

Abstract
Background
The differentiation of pancreatic ductal adenocarcinoma (PDAC) could be associated with prognosis and may influence the choices of clinical management. No applicable methods could reliably predict the tumor differentiation preoperatively. Thus, the aim of this study was to compare the metabonomic profiling of pancreatic ductal adenocarcinoma with different differentiations and assess the feasibility of predicting tumor differentiations through metabonomic strategy based on nuclear magnetic resonance spectroscopy.
Methods
By implanting pancreatic cancer cell strains Panc-1, Bxpc-3 and SW1990 in nude mice in situ, we successfully established the orthotopic xenograft models of PDAC with different differentiations. The metabonomic profiling of serum from different PDAC was achieved and analyzed by using 1H nuclear magnetic resonance (NMR) spectroscopy combined with the multivariate statistical analysis. Then, the differential metabolites acquired were used for enrichment analysis of metabolic pathways to get a deep insight.
Results
An obvious metabonomic difference was demonstrated between all groups and the pattern recognition models were established successfully. The higher concentrations of amino acids, glycolytic and glutaminolytic participators in SW1990 and choline-contain metabolites in Panc-1 relative to other PDAC cells were demonstrated, which may be served as potential indicators for tumor differentiation. The metabolic pathways and differential metabolites identified in current study may be associated with specific pathways such as serine-glycine-one-carbon and glutaminolytic pathways, which can regulate tumorous proliferation and epigenetic regulation.
Conclusion
The NMR-based metabonomic strategy may be served as a non-invasive detection method for predicting tumor differentiation preoperatively.
Keywords: Pancreatic ductal adenocarcinoma, Nuclear magnetic resonance, Metabonomics, Tumor differentiation
Background
Pancreatic ductal adenocarcinoma (PDAC) is one of the most malignant tumors with an extremely poor prognosis. Only about 7% of patients can be survived in 5 years, making PDAC the fourth leading cause of death among tumors [1]. Many risk factors have been correlated with prognosis, including tumor size [2, 3], lymph node metastasis [3, 4], nerve plexus invasion [5, 6], vascular invasion [6, 7], tumor differentiation [2, 3, 8], surgical margin status [3, 9] and specific molecular prognostic factors [10, 11]. Thereinto, poorly differentiated/high grade tumors are closely associated with poor outcome of the patients [12]. Furthermore, previous researches also linked tumor histological grading to an increased risk of early death within 1 year [13, 14]. As an important component of early mortality risk score, tumor differentiation can help to assessing short-term tumor-related mortality [14, 15]. Given the important role of tumor differentiation in PDAC management, increased interest in preoperative tumor differentiation assessment were emerged in order to identify high-risk patients, which can benefit the most from neoadjuvant treatment [13, 16–19], even over than upfront surgery [20, 21]. Thus, notarizing differentiation of tumors preoperatively can provide constructive information for prognostic evaluation and management of PDAC [22].
Conventionally, the preoperative assessments of tumor differentiation were conducted by tissue histological observations derived from fine needle aspiration. This method has been realized to be an effective way to grade the pancreatic neuroendocrine tumors and intraductal papillary mucinous neoplasms [23, 24]. However, this technique is highly invasive for many patients and the achievable samples are too limited to give a reliable histological grading, making this technique still being far away from application in clinical PDAC differentiation assessment [19]. Thus, it would be of great importance to develop an easily acceptable and reliable method to assess the differentiation of PDAC preoperatively.
Nuclear magnetic resonance (NMR) spectroscopy-based metabonomic technique is a promising diagnostic tool with the advantages of high sensitivity, non-invasion and high throughput. This technique can analyze the disease-related metabonomic differences occurred in various types of biosamples (etc. tissues, body fluids and cells) to identify differential metabolites and further biomarkers contributed to establishment of recognition models for diagnosis. At present, NMR-based diagnostic strategy has demonstrated a favorable clinical performance in many diseases [25–31]. Particularly noticeable, magnetic resonance spectroscopy have also been recommended for diagnosis of brain, prostate and breast cancer in European cancer conference [29]. In addition, by using NMR-based methods, many reports on detecting PDAC in vivo or in vitro have showed an encouraging result to distinguish PDAC from not only the normal but also other benign lesions [32–35]. Therefore, in present study, we used 1H NMR spectroscopy to analyze serum metabonomes from PDAC mice models established by implantations of Panc-1, BxPC-3 and SW1990 (being poor, poor to moderate and moderate to well differentiated [36–39], respectively) cell strains on pancreas, thus, to assess the feasibility of this strategy in predicting the differentiation of tumor.
Methods
Cell culture and animals feeding
PDAC cell strains (Panc-1, BxPC-3 and SW1990, Catalog NO. SCSP-535, TCHu 12 and TCHu 201) were obtained from Shanghai Institute of Cell Biology, Chinese Academy of Sciences (Shanghai, China) authenticated with short tandem repeat test and mycoplasma culture. At the circumstance of 5% CO2 and 37 °C, these strains were incubated in dulbecco’s modified eagle medium (DMEM, Gibco, Thermo Fisher Scientific Inc., USA) added with 10% fetal bovine serum (Gibco) in cell incubator (3110, Thermo Scientific). Then, cells were digested by 0.125% trypsinogen (Life Technologies, Grand Island, NY, USA) for the passage with the ratio of 1:2-4 every 2-3 days. BALB/c nude mice (male, 4 weeks, weighing 18-20 g), purchased from Shanghai Slac laboratory animals Co., Ltd. (NO: SCXK (HU) 2012-0002), were bred in Fujian Medical University Animals Centre (Fuzhou, china) with a standard SPF-grade laboratory conditions.
Establishment of animal models
This experimental protocol was in accordance with the principles of National Institutes of Health guide for the care and use of laboratory animals and approved by Ethical Committee of Fujian Medical University. Three PDAC cell strains in the exponential phase were digested with 0.125% trypsinogen, washed by phosphate buffered saline (PBS) for three times, then collected and resuspended in PBS (1 × 107 cells per milliliter). After skin degerming, the cell suspension liquids were subcutaneously injected into the axilla of mice (one cell strain each mouse), followed by a month of normal feeding. The tumors with a size of 5 to 10 mm in diameter generated in the injected positions of mice. Consequently, the mice were executed by a mercy killing, and the tumor tissues of Panc-1, BxPC-3 and SW1990 were carefully collected and divided into pieces of 1 mm3 for implantation in situ.
Forty-five mice were randomly divided into 3 groups using random number table. Before surgery, all mice have a 12-h fasting without drink-deprivation. A 2-cm horizontal incision was made on the middle of abdominal wall to expose the pancreas. One piece of tumors was placed on the body or tail of pancreas and fixed with biogum (BaiYun medical glue Co., Ltd., Guangzhou, China), followed by carefully organ restoration and suture. Three groups were dealt with tumor tissues of Panc-1, BxPC-3 and SW1990, respectively (n = 15 for each).
