

Abstract
Acquired thrombotic thrombocytopenic purpura is a rare and severe disease characterized by auto-antibodies directed against “A Disintegrin And Metalloproteinase with Thrombospondin type 1 repeats, 13th member" (ADAMTS13), a plasma protein involved in hemostasis. Involvement of CD4+ T cells in the pathogenesis of the disease is suggested by the IgG isotype of the antibodies. However, the nature of the CD4+ T-cell epitopes remains poorly characterized. Here, we determined the HLA-DR-restricted CD4+ T-cell epitopes of ADAMTS13. Candidate T-cell epitopes were predicted in silico and binding affinities were confirmed in competitive enzyme-linked immunosorbent assays. ADAMTS13-reactive CD4+ T-cell hybridomas were generated following immunization of HLA-DR1 transgenic mice (Sure-L1 strain) and used to screen the candidate epitopes. We identified the ADAMTS131239–1253 peptide as the single immunodominant HLA-DR1-restricted CD4+ T-cell epitope. This peptide is located in the CUB2 domain of ADAMTS13. It was processed by dendritic cells, stimulated CD4+ T cells from Sure-L1 mice and was recognized by CD4+ T cells from an HLA-DR1-positive patient with acute thrombotic thrombocytopenic purpura. Interestingly, the ADAMTS131239–1253 peptide demonstrated promiscuity towards HLA-DR11 and HLA-DR15. Our work paves the way towards the characterization of the ADAMTS13-specific CD4+ T-cell response in patients with thrombotic thrombocytopenic purpura using ADAMTS131239–1253-loaded HLA-DR tetramers.
Introduction
Thrombotic thrombocytopenic purpura (TTP) is a rare and severe autoimmune disease characterized by the occurrence of IgG autoantibodies against the metalloprotease “A Disintegrin And Metalloproteinase with Thrombospondin type 1 repeats, 13th member” (ADAMTS13).1–3 ADAMTS13 cleaves multimers of von Willebrand factor, a glycoprotein involved in hemostasis. Inhibition of ADAMTS13 by IgG leads to an accumulation of hyper-adhesive von Willebrand factor multimers causing microthrombi that occlude the lumen of the capillaries in the microcirculation, thus inducing red cell hemolysis and ischemia of downstream organs. TTP is thus characterized by a combination of microangiopathic hemolytic anemia, peripheral thrombocytopenia and organ failure of variable severity with typically neurological involvement.4
The physiopathological mechanisms underlying TTP and responsible for the synthesis of anti-ADAMTS13 antibodies, and particularly the mechanisms involved in the loss of tolerance of the immune system towards ADAMTS13, are poorly understood. Polyclonal anti-ADAMTS13 antibodies are directed against different domains of ADAMTS13.5 In most patients, anti-ADAMTS13 antibodies are of the IgG isotype with a predominance of the IgG4 subclass.6 IgG from all patients recognize immunodominant B-cell epitopes located in the spacer domain of ADAMTS13.7 The B-cell epitopes have been proposed to be located between the 660–661 and 665 amino-acids.8 The fact that anti-ADAMTS13 antibodies are of the IgG isotype, of high affinity and have undergone affinity maturation, strongly suggests the requirement of CD4+ T-cell help in the development of the disease.9 Besides, the HLA-DRB1*11 (DR11) haplotype was independently identified as a strong risk factor by three research groups.10–12 However, while CD4+ T cells are thought to play a major role, the specificity and the properties of the CD4+ T lymphocytes involved in the pathogenesis of TTP have not been studied. Importantly, the HLA restriction hints at the existence of immunodominant peptides in ADAMTS13.
Naïve CD4+ T-cell activation is initiated by the interaction of the T-cell receptor (TCR) with a peptide/MHC class II complex on professional antigen-presenting cells. Extracellular antigens are endocytosed, degraded into peptides in the early endosome and loaded onto MHC class II αβ heterodimer molecules. Sorvillo et al. identified, through mass spectrometry, ADAMTS13-derived peptides that are localized within the HLA-DR molecules expressed by dendritic cells derived from human monocytes (Mo-DC) and incubated with ADAMTS13 in vitro.13 The identified peptides belonged mainly to the CUB1 and CUB2 domains of ADAMTS13. The core sequence FINVAPHAR from the CUB2 domain was the only sequence detected for six donors with the DR11 haplotype among the 17 donors included in the study. In addition, the same group recently demonstrated that a peptide containing the FINVAPHAR amino-acid sequence is able to stimulate CD4+ T cells from an HLA-DR11 TTP patient.14 These studies did not, however, investigate the recognition of other peptides by CD4+ T cells. To address this issue, we focused on the HLA-DR1 haplotype for which a humanized transgenic HLA-DR1 mouse is available, and investigated the CD4+ T-cell epitopes for the HLA-DR1 allele. The identified epitopes were then studied in the context of the HLA-DR11 allele.
