular apparatus and tubuloglomerular feedback (TGF) [32]. Further, the mRNA expression of nNOS has been reported in the thin limb of the loop of Henle and the medullary collecting duct [33]. eNOS is expressed widely in the epithelial cells of intrarenal vessels except in the venous system and regulates renal vascular tone [34]. In the tubular segments, high levels of eNOS mRNA have been reported in the proximal tubule, medullary thick ascending limb of the loop of Henle (mTAL), and collecting duct [24, 35].
iNOS is considered to have a wide distribution in the tubular epithelium. Generally, its expression is only induced after appropriate stimulation; however, in situ hybridisation studies have identified constitutive expression of iNOS mRNA in normal rat kidneys. iNOS mRNA has been detected in the S3 segment of the proximal tubule, mTAL, distal convoluted tubule, cortical collecting duct, and inner medullary collecting duct [36].
4. Physiological Functions of NO in Nephron Transport
It is well known that NO has a wide variety of physiological functions in the kidney based on the unique NOS population. For example, as a potent vasodilator, NO contributes to maintain a low vascular tone necessary for normal renal blood flow [37, 38]. On the other hand, NO released from the macula densa is involved in rennin secretion and TGF response via vasoconstriction of the afferent arterioles [39–41]. Besides the regulatory roles of NO in intrarenal haemodynamics and glomerular microcirculation, in vivo animal studies have shown the physiological importance of NO in renal Na+ transport [42, 43]. In these studies, infusion of NOS inhibitors into the kidney reduced water and Na+excretion, whereas the stimulation of NO production increased excretion. No significant change in the glomerular filtration rate (GFR) or renal blood flow (RBF) was observed with regard to the natriuretic and diuretic effects of NO, suggesting that NO could directly suppress nephron transport.
Considering the fact that NOS KO mice were significantly hypertensive and showed various cardiovascular abnormalities [44, 45], NO and NOS in nephron transport are possible key factors to elucidate the aetiology of Na+ retention and resultant hypertension. However, the functional significance of NO on individual nephron segments has recently been questioned [43, 45–47]. Despite the overall inhibitory effect of NO in nephron transport, several studies have demonstrated some controversial results for each nephron segment [43, 48–53]. Above all, the renal proximal tubule, which is the most important nephron portion for water and Na+ reabsorption, is intimately involved in NO-mediated renal Na+ transport regulation.
5. Effects of NO on Proximal Tubular Na+ Transport
Proximal tubules reabsorb approximately two-thirds of water and Na+ filtered in the glomeruli along with glucose, phosphate, amino acids and bicarbonates (HCO3−). Most of these solutes are carried via a series of Na+-coupled cotransporters or exchangers, which utilise the transmembrane Na+ gradient generated by NKA on the basolateral membrane [42]. Proximal tubule Na+ transport is thus primarily driven by NKA and predominantly accomplished by NHE3 on the apical membrane and NBCe1 on the basolateral membrane (Figure 1).
Figure 1.
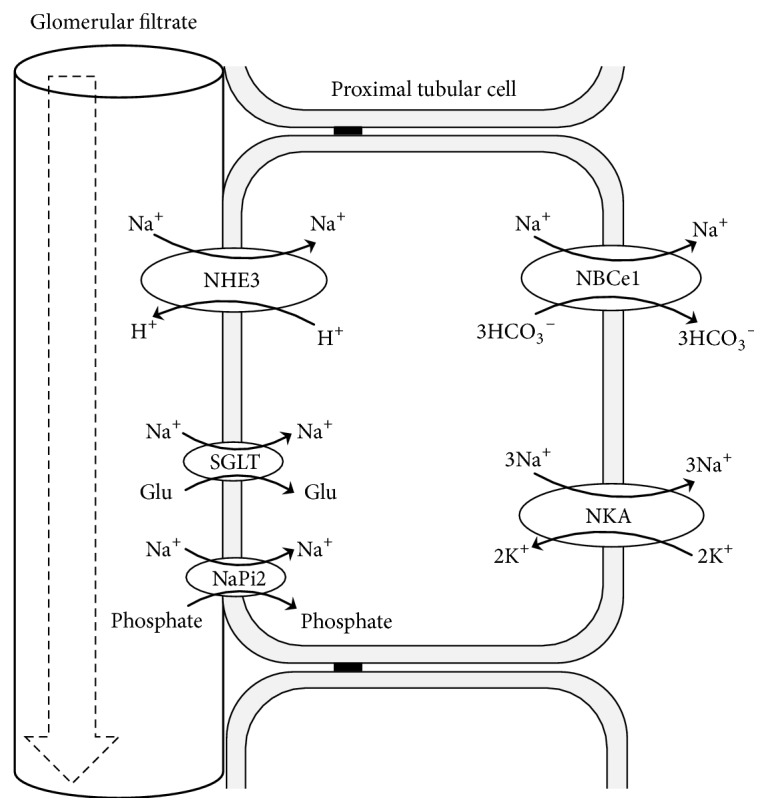
Proximal tubular Na + reabsorption. Active Na+ transport mediated by basolateral NKA provides the driving force for proximal tubular Na+ transport. Transporters in the apical membrane include NHE3, SGLT, and NaPi2. NHE3 is considered to be responsible for most of the Na+ reabsorption from the glomerular filtrate. NBCe1 at the basolateral membrane plays an important role not only in Na+ homeostasis but also in the systemic acid/base balance. NKA, Na+/K+ ATPase; NHE3, Na+/H+ exchanger isoform 3; NBCe1, Na+/HCO3− cotransporter; Glu, glucose; SGLT, Na+/glucose cotransporter; NaPi2, Na+/phosphate cotransporter type 2.