Tissues samples collection and preparation
Thirty days after surgeries, 1 mL of blood from each group was collected by aortic puncture under continuous airway anesthesia of isoflurane (Jiupai pharmaceutical Co., Ltd., Shijiazhuang, China) and stored in clear 1.5-mL Eppendorf tubes. After standing for 30 min, the blood went through a 10-min centrifugation at 10,000 g and 4 °C. The supernate was collected and immediately frozen by liquid nitrogen and stored at −80 °C. For the detection of 1H NMR spectroscopy, 400 μL of serum were melted on the surface of ice, and then mixed with 200 μL of 90 mM deuterated phosphate buffer (NaH2PO4 and K2HPO4, pH 7.4). The mixture of serum and buffer were centrifuged again, and finally, 550 μL of the supernate was moved into 5-mm NMR tubes (ST500, NORELL, Inc., Morganton, North Carolina, USA).
Detection of 1H NMR spectroscopy and preprocessing
The 1H NMR spectroscopy of serum samples were performed on a Varian NMR system (Agilent Technologies Co, Palo Alto, California, USA) with a 500.13 MHz of proton frequency at the temperature of 298 K. For each sample, a water-suppressed CPMG (Carr-Purcell-Meiboom-Gill) spin-echo pulse sequence (RD-90°-(τ-180°-τ)n-ACQ) was used to acquire the NMR spectrum. Herein, a total of 64 scans with a spectral width of 6 KHz and a data point of 12 K were accumulated for all spectra. Spin-echo loop time (2nτ) of 70 ms was applied with a relaxation delay of 2.0 s. The NMR spectra were processed by using MestReNova (V9.0.1, Mestrelab Research S. L., Spain). In order to increase the signal-to-noise ratio, all free induction decays were multiplied by an exponential weighting function equivalent to a 1 Hz line-broadening and subsequently disposed with Fourier transformation. To make the spectra more comparable, the manual phase rectifications and baseline corrections were conducted by using MestReNova. The chemical shifts were referenced to the double-peak of endogenic lactate at δ1.33 for metabolites identification. Automatically, the spectral regions δ9.0-0.5 of the processed NMR spectra were segmented into scatter integral regions of 0.002 ppm with a removal of spectral region δ6.40-5.50 and δ5.19-4.36 to eliminate the impacts of residual water signal and urea signal, respectively. Finally, for each spectrum, the integrated data were normalized to the total sum of the spectrum in favour of multivariate statistical analysis.
Multivariate statistical analysis
The multivariate statistical analysis, including principal component analysis (PCA), partial least squares discriminant analysis (PLS-DA) and orthogonal partial least squares discriminant analysis (OPLS-DA), were performed in SIMCA-P+ (V14.0 Umetrics, Sweden) to analyze the metabonomic differences between three PDAC groups. PCA, performed in the approach of mean-centered scaling, could simplify the normalized date into several components, which can roughly evaluate the clusters distributions and identify the existence of outlines. PLS-DA and OPLS-DA, which can be classified as supervised multivariate statistical analysis, were conducted in the approach of parato-scaling approach for better extraction and maximization of the metabonomic differences between PDAC groups. Furthermore, the OPLS-DA models coefficients, which were back-calculated from the coefficients, incorporated with the weight of the variables, and then to be plotted with color-coded coefficients to enhance interpretability of the models. As a result, the metabolites responsible for the metabonomic differences between groups can be extracted from the corresponding color-coded loading plots and displayed visually. By assistance of MATLAB (V7.1, the Mathworks Inc., Natick, USA), the color-coded coefficient loading plots were drew and color-coded according to the absolute value of coefficient. That meant, in the loading plots, a warm-toned color (i.e. red) represents for the metabolites being positive or negative significant in distinguishing different groups while a cool-toned color (i.e. blue) corresponds to the metabolites not being significant in discriminations. Moreover, to screen out differential metabolites, the cutoff value of correlation coefficients (|r| > 0.576) was determined according to the statistical significance of the Pearson correlation coefficient test at the level of P < 0.05 and df (degree of freedom) =10. In order to assess the quality and validity of models, the 10-fold cross validation and response permutation testing (n = 200) were performed and the corresponding parameters R2 and Q2 in the permutated plots presented the degree of model fitting and the potentially predictive ability of models, respectively.
The metabolic pathways and interactions analysis
The differential metabolites derived from multivariate statistical analysis were further analyzed for the metabolic pathways by using KEGG (www.genome.jp/kegg) and MBROLE 1.0 (http://csbg.cnb.csic.es/mbrole/) [40, 41].
Results
NMR spectral profiles of serum samples from Panc-1, BxPC-3, SW1990 groups
After visual confirmation for tumorgenesis, 12, 13, and 11 serum samples from Panc-1, BxPC-3 and SW1990 groups were included for the detections with 1H NMR spectroscopy, respectively. Typical one-dimensional 500-MHz 1H NMR spectra of serum samples from models induced by the different differentiated PDAC cells are presented in Fig. 1, which provided an integrated overview of all metabolites. Forty-seven metabolites were identified from the NMR spectra (Table 1) based on the relative literatures and public databases [42, 43]. A certain degree of metabolic differences could be noticed between different PDAC groups visually such as ethanol and phosphocholine. But considering the high similarity of spectra, the metabonomic information acquired was quite limited and the multivariate statistic analysis will help to extract the detailed information.
Fig. 1.

Representative 500 MHz 1H CPMG NMR spectra of serum samples from pancreatic cancer mice induced by the different differentiated cells. The spectral regions of δ5.5-9.0 (in the dashed box) were magnified 20 times compared with the regions of δ0.0-5.5 for the purpose of clarity. The abbreviations for peak assignments were noted in Table 1
Table 1.