Methods
Peptides and antigen
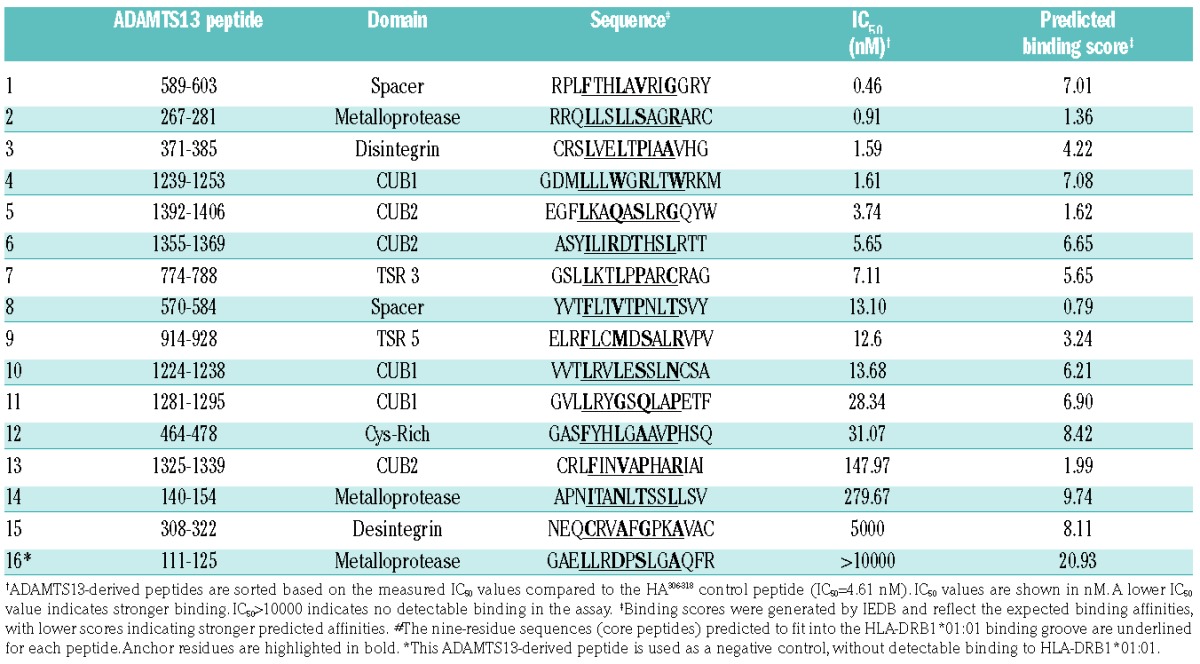
ADAMTS13-derived peptides were selected based on their capacity to bind to HLA-DRB1*01:01 molecules. HLA class II–binding predictions of human ADAMTS13 (GenBank reference: AAL11095.1) were made on November 10, 2011, using the Immune Epitope Database analysis resource consensus tool, on the dedicated website www.iedb.org.15–17 Strong HLA-DRB1*01:01 binders were predicted in silico using overlapping 15-mer peptides that span the whole ADAMTS13 sequence. Altogether, 99 15-mer peptides were predicted to be strong binders to HLA-DRB1*01:01 with binding scores below 10 (i.e., with a probability of being good binders greater than 90%). Some of the predicted peptides shared common HLA-DRB1*01:01-binding core sequences (9-mer peptides). When considering only unique core sequences and after exclusion of two peptides located in the prodomain of ADAMTS13, the list came down to 15 9-mer core peptides (Table 1). The peptides were synthesized at greater than 80% purity (GL Biochem, Shanghai, China) and included the 9-mer core sequences with addition of the three residues from the N-terminal end and the three residues of the C-terminal end. Individual peptides were solubilized at 1 mg/mL in dimethylsulfoxide/water. Recombinant full-length human ADAMTS13 (rhADAMTS13) was a kind gift from Baxter (Vienna, Austria).18
Table 1.
Affinity of ADAMTS13-derived peptides for HLA-DRB1*01:01 molecules.
HLA-peptide-binding assays
HLA-DR molecules were purified from homozygous Epstein-Barr virus cell lines by affinity-chromatography using the monomorphic monoclonal antibody L243. The binding to HLA-DR molecules was assessed by competitive enzyme-linked immunosorbent assay (ELISA), using an automated workstation, as previously reported.19,20 Briefly, HLA heterodimers were incubated with a biotinylated indicator peptide and serial dilutions of competitor peptides. As reference, the unlabeled form of biotinylated reporter peptide was used as an internal control. After 24 h incubation at 37°C, samples were neutralized with 450 mM Tris HCl (pH 7.5) (Sigma, St Quentin-Fallavier, France), 0.3% bovine serum albumin (Sigma), and 1 mM n-dodecyl β-D-maltoside buffer (Sigma) and applied to 96-well MaxiSorp ELISA plates (Nunc A/S, Roskilde, Denmark) coated with 10 μg/mL L243. Bound biotinylated peptide was detected by streptavidin-alkaline phosphatase conjugate (GE Healthcare, Saclay, France) after adding 4-methylumbelliferyl phosphate substrate (Sigma). Emitted fluorescence was measured at 450 nm upon excitation at 365 nm. The peptide concentration that prevented binding of 50% of the biotinylated peptide (IC50) was evaluated. The sequence of the biotinylated reporter hemagglutinin (HA) peptide was 306PKYVKQNTLKLAT318; the mean IC50 values of the HA306–318 peptide for HLA DRB1*01:01 and DRB1*11:01 binding were 4.61 and 37.42 nM, respectively. For HLA DRB1*15:01, the sequence of the reporter biotinylated peptide A3152–166 was 152EAEQLRAYLDGTGVE166 with an IC50 value of 41.46 nM. Means and standard errors of the means were calculated from three independent experiments. Binding data are estimated as relative affinities defined as the ratio of the IC50 of each individual peptide to the IC50 of the reference peptide. Peptides with relative affinities of 20 or less were considered as strong binders to the HLA-DRB1 molecules and were retained for the rest of the study.
Human samples
Heparinized blood from healthy donors (EFS, Ile-de-France, Créteil, France) or from TTP patients (French national reference center for TMA, Prof. P. Coppo, St Antoine Hospital, Paris, France and Dr L. Galicier, St Louis Hospital, Paris, France) was obtained after written informed consent. The use of samples for research purposes had been approved by the ethical committee from Saint-Antoine Hospital (Authorization n. 04807/12/2005). The criteria for diagnosing acquired TTP were those previously reported,4 and included a severe deficiency (<5% of normal activity) of ADAMTS-13 activity along with a detectable inhibitor and/or detectable serum antibodies as assessed by ELISA (Technozym R ADAMTS13 INH, Technoclone, Vienna, Austria; threshold positivity >15 U/mL).
Blood processing
All blood samples were shipped at room temperature within 4 h of being drawn. HLA-DR screening was performed by molecular typing using the Olerup SSP HLA*DR kit (Olerup SSP, Stockholm, Sweden) after extracting DNA from total blood (Qiagen, Vienna, Austria). Peripheral blood mononuclear cells were isolated by density gradient centrifugation on lymphocyte separation medium (PAA Laboratories GmbH, Les Mureaux, France) and immediately frozen in pooled human male AB serum containing 10% dimethylsulfoxide (Sigma).