Although the effect of NO on proximal tubular Na+ transport is still controversial, a number of animal studies have reported its overall inhibitory effect. For example, in rat studies, the NO donor sodium nitroprusside (SNP) reduced proximal tubule fluid absorption (Jv), whereas the NO inhibitor NG-monomethyl-L-arginine (l-NAME) increased Jv, suggesting the inhibitory effect of NO on proximal tubular Na+ reabsorption [50, 54]. Further, using the microperfusion technique, Vallon et al. reported higher fluid and chloride reabsorption rates in the proximal tubules of nNOS KO mice than in the tubules of wild-type mice [51]. In contrast, Wang observed that both rats and mice treated with l-NAME presented significant diuresis and natriuresis due to decreased Jv and renal tubular absorption of HCO3− (JHCO3), indicating the stimulatory effect of NO on proximal tubular transport [52, 53]. Moreover, they suggested that nNOS and iNOS could directly stimulate proximal tubular Na+ transport on the basis of lower Jv and JHCO3 in respective gene KO mice [55].
One possible reason for the discrepancy in the results on the effects of NO on proximal tubular transport is that the drug effects are modified by other regulatory and/or compensatory mechanisms occurring in a specific experimental condition. For example, effects of l-NAME on proximal tubular transport can be inhibited by denervation of the kidney, suggesting that the results reflect the differences in neural activity [54]. Additionally, the systemic administration of NOS inhibitors has been reported to increase renal perfusion pressure and induce flow-dependent changes in Jv and JHCO3 [56]. Furthermore, KO mice studies involve the influence of removal of the NOS gene from not only the kidney but also all from other tissues, which can modify a variety of regulatory mechanisms for proximal tubular transport, such as hormonal regulation, renal autoregulation, and neural regulation. Therefore, it is unclear whether the data obtained from these in vivo studies truly reflect the direct effect of NO on specific nephron segments. To delineate the influence of such interaction and regulatory mechanisms, in vitro microperfusion studies should be performed.
6. NO as a Potential Mediator of Ang II
Ang II, a powerful effector of the renin-angiotensin system, controls blood pressure partially by regulating water and Na+ homeostasis [57–59]. Its various effects are mediated by the specific cell-surface receptors AT1 and AT2. The AT1 receptor is ubiquitously expressed and mediates most of the physiological functions of Ang II, including vasoconstriction, stimulation of aldosterone release and sympathetic nerve activity, promotion of cell growth, and inflammation, whereas the AT2 receptor functionally antagonises the effects of the AT1 receptor [60, 61].
Studies have provided evidence of functional interaction between Ang II and NO in the vascular system, where Ang II regulates the NO/cGMP signalling pathway both by the AT1 and by AT2 receptors. Ang II, via the AT2 receptor, stimulates bradykinin-dependent NO production [62]. In addition, via AT1 receptors, it also triggers the Ca2+/calmodulin-dependent eNOS activation in bovine endothelial cells [63]. Then Ang II-induced NO exerts a vasodilating effect through the NO/cGMP signalling pathway to protect against Ang II vasoconstriction. Conversely, long-term infusion of Ang II is known to decrease NO bioavailability via the AT1 receptor by increasing NAD(P)H oxidase-mediated vascular superoxide production [64]. Further, the superoxide also reduces the activity of soluble guanylyl cyclase, inhibiting the NO/cGMP signalling pathway. Such functional interactions between Ang II and NO are regulated precisely at multiple levels, and the imbalance between Ang II and NO is considered to be the aetiology of many cardiovascular diseases including hypertension, atherosclerosis, and congestive heart failure.
In the proximal tubule of mice, rats, and rabbits, Ang II regulates Na+ transport in a biphasic manner such that low concentrations (picomolar to nanomolar) stimulate and high concentrations (nanomolar to micromolar) inhibit reabsorption [3, 6, 7, 65]. Although the responsible receptors had been debated [4, 6, 66, 67], studies using isolated proximal tubules from KO mice have demonstrated that the biphasic effect of Ang II on Na+ transport is predominantly mediated via the AT1A receptor, an isoform of the AT1 receptor [3, 65]. With regard to the stimulatory effect of Ang II, the involvement of the activation of protein kinase C (PKC) and/or the decrease in the intracellular cAMP concentration, which can activate the extracellular signal-regulated kinase (ERK) pathway, has been suggested [4, 68, 69]. On the other hand, it has been reported that activation of NO and its downstream effector cGMP by Ang II mediates the inhibitory effect by acting on the machinery for proximal tubular Na+ transport, NKA, NHE3, and NBCe1 [5, 70–72].
6.1. NKA
NKA is an active transporter on the basolateral membrane, which pumps Na+ from the cytoplasm to the interstitium against its concentration gradient, supplying the driving force for proximal tubular Na+ reabsorption. The impact of the NO/cGMP pathway on NKA activity has been observed in several cell lines. In cultured mouse proximal tubular cells (MCT cells), Guzman et al. found that endogenous NO/iNOS stimulation by LPS and interferon-gamma decreased the catalytic activity of ouabain-sensitive ATPase, consistent with the inhibitory effect of NO on NKA [73]. Further, in opossum kidney cells, NKA activity was inhibited by not only NO but also by a cGMP analogue, suggesting the possible presence of the NO/cGMP pathway in the regulation of proximal tubular NKA [74, 75]. In accordance with the data from cell experiments, Zhang et al. proposed the presence of crosstalk between the NO/cGMP pathway and Ang II signalling in rat proximal tubules [76]. Ang II stimulates both NKA activity and NO synthesis via the AT1 receptor at a low concentration, and as the peritubular concentration of Ang II rises, NO/cGMP signalling inhibits NKA activity, which is partially attributed to the biphasic regulation of Ang II for proximal tubular Na+ transport. In contrast, NO was shown to have no impact on NKA activity in LLC-PK1 cells (another cell line derived from pig kidney) [74, 77]. Taken together, these data suggest a definite inhibitory effect of NO/NOS on proximal tubular NKA; however, future studies are required to investigate the cell-specific heterogeneity in the regulation of NKA activity via the NO/cGMP signalling pathway.