The metabolites assignments from NMR spectra of serum from PDAC micea
| Abbreviation | Metabolites | 1H chemical shift(multiplicity)b |
|---|---|---|
| 1-MH | 1-Methylhistidine | 7.06(s), 7.78(s) |
| 3-HB | 3-Hydroxybutyrate | 1.20(d), 2.31(dd), 2.40(m), 4.16(m) |
| Ace | Acetate | 1.92(s) |
| AA | Acetoacetate | 2.28(s) |
| Act | Acetone | 2.24(s) |
| Ala | Alanine | 1.48(d) |
| All | Allantoin | 5.39(s) |
| Bet | Betaine | 3.27(s), 3.90(s) |
| Cho | Choline | 3.20(s) |
| Cit | Citrate | 2.53(d), 2.67(d) |
| Cr | Creatine | 3.04(s), 3.93(s) |
| Eth | Ethanol | 1.18(t), 3.61(q) |
| For | Formate | 8.46(s) |
| Fum | Fumarate | 6.52(s) |
| Glu | Glutamate | 2.08(m), 2.11(m), 2.35(m), 3.75(t) |
| Gln | Glutamine | 2.14(m), 2.45(m), 3.75(t) |
| G | Glycerol | 3.55(m), 3.66(dd), 3.78(m) |
| GPC | Glycerolphosphocholine | 3.23(s), 4.33(m) |
| Gly | Glycine | 3.56(s) |
| His | Histidine | 7.08(s), 7.82(s) |
| HOD | Residual water signal | 4.76(br) |
| IB | Isobutyrate | 1.07(d) |
| Ile | Isoleucine | 0.94(t), 1.01(d) |
| L1 | LDL | 0.86(br), 1.28(br) |
| L2 | VLDL | 0.89(br), 1.30(br), 1.58(br) |
| L3 | Unsaturated fatty acid | 2.04(br), 2.24(br), 2.76(br), 5.31(br) |
| Lac | Lactate | 1.33(d), 4.11(q) |
| Leu | Leucine | 0.96(d) |
| Lys | Lysine | 1.46(m), 1.73(m), 1.91(m), 3.03(m), 3.76(t) |
| Mal | Malonate | 3.11(s) |
| Met | Methionine | 2.14(s), 2.63(t) |
| MG | Methylguanidine | 2.83(s), 3.36(s) |
| Mol | Methanol | 3.36(s) |
| m-I | myo-Inositol | 3.52(dd), 3.61(dd), 4.07(m) |
| NAG | N-acetyl glycoprotein | 2.03(s) |
| Phe | Phenylalanine | 7.32(d), 7.37(t), 7.42(dd) |
| PC | Phosphocholine | 3.21(s) |
| Py | Pyruvate | 2.37(s) |
| Suc | Succinate | 2.40(s) |
| Thr | Threonine | 1.33(d), 4.26(m) |
| TMA | Trimethylamine | 2.89(s) |
| Trp | Tryptophan | 7.27(m), 7.30(s), 7.54(d), 7.73(d) |
| Tyr | Tyrosine | 6.90(d), 7.19(d) |
| Urea | Urea | 5.80(br) |
| Val | Valine | 0.99(d), 1.04(d) |
| α-Glc | α-Glucose | 3.42(t), 3.54(dd), 3.71(t), 3.73(m), 3.84(m), 5.24(d) |
| β-Glc | β-Glucose | 3.24(ddb), 3.41(t), 3.46(m), 3.49(t), 3.90(dd), 4.65(d) |
a PDAC pancreatic ductal adenocarcinoma
bmultiplicity:s, singlet; d, doublet; t, triplet; q, quartet; dd, doublets; m, multiplet; br, broad resonance
Metabonomic characteristics of serum from the PDAC groups
To show an overview of 1H NMR data collected from the serum of Panc-1, BxPC-3, and SW1990 groups, the PCA and PLS-DA were performed. The PCA scores plot showed a certain degree of separated trends between the three PDAC groups (Fig. 2a) though a little overlap or dispersity was demonstrated, indicating their obvious metabonomic differences. In further, a greater discrimination in cluster distributions of Panc-1, Bxpc-3 and SW1990 could be observed visually in PLS-DA scores plot (Fig. 2b), demonstrating a significant differences with each other.
Fig. 2.

The PCA (a) and PLS-DA (b) scores plots based on 1H NMR data of serums from PDAC groups. P, Panc-1; B, BxPC-3; SW, SW1990
To get deep insight into the metabolites responsible for the metabonomic alterations occurred in three PDAC groups, pair-wise comparisons were conducted by using the PLS-DA combined with orthogonal projection (OPLS-DA). The pronounced separations were demonstrated in OPLS-DA scores plots (Fig. 3 upper left panels) and the metabolites corresponding to the metabolic difference were marked in loading plots (Fig. 3 bottom panels). The summarized dominant metabolites, based on the cutoff value of correlation coefficient (|r| > 0.576), and the correlation coefficients were listed in detail based on their biochemical types (Table 2). Overall, the levels of metabolites belonged to glycolysis and glutaminolysis, alcohols and amino acids were lower in SW1990 group while the high concentrations of choline and its derivatives were noticeable in Panc-1 group. The favorable fit and prediction parameters (R2 and Q2) of the OPLS-DA models and the corresponding permutation test and probability (p-value) via CV-ANOVA also confirmed the strong predictive ability of the models to guarantee a reliable identification of characteristic metabolites.
Fig. 3.

OPLS-DA scores plots (upper left panels) and plots of permutation tests (n = 200) (upper right panels) derived from 1H NMR spectra of serum samples and corresponding coefficient loading plots (bottom panels) from the pair-wise comparisons between Panc-1, Bxpc-3 and SW1990 groups. a. Panc-1 vs SW1990, b. BxPC-3 vs SW1990, c. Panc-1 vs BxPC-3. The color map shows the significance of metabolites variations between the two classes. Keys of the assignments were shown in Table 1. P, Panc-1; B, BxPC-3; SW, SW1990
Table 2.
OPLS-DA coefficients of metabolites in different pair-comparisons derived from NMR-data
| Metabolites | ra | ||
|---|---|---|---|
| BxPC-3 vs SW1990 | Panc-1 vs SW1990 | Panc-1 vs BxPC-3 | |
| Glycolysis and glutaminolysis | |||
| α-Glucose | −0.788 | −0.631 | – |
| β-Glucose | −0.735 | −0.842 | – |
| Citrate | 0.817 | 0.921 | – |
| Glutamate | 0.808 | 0.747 | −0.789 |
| Glutamine | 0.767 | 0.856 | – |
| Lactate | 0.906 | 0.905 | – |
| pyruvate | −0.880 | – | 0.793 |
| Succinate | – | – | – |
| Carboxylic acids and derivatives | |||
| Acetate | – | – | – |
| Formate | – | – | – |
| Fumarate | – | – | – |
| Isobutyrate | – | – | −0.709 |
| Malonate | – | – | −0.648 |
| Alcohols | |||
| Ethanol | 0.879 | – | −0.804 |
| Methanol | 0.702 | 0.760 | 0.667 |
| myo-Inositol | 0.889 | 0.817 | −0.877 |
| Glycerol | 0.935 | 0.784 | −0.916 |
| Lipid | |||
| LDL | −0.899 | −0.847 | 0.912 |
| VLDL | −0.774 | 0.720 | 0.921 |
| Unsaturated fatty acid | −0.899 | −0.847 | 0.912 |
| ketone body | |||
| 3-Hydroxybutyrate | 0.747 | – | −0.636 |
| Acetoacetate | – | – | – |
| Acetone | −0.760 | – | 0.912 |
| Choline and derivatives | |||
| Choline | – | −0.836 | −0.841 |
| Glycerolphosphocholine | 0.671 | −0.912 | −0.894 |
| Phosphocholine | 0.736 | −0.832 | −0.892 |
| Amino acid | |||
| Non-essential amino acid | |||
| 1-methylhistidine | – | – | −0.651 |
| Alanine | 0.750 | 0.778 | – |
| Betaine | 0.812 | 0.834 | −0.769 |
| Creatine | 0.930 | 0.826 | −0.849 |
| Glycine | 0.871 | 0.674 | – |
| Histidine | 0.776 | 0.602 | – |
| Tyrosine | 0.832 | 0.859 | – |
| Essential amino acid | |||
| Isoleucine | 0.749 | 0.795 | – |
| Leucine | 0.707 | 0.775 | – |
| Lysine | 0.886 | 0.822 | −0.780 |
| Methionine | – | 0.645 | – |
| Phenylalanine | 0.878 | 0.813 | −0.642 |
| Threonine | 0.630 | 0.794 | 0.730 |
| Tryptophan | 0.846 | 0.847 | −0.673 |
| Valine | 0.839 | 0.858 | – |
| Others | |||
| Methylguanidine | 0.650 | 0.732 | – |
| Allantoin | −0.687 | – | – |
| N-acetyl glycoprotein | – | 0.661 | 0.914 |
| Trimethylamine | −0.855 | 0.750 | 0.782 |
aCorrelation coefficients, positive and negative signs indicate positive and negative correlation in the concentrations. |r| > 0.576 was the cutoff value for significance based on discrimination significance of p = 0.05 and df = 10. “-” means |r| < 0.576
The biochemical pathways related with the metabonomic difference between PDAC groups
For better understanding of the bioinformation contained in discriminatory metabolites, the biochemical pathways were identified based on the differential metabolites derived from OPLS-DA of pair-comparisons and those with p-value less than 0.01 were demonstrated on Fig. 4. The p-value for pathway identification were calculated automatically by the MBROLE [40].