Immunization of mice
The animals used in this study were HLA-A2.1-/HLA-DR1-transgenic H-2 class I-/class II (IA β−/− β2m−/−)-knockout mice (Sure-L1 mice), 8 to 12 weeks old.21 The mice were immunized subcutaneously with 150 μg of rhADAMTS13 in complete Freund’s adjuvant (Sigma), and 15 days later with 150 μg rhADAMTS13 in incomplete Freund’s adjuvant (Sigma). Blood was drawn by retro-orbital bleeding 4 days after the second administration of rhADAMTS13 and cells from the spleen and draining lymph nodes were collected. Animals were handled in agreement with local ethical authorities (Comité Régional d’Éthique p3/2005/002).
Titration of anti-ADAMTS13 IgG
Antibodies against human ADAMTS13 were measured in mice serum using ELISA. Briefly, ELISA plates were coated with rhADAMTS13 (1 μg/mL) for 1 h at 37°C and subsequently blocked for an additional hour with phosphate-buffered saline -1% bovine serum albumin (PBS-BSA). Serum serially diluted in PBS-BSA (dilution factor: 1/3) was then incubated for 1 h at room temperature. Bound IgG was revealed using a secondary horseradish peroxidase-coupled goat F(ab′)2 anti-mouse IgG antibody (Southern Biotech, Birmingham, AL, USA) and substrate. The mouse monoclonal anti-human ADAMTS13 IgG monoclonal antibody 20A5 (HYCULT, Uden, NL) was used as a positive control.
HLA-DRB1*01:01-positive antigen-presenting cells
Three different sources of antigen-presenting cells were used for T-cell presentation assays: (i) purified splenocytes from Sure-L1 mice inactivated with mitomycin C (50 μg/mL, Sigma), (ii) immature Mo-DC from HLA-DRB1*01:01 healthy donors inactivated with mitomycin C (50 μg/mL), and (iii) artificial antigen-presenting cells derived from the NIH-3T3 cell line expressing HLA-DRB1*01:01 (AAPCDR1). To generate AAPCDR1, NIH-3T3 cells were transduced with the common HLA-DRα chain and the specific HLA-DRβ1*01:01 chain, and with the co-stimulatory and adhesion human molecules B7.1, ICAM-1 and LFA-3.22 AAPCDR1 does not produce murine interleukin-2 and does not, therefore, require inactivation with mitomycin C. Its ability to present and stimulate HLA-DRB1*01:01-restricted CD4+ T cells efficiently has been demonstrated.22 In the case of Mo-DC, human monocytes were isolated from peripheral blood mononuclear cells using anti-CD14 magnetic beads (Miltenyi Biotec, Bergisch Gladbach, Germany). Monocytes were cultured in RPMI-1640 medium supplemented with 10% human AB Serum (Sigma), L-glutamine and antibiotics (Life Technologies - Invitrogen, Carlsbad, CA, USA) in the presence of 500 IU human recombinant interleukin 4 (ImmunoTools, Friesoythe, Germany) and 1000 IU human recombinant granulocyte-macrophage colony-stimulating factor (ImmunoTools) per 106 cells. After 5 days of culture, the non-adherent immature Mo-DC-enriched fraction was harvested and the immature status was confirmed by the expression of the following surface phenotypic markers: CD40, CD80, CD86, CD83, CD1a and HLA-DR (Becton Dickinson, Le Pont de Claix, France), by flow cytometry as explained previously.23
Proliferation response to ADAMTS13-derived peptide
Cells from the spleen and draining lymph nodes from immunized Sure-L1 mice were pooled. Cells (105 cells/well) were incubated alone, with each individual synthetic ADAMTS13-derived peptide (10 μg/mL) or with a pool containing all the peptides (2.5 μg/mL of each peptide) in AIM-V medium (Life Technologies) for 4 days. Cell proliferation was determined by incorporation of 3H-thymidine (1 μCi per well, specific activity of 6.7 Ci/mmol) added during the final 16 h and measured as counts per minute (cpm) on a micro-β-counter (Perkin Elmer Applied Biosystems). Stimulation indices (SI) were calculated as the ratio between the cpm in the presence of a stimulus, and the average cpm of the same cells incubated alone.
Generation of mouse ADAMTS13-specific CD4+ T-cell hybridomas
Cells from draining lymph nodes of immunized Sure-L1 mice were stimulated for 72 h with rhADAMTS13 (1 μg/mL) and fused with the BWZ.36 fusion (TCR−/−) partner cell line24 (a kind gift from Prof N Shastri, University of California, Berkeley, CA, USA) using polyethylene glycol. ADAMTS13-specific CD4+ T-cell hybridomas were obtained after culture in HAT and HT selective media (both from Sigma). Specificity for ADAMTS13 was evaluated using a T-cell stimulation assay.
T-cell hybridoma stimulation assay
Mouse HLA-DR1-restricted CD4+ T-cell hybridomas (105 cells/well) were cultured in plates of 96 round-bottomed wells with 104 (ratio 1:10) DRB1*01:01 antigen-presenting cells (splenocytes or immature Mo-DC) in the presence of rhADAMTS13 (10 μg/mL) for 24 h at 37°C. Murine interleukin-2 secreted by activated T cells was measured in the supernatant using an ELISA kit according to the manufacturer’s protocol (eBioscience, Paris, France). The positive control was concanavalin A (Sigma) at a concentration of 1 μg/mL. For each hybridoma cell line growing in HAT media, the stimulation index was calculated. Hybridoma clones with an at least 2-fold increase in interleukin-2 release (ratio between ADAMTS13-stimulated cultures and control medium) were considered to be ADAMTS13-specific and were subsequently sub-cloned by limiting dilution (0.5 cells/well). Among ADAMTS13-specific T-cell hybridomas, screening for peptide specificity was realized with AAPCDR1 co-cultured for 24 h with peptides at 10 μg/mL, and interleukin-2 secretion by activated T-cell hybridomas was measured in the supernatant with the same ELISA kit.