6.2. NHE3
NHE3, the epithelial isoform of the Na+/H+ exchanger, is abundantly expressed in the apical membrane of renal proximal tubules and is responsible for most of the Na+/H+ change activity in the region [78–80]. In support of in vivo data that demonstrated NO-induced natriuresis, NO has been shown to inhibit NHE3 activity in the proximal tubule [81–83]. Roczniak et al. first investigated the effect of NO on rabbit proximal tubules by using the NO donor sodium nitroprusside (SNP) and concluded that NO inhibits Na+/H+ exchange activity via cGMP elevation [83]. In addition, it has been reported that chronic blockade of NO synthesis by l-NAME results in increased expression of NHE3, suggesting the regulatory role of endogenous NO in the expression of Na+ transporters in the kidneys [84].
The biphasic effect of Ang II via the AT1 receptor on proximal tubular NHE3 activity has been observed in several animal studies [6, 85, 86]. It has been suggested that while stimulation of NHE3 at a low Ang II concentration is complicated in the PKC, PKA, PI3 kinase, or phospholipase C (PLC)-calmodulin pathway [1, 87–89], the inhibitory effect of a high Ang II concentration is mediated by the activation of the cGMP/cGMP-dependent protein kinase (PKG) pathway [1].
6.3. NBCe1
NBCe1 is predominantly expressed in the basolateral membrane of renal proximal tubules, and it mediates most of the Na+-coupled HCO3− cotransport, playing a pivotal role in Na+ homeostasis and systemic acid/base balance [90, 91]. Similar to NKA and NHE3, microperfusion studies demonstrated that the biphasic effect of Ang II on NBCe1 is observed in mouse and rat proximal tubules [3, 69, 92]. Further, our recent study revealed that the NO/cGMP/cGMP-dependent kinase II (cGKII) pathway mediates the inhibitory effect of Ang II on mouse proximal tubular NBCe1. Thus, the inhibitory effect of Ang II was lost in proximal tubules of cGKII-KO mice, and neither a SNP nor a cGMP analogue phosphorylated ERK. The overall data on NKA, NHE3, and NBCe1 obtained from animal studies support the inhibitory effect of NO on proximal tubular Na+ transport.
7. Human-Specific Effects of NO and Ang II on Proximal Tubular Na+ Transport
The effects of NO and Ang II on human proximal tubular transport have been widely discussed. Previous studies in humans demonstrated that systemic administration of NOS inhibitors decreased the fractional excretion of Na+ (FENa) and lithium (FELi), which was consistent with the inhibitory effect of NO on proximal tubular Na+ reabsorption [13–15]. However, it has been reported that the NO derivative failed to induce natriuresis [93] and was associated with Na+ retention and plasma volume expansion in humans [94], suggesting the presence of human-specific NO effects. Moreover, Rosenbaek et al. recently confirmed that sodium nitrate also failed to induce natriuresis in humans [95].
Interestingly, our functional analysis of isolated proximal tubules obtained from nephrectomy surgery demonstrated that Ang II dose-dependently stimulated human renal proximal tubular transport via ERK phosphorylation through the NO/cGMP pathway [92]. Thus, Ang II in human proximal tubules stimulates basolateral NBCe1 activity in a monophasic manner, unlike in animal proximal tubules. Furthermore, in luminal perfused human proximal tubules, we found that Ang II dose-dependently stimulated apical NHE3 activity and JHCO3. The exact reasons for such difference among species in the regulation of proximal tubular Na+ transport associated with the NO/cGMP pathway (Figure 2) are still unknown, but a high peritubular Ang II concentration [96], via its monophasic effect, would be more strongly associated with human hypertension [97].
Figure 2.

Species difference in Ang II-mediated regulation of proximal tubular Na + transport. In humans, Ang II dose-dependently stimulates the NO/cGMP pathway via the AT1 receptor over a wide range of concentrations (from low to high). The signalling cascade phosphorylates ERK, resulting in the stimulation of proximal tubular Na+ reabsorption, although the interplay between Ang II and NO signalling pathway on the NKA activity is still unconfirmed. In animal proximal tubules (mice, rats, and rabbits), a high concentration of Ang II also stimulates the NO/cGMP pathway, but the subsequent signalling cascade activates cGKII to exert inhibitory effects on Na+ reabsorption. As for NHE3, the inhibitory effect of Ang II is involved in the cGMP-dependent protein kinase, which is independent of the NO/cGMP pathway. Ang II, angiotensin II; AT1, angiotensin II receptor type 1; NO, nitric oxide; cGMP, guanosine 3′,5′-cyclic monophosphate; ERK, extracellular signal-regulated kinase; cGKII, cGMP-dependent protein kinase type II; NKA, Na+/K+ ATPase; NHE3, Na+/H+ exchanger isoform 3; NBCe1, Na+/HCO3− cotransporter.
8. Conclusion
NO plays a fundamental role in the regulation of proximal tubular Na+ transport. Although the direct effect of NO on proximal tubular transport has been intensely debated, the crosstalk with Ang II signalling and the interaction with Na+ transporters are possible key factors in solving this issue. The species difference in NO-mediated Ang II function is noteworthy. Our recent study identified human-specific monophasic regulation of proximal tubular transport through crosstalk between NO and Ang II signalling. Although its physiological significance is still unknown, the newly identified pathway that mediates the dose-dependent stimulatory effect of Ang II is expected to be a human-specific therapeutic target of hypertension. Future studies are necessary to clarify whether such a phenomenon is reproduced in other nephron segments, which has not yet been fully investigated.
Acknowledgments
This review was supported in part by the grants from the Tokyo Society of Medical Science and the Ministry of Education, Culture, Sports, Science and Technology of Japan (no. 16H06750).
Conflicts of Interest
The authors declare that there are no conflicts of interest regarding the publication of this paper.