Fig. 4.
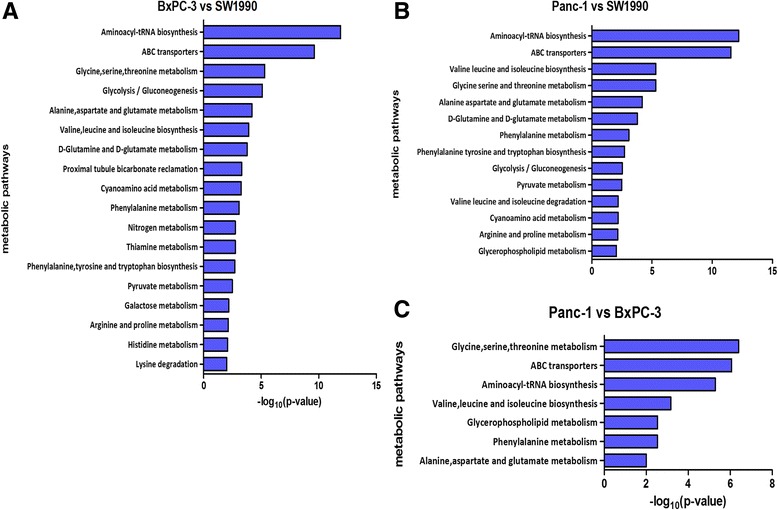
The corresponding pathways drived from the differential metabolites from different pair-comparisons. a BxPC-3 vs SW1990, b Panc-1 vs SW1990, c Panc-1 vs BxPC-3. This pathway analysis was performed in MBROLE online services based on KEGG database. The pathways with P-value <0.01(y-axes) and corresponding P-values (x-axes) in different pair-comparison were identified and listed
In the analysis to compare SW1990 with Bxpc-3, the numerous amino acid-related pathways were noticeable, including metabolism of essential and non-essential amino acids, the biosynthesis of aminoacyl-tRNA and ABC transporters. In addition, the pathways related with glycolysis involving pyruvate, galactose, glutamine and glutamate were also identified as differential features to distinguish the Bxpc-3 from the SW1990. Meanwhile, except the pathways of lysine, histidine and thiamine metabolisms, most pathways involved in Bxpc-3 vs SW1990 were also identified in the comparison between Panc-1 and Sw1990. In addition, the pathways of glycerophospholipid metabolism and the degradation of valine, leucine and isoleucine were also identified to be a signature contributed to distinguish Panc-1 from SW1990. In term of metabolic diversity between Panc-1 and BxPC-3, the metabolic discrimination seems to be quite limited where only a few pathways related with amino acids and glycerophospholipid metabolism were identified.
Discussion
In this study, we tried to evaluate the potential value of non-targeted NMR strategy to predict the tumor differentiation. Since many factors (e.g., drugs, operations) could influence the metabonomic characteristics of serum from patients. We chose three PDAC strains, Panc-1, BxPC-3 and SW1990 which can form tumors in vivo with typical histopathologic characters from poor, poor to moderate and moderate to well differentiation respectively [36–39] to establish PDAC models for research. By using animal models, the interference factors can be furthest eliminated. It is beneficial to purify serum metabonomic alteration caused by tumor with different differentiation and also specify the association between tumor differentiation and serum metabonomes. To amplify the metabolic difference between the tumors in different differentiations, all groups were compared directly. Given most of clinical patients were diagnosed with moderately differentiated PDAC and the significant clinical value for the identification of tumors in poor differentiation, we focus on the metabonomic difference between SW1990 and other two strains.
Comparative low levels of lactate, glutamate and glutamine indicate a poor differentiation
In present study, we found that the high concentration of citrate, lactate, glutamate and glutamine can help to distinguish the SW1990 from Panc-1 and Bxpc-3. Being well known, the tumor metabolic reprogramming has been validated to be the cornerstone for malignant transformation and one common composition in this process is the aerobic glycolysis (Warburg effect). Through the aerobic glycolysis, rather than tricarboxylic acid (TCA) cycle, the tumor cells derive the predominant ATP/energy and generate extensive lactate from pyruvate to result in environmental acidosis which promote the spreading of the tumor cells [44]. Meanwhile, the lactate generated from hypoxic PDAC can be taken up by normoxic PDAC cells nearby as fuel to maintain proliferation, creating a phenomenon called tumor symbiosis [45]. Thus, the tumor metabolic impact upon the level of lactate in peripheral circulation may be determined by the dynamic balance of release and uptake of lactate around tumor microenvironment. Our outcome indicates that the tumor with a poorer differentiation could induce a lower concentration of lactate in serum relative to that with a better differentiation, which may be due to a stronger ability of lactate recirculation. It’s also implied by inconsistent variation trends of lactate in serum reported by previous studies [46, 47]. In addition, due to the breakdown of TCA cycle, glutaminolysis is enhanced in PDAC cells to generate TCA intermediates (e.g. malate, oxaloacetate and citrate) which is called anaplerosis reaction, and subsequently served as building blocks for synthesis of lipid and non-essential amino acids [48]. Besides, glutamine can also act as fuel to support energy metabolism through aspartate, oxaloacetate and pyruvate transformation process, thus promoting growth of pancreatic cancer via Kras-regulated metabolic pathway [49]. Therefore, the significantly low levels of glutamine, glutamate and citrate may indicate that the tumor with poorer differentiation may provide a more dramatic glutaminolysis and deprive more glutamine and glutamate from peripheral circulation.