Generation of ADAMTS13-specific human CD4+ T-cell lines
Using the accelerated co-cultured dendritic cell assay,25 peripheral blood mononuclear cells from a TTP patient were stimulated for 10 days with the pool of 16 ADAMTS13-derived peptides (2.5 μg/mL for each peptide). Briefly, cells (5×106 cells/mL) were thawed and cultured in AIM-V medium supplemented with 1000 U/mL of granulocyte-macrophage colony-stimulating factor and 500 U/mL of interleukin-4 (ImmunoTools). After 24 h (day 1), the pool of ADAMTS13-derived peptides was added along with tumor necrosis factor-α (1000 U/mL), interleukin-1β (10 ng/mL), interleukin-7 (0.5 ng/mL) (ImmunoTools) and prostaglandin E2 (1 μM; Merck Calbiochem, Saint-Quentin-en-Yvelines, France). On day 10, non-adherent cells were collected, washed, and CD4+ T lymphocytes were isolated by negative magnetic purification with a CD4+ T-cell isolation kit (Miltenyi). Cells were then incubated with irradiated DRB1*01 Mo-DC (45 Gy) from healthy donors (ratio 1:10), pulsed with the pool of ADAMTS13-derived peptides, in fresh AIM-V at 37°C for 6 h. Subsequently, CD4+ T cells reactive to ADAMTS13-derived peptides were isolated using the interferon (IFN)-γ secretion assay-cell detection kit according to the manufacturer’s protocol (Miltenyi). Cell lines were obtained after culture of the IFN-γ-secreting cells in AIM-V medium supplemented with 10% human AB serum, interleukin-2 (20 U/mL), interleukin-4 (5 ng/mL), anti-CD3 monoclonal antibody (OKT3, 30 ng/mL) (ImmunoTools) and irradiated (45 Gy) peripheral blood mononuclear cells from two unrelated donors. Cells were fed every 7 days with fresh cytokines. The stimulated CD4+ T cells were tested after ~3 weeks for peptide specificity by IFN-γ enzyme-linked immunospot (ELISPOT) assays after incubation with peptide-pulsed or non-pulsed antigen-presenting cells.
Enzyme-linked immunospot assays
The human IFN-γ ELISPOT assay was performed according to the manufacturer’s recommendations. Ninety-six–well polyvinylidine fluoride plates (Millipore-Merck) were pre-wetted with ethanol. The individual ADAMTS13-derived peptides (10 μg/mL), the peptide pool, or medium alone were plated in triplicate wells, into the plates that had been pre-coated overnight with the anti-IFN-γ capture antibody (U-CyTech, Utrecht, the Netherlands) and blocked with AIM-V plus 10% human serum. Irradiated AAPCDR1 (45 Gy) were then seeded and incubated (3×104/well) with the CD4+ T-cell line (3×105/well) for 24 h at 37°C in a CO2 incubator. Following removal of peripheral blood mononuclear cells, IFN-γ secretion was visualized with a biotin-conjugated anti–IFN-γ antibody (U-CyTech), alkaline phosphatase-conjugated ExtrAvidin, and Sigmafast 5-bromo-4-chloro-3-indolyl phosphate/nitro blue tetrazolium (BCIP/NBT) tablets (Sigma). Plates were then air dried and scanned on an ImmunoSpot Analyzer (CTL, Shaker Heights, OH, USA). Spot-forming cells were automatically calculated by ImmunoSpot Software using the SmartCount™ and Autogate™ functions.26 ELISPOT readouts are expressed as spot-forming cells/106 CD4+ T-cells. Peptides that generated at least twice the sport-forming cells over the control wells were defined as eliciting a positive response.
Results
Few ADAMTS13-derived peptides are strong binders to HLA-DRB1*01:01 molecules
We hypothesized that the T-cell epitopes of rhADAMTS13 correspond to ADAMTS13-derived peptides that bind stably to the groove of class II MHC molecules. The candidate peptides should thus bind with a high affinity to HLA-DR molecules. Using the IEDB consensus method prediction tool for MHC class II binding, we first predicted the array of ADAMTS13-derived 15-mer peptides that bind to the HLA-DRB1*01:01 (DR1) allele. Altogether, 15 15-mer peptides with unique core sequences were predicted to be strong binders to HLA-DR1 with binding scores below 10 (Table 1). The corresponding synthetic 15 peptides were used in binding assays to determine in vitro affinities for purified HLA-DRB1*01:01 molecules by competition ELISA. The calculated IC50 values ranged from 0 to 5000 nM (Table 1). Five peptides bound to DR1 with an IC50 below 4 nM, an affinity greater than that of the influenza HA306–318 control peptide (IC50 = 4.61 nM). Three peptides revealed a poor binding capacity with IC50 values 20-fold greater than that of the HA control peptide, despite a good IEDB-predicted binding score.
ADAMTS13-derived peptides activate ADAMTS13-specific mouse CD4+ T-cell hybridomas restricted to HLA-DRB1*01:01
To assess whether the ADAMTS13-derived HLA-DR1 binding peptides are T-cell epitopes and are presented to T cells by antigen-presenting cells, we generated ADAMTS13-specific CD4+ T-cell hybridomas restricted to the HLA-DR1 allele. Induction of an anti-rhADAMTS13 immune response in a humanized transgenic HLA-DR1 mouse was confirmed by ELISA (Figure 1A). Cells from the draining lymph nodes were fused to a TCR−/− partner (BWZ36) and 95 CD4+ T-cell hybridomas were selected. Among them, 26 hybridomas produced interleukin-2 following incubation with splenocytes from Sure-L1 mice in the presence of rhADAMTS13. The 26 hybridomas were subsequently sub-cloned.
Figure 1.

Identification of HLA-DR1-restricted T-cell epitopes using ADAMTS13-specific CD4+ T-cell hybridomas. (A) Anti-ADAMTS13 IgG titers in Sure-L1 mice immunized with rhADAMTS13. The serum from an HLA-DR1-transgenic Sure-L1 mouse immunized with rhADAMTS13 was incubated in serial dilutions on ADAMTS13-coated ELISA wells. ADAMTS13-bound IgG were detected using horseradish peroxidase-coupled polyclonal goat anti-mouse IgG and substrate. Results are expressed in arbitrary units as optical density measured at 492 nm. (B) Activation of ADAMTS13-specific CD4+ T-cell hybridomas. A representative ADAMTS13-specific CD4+ T-cell hybridoma (clone 2G10δ) was incubated with AAPCDR1 (10:1 ratio) and ADAMTS13-derived peptides as listed in Table 1. Stimulation indices represent the ratio of IL-2 secreted by the T cells measured by ELISA upon incubation with ADAMTS13 peptides over IL-2 secreted in the absence of peptide. Means±SD are from two independent experiments. (C) Delineation of the core peptide for ADAMTS13-specific CD4+ T-cell hybridomas. A representative ADAMTS13-specific CD4+ T-cell hybridoma (clone 2G10δ) was incubated with AAPCDR1 and overlapping 15-mer peptides spanning the 1235–1256 peptide sequence. Stimulation indices were assessed as explained above.