References
- 1.Banday A. A., Lokhandwala M. F. Angiotensin II-mediated biphasic regulation of proximal tubular Na+/H+ exchanger 3 is impaired during oxidative stress. American Journal of Physiology-Renal Physiology. 2011;301(2):F364–F370. doi: 10.1152/ajprenal.00121.2011. [DOI] [PubMed] [Google Scholar]
- 2.Coppola S., Frömter E. An electrophysiological study of angiotensin II regulation of Na-HCO3 cotransport and K conductance in renal proximal tubules - II. Effect of micromolar concentrations. Pflügers Archiv - European Journal of Physiology. 1994;427(1-2):151–156. doi: 10.1007/BF00585954. [DOI] [PubMed] [Google Scholar]
- 3.Horita S., Zheng Y., Hara C., et al. Biphasic regulation of Na+-HCO3- cotransporter by angiotensin II type 1A receptor. Hypertension. 2002;40(5):707–712. doi: 10.1161/01.HYP.0000036449.70110.DE. [DOI] [PubMed] [Google Scholar]
- 4.Houillier P., Chambrey R., Achard J. M., Froissart M., Poggioli J., Paillard M. Signaling pathways in the biphasic effect of angiotensin II on apical Na/H antiport activity in proximal tubule. Kidney International. 1996;50(5):1496–1505. doi: 10.1038/ki.1996.464. [DOI] [PubMed] [Google Scholar]
- 5.Zhang C., Mayeux P. R. NO/cGMP signaling modulates regulation of Na+-K+-ATPase activity by angiotensin II in rat proximal tubules. American Journal of Physiology. 2001;280(3):F474–F479. doi: 10.1152/ajprenal.2001.280.3.F474. [DOI] [PubMed] [Google Scholar]
- 6.Harris P. J., Young J. A. Dose-dependent stimulation and inhibition of proximal tubular sodium reabsorption by angiotensin II in the rat kidney. Pflügers Archiv - European Journal of Physiology. 1977;367(3):295–297. doi: 10.1007/BF00581370. [DOI] [PubMed] [Google Scholar]
- 7.Schuster V. L., Kokko J. P., Jacobson H. R. Angiotensin II directly stimulates sodium transport in rabbit proximal convoluted tubules. The Journal of Clinical Investigation. 1984;73(2):507–515. doi: 10.1172/JCI111237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Palmer R. M. J., Ferrige A. G., Moncada S. Nitric oxide release accounts for the biological activity of endothelium-derived relaxing factor. Nature. 1987;327(6122):524–526. doi: 10.1038/327524a0. [DOI] [PubMed] [Google Scholar]
- 9.Ignarro L. J., Buga G. M., Wood K. S., Byrns R. E., Chaudhuri G. Endothelium-derived relaxing factor produced and released from artery and vein is nitric oxide. Proceedings of the National Academy of Sciences of the United States of America. 1987;84(24):9265–9269. doi: 10.1073/pnas.84.24.9265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Tsutsui M., Tanimoto A., Tamura M., et al. Significance of nitric oxide synthases: Lessons from triple nitric oxide synthases null mice. Journal of Pharmacological Sciences. 2015;127(1):42–52. doi: 10.1016/j.jphs.2014.10.002. [DOI] [PubMed] [Google Scholar]
- 11.Chaki S., Inagami T. New signaling mechanism of angiotensin II in neuroblastoma Neuro-2A cells: Activation of soluble guanylyl cyclase via nitric oxide synthesis. Molecular Pharmacology. 1993;43(4):603–608. [PubMed] [Google Scholar]
- 12.Côté F., Laflamme L., Payet M. D., Gallo-Payet N. Nitric oxide, a new second messenger involved in the action of angiotensin II on neuronal differentiation of NG108-15 cells. Endocrine Research. 1998;24(3-4):403–407. doi: 10.3109/07435809809032622. [DOI] [PubMed] [Google Scholar]
- 13.Bech J. N., Nielsen C. B., Pedersen E. B. Effects of systemic NO synthesis inhibition on RPF, GFR, UNa, and vasoactive hormones in healthy humans. American Journal of Physiology-Endocrinology and Metabolism. 1996;270(5):F845–F851. doi: 10.1152/ajprenal.1996.270.5.F845. [DOI] [PubMed] [Google Scholar]
- 14.Broere A., Van Den Meiracker A. H., Boomsma F., Derkx F. H., Veld A. J., Schalekamp M. A. Human renal and systemic hemodynamic, natriuretic, and neurohumoral responses to different doses of L-NAME. American Journal of Physiology. 1998;275(6):F870–F877. doi: 10.1152/ajprenal.1998.275.6.F870. [DOI] [PubMed] [Google Scholar]
- 15.Montanari A., Tateo E., Fasoli E., et al. Dopamine-2 receptor blockade potentiates the renal effects of nitric oxide inhibition in humans. Hypertension. 1998;31(1):277–282. doi: 10.1161/01.HYP.31.1.277. [DOI] [PubMed] [Google Scholar]
- 16.Siragy H. M., Inagami T., Carey R. M. NO and cGMP mediate angiotensin AT2 receptor-induced renal renin inhibition in young rats. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology. 2007;293(4):R1461–R1467. doi: 10.1152/ajpregu.00014.2007. [DOI] [PubMed] [Google Scholar]
- 17.Hall A. V., et al. Structural organization of the human neuronal nitric oxide synthase gene (NOS1) The Journal of Biological Chemistry. 1994;269(52):33082–33090. [PubMed] [Google Scholar]
- 18.Kone B. C. Nitric oxide synthesis in the kidney: Isoforms, biosynthesis, and functions in health. Seminars in Nephrology. 2004;24(4):299–315. doi: 10.1016/j.semnephrol.2004.04.002. [DOI] [PubMed] [Google Scholar]
- 19.Nakane M., Pollock J. S., Klinghofer V., et al. Functional Expression of Three Isoforms of Human Nitric Oxide Synthase in Baculovirus-Infected Insect Cells. Biochemical and Biophysical Research Communications. 1995;206(2):511–517. doi: 10.1006/bbrc.1995.1073. [DOI] [PubMed] [Google Scholar]