Comparative low levels of amino acids in serum imply poor differentiation
Likewise, the higher concentrations of amino acids could also contribute to the distinguishing of the SW1990 from Panc-1 and Bxpc-3, which could serve as key participants in the cancer metabolism reprogramming. Under the influence of the abnormal expression of oncogenes and tumor suppress genes, the anabolic metabolism and transport of amino acid were tremendously enhanced for rapid proliferation of cancer cells. To provide required nutrients for cancer growth, the catabolic metabolism of whole-body tissue would be enhanced, leading to an increased circulating amino acids at the early stage of PDAC [50]. But the catabolic metabolism cannot maintain in a high level for a long time and end in a severe nutritional imbalance called cachexia, thus creating a decrease of amino acids in serum at last. In this process, L-type amino-acid transporter 1 (LAT-1), the most important transporter of neutral amino acids, plays a key role in internalized transportation of essential amino acids (EAAs) in PDAC. As previous reports demonstrated, the overexpression of LAT-1 can promote cancer growth via mammalian target-of-rapamycin (mTOR) and serve as a prognostic factor in PDAC [51, 52]. Thus, the higher concentration of EAAs in SW1990 group than in Panc-1 and BxPC-3 group indicates that the tumors with poor differentiation may have a higher expression of LAT1 and nutritional stress from rapid proliferation, which can associated with poor prognosis.
With regard to the non-essential amino acids (NEAAs), several pathways were involved to enhance their biosynthesis and utilization for cell proliferation. As noted above, the accumulated glycolysis intermediates could also promote the biosynthesis of glycine, serine and threonine through 3-phospho-D-glycerate pathway. In addition, the increased glutaminolysis provides numerous substrates (e.g. isocitrate, malate, alpha-ketoglutaric acid) not only to supply the lipids synthesis but also to promote the biosynthesis of alanine and aspartate. Besides being used as building blocks and fuels for cell proliferation, NEAAs have been indicated to bridge the interplay metabolism and epigenetics, thus serve as programmed switch for cell differentiation [53]. For instance, several NEAAs including glycine could be associated with gene signatures of cell proliferation and Myc target activation through the serine-glycine-one-carbon pathway (SGOC pathway), which contribute significantly to energy generation and biosynthesis of NADPH and purine [54]. In addition, the mTOR-dependent induction of SGOC pathways can also lead to DNA methylation and tumorigenesis under the cooperatively oncogenic function of the loss of liver kinase B1 and activation of Kras, which highly involved in epigenetics [55]. Thus, NEAAs are highly associated with genesis, progression and epigenetics, and their relative concentration in serum may be indicators for the differentiation of PDAC.
Relative high concentration metabolites of choline metabolism may imply a poor differentiation
Impressively, the high correlation coefficient of choline groups in the pair-comparison of Panc-1 vs BxPC-3 and Panc-1 vs SW1990 implied that relatively high concentration of choline-like metabolites including phosphocholine (PC) and glycerolphosphocholine (GPC) may be significant metabolic features for poor differentiation of PDAC. According to previous study, the tumor-associated choline metabolism plays a key role in cell malignant transformation, tumor migration and metastasis [56, 57], characterized by elevated level of PC and total choline in tissue [45, 46]. Thereinto, the overexpression of choline kinase-α (Chk-a) induced by hypoxia-inducible factor (HIF) accounts for the increase of cellular PC and total choline [58], generating excessive phosphatidylcholine for biosynthesis of cell membrane. In addition, the EDI3-intermediated choline metabolism, a pathways verified in other solid tumor, can not only cleave GPC to form choline to supplement Kennedy pathway, but also generate glycerol-3-phosphate and its sequentially downstream intermediators for cellular signaling to regulate migration, invasion, proliferation and differentiation [57]. Other research detecting serum from PDAC patients also indicated that the choline metabolism were obviously altered and could potentially serve as biomarkers to detect PDAC in early stage. Thus, the difference of choline metabolism in PDAC could reflect and create a handful of regulatory functions on tumor progression and differentiation.
There are some pitfalls in this study. The PDAC models were established by using three PDAC cell lines which could only represent a part of metabonome landscape of pancreatic cancer, which inevitably lower the level of evidence provided from our study. The heterogeneity of pancreatic cancer in patients may compromise the directly clinical transformation application of our results. Thus, the further validation based on a large patient cohort will be performed in the future.
Conclusions
In this study, we compared the serum metabonomic profiling between PDAC with different differentiations and successfully established pattern recognition models to distinguish with each other. The lower concentration of amino acids, glycolytic and glutaminolytic participators may serve as the predictors for poor differentiation of tumor. Thus, NMR-based metabonomic strategy can be a promising non-invasive approach to predict tumor differentiation preoperatively.
Acknowledgements
No applicable
Funding
This work is sponsored by a. the National Natural Science Foundation of China (No. 81272581, 31671920); b. the United Fujian Provincial Health and Education Project for Tackling the Key Research (No.WKJ-FJ-10); c. National Key Clinical Specialty Discipline Construction Program of China and Key Clinical Specialty Discipline Construction Program of Fujian.
Availability of data and materials
The datasets generated and analyzed during the current study are not publicly available due to no available and appropriate public database can be achieved, but are available from the corresponding author on reasonable request.
Abbreviations
- 1-MH
1-Methylhistidine
- 3-HB
3-Hydroxybutyrate
- AA
Acetoacetate
- Ace
Acetate
- Act
Acetone
- Ala
Alanine
- All
Allantoin
- Bet
Betaine
- Cho
Choline
- Cit
Citrate
- CPMG
Carr-Purcell-Meiboom-Gill
- Cr
Creatine
- EAAs
Essential amino acids
- Eth
Ethanol
- For
Formate
- Fum
Fumarate
- G
Glycerol
- Gln
Glutamine
- Glu
Glutamate
- Gly
Glycine
- GPC
Glycerolphosphocholine
- His
Histidine
- HOD
Residual water signal
- IB
Isobutyrate
- Ile
Isoleucine
- L1
LDL
- L2
VLDL
- Lac
Lactate
- LAT-1
L-type amino-acid transporter 1
- Leu
Leucine
- Lys
Lysine
- Mal
Malonate
- Met
Methionine
- MG
Methylguanidine
- m-I
myo-Inositol
- Mol
Methanol
- mTOR
Mammalian target-of-rapamycin
- NAG
N-acetyl glycoprotein
- NEAAs
Non-essential amino acids
- NMR
Nuclear magnetic resonance
- OPLS-DA
Orthogonal partial least squares discriminant analysis
- PC
Phosphocholine
- PCA
Principal component analysis
- PDAC
Pancreatic ductal adenocarcinoma
- Phe
Phenylalanine
- PLS-DA
Partial least squares discriminant analysis
- Py
Pyruvate
- SGOC pathway
Serine-glycine-one-carbon pathway
- Suc
Succinate
- TCA
Tricarboxylic acid
- Thr
Threonine
- TMA
Trimethylamine
- Trp
Tryptophan
- Tyr
Tyrosine
- Val
Valine
- α-Glc
α-Glucose
- β-Glc
β-Glucose
Authors’ contributions
SW established the animal models, interpreted the metabonomic data regarding PDAC and was the main writer for this manuscript. BZ was in charge of NMR detection of samples and the disposition of spectrum. JF designed the protocol of NMR detection, check the whole experimental data and was a major contributor in writing the manuscript. WH, XL and JB were major contributors to animal models and the preparation of samples. HH was the initiator of this study, design the whole experimental protocol and as the ultimate checker for this manuscript. All authors have read and approved the final version of this manuscript.