We next evaluated the activation of the T-cell hybridomas by the 15-mer peptides. The ADAMTS131239–1253 peptide was the only peptide to induce a significant secretion of interleukin-2 in all the hybridoma cell lines tested (6 of the 26 rhADAMTS13-reactive T-cell hybridomas; a representative result is shown for 1 T cell-hybridoma in Figure 1B). To delineate the T-cell epitope recognized by the T-cell hybridomas more precisely, overlapping 15-mer peptides that span amino acids 1235 to 1256 were synthesized and assessed for their capacity to activate one of the T-cell hybridomas (Clone 2G10δ). Significant T-cell activation was observed for peptides 1237–1251, 1238–1252, 1239–1253, 1240–1254 and 1241–1255 (Figure 1C). Conversely, peptides 1235–1249, 1236–1250 and 1242–1256 failed to activate the T-cell hybridoma. Similar results were obtained with another T-cell hybridoma (clone 1F5γ, data not shown). These results demonstrate that the minimal core peptide sequence recognized by the ADAMTS13-specific DRB1*01:01-restricted T-cell hybridomas is 1241MLLLWGRLTWR1251. Anchor residues for binding of peptide 1239–1353 to the HLA groove are highlighted in Table 1: L1, W4, R6 and W9.
The ADAMTS131239–1253 peptide is recognized by CD4+ T cells from humanized Sure-L1 HLA-DRB1*01:01 mice
We next asked whether other peptides than the one identified using the hybridoma technology may be recognized by polyclonal CD4+ T cells from Sure-L1 mice immunized with rhADAMTS13. Splenocytes and cells from the draining lymph nodes from three immunized Sure-L1 mice were stimulated with the individual ADAMTS13-derived peptides previously identified to bind to HLA-DRB1*01:01 with a high affinity. Significant cell proliferation was observed in the case of ADAMTS131239–1253 peptide (Figure 2A), confirming that this peptide is a dominant CD4+ T-cell epitope, at least in mice.
Figure 2.

The ADAMTS131239–1253 peptide is an immunodominant T-cell epitope both for Sure-L1 mice as well as for a patient with TTP. (A) Stimulation by ADAMTS13-derived peptides of spleen and lymph node cells from Sure-L1 mice immunized with rhADAMTS13. Cells (3×105/well) were incubated with each individual peptide (10 μg/mL), with the pool of peptides (Peptide mix) or with rhADAMTS13. Proliferation was assessed by incorporation of tritiated thymidine. Proliferation indices are defined as the ratio of cpm of stimulated cells versus cpm of control cells, and are expressed as means±SD for two Sure-L1 mice. (B) The ADAMTS131239–1253 peptide is processed and presented to T cells by human antigen-presenting cells. A representative ADAMTS13-specific CD4+ T-cell hybridoma (clone 1F5γ) was incubated with immature dendritic cells from a healthy HLA-DR1+ blood donor and with the ADAMTS131239–1253 peptide, with rhADAMTS13 or with the ADAMTS13111–125 peptide. Stimulation indices represent the ratio of secretion of IL-2 by T cells stimulated in the presence of antigen versus the secretion of IL-2 by T cells incubated with Mo-DC alone. Means±SD are from two independent experiments. Similar results were obtained with other ADAMTS13-CD4+ T-cell hybridoma clones (not shown). The peptide incubated alone or with DR15+ artificial antigen-presenting cells failed to activate the T cells (not shown). (C) ADAMTS13 epitope specificity of CD4+ T-cell lines from a patient with acquired TTP. CD4+ T-cell lines were generated after stimulation with pooled ADAMTS13-derived peptides as defined in Table 1. T cells (3×105/well) were then incubated with AAPCDR1 (3×104 cells/well) alone or pulsed with individual peptide (10 μg/mL), or with the peptide pool (Peptide mix). The number of cells producing interferon-γ was then assessed by ELISPOT and is expressed as the mean number of spot-forming cells (SFC) per million cells calculated in the case of peptide-stimulated T cells minus the SFC obtained in the case of unstimulated cells. Means±SD are from two independent experiments. *The 111–125 ADAMTS13-derived peptide that did not bind to HLA-DRB1*01:01 was used as a negative control.
The ADAMTS13 T-cell epitope identified in humanized Sure-L1 HLA-DRB1*01:01 mice is presented by human antigen-presenting cells
We then investigated whether the ADAMTS131239–1253 peptide is generated by professional human antigen-presenting cells and presented to CD4+ T cells in an HLA-DRB1*01:01 context. To this end, immature HLA-DRB1*01:01 Mo-DC were incubated with rhADAMTS13 and with T-cell hybridomas of known specificity for the 1239–1253 peptide. T cells incubated with Mo-DC in the presence of the ADAMTS131239–1253 peptide presented with a stimulation index of 22.2±1.9 (Figure 2B). In contrast, incubation of the cells with the ADAMTS13111–125 control peptide did not result in T-cell activation (stimulation index: 1.1±0.0). Interestingly, incubation of T cells and Mo-DC with rhADAMTS13 induced T-cell activation with a stimulation index of 3.3±0.6, indicating that rhADAMTS13 is efficiently processed by human antigen-presenting cells, and that the ADAMTS131239–1253 peptide is readily generated.