- 20.Kone B. C. Localization and regulation of nitric oxide synthase isoforms in the kidney. Semin Nephrol. 1999;19(3):230–241. [PubMed] [Google Scholar]
- 21.Bredt D. S. Endogenous nitric oxide synthesis: Biological functions and pathophysiology. Free Radical Research. 1999;31(6):577–596. doi: 10.1080/10715769900301161. [DOI] [PubMed] [Google Scholar]
- 22.Mount P. F., Power D. A. Nitric oxide in the kidney: Functions and regulation of synthesis. Acta Physiologica. 2006;187(4):433–446. doi: 10.1111/j.1748-1716.2006.01582.x. [DOI] [PubMed] [Google Scholar]
- 23.Schmidt H. H. H. W., Smith R. M., Nakane M., Murad F. Ca2+/calmodulin-dependent NO Synthase type I: A biopteroflavoprotein with Ca2+/calmodulin-independent diaphorase and reductase activities. Biochemistry. 1992;31(12):3243–3249. doi: 10.1021/bi00127a028. [DOI] [PubMed] [Google Scholar]
- 24.Han K.-H., Lim J.-M., Kim W.-Y., Kim H., Madsen K. M., Kim J. Expression of endothelial nitric oxide synthase in developing rat kidney. American Journal of Physiology-Renal Physiology. 2005;288(4):F694–F702. doi: 10.1152/ajprenal.00085.2004. [DOI] [PubMed] [Google Scholar]
- 25.Liu S. F., Adcock I. M., Old R. W., Barnes P. J., Evans T. W. Lipopolysaccharide Treatment in Vivo Induces Widespread Tissue Expression of Inducible Nitric Oxide Synthase mRNA. Biochemical and Biophysical Research Communications. 1993;196(3):1208–1213. doi: 10.1006/bbrc.1993.2380. [DOI] [PubMed] [Google Scholar]
- 26.Saha R. N., Pahan K. Regulation of inducible nitric oxide synthase gene in glial cells. Antioxidants & Redox Signaling. 2006;8(5-6):929–947. doi: 10.1089/ars.2006.8.929. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Albakri Q. A., Stuehr D. J. Intracellular assembly of inducible NO synthase is limited by nitric oxide-mediated changes in heme insertion and availability. The Journal of Biological Chemistry. 1996;271(10):5414–5421. doi: 10.1074/jbc.271.10.5414. [DOI] [PubMed] [Google Scholar]
- 28.Boveris A., Alvarez S., Navarro A. The role of mitochondrial nitric oxide synthase in inflammation and septic shock. Free Radical Biology & Medicine. 2002;33(9):1186–1193. doi: 10.1016/S0891-5849(02)01009-2. [DOI] [PubMed] [Google Scholar]
- 29.Bachmann S., Bosse H. M., Mundel P. Topography of nitric oxide synthesis by localizing constitutive NO synthases in mammalian kidney. The American Journal of Physiology—Renal Fluid and Electrolyte Physiology. 1995;268(5):F885–F898. doi: 10.1152/ajprenal.1995.268.5.F885. [DOI] [PubMed] [Google Scholar]
- 30.Ollerstam A., Persson A. E. G. Macula densa neuronal nitric oxide synthase. Cardiovascular Research. 2002;56(2):189–196. doi: 10.1016/S0008-6363(02)00536-9. [DOI] [PubMed] [Google Scholar]
- 31.Ito S., Ren Y. Evidence for the role of nitric oxide in macula densa control of glomerular hemodynamics. The Journal of Clinical Investigation. 1993;92(2):1093–1098. doi: 10.1172/JCI116615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Wilcox C. S. Role of macula densa NOS in tubuloglomerular feedback. Current Opinion in Nephrology and Hypertension. 1998;7(4):443–449. doi: 10.1097/00041552-199807000-00016. [DOI] [PubMed] [Google Scholar]
- 33.Terada Y., Tomita K., Nonoguchi H., Marumo F. Polymerase chain reaction localization of constitutive nitric oxide synthase and soluble guanylate cyclase messenger RNAs in microdissected rat nephron segments. The Journal of Clinical Investigation. 1992;90(2):659–665. doi: 10.1172/JCI115908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Kone B. C., Baylis C. Biosynthesis and homeostatic roles of nitric oxide in the normal kidney. American Journal of Physiology-Renal Physiology. 1997;272(5):F561–F578. doi: 10.1152/ajprenal.1997.272.5.F561. [DOI] [PubMed] [Google Scholar]
- 35.Tojo A., Welch W. J., Bremer V., et al. Colocalization of demethylating enzymes and NOS and functional effects of methylarginines in rat kidney. Kidney International. 1997;52(6):1593–1601. doi: 10.1038/ki.1997.490. [DOI] [PubMed] [Google Scholar]
- 36.Ahn Kyu Y., Mohaupt M. G., Madsen K. M., Kone B. C. In situ hybridization localization of mRNA encoding inducible nitric oxide synthase in rat kidney. American Journal of Physiology. 1994;267(5):F748–F757. doi: 10.1152/ajprenal.1994.267.5.F748. [DOI] [PubMed] [Google Scholar]
- 37.Navar L. G., Inscho E. W., Majid S. A., Imig J. D., Harrison-Bernard L. M., and Mitchell K. D. Paracrine regulation of the renal microcirculation. Physiological Reviews. 1996;76(2):425–536. doi: 10.1152/physrev.1996.76.2.425. [DOI] [PubMed] [Google Scholar]
- 38.Sonntag M., Deussen A., Schrader J. Role of nitric oxide in local blood flow control in the anaesthetized dog. Pflügers Archiv - European Journal of Physiology. 1992;420(2):194–199. doi: 10.1007/BF00374990. [DOI] [PubMed] [Google Scholar]
- 39.Castrop H., Schweda F., Mizel D., et al. Permissive role of nitric oxide in macula densa control of renin secretion. American Journal of Physiology-Renal Physiology. 2004;286(5):F848–F857. doi: 10.1152/ajprenal.00272.2003. [DOI] [PubMed] [Google Scholar]