Ethics approval
This experimental protocol was in accordance with the principles of National Institutes of Health guide for the care and use of laboratory animals and approved by Ethical Committee of Fujian Medical University.
Consent for publication
No applicable
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Shi Wen, Email: oopsreach@126.com.
Bohan Zhan, Email: zbh119110@qq.com.
Jianghua Feng, Phone: +86-592-2183301, Email: jianghua.feng@xmu.edu.cn.
Weize Hu, Email: sirius0808@126.com.
Xianchao Lin, Email: linxc07@163.com.
Jianxi Bai, Email: 810644415@qq.com.
Heguang Huang, Phone: +86-13705947538, Email: heguanghuang2@163.com, Email: hhuang2@aliyun.com.
References
- 1.Siegel RL, Miller KD, Jemal A. Cancer statistics, 2015. CA Cancer J Clin. 2015;65(1):5–29. doi: 10.3322/caac.21254. [DOI] [PubMed] [Google Scholar]
- 2.Sohn T, Yeo C, Lillemoe K, Koniaris L, Kaushal S, Sauter P, Coleman J, Hruban R, Cameron J. Resected adenocarcinoma of the pancreas - 616 patients: results, outcome and prognostic indicators. Gastroenterology. 2000;118(4):A1059. doi: 10.1016/S0016-5085(00)86385-4. [DOI] [PubMed] [Google Scholar]
- 3.Winter J, Cameron J, Campbell K, Chang D, Coleman J, Hodgin M, Sauter P, Hruban R, Riall T, Schulick R. 1423 Pancreaticoduodenectomies for pancreatic cancer: a single-institution experience. J Gastrointest Surg. 2006;10(9):1199–1211. doi: 10.1016/j.gassur.2006.08.018. [DOI] [PubMed] [Google Scholar]
- 4.Makowiec F, Riediger H, Fischer E, Keck T, Opitz OG, Adam U, Hopt UT. M1547 the lymph node-ratio is the strongest factor predicting survival after resection of pancreatic cancer. Gastroenterology. 2008;134(4):A-870. [Google Scholar]
- 5.Nagai S, Fujii T, Kodera Y, Kanda M, Sahin TT, Kanzaki A, Yamada S, Sugimoto H, Nomoto S, Takeda S. Impact of operative blood loss on survival in invasive Ductal Adenocarcinoma of the pancreas. Pancreas. 2010;40(1):3–9. doi: 10.1097/MPA.0b013e3181f7147a. [DOI] [PubMed] [Google Scholar]
- 6.Shimada K, Sano T, Sakamoto Y, Kosuge T. Clinical implications of combined portal vein resection as a palliative procedure in patients undergoing pancreaticoduodenectomy for pancreatic head carcinoma. Ann Surg Oncol. 2006;13(12):1569–1578. doi: 10.1245/s10434-006-9143-4. [DOI] [PubMed] [Google Scholar]
- 7.Mitsuro K, Tsutomu F, Sahin TT, Akiyuki K, Shunji N, Suguru Y, Hiroyuki S, Shuji N, Shin T, Yasuhiro K. Invasion of the splenic artery is a crucial prognostic factor in carcinoma of the body and tail of the pancreas. Ann Surg. 2010;251(3):483–487. doi: 10.1097/SLA.0b013e3181cf9171. [DOI] [PubMed] [Google Scholar]
- 8.Sugiura T, Uesaka K, Mihara K, Sasaki K, Kanemoto H, Mizuno T, Okamura Y. Margin status, recurrence pattern, and prognosis after resection of pancreatic cancer. Surgery. 2013;154(5):1078–1086. doi: 10.1016/j.surg.2013.04.015. [DOI] [PubMed] [Google Scholar]
- 9.Neoptolemos JP, Stocken DD, Dunn JA, Almond J, Beger HG, Pederzoli P, Bassi C, Dervenis C, Fernandez-Cruz L, Lacaine F, et al. Influence of resection margins on survival for patients with pancreatic cancer treated by adjuvant chemoradiation and/or chemotherapy in the ESPAC-1 randomized controlled trial. Ann Surg. 2001;234(6):758–768. doi: 10.1097/00000658-200112000-00007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Saukkonen K, Hagstrom J, Mustonen H, Juuti A, Nordling S, Kallio P, Alitalo K, Seppanen H, Haglund C. PROX1 and beta-catenin are prognostic markers in pancreatic ductal adenocarcinoma. BMC Cancer. 2016;16:472. doi: 10.1186/s12885-016-2497-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Reiser-Erkan C, Erkan M, Pan Z, Bekasi S, Giese NA, Streit S, Michalski CW, Friess H, Kleeff J. Hypoxia-inducible proto-oncogene Pim-1 is a prognostic marker in pancreatic ductal adenocarcinoma. Cancer Biol Ther. 2008;7(9):1352–1359. doi: 10.4161/cbt.7.9.6418. [DOI] [PubMed] [Google Scholar]
- 12.Neoptolemos JP, Stocken DD, Friess H, Bassi C, Dunn JA, Hickey H, Beger H, Fernandez-Cruz L, Dervenis C, Lacaine F, et al. A randomized trial of chemoradiotherapy and chemotherapy after resection of pancreatic cancer. N Engl J Med. 2004;350(12):1200–1210. doi: 10.1056/NEJMoa032295. [DOI] [PubMed] [Google Scholar]
- 13.Barugola G, Partelli S, Marcucci S, Sartori N, Capelli P, Bassi C, Pederzoli P, Falconi M. Resectable pancreatic cancer: who really benefits from resection? Ann Surg Oncol. 2009;16(12):3316–3322. doi: 10.1245/s10434-009-0670-7. [DOI] [PubMed] [Google Scholar]
- 14.Hsu CC, Wolfgang CL, Laheru DA, Pawlik TM, Swartz MJ, Winter JM, Robinson R, Edil BH, Narang AK, Choti MA, et al. Early mortality risk score: identification of poor outcomes following upfront surgery for resectable pancreatic cancer. J Gastrointest Surg. 2012;16(4):753–761. doi: 10.1007/s11605-011-1811-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Joliat GR, Petermann D, Demartines N, Schafer M. External assessment of the early mortality risk score in patients with adenocarcinoma undergoing pancreaticoduodenectomy. HPB (Oxford) 2015;17(7):605–610. doi: 10.1111/hpb.12422. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Mornex F, Girard N, Delpero JR, Partensky C. Radiochemotherapy in the management of pancreatic cancer--part I: neoadjuvant treatment. Semin Radiat Oncol. 2005;15(4):226–234. doi: 10.1016/j.semradonc.2005.04.008. [DOI] [PubMed] [Google Scholar]
- 17.Quiros RM, Brown KM, Hoffman JP. Neoadjuvant therapy in pancreatic cancer. Cancer Investig. 2007;25(4):267–273. doi: 10.1080/07357900701206356. [DOI] [PubMed] [Google Scholar]
- 18.Varadhachary GR, Tamm EP, Abbruzzese JL, Xiong HQ, Crane CH, Wang H, Lee JE, Pisters PW, Evans DB, Wolff RA. Borderline resectable pancreatic cancer: definitions, management, and role of preoperative therapy. Ann Surg Oncol. 2006;13(8):1035–1046. doi: 10.1245/ASO.2006.08.011. [DOI] [PubMed] [Google Scholar]