The ADAMTS131239–1253 peptide is a dominant T-cell epitope in a patient with thrombotic thrombocytopenic purpura with the HLA-DRB1*01:01 allele
We then assessed whether the ADAMTS131239–1253 peptide is recognized by CD4+ T cells from patients with acquired TTP. CD4+ T-lymphocytes were isolated from the blood of a 43-year -old HLA-DRB1*01:01 male, 14 days after the onset of acute acquired TTP. ADAMTS13 activity in this patient was less than 5% of the normal value due to the presence of inhibitory antibodies directed to ADAMTS13. A CD4+ T-cell line specific for ADAMTS13-derived peptides was amplified from the patient’s peripheral blood mononuclear cells as explained. After 32 days of culture, the CD4+ T-cell line produced IFN-γ, as assessed by ELISPOT, following incubation with Mo-DC from HLA-DR1 healthy donors, pulsed with the pool of ADAMTS13-derived peptides with high binding affinity for HLA-DR1, but failed to produce IFN-γ when incubated with Mo-DC alone (data not shown). We then incubated the T-cell lines with AAPCDR1 and with each individual peptide. Together with the ADAMTS131392–1406 peptide, the ADAMTS131239–1253 was the only peptide able to activate human IFN-γ-producing CD4+ T cells (Figure 2C). Similar results were obtained when the T cells and ADAMTS131239–1253 peptide were incubated in the presence of HLA-DR1 Mo-DC (data not shown). Further evaluation of the ADAMTS131239–1253 peptide by IEDB predicted good binding scores to HLA-DR11, 15, 13, 8, 3 and 4, which was confirmed in competitive ELISA at least in the case of HLA-DR11 and HLA-DR15 (Table 2). T-cell lines were also generated from one female TTP patient in acute phase (aged 19, Figure 3A), one female TTP patient in remission phase (aged 34, Figure 3B) and one 25-year old, healthy male donor (Figure 3C), all with the HLA-DRB1*11:01 allele. Reactivity towards the ADAMTS131239–1253 peptide was detected in the case of the three individuals. Together, our data suggest that the ADAMTS131239–1253 peptide is a dominant and promiscuous T-cell epitope in TTP patients.
Table 2.
Affinity of ADAMTS131239–1253 peptide for HLA-DRB1 molecules.
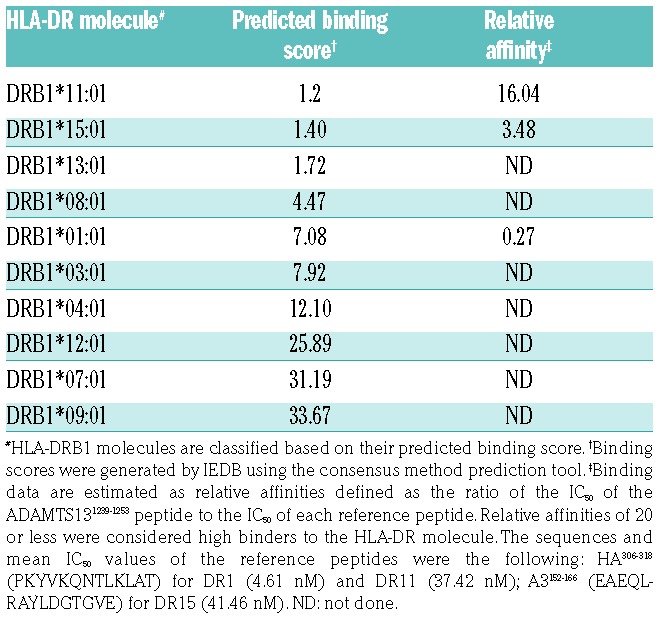
Figure 3.

The ADAMTS131239–1253 peptide is an immunodominant T-cell epitope for HLA-DRB1*11 individuals. CD4+ T-cell lines from HLA-DRB1*11 TTP patients at (A) acute phase or (B) in remission phase of the disease, or from (C) a HLA-DRB1*11 healthy donor, were stimulated with the ADAMTS13-derived peptides identified as immunodominant for HLA-DRB1*01. T cells (3×105 cells/well) were incubated with AAPCDR11 (3×104 cells/well) alone or pulsed with individual peptide (10 μg/mL), or with the peptide pool (Peptide mix). The number of cells producing interferon-γ was then assessed by ELISPOT and is expressed as mean number of spot-forming cells (SFC) per million cells calculated in the case of peptide-stimulated T-cells minus the SFC obtained in the case of unstimulated cells. *The 111–125 ADAMTS13-derived peptide is used as a negative control, without detectable binding to HLA-DRB1*11:01 (data not shown).
Discussion
In the present study, we delineated the ADAMT13-derived peptides that are potent T-cell epitopes in the context of the HLA-DR1 haplotype. An array of ADAMTS13-derived peptides that are processed and presented by Mo-DC had been previously reported in the case of healthy donors with different HLA-DR haplotypes.13 As many as 11 core peptides had thus been retrieved from the HLA-DR-binding groove; the peptides spanned six different domains of ADAMTS13. In order to identify HLA-DR-restricted T-cell epitopes of ADAMTS13, we took advantage of the humanized Sure-L1 mouse strain. Sure-L1 mice are invalidated for murine MHC class I and II molecules and transgenic for HLA-DRB1*01:01. HLA transgenic mice have been proven to be an appropriate human counterpart for the characterization of T-cell epitopes derived from pathogens21 as well as therapeutic proteins.27 In agreement with this, our results demonstrate that the ADAMTS131239–1253 peptide, identified as an immunodominant T-cell epitope using in silico prediction, in vitro binding assays and T-cell hybridoma generation, is processed by human Mo-DC in vitro and activates CD4+ T cells both in vivo in Sure-L1 mice, and ex vivo in the case of cells from patients with TTP.