- 40.Sayago C. M., Beierwaltes W. H. Nitric oxide synthase and cGMP-mediated stimulation of renin secretion. American Journal of Physiology. Regulatory, Integrative and Comparative Physiology. 2001;281(4):R1146–11451. doi: 10.1152/ajpregu.2001.281.4.R1146. [DOI] [PubMed] [Google Scholar]
- 41.Ren Y., Garvin J. L., Carretero O. A. Role of macula densa nitric oxide and cGMP in the regulation of tubuloglomerular feedback. Kidney International. 2000;58(5):2053–2060. doi: 10.1046/j.1523-1755.2000.00377.x. doi: 10.1046/j.1523-1755.2000.00377.x. [DOI] [PubMed] [Google Scholar]
- 42.Garvin J. L., Herrera M., Ortiz P. A. Regulation of renal NaCl transport by nitric oxide, endothelin, and ATP: Clinical implications. Annual Review of Physiology. 2011;73:359–376. doi: 10.1146/annurev-physiol-012110-142247. [DOI] [PubMed] [Google Scholar]
- 43.Ortiz P. A., Garvin J. L. Role of nitric oxide in the regulation of nephron transport. American Journal of Physiology-Renal Physiology. 2002;282(5):F777–F784. doi: 10.1152/ajprenal.00334.2001. [DOI] [PubMed] [Google Scholar]
- 44.Morishita T., et al. Nephrogenic diabetes insipidus in mice lacking all nitric oxide synthase isoforms. Proceedings of the National Academy of Sciences of the United States. 2005;102(30):10616–10621. doi: 10.1073/pnas.0502236102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Stauss H. M., Gödecke A., Mrowka R., Schrader J., Persson P. B. Enhanced blood pressure variability in eNOS knockout mice. Hypertension. 1999;33(6):1359–1363. doi: 10.1161/01.HYP.33.6.1359. [DOI] [PubMed] [Google Scholar]
- 46.Pearce D., Soundararajan R., Trimpert C., Kashlan O. B., Deen P. M., Kohan D. E. Collecting Duct Principal Cell Transport Processes and Their Regulation. Clinical Journal of the American Society of Nephrology. 2015;10(1):135–146. doi: 10.2215/CJN.05760513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Herrera M., Ortiz P. A., Garvin J. L. Regulation of thick ascending limb transport: Role of nitric oxide. American Journal of Physiology-Renal Physiology. 2006;290(6):F1279–F1284. doi: 10.1152/ajprenal.00465.2005. [DOI] [PubMed] [Google Scholar]
- 48.Good D. W., George T., Wang D. H. Angiotensin II inhibits absorption via a cytochrome P-450-dependent pathway in MTAL. American Journal of Physiology - Renal Physiology. 1999;276(5):F726–F736. doi: 10.1152/ajprenal.1999.276.5.F726. [DOI] [PubMed] [Google Scholar]
- 49.Ortiz P. A., Garvin J. L. Autocrine effects of nitric oxide on HCO3- transport by rat thick ascending limb. Kidney International. 2000;58(5):2069–2074. doi: 10.1046/j.1523-1755.2000.00379.x. [DOI] [PubMed] [Google Scholar]
- 50.Eitle E., Hiranyachattada S., Wang H., Harris P. J. Inhibition of proximal tubular fluid absorption by nitric oxide and atrial natriuretic peptide in rat kidney. American Journal of Physiology-Cell Physiology. 1998;274(4):C1075–C1080. doi: 10.1152/ajpcell.1998.274.4.C1075. [DOI] [PubMed] [Google Scholar]
- 51.Vallon V., Traynor T., Barajas L., Huang Y. G., Briggs J. P., Schnermann J. Feedback control of glomerular vascular tone in neuronal nitric oxide synthase knockout mice. Journals of the American Society of Nephrology. 2001;12(8):1599–1606. doi: 10.1681/ASN.V1281599. [DOI] [PubMed] [Google Scholar]
- 52.Wang T. Nitric oxide regulates HCO3 and Na+ transport by a cGMP-mediated mechanism in the kidney proximal tubule. American Journal of Physiology-Renal Physiology. 1997;272(2):F242–F248. doi: 10.1152/ajprenal.1997.272.2.F242. [DOI] [PubMed] [Google Scholar]
- 53.Wang T., Inglis F. M., Kalb R. G. Defective fluid and HCO3/- absorption in proximal tubule of neuronal nitric oxide synthase-knockout mice. American Journal of Physiology-Renal Physiology. 2000;279(3):F518–F524. doi: 10.1152/ajprenal.2000.279.3.F518. [DOI] [PubMed] [Google Scholar]
- 54.Wu X. C., Harris P. J., Johns E. J. Nitric oxide and renal nerve-mediated proximal tubular reabsorption in normotensive and hypertensive rats. The American Journal of Physiology. 1999;277(4):F560–F566. doi: 10.1152/ajprenal.1999.277.4.F560. [DOI] [PubMed] [Google Scholar]
- 55.Wang T. Role of iNOS and eNOS in modulating proximal tubule transport and acid-base balance. American Journal of Physiology. Renal Physiology. 2002;283(4):F658–F662. doi: 10.1152/ajprenal.00243.2001. [DOI] [PubMed] [Google Scholar]
- 56.Haas J. A., Khraibi A. A., Perrella M. A., Knox F. G. Role of renal interstitial hydrostatic pressure in natriuresis of systemic nitric oxide inhibition. The American Journal of Physiology. 1993;264(3):F411–F414. doi: 10.1152/ajprenal.1993.264.3.F411. [DOI] [PubMed] [Google Scholar]
- 57.Yang J., Chen C., Ren H., et al. Angiotensin II AT 2 receptor decreases AT 1 receptor expression and function via nitric oxide/cGMP/Sp1 in renal proximal tubule cells from Wistar-Kyoto rats. Journal of Hypertension. 2012;30(6):1176–1184. doi: 10.1097/HJH.0b013e3283532099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Wang T., Chan Y. L. Mechanism of angiotensin II action on proximal tubular transport. Journal of Pharmacology and Experimental Therapeutics. 1990;252(2):689–695. [PubMed] [Google Scholar]