- 19.Larghi A, Correale L, Ricci R, Abdulkader I, Monges G, Iglesias-Garcia J, Giovannini M, Attili F, Vitale G, Hassan C, et al. Interobserver agreement and accuracy of preoperative endoscopic ultrasound-guided biopsy for histological grading of pancreatic cancer. Endoscopy. 2015;47(4):308–314. doi: 10.1055/s-0034-1390912. [DOI] [PubMed] [Google Scholar]
- 20.Nurmi A, Haglund C, Mustonen H, Seppänen H. Neoadjuvant therapy offers longer survival for pancreatic cancer patients over upfront surgery. Pancreatology. 2017;17(3):S73. doi: 10.1016/j.pan.2017.05.230. [DOI] [Google Scholar]
- 21.Mokdad AA, Minter RM, Zhu H, Augustine MM, Porembka MR, Wang SC, Yopp AC, Mansour JC, Choti MA, Polanco PM. Neoadjuvant therapy followed by resection versus upfront resection for Resectable pancreatic cancer: a propensity score matched analysis. J Clin Oncol. 2016;35(4):515-22. [DOI] [PubMed]
- 22.Network NCC . NCCN clinical practice guidelines in oncology: pancreatic Adenocarcinoma(version 2.2014) Fort Washington: National Comprehensive Cancer Network; 2014. [Google Scholar]
- 23.Farrell JM, Pang JC, Kim GE, Tabatabai ZL. Pancreatic neuroendocrine tumors: accurate grading with Ki-67 index on fine-needle aspiration specimens using the WHO 2010/ENETS criteria. Cancer Cytopathol. 2014;122(10):770–778. doi: 10.1002/cncy.21457. [DOI] [PubMed] [Google Scholar]
- 24.Pitman MB, Centeno BA, Genevay M, Fonseca R, Mino-Kenudson M. Grading epithelial atypia in endoscopic ultrasound-guided fine-needle aspiration of intraductal papillary mucinous neoplasms: an international interobserver concordance study. Cancer Cytopathol. 2013;121(12):729–736. doi: 10.1002/cncy.21334. [DOI] [PubMed] [Google Scholar]
- 25.Brindle JT, Antti H, Holmes E, Tranter G, Nicholson JK, Bethell HWL, Clarke S, Schofield PM, Mckilligin E, Mosedale DE. Rapid and noninvasive diagnosis of the presence and severity of coronary heart disease using 1H-NMR-based metabonomics. Nat Med. 2002;8(12):1439–1444. doi: 10.1038/nm1202-802. [DOI] [PubMed] [Google Scholar]
- 26.Bartella L, Morris EA, Dershaw DD, Liberman L, Thakur SB, Moskowitz C, Guido J, Huang W. Proton MR spectroscopy with choline peak as malignancy marker improves positive predictive value for breast cancer diagnosis: preliminary study. Radiology. 2006;239(3):686–692. doi: 10.1148/radiol.2393051046. [DOI] [PubMed] [Google Scholar]
- 27.Huzjan R, Sala E, Hricak H. Magnetic resonance imaging and magnetic resonance spectroscopic imaging of prostate cancer. Nat Clin Pract Urol. 2005;2(9):434–442. doi: 10.1038/ncpuro0296. [DOI] [PubMed] [Google Scholar]
- 28.Khan SA, Cox IJ, Thillainayagam AV, Bansi DS, Thomas HC, Taylor-Robinson SD. Proton and phosphorus-31 nuclear magnetic resonance spectroscopy of human bile in hepatopancreaticobiliary cancer. Eur J Gastroenterol Hepatol. 2005;17(7):733–738. doi: 10.1097/00042737-200507000-00007. [DOI] [PubMed] [Google Scholar]
- 29.Kwock L, Smith JK, Castillo M, Ewend MG, Collichio F, Morris DE, Bouldin TW, Cush S. Clinical role of proton magnetic resonance spectroscopy in oncology: brain, breast, and prostate cancer. Lancet Oncol. 2006;7(10):859–868. doi: 10.1016/S1470-2045(06)70905-6. [DOI] [PubMed] [Google Scholar]
- 30.Rehman L, Rehman UL, Azmat SK, Mohammad Hashim AS. Magnetic resonance spectroscopy: novel non-invasive technique for diagnosing brain tumors. J Coll Physicians Surg Pak. 2015;25(12):863–866. [PubMed] [Google Scholar]
- 31.Thomas MA, Wyckoff N, Yue K, Binesh N, Banakar S, Chung HK, Sayre J, DeBruhl N. Two-dimensional MR spectroscopic characterization of breast cancer in vivo. Technol Cancer Res Treat. 2005;4(1):99–106. doi: 10.1177/153303460500400113. [DOI] [PubMed] [Google Scholar]
- 32.Kaplan O, Kushnir T, Askenazy N, Knubovets T, Navon G. Role of nuclear magnetic resonance spectroscopy (MRS) in cancer diagnosis and treatment: 31P, 23Na, and 1H MRS studies of three models of pancreatic cancer. Cancer Res. 1997;57(8):1452–1459. [PubMed] [Google Scholar]
- 33.Bathe OF, Shaykhutdinov R, Kopciuk K, Weljie AM, McKay A, Sutherland FR, Dixon E, Dunse N, Sotiropoulos D, Vogel HJ. Feasibility of identifying pancreatic cancer based on serum metabolomics. Cancer Epidemiol Biomarkers Prev. 2011;20(1):140–147. doi: 10.1158/1055-9965.EPI-10-0712. [DOI] [PubMed] [Google Scholar]
- 34.Zhang L, Jin H, Guo X, Yang Z, Zhao L, Tang S, Mo P, Wu K, Nie Y, Pan Y, et al. Distinguishing pancreatic cancer from chronic pancreatitis and healthy individuals by (1)H nuclear magnetic resonance-based metabonomic profiles. Clin Biochem. 2012;45(13-14):1064–1069. doi: 10.1016/j.clinbiochem.2012.05.012. [DOI] [PubMed] [Google Scholar]
- 35.Fang F, He X, Deng H, Chen Q, Lu J, Spraul M, Yu Y. Discrimination of metabolic profiles of pancreatic cancer from chronic pancreatitis by high-resolution magic angle spinning 1H nuclear magnetic resonance and principal components analysis. Cancer Sci. 2007;98(11):1678–1682. doi: 10.1111/j.1349-7006.2007.00589.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Tan MH, Nowak NJ, Loor R, Ochi H, Sandberg AA, Lopez C, Pickren JW, Berjian R, Douglass HO, Chu TM. Characterization of a new primary human pancreatic tumor line. Cancer Investig. 1986;4(1):15–23. doi: 10.3109/07357908609039823. [DOI] [PubMed] [Google Scholar]
- 37.Kyriazis AP, Sandberg AA, Kyriazis AA, Sloane NH, Lepera R. Establishment and characterization of human pancreatic adenocarcinoma cell line SW-1990 in tissue culture and the nude mouse. Cancer Res. 1983;43(9):4393–4401. [PubMed] [Google Scholar]
- 38.Lieber M, Mazzetta J, Nelson-Rees W, Kaplan M, Todaro G. Establishment of a continuous tumor-cell line (panc-1) from a human carcinoma of the exocrine pancreas. Int J Cancer. 1975;15(5):741–747. doi: 10.1002/ijc.2910150505. [DOI] [PubMed] [Google Scholar]