The in silico selection of the ADAMTS13-derived peptides was based on the hypothesis that the T-cell epitope of rhADAMTS13 binds with a high affinity to HLA-DR molecules. The prediction efficiency of IEDB for HLA class II–binding peptides has been validated for various antigens, including human plasma proteins, and correlates adequately with HLA binding, as measured by competitive ELISA.15–17 For instance, T-cell epitopes of the human pro-coagulant factor VIII were identified using the hybridoma technology in an HLA-DR15 transgenic mouse model upon screening a peptide library spanning the entire sequence of the protein.27 Our unpublished data demonstrate a strong correlation between the predicted IEDB binding score for HLA-DR15 and the CD4+ T-cell epitope found in this work through hybridoma technology (area under the receiver operating characteristic curve=0.90). In silico prediction may nevertheless be over-predictive and positively select peptides that do not activate T cells.28
In the case of ADAMTS13, HLA-DR11 has been identified as a major risk factor for TTP. Sorvillo et al. identified 11 hypothetical T-cell epitopes following purification from HLA-DR molecules expressed by ADAMTS13-loaded Mo-DC from healthy donors with several HLA haplotypes.13 The ADAMTS13-derived core sequence 1328FINVAPHAR1336 was shown to be preferentially presented by the HLA-DR11 molecule,13 and stimulated CD4+ T cells from at least one TTP patient.14 In our hands, the ADAMTS131325–1339 peptide, that contains the same core sequence, was predicted to be a strong binder to HLA-DR1 peptide. However, it failed to bind HLA-DRB1*01:01 and to activate human CD4+ T cells from a DRB1*01:01 patient. Besides, the ADAMTS131325–1339 peptide did not activate CD4+ T cells from two TTP patients with the HLA-DRB11 allele. Whether this results from differences in peptide length and in the nature of the residues adjacent to the core peptide remains to be determined.29,30
Our rationale for focusing our study on the HLA-DRB1*01:01 allele is justified by the availability of an appropriate transgenic HLA-DRB1*01:01 mouse model, and by the fact that the HLA-DRB1*01:01 allele is found in 18% of the patients with acquired TTP10 and in 15 to 30% of the Caucasian population.31 Interestingly, 11 of the 15 peptides predicted to bind HLA-DR1 by IEDB were also predicted to have a high affinity for the HLA-DRB1*11:01 allele. In particular, the unique immunodominant HLA-DRB1*01:01-associated T-cell epitope characterized in our work, ADAMTS131239–1253, had an excellent binding score to HLA-DRB1*11:01 (score of 1.2) and HLA-DRB1*15:01, strong affinity for HLA-DRB1*11:01 and HLA-DRB1*15:01 in competitive ELISA (Table 2), and was a target epitope of CD4+ T cells from two TTP patients and one healthy donor with the HLA-DRB11 allele. Of note, the ADAMTS131240–1252 peptide, which shares the identical core peptide with the ADAMTS131239–1253 peptide, was identified by the study of Sorvillo et al. in the case of Mo-DC from a patient with the HLA-DR4/DR13 haplotype.13 Together with our IEDB prediction, the data suggest that the ADAMTS131239–1253 peptide exhibits promiscuity for at least three HLA-DR alleles: DR1, DR11 and DR15. Additional work including more HLA-DR alleles is required to determine whether pathogenic recognition of ADAMTS13 by CD4+ T cells is principally directed to promiscuous immunodominant epitopes such as the ADAMTS131239–1253 peptide, or whether private T-cell epitopes dominate in the case of HLA-DR alleles that are predominantly associated with acquired TTP.
We cannot exclude the existence of additional immunodominant T-cell epitopes potentially overlooked in our experimental approach. Besides, additional work is required to link our findings to the Th2 polarization of CD4+ T cells and production of anti-ADAMTS13 autoantibodies. Nevertheless, identification of the ADAMTS131239–1253 peptide as a promiscuous immunodominant T-cell epitope may have clinical implications at term. It should pave the way towards the characterization of the ADAMTS13-specific CD4+ T-cell response in TTP patients using ADAMTS131239–1253-loaded HLA-DR tetramers. Such a tool would provide new insights into the mechanisms involved in the breakdown of tolerance to ADAMTS13 and should be of clinical relevance for following, in a quantitative manner, populations of ADAMTS13-specific T cells in the course of TTP treatment and anticipate relapse. In particular, confirmation of our results would lead to identification of prognostic factors (persistence of specific Th17 T cells, generation of regulatory T cells) that would help in the decision process regarding pre-emptive use of rituximab to prevent relapse.32
Supplementary Material
Acknowledgments
We thank Professor John Robinson (Newcastle University, Newcastle upon Tyne, United Kingdom) for his advice about generating the ADAMTS13-specific T-cell hybridomas.
Footnotes
Check the online version for the most updated information on this article, online supplements, and information on authorship & disclosures: www.haematologica.org/content/102/11/1833
Funding
This study was supported by Institut National de la Santé et de la Recherche Médicale, Centre National de la Recherche Scientifique, Université Pierre et Marie Curie – Paris 6 and the 2012 Prof. Heimburger Award from CSL-Behring (Marburg, Germany). LG was the recipient of a « Poste d’accueil INSERM» fellowship and Bourse Marcel Simon - Société Française de Médecine Interne (SNFMI).