- 59.Edwards R. M., Stack E. J. Angiotensin II inhibits glomerular adenylate cyclase via the angiotensin II receptor subtype 1 (AT1. Journal of Pharmacology and Experimental Therapeutics. 1993;266(2):506–510. [PubMed] [Google Scholar]
- 60.Schmieder R. E., Hilgers K. F., Schlaich M. P., Schmidt B. M. Renin-angiotensin system and cardiovascular risk. The Lancet. 2007;369(9568):1208–1219. doi: 10.1016/S0140-6736(07)60242-6. [DOI] [PubMed] [Google Scholar]
- 61.de Gasparo M., Catt K. J., Inagami T., Wright J. W., Unger T. International union of pharmacology. XXIII. The angiotensin II receptors. Pharmacological Reviews. 2000;52(3):415–472. [PubMed] [Google Scholar]
- 62.Gohlke P., Pees C., Unger T. AT2 receptor stimulation increases aortic cyclic GMP in SHRSP by a kinin-dependent mechanism. Hypertension. 1998;31(1):349–355. doi: 10.1161/01.HYP.31.1.349. [DOI] [PubMed] [Google Scholar]
- 63.Saito S., Hirata Y., Emori T., Imai T., Marumo F. Angiotensin II activates endothelial constitutive nitric oxide synthase via AT1 receptors. Hypertension Research. 1996;19(3):201–206. doi: 10.1291/hypres.19.201. [DOI] [PubMed] [Google Scholar]
- 64.Mollnau H., Wendt M., Szöcs K., et al. Effects of angiotensin II infusion on the expression and function of NAD(P)H oxidase and components of nitric oxide/cGMP signaling. Circulation Research. 2002;90(4):E58–E65. doi: 10.1161/01.res.0000012569.55432.02. [DOI] [PubMed] [Google Scholar]
- 65.Zheng Y., Horita S., Hara C., et al. Biphasic regulation of renal proximal bicarbonate absorption by luminal AT1A receptor. Journal of the American Society of Nephrology. 2003;14(5):1116–1122. doi: 10.1097/01.ASN.0000064700.58048.C1. [DOI] [PubMed] [Google Scholar]
- 66.Carey R. M., Jin X.-H., Wang Z.-Q., Siragy H. M. Nitric oxide: a physiological mediator of the type 2 (AT2) angiotensin receptor. Acta Physiologica Scandinavica. 2000;168(1):65–71. doi: 10.1046/j.1365-201x.2000.00660.x. [DOI] [PubMed] [Google Scholar]
- 67.Haithcock D., et al. Renal proximal tubular AT2 receptor: signaling and transport. Journal of the American Society of Nephrology. 1999;10(supplement 11):S69–S74. [PubMed] [Google Scholar]
- 68.Li Y., Yamada H., Kita Y., et al. Arachidonic acid metabolites inhibit the stimulatory effect of angiotensin II in renal proximal tubules. Hypertension Research. 2008;31(12):2155–2164. doi: 10.1291/hypres.31.2155. [DOI] [PubMed] [Google Scholar]
- 69.Li Y., Yamada H., Kita Y., et al. Roles of ERK and cPLA2 in the angiotensin II-mediated biphasic regulation of Na+-HCO3− transport. Journal of the American Society of Nephrology. 2008;19(2):252–259. doi: 10.1681/asn.2007030289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Shah S., Hussain T. Enhanced angiotensin II-induced activation of Na+, K +-ATPase in the proximal tubules of obese Zucker rats. Clinical and Experimental Hypertension. 2006;28(1):29–40. doi: 10.1080/10641960500386650. [DOI] [PubMed] [Google Scholar]
- 71.Becker M., Umrani D., Lokhandwala M. F., Hussain T. Increased renal angiotensin II AT1 receptor function in obese Zucker rat. Clinical and Experimental Hypertension. 2003;25(1):35–47. doi: 10.1081/CEH-120017739. [DOI] [PubMed] [Google Scholar]
- 72.Bharatula M., Hussain T., Lokhandwala M. F. Angiotensin II AT1 receptor/signaling mechanisms in the biphasic effect of the peptide on proximal tubular Na+,K+-ATPase, proximal tubules, kidney. Clinical and Experimental Hypertension. 1998;20(4):465–480. doi: 10.3109/10641969809053225. [DOI] [PubMed] [Google Scholar]
- 73.Guzman N. J., Fang M.-Z., Tang S.-S., Ingelfinger J. R., Garg L. C. Autocrine inhibition of Na+/K+-ATPase by nitric oxide in mouse proximal tubule epithelial cells. The Journal of Clinical Investigation. 1995;95(5):2083–2088. doi: 10.1172/JCI117895. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Liang M., Knox F. G. Nitric oxide activates PKCalpha and inhibits Na+-K+-ATPase in opossum kidney cells. The American Journal of Physiology. 1999;277(6):F859–F865. doi: 10.1152/ajprenal.1999.277.6.F859. [DOI] [PubMed] [Google Scholar]
- 75.Liang M., Knox F. G. Nitric oxide reduces the molecular activity of Na+,K+-ATPase in opossum kidney cells. Kidney International. 1999;56(2):627–634. doi: 10.1046/j.1523-1755.1999.00583.x. [DOI] [PubMed] [Google Scholar]
- 76.Zhang C., Mayeux P. R. NO/cGMP signaling modulates regulation of Na+-K+-ATPase activity by angiotensin II in rat proximal tubules. American Journal of Physiology-Renal Physiology. 2001;280(3):F474–F479. doi: 10.1152/ajprenal.2001.280.3.F474. [DOI] [PubMed] [Google Scholar]
- 77.Liang M., Knox F. G. Production and functional roles of nitric oxide in the proximal tubule. American Journal of Physiology. Regulatory, Integrative and Comparative Physiology. 2000;278(5):R1117–R1124. doi: 10.1152/ajpregu.2000.278.5.R1117. [DOI] [PubMed] [Google Scholar]
- 78.Azuma K. K., Balkovetz D. F., Magyar C. E., et al. Renal Na+/H+ exchanger isoforms and their regulation by thyroid hormone. The American Journal of Physiology. 1996;270(2):C585–C592. doi: 10.1152/ajpcell.1996.270.2.C585. [DOI] [PubMed] [Google Scholar]