- 39.Deer EL, Jessica GH, Coursen JD, Shea JE, Josephat N, Scaife CL, Firpo MA, Mulvihill SJ. Phenotype and genotype of pancreatic cancer cell lines. Pancreas. 2010;39(4):425–435. doi: 10.1097/MPA.0b013e3181c15963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Chagoyen M, Pazos F. MBRole: enrichment analysis of metabolomic data. Bioinformatics. 2011;27(5):730–731. doi: 10.1093/bioinformatics/btr001. [DOI] [PubMed] [Google Scholar]
- 41.Kanehisa M, Goto S, Sato Y, Kawashima M, Furumichi M, Tanabe M. Data, information, knowledge and principle: back to metabolism in KEGG. Nucleic Acids Res. 2014;42(Database issue):D199–D205. doi: 10.1093/nar/gkt1076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Wishart DS, Knox C, Guo AC, Eisner R, Young N, Gautam B, Hau DD, Psychogios N, Dong E, Bouatra S, et al. HMDB: a knowledgebase for the human metabolome. Nucleic Acids Res. 2009;37(Database issue):D603–D610. doi: 10.1093/nar/gkn810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Cui Q, Lewis IA, Hegeman AD, Anderson ME, Li J, Schulte CF, Westler WM, Eghbalnia HR, Sussman MR, Markley JL. Metabolite identification via the Madison Metabolomics consortium database. Nat Biotechnol. 2008;26(2):162–164. doi: 10.1038/nbt0208-162. [DOI] [PubMed] [Google Scholar]
- 44.Hirschhaeuser F, Sattler UG, Mueller-Klieser W. Lactate: a metabolic key player in cancer. Cancer Res. 2011;71(22):6921–6925. doi: 10.1158/0008-5472.CAN-11-1457. [DOI] [PubMed] [Google Scholar]
- 45.Guillaumond F, Leca J, Olivares O, Lavaut MN, Vidal N, Berthezene P, Dusetti NJ, Loncle C, Calvo E, Turrini O, et al. Strengthened glycolysis under hypoxia supports tumor symbiosis and hexosamine biosynthesis in pancreatic adenocarcinoma. Proc Natl Acad Sci U S A. 2013;110(10):3919–3924. doi: 10.1073/pnas.1219555110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.OuYang D, Xu J, Huang H, Chen Z. Metabolomic profiling of serum from human pancreatic cancer patients using 1H NMR spectroscopy and principal component analysis. Appl Biochem Biotechnol. 2011;165(1):148–154. doi: 10.1007/s12010-011-9240-0. [DOI] [PubMed] [Google Scholar]
- 47.Kobayashi T, Nishiumi S, Ikeda A, Yoshie T, Sakai A, Matsubara A, Izumi Y, Tsumura H, Tsuda M, Nishisaki H, et al. A novel serum metabolomics-based diagnostic approach to pancreatic cancer. Cancer Epidemiol Biomarkers Prev. 2013;22(4):571–579. doi: 10.1158/1055-9965.EPI-12-1033. [DOI] [PubMed] [Google Scholar]
- 48.Soga T. Cancer metabolism: key players in metabolic reprogramming. Cancer Sci. 2013;104(3):275–281. doi: 10.1111/cas.12085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Son J, Lyssiotis CA, Ying H, Wang X, Hua S, Ligorio M, Perera RM, Ferrone CR, Mullarky E, Shyh-Chang N, et al. Glutamine supports pancreatic cancer growth through a KRAS-regulated metabolic pathway. Nature. 2013;496(7443):101–105. doi: 10.1038/nature12040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Mayers JR, Wu C, Clish CB, Kraft P, Torrence ME, Fiske BP, Yuan C, Bao Y, Townsend MK, Tworoger SS, et al. Elevation of circulating branched-chain amino acids is an early event in human pancreatic adenocarcinoma development. Nat Med. 2014;20(10):1193–1198. doi: 10.1038/nm.3686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Fuchs BC, Bode BP. Amino acid transporters ASCT2 and LAT1 in cancer: partners in crime? Semin Cancer Biol. 2005;15(4):254–266. doi: 10.1016/j.semcancer.2005.04.005. [DOI] [PubMed] [Google Scholar]
- 52.Kaira K, Sunose Y, Arakawa K, Ogawa T, Sunaga N, Shimizu K, Tominaga H, Oriuchi N, Itoh H, Nagamori S, et al. Prognostic significance of L-type amino-acid transporter 1 expression in surgically resected pancreatic cancer. Br J Cancer. 2012;107(4):632–638. doi: 10.1038/bjc.2012.310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Phang JM, Liu W, Hancock C. Bridging epigenetics and metabolism: role of non-essential amino acids. Epigenetics. 2013;8(3):231–236. doi: 10.4161/epi.24042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Tedeschi PM, Markert EK, Gounder M, Lin H, Dvorzhinski D, Dolfi SC, Chan LL, Qiu J, DiPaola RS, Hirshfield KM, et al. Contribution of serine, folate and glycine metabolism to the ATP, NADPH and purine requirements of cancer cells. Cell Death Dis. 2013;4:e877. doi: 10.1038/cddis.2013.393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Kottakis F, Nicolay BN, Roumane A, Karnik R, Gu H, Nagle JM, Boukhali M, Hayward MC, Li YY, Chen T, et al. LKB1 loss links serine metabolism to DNA methylation and tumorigenesis. Nature. 2016;539(7629):390-95. [DOI] [PMC free article] [PubMed]
- 56.Glunde K, Bhujwalla ZM, Ronen SM. Choline metabolism in malignant transformation. Nat Rev Cancer. 2011;11(12):835–848. doi: 10.1038/nrc3162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Stewart JD, Marchan R, Lesjak MS, Lambert J, Hergenroeder R, Ellis JK, Lau CH, Keun HC, Schmitz G, Schiller J, et al. Choline-releasing glycerophosphodiesterase EDI3 drives tumor cell migration and metastasis. Proc Natl Acad Sci U S A. 2012;109(21):8155–8160. doi: 10.1073/pnas.1117654109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Penet MF, Shah T, Bharti S, Krishnamachary B, Artemov D, Mironchik Y, Wildes F, Maitra A, Bhujwalla ZM. Metabolic imaging of pancreatic ductal adenocarcinoma detects altered choline metabolism. Clin Cancer Res. 2015;21(2):386–395. doi: 10.1158/1078-0432.CCR-14-0964. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets generated and analyzed during the current study are not publicly available due to no available and appropriate public database can be achieved, but are available from the corresponding author on reasonable request.