References
- 1.Tsai HM, Lian EC. Antibodies to von Willebrand factor-cleaving protease in acute thrombotic thrombocytopenic purpura. N Engl J Med. 1998;339(22):1585–1594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Furlan M, Robles R, Solenthaler M, et al. Acquired deficiency of von Willebrand factor-cleaving protease in a patient with thrombotic thrombocytopenic purpura. Blood. 1998;91(8):2839–2846. [PubMed] [Google Scholar]
- 3.Levy GG, Nichols WC, Lian EC, et al. Mutations in a member of the ADAMTS gene family cause thrombotic thrombocytopenic purpura. Nature. 2001;413(6855): 488–494. [DOI] [PubMed] [Google Scholar]
- 4.Coppo P, Bengoufa D, Veyradier A, et al. Severe ADAMTS13 deficiency in adult idiopathic thrombotic microangiopathies defines a subset of patients characterized by various autoimmune manifestations, lower platelet count, and mild renal involvement. Medicine (Baltimore). 2004;83(4):233–244. [DOI] [PubMed] [Google Scholar]
- 5.Zheng XL, Wu HM, Shang D, et al. Multiple domains of ADAMTS13 are targeted by autoantibodies against ADAMTS13 in patients with acquired idiopathic thrombotic thrombocytopenic purpura. Haematologica. 2010;95(9):1555–1562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Ferrari S, Palavra K, Gruber B, et al. Persistence of circulating ADAMTS13-specific immune complexes in patients with acquired thrombotic thrombocytopenic purpura. Haematologica. 2014;99(4):779–787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Thomas MR, de Groot R, Scully MA, et al. Pathogenicity of anti-ADAMTS13 autoantibodies in acquired thrombotic thrombocytopenic purpura. EBioMedicine 2015;2(8): 942–952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Pos W, Crawley JTB, Fijnheer R, et al. An autoantibody epitope comprising residues R660, Y661, and Y665 in the ADAMTS13 spacer domain identifies a binding site for the A2 domain of VWF. Blood. 2010;115(8):1640–1649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Pos W, Luken BM, Sorvillo N, et al. Humoral immune response to ADAMTS13 in acquired thrombotic thrombocytopenic purpura. J Thromb Haemost. 2011;9(7):1285–1291. [DOI] [PubMed] [Google Scholar]
- 10.Coppo P, Busson M, Veyradier A, et al. HLA-DRB1*11: a strong risk factor for acquired severe ADAMTS13 deficiency-related idiopathic thrombotic thrombocytopenic purpura in Caucasians. J Thromb Haemost JTH. 2010;8(4):856–859. [DOI] [PubMed] [Google Scholar]
- 11.Scully M, Brown J, Patel R, et al. Human leukocyte antigen association in idiopathic thrombotic thrombocytopenic purpura: evidence for an immunogenetic link. J Thromb Haemost. 2010;8(2):257–262. [DOI] [PubMed] [Google Scholar]
- 12.John M-L, Hitzler W, Scharrer I. The role of human leukocyte antigens as predisposing and/or protective factors in patients with idiopathic thrombotic thrombocytopenic purpura. Ann Hematol. 2012;91(4):507–510. [DOI] [PubMed] [Google Scholar]
- 13.Sorvillo N, van Haren SD, Kaijen PH, et al. Preferential HLA-DRB1*11-dependent presentation of CUB2-derived peptides by ADAMTS13-pulsed dendritic cells. Blood. 2013;121(17):3502–3510. [DOI] [PubMed] [Google Scholar]
- 14.Verbij FC, Turksma AW, de Heij F, et al. CD4+ T cells from patients with acquired thrombotic thrombocytopenic purpura recognize CUB2 domain-derived peptides. Blood. 2016;127(12):1606–1609. [DOI] [PubMed] [Google Scholar]
- 15.Wang P, Sidney J, Dow C, et al. A systematic assessment of MHC class II peptide binding predictions and evaluation of a consensus approach. PLoS Comput Biol. 2008;4(4): e1000048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Wang P, Sidney J, Kim Y, et al. Peptide binding predictions for HLA DR, DP and DQ molecules. BMC Bioinformatics. 2010;11:568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Vita R, Overton JA, Greenbaum JA, et al. The immune epitope database (IEDB) 3.0. Nucleic Acids Res. 2015;43(Database issue):D405–412. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Plaimauer B, Kremer Hovinga JA, Juno C, et al. Recombinant ADAMTS13 normalizes von Willebrand factor-cleaving activity in plasma of acquired TTP patients by overriding inhibitory antibodies. J Thromb Haemost. 2011;9(5):936–944. [DOI] [PubMed] [Google Scholar]
- 19.Texier C, Pouvelle-Moratille S, Busson M, et al. Complementarity and redundancy of the binding specificity of HLA-DRB1, -DRB3, -DRB4 and -DRB5 molecules. Eur J Immunol. 2001;31(6):1837–1846. [DOI] [PubMed] [Google Scholar]
- 20.Texier C, Pouvelle S, Busson M, et al. HLA-DR restricted peptide candidates for bee venom immunotherapy. J Immunol. 2000;164(6):3177–3184. [DOI] [PubMed] [Google Scholar]
- 21.Pajot A, Michel M-L, Fazilleau N, et al. A mouse model of human adaptive immune functions: HLA-A2.1-/HLA-DR1-transgenic H-2 class I-/class II-knockout mice. Eur J Immunol. 2004;34(11):3060–3069. [DOI] [PubMed] [Google Scholar]
- 22.Garnier A, Hamieh M, Drouet A, et al. Artificial antigen-presenting cells expressing HLA class II molecules as an effective tool for amplifying human specific memory CD4(+) T cells. Immunol Cell Biol. 2016;94(7):662–672. [DOI] [PubMed] [Google Scholar]
- 23.Bayry J, Lacroix-Desmazes S, Carbonneil C, et al. Inhibition of maturation and function of dendritic cells by intravenous immunoglobulin. Blood. 2003;101(2):758–765. [DOI] [PubMed] [Google Scholar]
- 24.Sanderson S, Shastri N. LacZ inducible, antigen/MHC-specific T cell hybrids. Int Immunol. 1994;6(3):369–376. [DOI] [PubMed] [Google Scholar]
- 25.Martinuzzi E, Afonso G, Gagnerault M-C, et al. acDCs enhance human antigen-specific T-cell responses. Blood. 2011;118(8):2128–2137. [DOI] [PubMed] [Google Scholar]
- 26.Lehmann PV. Image analysis and data management of ELISPOT assay results. Methods Mol Biol. 2005;302:117–32. [DOI] [PubMed] [Google Scholar]
- 27.Steinitz KN, van Helden PM, Binder B, et al. CD4+ T-cell epitopes associated with antibody responses after intravenously and subcutaneously applied human FVIII in humanized hemophilic E17 HLA-DRB1*1501 mice. Blood. 2012. 26;119(17):4073–4082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Lundegaard C, Lund O, Nielsen M. Predictions versus high-throughput experiments in T-cell epitope discovery: competition or synergy? Expert Rev Vaccines. 2012;11(1):43–54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Carson RT, Vignali KM, Woodland DL, et al. T cell receptor recognition of MHC class II-bound peptide flanking residues enhances immunogenicity and results in altered TCR V region usage. Immunity. 1997;7(3):387–399. [DOI] [PubMed] [Google Scholar]
- 30.Holland CJ, Cole DK, Godkin A. Re-Directing CD4(+) T cell responses with the flanking residues of MHC class II-bound peptides: the core is not enough. Front Immunol. 2013;4:172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.González-Galarza FF, Takeshita LYC, Santos EJM, et al. Allele frequency net 2015 update: new features for HLA epitopes, KIR and disease and HLA adverse drug reaction associations. Nucleic Acids Res. 2015;43(Database issue):D784–788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Hie M, Gay J, Galicier L, et al. Preemptive rituximab infusions after remission efficiently prevent relapses in acquired thrombotic thrombocytopenic purpura. Blood. 2014;124(2):204–210. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.