- 79.Biemesderfer D., Pizzonia J., Abu-Alfa A., et al. NHE3: A Na+/H+ exchanger isoform of renal brush border. American Journal of Physiology. 1993;265(5):F736–F742. doi: 10.1152/ajprenal.1993.265.5.F736. [DOI] [PubMed] [Google Scholar]
- 80.Bobulescu I. A., Moe O. W. Luminal Na+/H+ exchange in the proximal tubule. Pflügers Archiv - European Journal of Physiology. 2009;458(1):5–21. doi: 10.1007/s00424-008-0595-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Gill R. K., Saksena S., Syed I. A., et al. Regulation of NHE3 by nitric oxide in Caco-2 cells. American Journal of Physiology-Gastrointestinal and Liver Physiology. 2002;283(3):G747–G756. doi: 10.1152/ajpgi.00294.2001. [DOI] [PubMed] [Google Scholar]
- 82.Stoos B. A., Garvin J. L. Actions of nitric oxide on renal epithelial transport. Clinical and Experimental Pharmacology and Physiology. 1997;24(8):591–594. doi: 10.1111/j.1440-1681.1997.tb02097.x. [DOI] [PubMed] [Google Scholar]
- 83.Roczniak A., Burns K. D. Nitric oxide stimulates guanylate cyclase and regulates sodium transport in rabbit proximal tubule. American Journal of Physiology-Endocrinology and Metabolism. 1996;270(1):F106–F115. doi: 10.1152/ajprenal.1996.270.1.F106. [DOI] [PubMed] [Google Scholar]
- 84.Kim J. S., Choi K. C., Jeong M. H., Kim S. W., Oh Y., Lee J. Increased expression of sodium transporters in rats chronically inhibited of nitric oxide synthesis. Journal of Korean Medical Science. 2006;21(1):1–4. doi: 10.3346/jkms.2006.21.1.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Jourdain M., Amiel C., Friedlander G. Modulation of Na-H exchange activity by angiotensin II in opossum kidney cells. American Journal of Physiology-Cell Physiology. 1992;263(6):C1141–C1146. doi: 10.1152/ajpcell.1992.263.6.C1141. [DOI] [PubMed] [Google Scholar]
- 86.Geibel J., Giebisch G., Boron W. F. Angiotensin II stimulates both Na+-H+ exchange and Na+/HCO3- cotransport in the rabbit proximal tubule. Proceedings of the National Acadamy of Sciences of the United States of America. 1990;87(20):7917–7920. doi: 10.1073/pnas.87.20.7917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Du Cheyron D., Chalumeau C., Defontaine N., et al. Angiotensin II stimulates NHE3 activity by exocytic insertion of the transporter: Role of PI 3-kinase. Kidney International. 2003;64(3):939–949. doi: 10.1046/j.1523-1755.2003.00189.x. [DOI] [PubMed] [Google Scholar]
- 88.Liu F.-Y., Cogan M. G. Role of protein kinase C in proximal bicarbonate absorption and angiotensin signaling. American Journal of Physiology - Renal Fluid and Electrolyte Physiology. 1990;258(4):F927–F933. doi: 10.1152/ajprenal.1990.258.4.F927. [DOI] [PubMed] [Google Scholar]
- 89.Weinman E. J., Steplock D., Shenolikar S. Acute regulation of NHE3 by protein kinase A requires a multiprotein signal complex. Kidney International. 2001;60(2):450–454. doi: 10.1046/j.1523-1755.2001.060002450.x. [DOI] [PubMed] [Google Scholar]
- 90.Alper S. L. Genetic diseases of acid-base transporters. Annual Review of Physiology. 2002;64:899–923. doi: 10.1146/annurev.physiol.64.092801.141759. [DOI] [PubMed] [Google Scholar]
- 91.Romero M. F., Hediger M. A., Boulpaep E. L., Boron W. F. Expression cloning and characterization of a renal electrogenic Na+/HCO3- cotransporter. Nature. 1997;387(6631):409–413. doi: 10.1038/387409a0. [DOI] [PubMed] [Google Scholar]
- 92.Shirai A., Yamazaki O., Horita S., et al. Angiotensin II dose-dependently stimulates human renal proximal tubule transport by the nitric oxide/guanosine 3′, 5′-cyclic monophosphate pathway. Journal of the American Society of Nephrology. 2014;25(7):1523–1532. doi: 10.1681/ASN.2013060596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.White W. B., Halley S. E. Comparative renal effects of intravenous administration of fenoldopam mesylate and sodium nitroprusside in patients with severe hypertension. JAMA Internal Medicine. 1989;149(4):870–874. doi: 10.1001/archinte.1989.00390040080016. [DOI] [PubMed] [Google Scholar]
- 94.Parker J. D., Farrell B., Fenton T., Cohanim M., Parker J. O. Counter-regulatory responses to continuous and intermittent therapy with nitroglycerin. Circulation. 1991;84(6):2336–2345. doi: 10.1161/01.CIR.84.6.2336. [DOI] [PubMed] [Google Scholar]
- 95.Rosenbaek J. B., et al. Effect of sodium nitrite on renal function, sodium and water excretion and brachial and central blood pressure in healthy subjects. A dose-response study. American Journal of Physiology. Renal Physiology. 2017 doi: 10.1152/ajprenal.00400.2016. [DOI] [PubMed] [Google Scholar]
- 96.Navar L. G., Harrison-Bernard L. M., Wang C. T., Cervenka L., Mitchell K. D. Concentrations and actions of intraluminal angiotensin II. Journal of the American Society of Nephrology. 1999;10(Supplement 11):S189–S195. [PubMed] [Google Scholar]
- 97.Seki G., Nakamura M., Suzuki M., Satoh N., Horita S. Species differences in regulation of renal proximal tubule transport by certain molecules. World Journal of Nephrology. 2015;4(2):307–312. doi: 10.5527/wjn.v4.i2.307. [DOI] [PMC free article] [PubMed] [Google Scholar]